
Abstract
Stable isotopic compositions of carbon (δ13C) and oxygen (δ18O) in plants reflect growth conditions. Therefore, these isotopes might be good indicators of changes in environmental factors, such as variations in air temperature caused by climate change. It is predicted that climate change will lead to a greater increase in minimum air temperatures (primarily during the night) than in maximum air temperatures (primarily during the day) in many parts of Japan. In the present study, we investigated whether the δ13C and δ18O of the rice grain Koshihikari (Oryza sativa L.) from the northern latitudes (30.49°–37.14°) of Japan reflect variations in air temperature during grain filling and are related to the yield and proportion of first-grade rice (<15 % transparency, roundness, and cracking) as an indicator of quality. We revealed that rice δ13C was not correlated with mean maximum or minimum air temperatures for each prefecture. By contrast, rice δ18O was positively correlated with mean minimum air temperature, suggesting that rice δ18O reflects changes in night air temperature. We further showed that an increase in the mean minimum air temperature during grain filling had a negative effect on rice yield and quality. Our findings indicate that the δ18O of rice grain may be a good indicator of physiological changes in response to minimum air temperatures during grain filling.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Intergovernmental Panel on Climate Change (IPCC 2007) has developed future climate scenarios based on expected increases in temperature and atmospheric concentrations of greenhouse gases such as carbon dioxide (CO2), methane, and nitrous oxide. Increased concentrations of CO2, coupled with higher temperatures, are likely to have a considerable impact on the future dynamics of water and nutrients, and also on the structure and function of ecosystems (Cramer et al. 2001; Henry et al. 2005). This, in turn, will lead to substantial changes in agricultural crops (Peng et al. 2004; Yoshimoto et al. 2005; Masutomi et al. 2010), which are widely distributed and grown using the same agricultural management practices.
The Japanese archipelago extends over a distance of approximately 2,000 km, and its climate ranges from subtropical to subarctic (Japan Meteorological Agency 2005). There are concerns that climate change in the region will affect not only the production but also the quality of rice (Oryza sativa L.)—one of the most important basic food resources in Japan. The impact of climate change on rice production has been investigated extensively using crop models and climate change scenarios (Matthews et al. 1997; Aggarwal and Mall 2002; Tao et al. 2008). The results suggest that global warming will enhance rice production in northern Japan, but reduce production by at least 15–30 % in the south-central and southwestern parts of the country (Horie et al. 1995). In addition, high temperatures during the grain-filling period may reduce rice quality, as measured by grain size, transparency, roundness, and cracking (Yamakawa et al. 2007; Morita 2008; Hasegawa et al. 2011). However, the way in which high air temperature, especially at night, decreases the fertility of rice grains remains unclear (Cheng et al. 2009).
Carbon isotope discrimination (Δ13C) occurs in C3 plants, owing to series of isotope fractionations along the photosynthesis pathway. As a consequence, plant carbon is invariably depleted in 13C in respect to the isotopic composition of atmospheric CO2. The parameter Δ13C is an important indicator of all those conditions affecting the photosynthetic set point, i.e., the internal to atmospheric CO2 concentration ratio (Ci/Ca) (Farquhar et al. 1989). The δ13C of plants depends on fractionation during diffusion of CO2 into the leaf, and also on subsequent photosynthetic metabolism and water-use efficiency (WUE) independently (Farquhar et al. 1982; Kume et al. 2003; Hanba et al. 2010; Ma et al. 2012). Most carbon stored in mature rice grains originates from CO2 assimilation during the grain filling period, with the flag leaf as the most photosynthetically active (Murchie et al. 1999). Factors such as the photosynthesis rate of the flag leaf during this period could potentially determine grain yield (Dingkuhn et al. 1989). Furthermore, the plastic response of WUE to water availability has been demonstrated using instantaneous gas-exchange measurements, by analyses of plant δ13C (Ehleringer and Cooper 1988; Toft et al. 1989; Centritto et al. 2009). Therefore, rice δ13C may reflect an integration over time of photosynthesis and plant water status (with related factors such as sunshine hours, precipitation, humidity, and air temperature), as this is affected by water availability and evapotranspiration demand by the atmosphere.
Oxygen isotope discrimination (Δ18O) in C3 plants occurs mainly during transpiration at the leaf relating to the atmospheric to internal vapor pressure ratio (Barbour 2007). Although the oxygen stable isotopic composition (δ18O) of xylem water reflects purely soil water taken up by roots (Dawson and Ehleringer 1991), there are complex phenomena of remixing and back diffusion of the enriched water with xylem water (i.e., metabolic water) arriving to the leaves and permeating the chloroplasts, consequently determining the δ18O of metabolic water (Farquhar and Lloyd 1993). Additionally, the δ18O of soil water is normally closely linked to precipitation, which becomes enhanced with δ18O as the temperature increases at lower latitudes and altitudes (Dansgaard 1964; Mizota and Kusakabe 1994). The metabolic water retains the environmental signal (both soil water and transpiration effects) and is detectably affected by air temperature, which imprints the isotopic signature of the organic matter by means of isotopic exchange on the gem-diole groups of triose-phosphates (Samuel and Silver 1965; Sternberg et al. 1986; Roden and Farquhar 2012). Maximum air temperature (primarily during the day) has the potential to affect both photosynthesis and transpiration, whereas minimum air temperature (primarily at night) may affect transpiration, which promotes translocation in mature rice grains. Therefore, it is expected that the δ13C and δ18O of organic matter in rice grains will reflect physiological changes in response to warming impacts. If the δ13C and δ18O of rice grains reflect an increase in maximum and/or minimum air temperatures, they may provide direct indicators for changes in rice yield and quality under conditions of climate change.
The aim of the present study was to investigate whether stable carbon and oxygen isotopes of rice organic matter reflect variations in air temperature during grain filling, and are related to the yield and quality of rice. We analyzed the δ13C and δ18O of rice grains from the northern latitudes (30.49°–37.14°) of Japan in relation to mean maximum and minimum air temperatures, yield, and quality—as defined by the content of first-grade rice.
Materials and methods
Rice and rice-field water samples
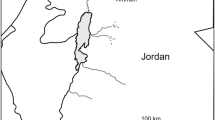
A total of 205 samples of the japonica rice cultivar Koshihikari (Oryza sativa L.) were obtained from 114 paddies within 21 prefectures in September 2007, and 91 paddies within 13 prefectures in September 2008, on the Japanese islands of Honshu, Shikoku, and Kyusyu (detailed in Fig. 1; Table 1). We selected and polished rice crops for which heading and flowering occurred between July and August. Each polished rice grain sample (50 g) was freeze-dried and ground into a powder for stable isotope analysis. In May 2007, water samples were filtered by a membrane filter (nominal pore size: 0.45 μm, ADVANTEC, Tokyo, Japan) and collected in 100 ml polyethylene bottles from the irrigation ditch of one rice field in each of the following locations (within the region 24.35–38.17°N): Sado (Niigata), Uonuma (Niigata), Murakami (Niigata), Matsusaka (Mie), Matsukawa (Nagano), Matsumoto (Nagano), Matsue (Shimane), Unnan (Shimane), and Ishigaki (Okinawa) in order to consider geographical changes in isotopic composition of source water (Fig. 1). The rice crops were irrigated and fertilized according to conventional management practices for rice farming in Japan. Intermittent irrigation was applied from approximately 1 month after transplanting to maintain wet soil conditions. Irrigation was stopped from approximately 1 month after ear emergence until harvest, because precipitation water provided sufficient water levels for crop production (e.g., Chapagain and Yamaji 2010).

Sampling locations of Koshihikari rice (Oryza sativa L.) and rice-field water in Japan. Closed and open circles represent locations of rice samples in 2007 and 2008, respectively. Open triangles represent locations of rice-field water samples in 2007
Meteorological and rice statistical data collection
Meteorological data (mean maximum and minimum air temperatures, precipitation, and sunshine hours during the grain-filling period) were obtained from local meteorological stations within a 15-km radius of each sampling point (Japan Meteorological Agency 2012). Where possible, these data were collected for the grain-filling period of Koshihikari rice, determined according to statistical reports (Ministry of Agriculture, Forestry and Fisheries 2012). Rice yield and grade data for 2007 and 2008 were collected from statistical reports (Ministry of Agriculture, Forestry and Fisheries 2012). We used the characteristics of first-grade rice (<15 % transparency, roundness, and cracking) as indicators of quality.
Stable isotope analysis
The δ13C of rice grain (δ13Crice) was determined using an elemental analyzer/isotope ratio mass spectrometer (EA/IRMS, Finnigan Delta V Advantage, interfaced with FlashEA 1112 HT; Thermo Fisher Scientific, Bremen, Germany) (Suzuki et al. 2008). Subsamples of rice (1.0 mg) were placed into tin capsules (5 × 9 mm) and loaded into the auto-sampler of the EA/IRMS. The obtained CO2 was separated on a PorapakQS column (length, 3 m) at 40 °C. The δ notation defined in Eq. (1) was used to describe the isotopic composition.
where R sample is the isotope ratio of the sample, and R standard is the isotope ratio of the international standard, Vienna Pee Dee Belemnite. Each sample was determined in duplicate, and analytical errors in δ13C had a standard deviation (SD) <0.1 ‰.
The δ18O of rice grain (δ18Orice) and rice-field water (δ18Owater) were determined by EA/IRMS in the pyrolysis mode. Subsamples of rice and rice-field water (0.5 mg and 1.0 μL, respectively) were placed into silver capsules (5 × 3 mm). Care was taken to avoid atmospheric contamination of the samples. All rice samples were dried overnight by lyophilization before isotope measurement. Pyrolysis was performed in a ceramic tube of glassy carbon at 1,450 °C under a continuous flow of He at 100 ml min−1. The gas obtained was separated from residual gases by molecular sieving through a 5-A˚ column (length, 3 m) at 50 °C (Werner et al. 1996; Böhlke et al. 2003; Leuenberger and Filot 2007). δ18O was calculated according to Eq. (1) and expressed relative to that of Vienna Standard Mean Ocean Water (V-SMOW). The measured δ18O values were obtained using known isotope benzoic acid standards (+71.4, +23.2 ‰) purchased from Indiana University (Indianapolis, IN). The accuracy of the obtained δ18O values was checked independently against V-SMOW and Vienna Standard Light Antarctic Precipitation standards distributed by the International Atomic Energy Agency. Four working standards—dibenzo-24-crown-8 (−15.7 ‰), dibenzo-18-crown-6 (+1.7 ‰), β-d-galactose pentaacetate (+12.7 ‰), and D-(+)-sucrose octaacetate (+26.8 ‰)—were determined every six samples to confirm the reproducibility of the measurements (Nakashita et al. 2008). The analytical error of the isotope measurements had a SD <0.3 ‰, with a minimum sample amount of 10 μmol oxygen.
Statistical analysis
Correlations between rice (δ13C, δ18O, yield, and quality) and meteorological data (mean maximum and minimum air temperatures, precipitation, and sunshine hours during the grain-filling period) were tested independently using Pearson’s product-moment coefficient with the average values for each prefecture. We excluded from our statistical analyses the rice yield and quality of 2007 data from Kagoshima and Miyazaki prefectures, because of typhoon damage. Correlations between the δ18Owater and the δ18Orice, and also between the mean minimum air temperature during grain filling and the δ18Orice and the δ18Owater with global mean 18O enrichment of carbohydrate relative to the source water (+27 ‰, Sternberg 1989), were tested using Pearson’s product-moment coefficient. In this study, the 18O enrichment of rice grain to source water was defined as δ18Orice–δ18Owater (e.g. Matsuo et al. 2013). We compared the theoretical values (δ18Owater + global mean 18O enrichment) with measured δ18Orice, because the difference between the theoretical and measured 18O enrichments may reveal physiological changes such as rice translocation and evaporative demands. For all tests, an α value of 0.05 was used to indicate statistical significance. All analyses were conducted using R ver. 2.15.0 (R Development Core Team 2012).
Results
Rice yields ranged from 2.05 to 6.27 t ha−1 in 2007, and from 4.67 to 6.35 t ha−1 in 2008 (Table 1). The proportion of first-grade rice in the rice yield (rice quality) ranged from 0 to 97 % in 2007, and from 30 % to 98 % in 2008. There was a positive correlation between yield and quality (Table 2). Sunshine hours during grain filling ranged from 144 to 276 h in 2007, and from 118 to 268 h in 2008. Sunshine hours were not correlated with yield or quality of rice (Table 2). Precipitation during grain filling ranged from 10 to 701 mm in 2007, and from 37 to 389 mm in 2008. Precipitation was also not correlated with yield or quality of rice (Table 2).
Rice δ13C ranged from −28.4 to −26.8 ‰ in 2007, and from −28.1 to −27.1 ‰ in 2008 (Table 1). Rice δ13C was not correlated with yield, quality, sunshine hours, or precipitation (Table 2). δ18Orice ranged from 20.8 to 24.1 ‰ in 2007, and from 20.4 to 23.9 ‰ in 2008 (Table 1). δ18Orice was not correlated with sunshine hours, yield, quality or precipitation (Table 2). The δ18Orice was not correlated with the δ13C (Table 2).
The mean maximum air temperatures ranged from 30.6 to 34.5 °C in 2007, and from 28.3 to 34.5 °C in 2008 (Table 1). The maximum air temperature was negatively correlated with rice yield (Fig. 2a), quality (Fig. 2b), but not with δ13C (Fig. 2c), or δ18Orice (Fig. 2d).

Relationships among mean maximum and minimum air temperatures during grain filling, and yield, quality, δ13C, and δ18O of Koshihikari rice (Oryza sativa L.). a Yield versus mean maximum air temperature, b quality versus mean maximum air temperature, c δ13C versus mean maximum air temperature, d δ18O versus mean maximum air temperature, e yield versus mean minimum air temperature, f quality versus mean minimum air temperature, g δ13C versus mean minimum air temperature, h δ18O versus mean minimum air temperature. Data are represented as mean ± SD
The mean minimum air temperatures ranged from 19.4 to 24.9 °C in 2007, and from 19.3 to 24.6 °C in 2008 (Table 1). The minimum air temperature was negatively correlated with rice yield (Fig. 2e) and quality (Fig. 2f), positively correlated with δ18Orice (Fig. 2h), but not correlated with rice δ13C (Fig. 2g).
There was a positive correlation between the δ18Owater and δ18Orice (Fig. 3). The 18O enrichment of rice relative to rice-field water decreased significantly with an increase in the mean minimum air temperature during grain filling (Fig. 4).

Relationship between the δ18O of rice-field water and Koshihikari rice (Oryza sativa L.) in different regions of Japan

Relationship between mean minimum air temperature during grain filling and the δ18O of Koshihikari rice (Oryza sativa L.) and rice-field water with global mean 18O enrichment of carbohydrate relative to the source water (+27 ‰)
Discussion
In the present study, the δ18Orice was correlated positively with the δ18Owater (Fig. 3), confirming that the oxygen atoms in the organic compounds of rice grain were derived mainly from the region-specific ambient water. The δ18O of precipitation and groundwater (i.e., source water for rice plants) varied in accordance with climatic and geographical changes relating to amount and thermal effects (Dansgaard 1964; Rozanski et al. 1992), and becomes enhanced at lower latitudes and altitudes in Japan (e.g., Mizota and Kusakabe 1994). Therefore, similar to variations in the δ18O of organic compounds (e.g., cellulose, sucrose, etc.) in trees to climate gradients in air temperature, rainfall, sunshine hours, and water availability during the growing season (Libby and Pandolfi 1974; Cernusak et al. 2003; Li et al. 2011), the δ18Orice may reflect temperature-induced changes in the δ18Owater. However, the δ18O of irrigation water may not fully reflect precipitation δ18O in some locations, because irrigation water is obtained from various sources—not only rivers, but also ground water and springs far away from the paddy fields. For instance, water used in paddy fields may be derived from precipitation in the mountains either directly or via ground water or springs. As a result, sources of water used in different paddy field regions need to be examined in order to reflect temperature-induced changes.
We found negative effects of mean minimum air temperature during grain filling on the δ18Orice (Fig. 2h), but no significant effects of mean maximum air temperature (Fig. 2d). Throughout many parts of Japan, global warming is predicted to lead to a greater increase in minimum air temperatures than in maximum air temperatures (Japan Meteorological Agency 2005). Therefore, the δ18Orice is potentially a good indicator for climate-change-induced variations in minimum air temperature. The mean enrichment of δ18Orice relative to source water was +31 ± 2 ‰, it is 4 ‰ higher than the global mean 18O enrichment of carbohydrate relative to source water of +27 ± 4 ‰ (Sternberg 1989). The oxygen isotope enrichment of rice grain decreased as the minimum air temperature increased and the difference between measured and global mean 18O enrichments decreased with increase in the minimum temperature (Fig. 4). A rise in minimum air temperature forecast to cause increase in respiration and translocation (Morita 2008), having the potential for increase in δ18O of the rice grain with degradation of starch. Additionally, the increase of minimum air temperature may induce an increase in soil and metabolic water temperatures, and may act to promote of smaller isotope enrichment during transpiration and photosynthesis compared to low water temperatures, because high water temperature increases stomatal conductance and leaf photosynthesis (Shimono et al. 2004). In this study, although we did not accumulate data for other environmental factors such as humidity, altitudes, and nutrients, they may help to demonstrate the effects of minimum temperature on δ18Orice. The δ18O of rice carbohydrate is also dependent on the actual metabolic status of plants as reported by Saurer et al. (1997).
Many physiological studies have indicated that decreases in grain size and quality, such as transparency, roundness, and cracking, are caused by abnormal growth of the endosperm under high temperatures (e.g., Morita 2008; Hasegawa et al. 2011). For example, rice grain size was shown to decrease when night air temperatures exceeded 21 °C during grain filling (Matsushima and Tsunoda 1957; Aimi et al. 1959). In the present study, we detected stronger negative effects from mean minimum air temperature during grain filling on rice yield and quality than from mean maximum air temperature (Fig. 2). Yamakawa et al. (2007) reported that several starch synthesis–related genes, such as granule-bound starch synthase I (GBSSI), branching enzymes (especially BEIIb), and a cytosolic pyruvate orthophosphate dikinase, are downregulated by high temperatures, whereas genes for starch-consuming α-amylases and heat shock proteins are upregulated. Therefore, an increase in minimum air temperature during grain filling can cause changes in the processes involved in amylose composition and translocation, and might lead to changes in the δ18Orice. Minimum air temperature could therefore be useful for predicting the yield and quality of rice, with changes in the δ18Orice reflecting the physiological conditions during grain filling.
In contrast to the δ18Orice, the δ13Crice was not correlated with minimum or maximum air temperatures, suggesting that the δ13Crice might not be a direct indicator of changes in air temperature. The δ13C of rice is known to be a good indicator of WUE (Kondo et al. 2004; Xu et al. 2009). In the present study, we observed a relatively small difference in rice δ13C among the prefectures surveyed (1.6 ‰), suggesting nearly constant water-use efficiency. The δ13Crice might reflect the nearly constant wet soil conditions maintained under the unified irrigation and fertilization practices of conventional rice farming in Japan, irrespective of the differences in plant-experienced atmospheric vapor demand, namely the leaf-to-air vapor pressure difference caused by different atmospheric temperatures, hours of sunshine, and precipitation. We determined no effects of precipitation on the yield and quality of rice; this finding could also be derived from cultivation under constantly wet soil conditions.
References
Aggarwal PK, Mall RK (2002) Climate change and rice yields in diverse agro-environments of India. II. Effect of uncertainties in scenarios and crop models on impact assessment. Clim Change 52:331–343
Aimi R, Sawamura H, Konno S (1959) Physiological studies on the mechanism of crop plants: the effect of the temperature upon the behaviour of carbohydrates and some related enzymes during the ripening of rice plant (in Japanese with English abstract). Jpn J Crop Sci 27: 405–407
Barbour MM (2007) Stable oxygen isotope composition of plant tissue: a review. Funct Plant Biol 34:83–94
Böhlke JK, Mroczkowski SJ, Coplen TB (2003) Oxygen isotopes in nitrate: new reference materials for 18O:17O:16O measurements and observations on nitrate-water equilibration. Rapid Commun Mass Spectrom 17:1835–1846
Centritto M, Lauteri M, Monteverdi MC, Serraj R (2009) Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage. J Exp Botany 60:2325–2339
Cernusak LA, Arthur DJ, Pate JS, Farquhar GD (2003) Water relations link carbon and oxygen isotope discrimination to phloem sap sugar concentration in Eucalyptus globulus. Plant Physiol 131:1544–1554
Chapagain T, Yamaji E (2010) The effects of irrigation method, age of seedling and spacing on crop performance, productivity and water-wise rice production in Japan. Paddy Water Environ 8:81–90
Cheng W, Sakai H, Yagi K, Hasegawa T (2009) Interactions of elevated [CO2] and night temperature on rice growth and yield. Agric For Meteorol 149:51–58
Cramer W, Bondeau A, Woodward FI, Prentice IC, Betts RA, Brovkin V, Cox PM, Fisher V, Foley JA, Friend AD, Kucharik C, Lomas MR, Ramankutty N, Sitch S, Smith B, White A, Young-Molling C (2001) Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models. Glob Change Biol 7:357–373
Dansgaard W (1964) Stable isotopes in precipitation. Tellus 16:436–468
Dawson TE, Ehleringer JR (1991) Streamside trees that do not use stream water. Nature 350:335–337
Dingkuhn M, De Datta SK, Dorffling K, Javellana C (1989) Varietal differences in leaf water potential, leaf net CO2 assimilation, conductivity, and water use efficiency in upland rice. Aust J Agric Res 40:1183–1192
Ehleringer JR, Cooper TA (1988) Correlations between carbon isotope ratio and microhabitat in desert plants. Oecologia 76:562–566
Farquhar GD, Lloyd J (1993) Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere. In Ehleringer JR, Hall AE, Farquhar GD (eds) Stable isotopes and plant carbon-water relations. Academic, New York, pp 44–70
Farquhar GD, O’Leary MH, Berry JA (1982) On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust J Plant Physiol 9:121–137
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol Plant Mol Biol 40:503–507
Hanba YT, Kobayashi T, Enomoto T (2010) Variations in the foliar δ13C and C3/C4 species richness in the Japanese flora of Poaceae among climates and habitat types under human activity. Ecol Res 25:213–224
Hasegawa T, Ishimaru T, Kondo M, Kuwagata T, Yoshimoto M, Fukuoka M (2011) Spikelet sterility of rice observed in the record hot summer of 2007 and the factors associated with its variation. J Agric Meterol 67:225–232
Henry HAL, Cleland EE, Field CB, Vitousek PM (2005) Interactive effects of elevated CO2, N deposition and climate change on plant litter quality in a California annual grassland. Oecologia 142:465–473
Horie T, Nakagawa H, Ohnishi M, Nakano J (1995) Rice production in Japan under current and future climates. In: Matthews RB, Kropff MJ, Bachelet D, van Laar HH (eds) Modelling the impact of climate change on rice production in Asia. CAB International, Oxon, pp 143–164
IPCC (2007) Summary for policymakers. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL (eds) Climate change 2007: the physical science basis. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, pp 1–18
Japan Meteorological Agency (2005) Climate change monitoring report. http://www.data.kishou.go.jp/climate/cpdinfo/climate_change/
Japan Meteorological Agency (2012) Meteorological statistical information. http://www.jma.go.jp/jma/menu/report.html
Kondo M, Pablico PP, Aragones DV, Agbisit R (2004) Genotypic variations in carbon isotope discrimination, transpiration efficiency, and biomass production in rice as affected by soil water conditions and N. Plant Soil 267:165–177
Kume A, Satomura T, Tsuboi N, Chiwa M, Hanba YT, Nkane K, Horikoshi T, Sakugawa H (2003) Effects of understory vegetation on the ecophysiological characteristics of an overstory pine, Pinus densiflora. For Ecol Manag 176:195–203
Leuenberger MC, Filot MS (2007) Temperature dependencies of high-temperature reduction on conversion products and their isotopic signatures. Rapid Commun Mass Spectrom 21:1587–1598
Li Q, Nakatsuka T, Kawamura K, Liu Y, Song H (2011) Regional hydroclimate and precipitation δ18O revealed in tree-ring cellulose δ18O from different tree species in semi-arid Northern China. Chem Geol 282:19–28
Libby LM, Pandolfi LJ (1974) Temperature dependence of isotope ratios of tree rings. Proc Natl Acad Sci USA 71:2482–2486
Ma JW, Sun W, Sun HI, Wang SM (2012) Stable carbon isotope characteristics of desert plants in the Junggar Basin, China. Ecol Res 27:115–124
Masutomi Y, Takahashi K, Harasawa H, Matsuoka Y (2010) Impact assessment of climate change on rice production in Asia in comprehensive consideration of process/parameter uncertainty in general circulation models. Agric Ecosyst Environ 131:281–291
Matsuo N, Ojika K, Shuyskaya E, Radjabov T, Toderich K, Yamanaka N (2013) Responses of the carbon and oxygen isotope compositions of desert plants to spatial variation in soil salinity in Central Asia. Ecol Res 28:717–723
Matsushima S, Tsunoda K (1957) Analysis of development factors determining yield and yield prediction in lowland rice: on the mechanism of ripening (6). Effect of the range daily temperature in different stages in growth upon the ripening of rice plants (in Japanese with English abstract). Jpn J Crop Sci 25: 203–206
Matthews RB, Kropff MJ, Horie T, Bachelet D (1997) Simulating the impact of climate change on rice production in Asia and evaluating options for adaptation. Agric Syst 54:399–425
Ministry of Agriculture, Forestry and Fisheries (2012) Statistical report on agriculture, forestry, and fisheries. http://www.maff.go.jp/j/tokei/index.html
Mizota C, Kusakabe M (1994) Spatial distribution of δD-δ18O values of surface and shallow groundwaters from Japan, South Korea and east China. Geochem J 28:387–410
Morita S (2008) Prospect for developing measures to prevent high-temperature damage to rice grain ripening (in Japanese with English abstract). Jpn J Crop Sci 77: 1–12
Murchie EH, Chen Y-Z, Hubbart S, Peng S, Horton P (1999) Interactions between senescence and leaf orientation determine in situ patterns of photosynthesis and photoinhibition in field-grown rice. Plant Physiol 119:553–563
Nakashita R, Suzuki Y, Akamatsu F, Iizumi Y, Korenaga T, Chikaraishi Y (2008) Stable carbon, nitrogen, and oxygen isotope analysis as a potential tool for verifying geographical origin of beef. Anal Chim Acta 617:148–152
Peng S, Huang J, Sheehy JE, Laza RC, Visperas RM, Zhong X, Centeno GS, Khush GS, Cassman GS (2004) Rice yields decline with higher night temperature from global warming. Proc Natl Acad Sci USA 101:9971–9975
R Development Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Roden JS, Farquhar GD (2012) A controlled test of the dual-isotope approach for the interpretation of stable carbon and oxygen isotope ratio variation in tree rings. Tree Physiol 32:490–503
Rozanski K, Araguás-Araguás L, Gonfiantini R (1992) Relation between longterm trends of oxygen-18 isotope composition of precipitation and climate. Science 258:981–985
Samuel D, Silver BL (1965) Oxygen isotope exchange reactions of organic compounds. Adv Phys Org Chem 3:1885–1895
Saurer M, Aellen K, Siegwolf R (1997) Correlating δ13C and δ18O in cellulose of trees. Plant Cell Environ 20:1543–1550
Shimono H, Hasegawa T, Fujimura S, Iwama K (2004) Responses of leaf photosynthesis and plant water status in rice to low water temperature at different growth stages. Field Crops Res 89:71–83
Sternberg LdaSL (1989) Oxygen and hydrogen isotope ratios in plant cellulose: mechanisms and application. In: Rundel P, Ehleringer JR, Nagy K (eds), Applications of stable isotope ratios to ecological research. Ecological Series Monographs 62. Springer, New York, pp 124–141
Sternberg LdaSL, DeNiro MJ, Savidge RA (1986) Oxygen isotope exchange between metabolites and water during biochemical reactions leading to cellulose synthesis. Plant Physiol 82:423–427
Suzuki Y, Chikaraishi Y, Ogawa NO, Ohkouchi N, Korenaga T (2008) Geographical origin of polished rice based on multiple element and stable isotope analyses. Food Chem 109:470–475
Tao F, Hayashi Y, Zhang Z, Sakamoto T, Yokozawa M (2008) Global warming, rice production, and water use in China: developing a probabilistic assessment. Agric For Meteorol 148:94–110
Toft NL, Anderson JE, Nowak RS (1989) Water use efficiency and carbon isotope composition of plants in a cold desert environment. Oecologia 80:11–18
Werner RA, Kornexl BE, Roßmann A, Schmidt H-L (1996) On-line determination of δ18O values of organic substances. Anal Chim Acta 319:159–164
Xu Y, This D, Pausch RC, Vonhof WM, Coburn JR, Comstock JP, McCouch SR (2009) Leaf-level water use efficiency determined by carbon isotope discrimination in rice seedlings: genetic variation associated with population structure and QTL mapping. Theor Appl Genet 118:1065–1081
Yamakawa H, Hirose T, Kuroda M, Yamaguchi T (2007) Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol 144:258–277
Yoshimoto M, Oue H, Kobayashi K (2005) Energy balance and water use efficiency of rice canopies under free-air CO2 enrichment. Agric For Meteorol 133:226–246
Acknowledgments
We thank Dr. T. Yanagihara, Dr. S. Yanagihara, Dr. Y. Iizumi, Mr. D. Tabei, Mr. I. Yamagata, Mr. H. Yamaguchi, and members of Syokukyo CORP, Yaeyama Branch of the Okinawa Prefectural Agricultural Experiment Station, Japan Rice Millers Association, ZEN-NOH Pearl Rice East Japan Corp., and the Agricultural Cooperative Association for supporting the collection of certified rice and water samples. This work was supported by the Program for Promotion of Basic Research Activities for Innovative Biosciences (PROBRAIN).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Akamatsu, F., Suzuki, Y., Nakashita, R. et al. Responses of carbon and oxygen stable isotopes in rice grain (Oryza sativa L.) to an increase in air temperature during grain filling in the Japanese archipelago. Ecol Res 29, 45–53 (2014). https://doi.org/10.1007/s11284-013-1097-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-013-1097-y