
Abstract
Restoration and conservation of species-rich nature reserves requires inclusion of landscape-scale connections and transport processes such as hydrologic flows and species dispersal. These are important because they provide suitable habitat conditions and an adequate species pool. This study aimed at identifying the key hydrologic flows and plant dispersal processes affecting a landscape with species-rich fen reserves where restoration measures are carried out to set back succession. It also intended to use this information for delineating the area relevant for conservation planning on an Operational Landscape Unit map. The study was carried out for complexes of fen ponds in former turbaries in the Vechtplassen area, The Netherlands. A number of recent insights on plant dispersal were integrated with knowledge on hydrologic flows in the present approach. The results showed that groundwater discharge to ensure mesotrophic, base-rich conditions, should be enhanced by restoring the groundwater recharge areas NE of the reserves. A nearby lake with suitable water chemistry was also identified as a key source of surface water to feed the fens in dry periods. Water dispersal was identified as important within the fen reserves, whereas dispersal by daily migrating dabbling ducks, typically occurring over 2–3 km, was the most important route connecting the reserves with the surrounding landscape. The delineation of the Operational Landscape Unit for this region provides a basis for conservation and restoration that take fundamental landscape connections and transport processes into account. This unique approach simultaneously considers hydrological transport processes as well as species dispersal in the larger landscape beyond the reserves themselves and therefore leads to greater success of restoration and conservation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Longitudinal, shallow water bodies with only moderate water flow, such as old river branches or oxbows, are characterized by abundant vegetation of submerged and emergent aquatic plants. In regions with a considerable precipitation surplus, e.g. in northwestern Europe, such meso- to eutrophic aquatic systems are inhabited by an array of plant communities which together are among the most species-rich plant assemblages in the temperate zone. They are characterized by long-term succession, during which the water body gradually develops from open water with submerged vegetation via floating-leaved communities towards floating rafts and quaking fens, ultimately leading to forest with alder, willow and/or birch (Chiarello et al. 1998). These systems can be found naturally in oxbows of medium-sized rivers, which are formed by river scouring, become gradually detached from the river and then gradually fill up with vegetation and organic matter, often culminating in fen peat. The dynamic meandering behavior of the river will keep forming new oxbows on the floodplain, so that the complete series of plant communities will remain present in the river reach at large in a dynamic mosaic of water bodies in different successional stages (Amoros 2001; Bornette et al. 1998).
In many parts of the world, temperate rivers and their floodplains have been modified drastically, with dikes separating substantial parts of the floodplain from river flooding and the river bed channelized for navigation. This practically prevents the formation of new oxbows or other water bodies remote from the river influence. Therefore, the assemblages of plant communities associated with the succession from open water to terrestrialized fen have become increasingly rare. This has happened, for instance, in the floodplains of the rivers Rhine and Meuse in The Netherlands. On the other hand, peat dredging in the fen meadow areas in the Western Netherlands, starting in the 18th century, has resulted in landscapes with rectilinear, narrow and shallow fen ponds, often called ‘turf ponds’. These fen pond complexes now show a remarkable botanical richness with plant communities characteristic for the succession from open water to fen wetland (Van Wirdum et al. 1992; Verhoeven and Bobbink 2001).
Although these artificial fen ponds could be seen as a refuge for the plant assemblages that are characteristic for oxbow landscapes in meandering river basins, the pond complexes miss the inherently dynamic features of river landscapes: no new fen ponds are formed after peat dredging has stopped in the 1920s and progressive succession leads to the predominance of final stages, i.e. wetland forest and grazed or mown meadow habitats. The initial and mid-successional stages are now gradually disappearing from the landscape, which reduces overall landscape diversity and results in the disappearance of typical species such as Carex diandra, Carex lasiocarpa, Scorpidium scorpioides and Hammarbya paludosa (Verhoeven and Bobbink 2001; Verhoeven 2014). Nature conservation agencies in The Netherlands have, therefore, invested in the (re-)excavation of fen ponds for restoration of the open-water habitats suitable for early-succession plant communities. These new ponds have been created or restored in the past 30 years. First monitoring of their species composition has shown that colonization by plant species has been slow. In a larger study of vegetation development in restored fen ponds in various turbary complexes in The Netherlands, slow colonization was shown to be correlated with deteriorated water quality, isolated location of the ponds, absence of seed banks and herbivory by musk rats and geese (Sarneel et al. 2011). The issues of water quality and plant diaspore availability are strongly related to landscape-scale processes of hydrology and plant dispersal.
The study of landscape-scale flows has received much attention in wetland science. Wetland structure and functioning is strongly governed by hydrological processes at the landscape scale. The wider landscape setting and the spatial position of the wetland determine to what extent the wetland is fed by groundwater and/or surface water, which has strong consequences for the hydroperiod, the biogeochemical processes and species composition of flora and fauna in the wetland (Mitsch and Gosselink 2015). The importance of landscape features for wetland structure and functioning has led to several theoretical concepts that have given much insight in these often complex relations. The Hydrogeomorphic approach or HGM (Cole 2016; Whigham et al. 1999) has emphasized the position of wetlands in the landscape, particularly in relation to the consequences of this position for the types of water feeding the wetland, the water retention time and hydroperiod. The Hydrogeologic Setting framework or HGS has classified fens according to their hydrogeologic landscape position, which correlated strongly to their plant species composition (Godwin et al. 2002). These concepts have addressed the major role of hydrologic flows across the landscape for wetland structure and functioning, but have not included the effects of species dispersal processes.
For our analysis of the landscape-scale factors that might hamper the colonization of restored fen ponds, we want to address the role of hydrologic flows but also that of plant dispersal. We earlier have proposed the Operational Landscape Unit approach (OLU) to identify the landscape components that are indispensable for maintaining the essential hydrologic flows as well as the key plant dispersal processes (Verhoeven et al. 2008). We developed the OLU approach for a river floodplain area of the Dinkel, a Dutch lowland stream in a sandy moraine landscape targeted for restoration of flooding regime and vegetation. The OLU encompassed the floodplain and the stream channel from where flooding could be restored, but also identified an upstream nature reserve to be reconnected to the stream as a source of plant diaspores. The OLU approach gives managers a clear perspective on the areas beyond the restoration sites themselves which should be considered in a management plan. Here we want to use the OLU approach for the fen complexes in the Vechtplassen near Utrecht to identify such landscape-scale hydrological and plant dispersal processes in a completely different landscape of peat meadows, canals and species-rich turf ponds. This area contains ponds with late-succession vegetation as well as restored and newly created ponds where processes controlling hydrogeochemistry and vegetation succession have been studied extensively (Beltman et al. 2011). Furthermore, a number of new insights in the importance of the dispersal vectors water, wind and waterfowl operating at this landscape scale and arising from recent studies in this area will be applied in the OLU delineation.
In this way, we test the feasibility of the OLU approach, developed in a stream valley landscape, for complexes of fen systems, which have much more moderate hydrological dynamics and quite different landscape settings compared to lowland streams with floodplain habitats where the approach was first developed. We evaluate the opportunities and limitations of the approach coming out of these two cases.
Study area
Turf pond complexes in The Netherlands, mostly excavated below the water table in the eighteenth and early nineteenth century, are characterized by very species-rich fen vegetation. As many as 35 different vegetation types have been described for such habitats in The Netherlands (Van Wirdum et al. 1992). There are three different succession series, which each characterize a specific water chemistry in the open-water stage, i.e. brackish, fresh mesotrophic and fresh eutrophic. The succession goes through seven stages (aquatic, floating raft, helophyte, brownmoss, transitional fen and carr) and has been described by space-for-time substitution of an extensive database of vegetation relevées (Van Wirdum et al. 1992; Verhoeven and Bobbink 2001, Online Resource 1).
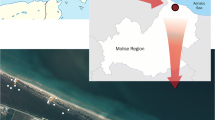
The eastern Vechtplassen area north of Utrecht, The Netherlands, contains turbaries in the polders Westbroek and Tienhoven (Fig. 1). This is an example of at least six major complexes of long, rectilinear ponds located in a zone between the upper sandy areas and the low coastal plain in The Netherlands (Beltman et al. 2011). The ponds have been created by peat dredging in the eighteenth–nineteenth centuries, and have filled up again with vegetation and peat since then. They occur in a peat-meadow landscape that had been reclaimed in the fourteenth–sixteenth centuries (Borger 1992). The turf ponds are still embedded in a landscape of peat meadows used for cattle grazing. The vegetation in such turf pond complexes is quite varied and species-rich.

Location of the Westbroek study area in The Netherlands and in the Loosdrechtse Plassen area. Topographic map
Historical setting
The species-rich plant communities in the terrestrializing fens in turf ponds are undergoing succession and can be attributed to the Phragmites and the brownmoss succession series, which are typically associated with mesotrophic and slightly eutrophic conditions (Van Wirdum et al. 1992; Verhoeven and Bobbink 2001). These conditions were quite common in The Netherlands until the 1950s and occurred in groundwater discharge areas and floodplain areas of streams and small rivers. Complexes of turf ponds with these terrestrializing fen systems can be found in The Netherlands where the Holocenic low fen areas border the higher sandy areas of Pleistocenic origin, e.g. Rottige Meente, De Wieden, De Weerribben and the Vechtplassen area.
In a study of historic aerial photographs of the 52-ha study area in the eastern Vechtplassen area spanning a 50-year time period (1937–1987), four successional stages were identified, i.e. open water, tall reed, short fen meadow and carr forest (Bakker et al. 1994). Many ponds in the Westbroek polder were still open water in 1937, but terrestrialized rather readily in a period of 20–40 years, with the share of open water decreasing and that of carr forest strongly increasing over time. By the end of the 1980s, the area had lost most of its early-succession stages, while some mid-succession species-rich fens were maintained by a mowing regime (Online Resource 2). These systems did, however, show signs of acidification, which is an indication of moving forward in succession to transitional fen, but was probably also enhanced because of atmospheric deposition of NHy and NOx (Paulissen et al. 2014). The State Forest Service, who owned and managed this fen area, decided to re-excavate a number of ponds to restore the early-succession vegetation types, so that the complete successional series would be safeguarded on the long term (Beltman et al. 1996). In the course of 30 years (1985–2015), at least 45 ponds were restored or created, strongly increasing the share of open-water ponds in the area. Initial monitoring activities indicated that recolonization of the ponds was slow and differed quite substantially between different ponds, which was attributed to dispersal issues, to changes in the water chemistry and to herbivory by invasive musk rats (Beltman et al. 2011; Sarneel et al. 2011).
Water flow and water chemistry
The Westbroek polder is a flat area with drained fenlands, situated close to a hill ridge of sandy moraines. The drainage occurs through parallel ditches about 50 m apart, which date back from the era of peatland reclamation in the fourteenth–fifteenth centuries (Borger 1992). The general direction of surface water flow, fed by rain water and groundwater discharge, is towards the south-west. Groundwater discharge from the sandy ridge toward the polder used to result in a water chemistry in the drainage ditches and the turf ponds which was favourable for the mesotrophic plant communities mentioned. The water discharging in the polder Westbroek from this aquifer is relatively low in nutrients, chloride and sulphate and rich in calcium and bicarbonate ions (Table 1).
However, these mesotrophic to moderately eutrophic systems became disturbed by polluted surface water since the 1960s. As groundwater resources in the hill ridge were increasingly used for drinking water abstraction, the amount of groundwater discharged in the polder decreased and more surface water supply was needed to keep water levels sufficiently high during dry periods in summer. This surface water originated from the Vecht river and, ultimately, from the Rhine and was polluted with nitrogen and phosphorus. It also had high concentrations of potassium, chloride and sulphate (Beltman et al. 1986; Beltman and Rouwenhorst 1991; Beltman and Verhoeven 1988; Van den Broek and Beltman 2000).
As a result of the surface water supply, the water chemistry in the polders Westbroek and Tienhoven changed considerably in a non-favourable direction, as can be seen in the composition of the supply water in the 1980s (Table 1). As the direction of surface water flow during such dry periods was reversed, i.e. towards the north-east, ditches the southern parts of both polders showed a strongly modified water chemistry with high sodium, potassium and chloride and low calcium and bicarbonate [Fig. 2, (Beltman and Rouwenhorst 1991)]. As a consequence, signs of deterioration of the quality of the submerged vegetation in ditches were observed in the early 1980s and there was increasing concern that the species-rich turf ponds would become negatively affected as well.

Water chemistry in the Westbroek polder ditches in summer 1983. The Stiff diagrams give the proportional concentrations as % of total electrical positive and negative charge. Water types dominated by Na, K and Cl occur the southern part of the polder, while water is relatively rich in Ca and HCO3 in the northern part (after Beltman and Rouwenhorst 1991)
The observed effects of the polluted river water in dry summer periods led to a decision by the water board to change the surface water source to feed the polder Westbroek in the early 1990s; the new source was the Breukeleveense Plas, a shallow lake just 1 km west of the polder (Fig. 3). This lake has good water quality which is chemically rather similar to the groundwater in the aquifer (see Table 1). The regional water regime controlled by pumping stations and weirs was modified in such a way that the water from the Breukeleveense Plas was directed towards the Westbroek polder in dry summer periods. This has improved the situation in the polder to the point that no signs of negative impacts on the submerged vegetation in the ditches are visible any longer.

The location of the groundwater recharge area feeding the aquifer which discharges in the Westbroek polder. The Breukeleveense Plas is used as a resource of surface area to be transported to Westbroek through canals during dry summer periods
This major change in the water management of the polder has prevented the precious plant communities in the turf ponds to be affected by the polluted river water. Table 1 shows that the water chemistry in the turf ponds remained favourable even in the 1980s, when the polluted water was penetrating the ditch system in dry summers. The slow flow rates in the exchange between turf ponds and ditches, as well as the timely measure of changing the surface water source to the polder have played a positive role (Beltman and Verhoeven 1988). The current water chemistry of the Westbroek turf ponds is characterized by low ammonium, phosphate and sulphate concentrations, which is favourable for the development of initial, submerged stages of the brownmoss succession series (Geurts et al. 2008).
Plant dispersal
The large number of plant species associated with the succession from open water to carr forest and the rapid turnover of these species during succession imply that dispersal to new or restored ponds is essential. In natural landscapes with oxbow lakes such dispersal would have been facilitated by the proximity of existing populations serving as propagule sources and by the ample availability of dispersal vectors water, wind and waterfowl. In the turf ponds, past dispersal capacities likely consisted also of dispersal by these same vectors, while the direct connections via surface water to nearby propagule sources compensated for the lack of flowing (river) water. In the current situation, where early-successional stages in the landscape are sparse and far apart, dispersal limitation has become an issue likely to limit the re-colonization of newly created ponds. Ponds that had terrestrialized before and are being re-excavated will have a better chance than newly created ponds to already contain diaspores left over in the seed bank of the shore or the pond bottom (Bekker et al. 1998, 1999; Klimkowska et al. 2010). However, due to excavation activities a large part of the existing seed bank is removed and colonization by dispersal in space also becomes important. Arrival of propagules to new habitats is often a time-consuming process, with species that possess traits enhancing long-distance dispersal arriving sooner and in greater numbers (Brederveld et al. 2011; Fraaije et al. 2015). However, also the distance between habitats and their connectivity plays an important role: nearby source populations facilitate colonization (Brederveld et al. 2011; Honnay et al. 2002). In the successional series relevant here, three main dispersal mechanisms determine the dispersal of plant species and the connectivity between habitat areas: dispersal by water, wind, and waterfowl.
For species characteristic of aquatic and shoreline vegetation, and for the early-succession, aquatic phases of the series, species dispersal by water is particularly important (Soons et al. 2017). Water is widely available as a dispersal vector, and, perhaps even more importantly, is likely to connect wet, suitable sites. Seeds of early-succession species may float for months, particularly in stagnant or slow-flowing water (Van den Broek et al. 2005). Vegetative fragments may float for much shorter periods of time, but may be highly effective in colonizing new areas (Boedeltje et al. 2003, 2004, 2008). A disadvantage of the turbary system in regard to water dispersal is that the water in the system hardly flows—the system resembles a set of interconnected lakes rather than a stream. Hence, dispersal by water is very slow and directed by wind, pushing the floating seeds downwind, particular in ditches and linear ponds (Sarneel et al. 2014; Soomers et al. 2013a). It has been shown that the orientation of the fen pond complexes towards the prevailing wind direction may affect dispersal distances to a considerable degree (Soomers et al. 2013a). The fens in our study area are orientated in a NE→SW direction, so that wind- enhanced hydrochorous dispersal is expected to be quite effective considering the prevailing south-westerly winds. Eventually, water-dispersed seeds in such systems are likely to be deposited at (northeastern) shorelines at times of receding water levels in spring, followed by a rapid germination and locally high seedling densities (Sarneel and Soons 2012; van Leeuwen et al. 2014). Another limiting factor to seed dispersal by water is that it only reaches sites that are connected via surface water so that dispersal between different catchments or hydrological units is not possible. The scale at which dispersal by water takes place is therefore highly site-specific, and dependent on the hydrological connections by surface water and the flow velocity and direction of the system.
Dispersal by wind is a common phenomenon in particular for later successional species. Increasingly terrestrial stages have concomitantly higher proportions of species with traits facilitating long-distance dispersal by wind (Soons 2006). The advantage of this mechanism is that wet, unsuitable patches can be bridged and suitable sites across the water can be connected (Soons et al. 2017). In addition, this dispersal mechanism is not limited to catchment boundaries or hydrological units. A disadvantage is that this process is not limited to wetlands but rather distributes seed across the landscape, so that dispersal is not directed towards suitable sites (Sarneel et al. 2014; Soomers et al. 2013b; Soons 2006). The scale at which dispersal by wind takes place is much less site-specific than for water dispersal, and much more dependent on the traits of the species dispersing: plant species that release their seeds from a great height and produce very light seeds (with a low terminal velocity) may disperse their seeds over hundreds to thousands of meters (Soons et al. 2004a, b; Soons and Ozinga 2005). Typical emergent species such as Phragmites and Typha, but also carr forest trees (Alnus, Betula, Salix) disperse very well by wind.
Dispersal by animals is the third relevant dispersal vector. As there are no large herbivores playing a significant role in the dispersal of plant seeds in these turbaries, the main groups of seed dispersing animals are fish and waterfowl. Fish are known to ingest and disperse seeds (Boedeltje et al. 2015; van Leeuwen et al. 2012), but the velocity of dispersal is limited in the same way as dispersal by water: only sites connected by surface water can be connected by this seed dispersal mechanism. Waterfowl therefore play the more important role: they are numerous, seasonally granivorous, while seeds survive gut passage in relatively great numbers, and often show daily migrations, thereby crossing hydrological or other landscape barriers (Kleyheeg et al. 2015, 2017b). Among the waterfowl frequenting the lakes, ponds and canals in the peat meadow areas there are large numbers of dabbling ducks, which have been shown to disperse seeds of a wide range of wetland and terrestrial plant species (Soons et al. 2016). The most common waterfowl species in the area is the mallard (Anas platyrhynchos). This species has a characteristic behavior of diurnal migrations between roosting sites (day) and foraging sites (night). In this way, they disperse large numbers of seeds between their subsequent nocturnal feeding sites (Kleyheeg et al. 2017a). The scale of this dispersal is again dependent on the landscape: in landscapes where more open water is present, travelling distances (and, hence, dispersal distances) are shorter than in landscapes with more scattered water bodies. In peat districts with much open water the scale is on the order of magnitude of a few km (Kleyheeg et al. 2017a).
OLU delineation
By delineating an OLU we want to map the source areas of the landscape-scale transport flows of water and plant diaspores which are essential for the conservation of the terrestrializing vegetation in the 52-ha turf pond complex in the Westbroek polder. It is first important to indicate the relevant spatial scale. In Fig. 1 the red line delineates this collection of 43 turf ponds, some of which are connected to one or more neighbouring ponds. For the OLU, however, we look beyond the spatial relations within this complex and emphasize the regional sources of high-quality water and of plant diaspores and the associated transport routes towards the complex. These spatial connections are the groundwater flows in the sandy aquifer below the 1–2 m peat layer, the regulated, slow surface water flows through canals and the seed movements through water, wind and waterfowl dispersal. These fluxes are realistically taking place within a perimeter of 10 km around the Westbroek turf ponds (see the red circle in Fig. 3).
The source areas of water with favourable chemistry encompass groundwater recharge areas which are connected to the aquifer feeding the turf pond complex, as well as the freshwater lake which is the source of surface water supplied to the pond complex during dry summer periods. The origin of the groundwater discharging in the Westbroek polder is primarily located in the recharge area in the higher sandy terrain just east of the polder (see the hatched area in Fig. 3, based on Beltman and Grootjans 1986; Schot and Molenaar 1992). The source area of suitable surface water in dry summers, i.e. the lake Breukeleveense Plas, is also marked on this map.
For the identification of source areas of plant diaspores, it is critical to quantify and map the presence of target species characteristic for the plant communities in the terrestrializing fen succession. Figure 4 indicates which km squares in the 10-km perimeter around the turf ponds contain at least one of 33 target species, as based on the national flora data base of The Netherlands (FLORON, Beltman et al. 2011, see Online Resource 3). It is clear that target species occur mostly in the northern and western parts of the area under consideration. The connections from these areas toward the pond complex via water are very limited, and water dispersal can be considered insignificant at this scale. The major dispersal route for plant diaspores of these species is through wind or transport by dabbling ducks. Wind dispersal distances are in general shorter and less directed than dispersal distances secured via waterfowl moving in between wetlands. Mallards are very common in the area and have daily migration patterns between roosting areas and feeding areas. The range of these migrations in similar peat landscapes is limited to ca. 2.5 km (Kleyheeg et al. 2017a). All water bodies and wetlands within 2.5 km of the Westbroek turf pond complex are indicated as dark blue patches on Fig. 4. All these water bodies are located in km squares with at least one target species.

Map combining all spatial information needed to delineate the Operational Landscape Unit with the OLU boundary shown by the black line. Distance of water bodies to the fen reserve area (‘Westbroek’) and km squares with at least one of 33 target species within 10 km of the area. The water bodies within 2500 m of the fen reserve are within the diurnal migration zone of dabbling ducks
Figure 4 also shows the delineation of the Operational Landscape Unit for the Westbroek turf ponds as a black line. In the west and south this demarcation line follows location of the water bodies within 2.5 km of the turf pond complex and includes the Breukeleveense Plas, which is a major source of high-quality surface water. The groundwater recharge area is included in the eastern part of the OLU and its boundary delineates the OLU there.
Discussion
The delineation of the Operational Landscape Unit in the Eastern Vechtplassen has clarified which areas outside the 52-ha complex of Westbroek turf ponds play a major role as sources of high-quality water and of plant diaspores, which are both indispensable for the conservation of the high botanical richness in Westbroek on the long term. There is considerable interest in the protection of this valuable wetland area and the State Forestry Service has invested quite a lot in management and restoration measures in the area itself. Mowing and clear-cutting regimes have been installed, ponds have been re-excavated or newly dug, and attention has been paid to water level management and connections between ponds. The OLU delineated here helps to ensure that connections with a number of important landscape elements beyond these reserves are maintained and/or restored as well.
The landscape-scale processes addressed in the OLU are the transports of water and plant diaspores which typically take place at a scale of 10–20 km. The hydrological processes are strongly related to the hydrogeomorphic setting of the wetlands and encompass groundwater flows between groundwater recharge and discharge areas as well as human-controlled surface water flows. The groundwater discharge in the turf pond complex is essential because it ensures the water chemistry required for high botanical richness. The OLU visualizes the groundwater recharge area and aquifer which should be protected from too intensive drinking water extraction which diminishes the discharge in the ponds. The surface water levels in the polder areas are completely controlled and result in slow flows among canals and water bodies. The OLU also visualizes the Breukeleveense Plas as a source of high-quality water which can be brought to the fen complex by keen surface water management. Surprisingly, surface water flows at this scale have only a minor role for plant dispersal by water in this region, because water flows are very slow and the various polders have water systems which are relatively isolated. Recent studies have revealed that the dispersal by dabbling ducks, particularly mallards, is the key factor for plant dispersal at this scale. No other common waterfowl species in the area have similar diurnal movements between roosting and feeding habitats. The range of the dabbling duck movements (2–3 km) was leading for the delineation of water bodies to include in the OLU.
As the management of the wetland areas, the surface water levels and the drinking water resources around the turf pond complex is carried out by different agencies, e.g. the State Forest Service, the water board (Waternet) and the drinking water company (Vitens), respectively, it is appropriate to try to link management plans and long-term objectives.
This case study has applied the OLU approach to a lowland area with species-rich fen ponds in a landscape setting of polders with controlled surface water hydrology located close to a sandy hill ridge. This area functions in a totally different way than the stream valley in the eastern Netherlands for which the OLU was first conceptualized and applied (Verhoeven et al. 2008). In that valley, the water flow in the stream was inherently orders of magnitude faster than in the canals and ditches in the Vechtplassen polders and much more variation in discharge occurred, leading to opportunities for restoration of flooding of a targeted former floodplain. In the delineated OLU, the targeted floodplain, a remote wetland reserve and the stream in between were included and the emphasis was on the reconnection of both the reserve and the former floodplain to the river, so that the hydrological functioning and the hydrochorous seed dispersal in the floodplain would be restored.
The OLU delineations in these two totally different wetland complexes targeted for restoration embedded in different landscape settings were both instrumental in identifying source areas and transport routes of water and plant diaspores beyond the targeted areas themselves. The innovative character of the OLU is that it addresses simultaneously two different sets of landscape-scale transport processes, i.e., hydrological flows and plant dispersal. These two types of transport processes are much more important than other flows at the landscape scale for creating the suitable conditions in terms of colonization potential in the restored or created wetlands in the areas that we studied so far. In other areas, point sources of airborne pollution could also be considered for inclusion in the OLU, but then in the sense that they would need to be sanitized. Our OLU approach is different from earlier landscape concepts which either addressed hydrogeomorphic features (e.g. the Hydrogeomorphic approach, or the Hydrogeologic setting, (Cole 2016; Godwin et al. 2002; Whigham et al. 1999), or landscape connectivity and patch-corridor relations (Alagador et al. 2012; Janssen et al. 2013; Nilsson et al. 2010).
The exercise of delineating OLUs has shown to be useful to recognize transport processes at the landscape scale beyond the area of nature reserves. The OLU delineation brings together information from different experts (wetland ecologists, hydrologists, landscape ecologists) in an iterative process. It enables agencies responsible for management and restoration to recognize which of these areas need to be considered in the management plan and for what reason. The priority of taking measures and the resources involved could then be determined and implemented in a more optimal way. In the case of the Westbroek turf ponds, particularly because so much has been invested in a long-term plan of managing the ponds themselves, it is appropriate to ensure that high-quality water and a large collection of plant diaspores continue to be delivered to the pond complex from the regional sources in the vicinity.
References
Alagador D, Trivino M, Cerdeira JO, Bras R, Cabeza M, Araujo MB (2012) Linking like with like: optimising connectivity between environmentally-similar habitats. Landsc Ecol 27:291–301
Amoros C (2001) The concept of habitat diversity between and within ecosystems applied to river side-arm restoration. Environ Manage 28:805–817
Bakker SA, Van den Berg NJ, Speleers BP (1994) Vegetation transitions of floating wetlands in a complex of turbaries between 1937 and 1989 as determined from aerial photographs with GIS. Plant Ecol 114:161–167
Bekker RM, Schaminee JHJ, Bakker JP, Thompson K (1998) Seed bank characteristics of Dutch plant communities. Acta Botanica Neerlandica 47:15–26
Bekker RM, Lammerts EJ, Schutter A, Grootjans AP (1999) Vegetation development in dune slacks: the role of persistent seed banks. J Veg Sci 10:745–754
Beltman B, Grootjans AP (1986) Distribution of nutrient-poor plant communities in relation to the groundwater regime and nutrient availability. CHO-TNO Proc Inform 34:59–80
Beltman B, Rouwenhorst TG (1991) Ecohydrology and fen plant-distribution in the Vechtplassen Area, the Netherlands. In: Nachtnebel HP (ed) Hydrological basis of Ecologically sound management of soil and groundwater. IAHS Publication, Wallingford
Beltman B, Verhoeven JTA (1988) Distribution of fen plant communities in relation to hydrochemical characteristics in the Vechtplassen area. In: Verhoeven JTA (ed) Structure of vegetation in relation to carbon and nutrient economy. SPB Academic Publischers, The Hague, pp 121–136
Beltman B, Bouter P, Rouwenhorst G (1986) Kationenuitwisseling bij toevoeging van ammonium aan veenbodem. Utrecht Plant Ecol News Rep 6:17–28
Beltman B, Van den Broek T, Van Maanen K, Vaneveld K (1996) Measures to develop a rich-fen wetland landscape with a full range of successional stages. Ecol Eng 7:299–314
Beltman B, Omtzigt NQ, Vermaat JE (2011) Turbary restoration meets variable success: does landscape structure force colonization success of wetland plants? Restor Ecol 19:185–193
Boedeltje G, Bakker JP, Bekker RM, Van Groenendael JM, Soesbergen M (2003) Plant dispersal in a lowland stream in relation to occurrence and three specific life-history traits of the species in the species pool. J Ecol 91:855–866
Boedeltje G, Bakker JP, Ten Brinke A, Van Groenendael JM, Soesbergen M (2004) Dispersal phenology of hydrochorous plants in relation to discharge, seed release time and buoyancy of seeds: the flood pulse concept supported. J Ecol 92:786–796
Boedeltje G, Ozinga WA, Prinzing A (2008) The trade-off between vegetative and generative reproduction among angiosperms influences regional hydrochorous propagule pressure. Glob Ecol Biogeogr 17:50–58
Boedeltje G, Spanings T, Flik G, Pollux BJA, Sibbing FA, Verberk WCEP (2015) Effects of seed traits on the potential for seed dispersal by fish with contrasting modes of feeding. Freshw Biol 60:944–959
Borger GJ (1992) Draining-digging-dredging; the creation of a new landscape in the peat areas of the low countries. In: Verhoeven JTA (ed) Fens and bogs in The Netherlands: vegetation, history, nutrient dynamics and conservation. Kluwer Academic Publishers, Dordrecht, pp 131–172
Bornette G, Amoros C, Piegay H, Tachet J, Hein T (1998) Ecological complexity of wetlands within a river landscape. Biol Conserv 85:35–45
Brederveld RJ, Jaehnig SC, Lorenz AW, Brunzel S, Soons MB (2011) Dispersal as a limiting factor in the colonization of restored mountain streams by plants and macroinvertebrates. J Appl Ecol 48:1241–1250
Chiarello E, Amoros C, Pautou G, Jolion JM (1998) Succession modeling of river floodplain landscapes. Environ Model Softw 13:75–85
Cole CA (2016) HGM: a call for model validation. Wetlands Ecol Manage 24:579–585
Fraaije RG, ter Braak CJ, Verduyn B, Verhoeven JT, Soons MB (2015) Dispersal versus environmental filtering in a dynamic system: drivers of vegetation patterns and diversity along stream riparian gradients. J Ecol 103:1634–1646
Geurts JJM, Smolders AJP, Verhoeven JTA, Roelofs JGM, Lamers LPM (2008) Sediment Fe: PO4 ratio as a diagnostic and prognostic tool for the restoration of macrophyte biodiversity in fen waters. Freshw Biol 53:2101–2116
Godwin KS, Shallenberger JP, Leopold DJ, Bedford BL (2002) Linking landscape properties to local hydrogeologic gradients and plant species occurrence in minerotrophic fens of New York State, USA: a hydrogeologic setting (HGS) framework. Wetlands 22:722–737
Honnay O, Bossuyt B, Verheyen K, Butaye J, Jacquemyn H, Hermy M (2002) Ecological perspectives for the restoration of plant communities in European temperate forests. Biodivers Conserv 11:213–242
Janssen R, Arciniegas GA, Verhoeven JTA (2013) Spatial evaluation of ecological qualities to support interactive land-use planning. Environ and Plann B-Plann Des 40:427–446
Kleyheeg E, van Leeuwen CHA, Morison MA, Nolet BA, Soons MB (2015) Bird-mediated seed dispersal: reduced digestive efficiency in active birds modulates the dispersal capacity of plant seeds. Oikos 124:899–907
Kleyheeg E, Treep J, De Jager M, Nolet B, Soons MB (2017a) Seed dispersal distributions resulting from landscape-dependent daily movement behaviour of a key vector species. J Ecol
Kleyheeg E, van Dijk JGB, Nolet BA, Tsopoglou-gkina D, Woud T, Boonstra D, Soons MB (2017b) Daily movement distances and home range sizes of mallards (Anas platyrhynchos) are strongly affected by landscape configuration. J Ecol
Klimkowska A, Van Diggelen R, Grootjans AP, Kotowski W (2010) Prospects for fen meadow restoration on severely degraded fens. Perspect Plant Ecol Evol Syst 12:245–255
Mitsch WJ, Gosselink JG (2015) Wetlands. Wiley, London, New York., p 456
Nilsson C, Brown RL, Jansson R, Merritt DM (2010) The role of hydrochory in structuring riparian and wetland vegetation. Biol Rev 85(4):837–858
Paulissen MP, Schaminee JH, During HJ, Wamelink G, Verhoeven JT (2014) Expansion of acidophytic late-successional bryophytes in Dutch fens between 1940 and 2000. J Veg Sci 25:525–533
Sarneel JM, Soons MB (2012) Post-dispersal probability of germination and establishment on the shorelines of slow-flowing or stagnant water bodies. J Veg Sci 23:517–525
Sarneel JM, Soons MB, Geurts JJ, Beltman B, Verhoeven JT (2011) Multiple effects of land-use changes impede the colonization of open water in fen ponds. J Veg Sci 22:551–563
Sarneel JM, Beltman B, Buijze A, Groen R, Soons MB (2014) The role of wind in the dispersal of floating seeds in slow-flowing or stagnant water bodies. J Veg Sci 25:262–274
Schot PP, Molenaar A (1992) Regional changes in groundwater-flow patterns and effects on groundwater composition. J Hydrol 130:151–170
Soomers H, Karssenberg D, Soons MB, Verweij PA, Verhoeven JTA, Wassen MJ (2013a) Wind and water dispersal of wetland plants across fragmented landscapes. Ecosystems 16:434–451
Soomers H, Karssenberg D, Verhoeven JTA, Verweij PA, Wassen MJ (2013b) The effect of habitat fragmentation and abiotic factors on fen plant occurrence. Biodivers Conserv 22:405–424
Soons MB (2006) Wind dispersal in freshwater wetlands: knowledge for conservation and restoration. Appl Veg Sci 9:271–278
Soons MB, Ozinga WA (2005) How important is long-distance seed dispersal for the regional survival of plant species? Divers Distrib 11:165–172
Soons MB, Heil GW, Nathan R, Katul GG (2004a) Determinants of long-distance seed dispersal by wind in grasslands. Ecology 85:3056–3068
Soons MB, Nathan R, Katul GG (2004b) Human effects on long-distance wind dispersal and colonization by grassland plants. Ecology 85:3069–3079
Soons MB, Brochet AL, Kleyheeg E, Green AJ (2016) Seed dispersal by dabbling ducks: an overlooked dispersal pathway for a broad spectrum of plant species. J Ecol 104:443–455
Soons MB, de Groot GA, Cuesta Ramirez MT, Fraaije RGA, Verhoeven JTA, de Jager M (2017) Directed dispersal by an abiotic vector: wetland plants disperse their seeds selectively to suitable sites along the hydrological gradient via water. Funct Ecol. doi:10.1111/1365-2435.12771
Van den Broek T, Beltman B (2000) Natuurontwikkeling in het Noorderpark—5 jaar monitor onderzoek in nieuw gegraven petgaten. Faculteit Biologie, Onderzoeksgroep Landschapsoecologie, Utrecht, p 63
Van den Broek T, Van Diggelen R, Bobbink R (2005) Variation in seed buoyancy of species in wetland ecosystems with different flooding dynamics. J Veg Sci 16:579–586
van Leeuwen CH, Tollenaar ML, Klaassen M (2012) Vector activity and propagule size affect dispersal potential by vertebrates. Oecologia 170:101–109
van Leeuwen CHA, Sarneel JM, van Paassen J, Rip WJ, Bakker ES (2014) Hydrology, shore morphology and species traits affect seed dispersal, germination and community assembly in shoreline plant communities. J Ecol 102:998–1007
Van Wirdum G, Den Held AJ, Schmitz M (1992) Terrestrializing fen vegetation in former turbaries in The Netherlands. In: Verhoeven JTA (ed) Fens and bogs in The Netherlands: vegetation, history, nutrient dynamics and conservation. Kluwer Academic Publishing, Dordrecht, pp 323–360
Verhoeven JTA (2014) Wetlands in Europe: perspectives for restoration of a lost paradise. Ecol Eng 66:6–9
Verhoeven JTA, Bobbink R (2001) Plant diversity of fen landscapes in The Netherlands. In: Gopal B (ed) Biodiversity in wetlands: assessment, function and conservation. Backhuys Publishers, Leiden, pp 65–87
Verhoeven JTA, Soons MB, Janssen R, Omtzigt N (2008) An Operational Landscape Unit approach for identifying key landscape connections in wetland restoration. J Appl Ecol 45:1496–1503
Whigham DF, Lee LC, Brinson MM, Rheinhardt RD, Rains MC, Mason JA, Kahn H, Ruhlman MB, Nutter WL (1999) Hydrogeomorphic (HGM) assessment—a test of user consistency. Wetlands 19:560–569
Acknowledgements
The authors would like to thank Nancy Omtzigt (Geodan BV) and Jasper Dekkers (Vrije Universiteit Amsterdam) for their help in preparing the maps. Dr. Winnie Rip (Waternet, Amsterdam) kindly provided recent data on water chemistry for the various water bodies.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Verhoeven, J.T.A., Beltman, B., Janssen, R. et al. Delineating landscape-scale processes of hydrology and plant dispersal for species-rich fen conservation: the Operational Landscape Unit approach. Wetlands Ecol Manage 25, 761–774 (2017). https://doi.org/10.1007/s11273-017-9549-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-017-9549-8