
Abstract
Ferns and lycophytes are excellent model organisms to investigate the effect of climate change on species niches, due to their high sensitivity to changes in temperature and precipitation. In Honduras, this species group is particularly diverse with over 713 known taxa. However, we currently lack a quantitative assessment of the impact that changes in climate might have on these species. This study aimed (1) to explore the structure of the available climatic conditions across the entire country; (2) to determine the extent to which the available climatic space is occupied by different fern and lycophyte species; and (3) to assess which regions of climatic space will be most affected according to scenarios of climate change. We analyzed a newly developed database of herbarium fern records (n = 11,791) and used ordination models to compare the climatic niche of all Honduran fern and lycophyte taxa under present and future climates. We found that species were distributed throughout most of the available climatic space. Under RCP2.6 and RCP8.5 projections 128 to 391 species will have their average niche positions outside of the predicted available climatic space by the years 2049–2099. Particularly affected will be species that grow in low temperature and high precipitation areas, which represent high altitude parks, such as Celaque National Park. Epiphytes showed smaller variance in their climatic niches than terrestrial species. However, terrestrial species showed higher loss in climate space (e.g., 82.9% vs. 17.1% for RCP8.5) and are more likely to be at risk in the future. In general, Honduras is expected to become drier, with more extreme events of severe drought. Considering that ferns are particularly sensitive to changes in climate, if tested in the future, they could be used as a proxy for other Honduran plant taxa.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The climatic niche—i.e., the climatic conditions associated with the occurrence of a given species—has come to the forefront of much of recent ecological and evolutionary research, given that it provides a natural intersection of many distinct fields, from ecophysiology and invasion biology to biogeography and macroecology (Petitpierre et al. 2012; Khaliq et al. 2015; Wasof et al. 2015; Atwater et al. 2018). For instance, climatic niches are at the core of the concept of phylogenetic niche conservatism (Wiens and Donoghue 2004), which has been instrumental to understand the biogeographical distributions of a variety of taxa, as well as the origin of latitudinal gradients of species diversity (e.g., Buckley et al. 2010; Kerkhoff et al. 2014). Instead of considering all biotic and abiotic dimensions that can be part of Hutchinson’s ecological niche as an n-dimensional hypervolume (Hutchinson 1957), the focus on a subset of climatic variables that make up the climatic niche is both practical and heuristic. The increasing availability of high-quality climatic layers in recent years, including past, present, and future climatic conditions (Brown et al. 2018; Karger et al. 2017; Sugiyama et al. 2010), has allowed for the first time in recent decades to readily analyze climate data across large geographical regions. In addition, the emphasis on abiotic conditions can provide an overall framework against which biotic conditions such as ecological interactions can be tested against. Indeed, the preeminence of the study of climatic niches is likely to increase in the near future, as we seek to understand how species will respond to the challenges imposed by anthropogenic climate change (Quintero and Wiens 2013; Jezkova and Wiens 2016; Herrera et al. 2018; Meyer and Pie 2022).
A common way in which studies on climatic niche evolution are carried out involves extracting bioclimatic conditions associated with occurrence data of a set of species, calculating species means and then use ordination patterns to understand the structure of interspecific variation in their climatic niches (e.g., Kamilar and Muldoon 2010; Duran and Pie 2015; Ramírez-Barahona et al. 2016; Pie et al. 2017). Although it has provided important insight into how climatic niches evolve in different lineages, one important limitation of this approach is that it does not explicitly take into account the fact that climatic conditions are not isotropic. Rather, some combinations of climatic conditions might be more common in geographical space, whereas other combinations might simply not occur in nature. Such variation in the relative commonness of environmental conditions (environmental prevalence, sensu Meyer and Pie 2018) can constrain the climatic niches that different species are able to occupy. For instance, interspecific variation in climatic niche breadth might ultimately be the result of the prevalence of different combinations of climatic variables rather than physiological tolerances (Pie et al. 2021). Therefore, it is important that climatic niches should be interpreted with respect to the actual distribution of climatic niche space in which they occur.
Ferns and lycophytes are important components of global plant diversity. Current data suggest that there are approximately 13,000 fern and lycophyte taxa globally (PPGI 2016). They are particularly diverse in the humid tropics (Suissa et al. 2021), making up around 8.2% (1,358) of all vascular plants in Central America (16,335) (Moran and Riba 1995; Ulloa et al. 2017), and in some cases can account for a very large proportion of species in forest inventories (Linares-Palomino et al. 2009). It is believed that approximately 25% of ferns are epiphytes (i.e., plants that live on other plants for physical support but are not parasitic) and it has been shown that they are the third most species-rich group of vascular epiphytes globally (Zotz 2013). In Mesoamerica, for example, the most species-rich countries for ferns are Costa Rica, Mexico, and Panama (Reyes et al. 2021a). Given the prevalence of reproduction through spores, ferns in particular tend to be excellent dispersers and therefore are often believed to be more likely to occur whenever there are suitable conditions for their establishment and growth (although this notion has yet to be tested experimentally, Wild and Gagnon 2005). For instance, dispersal by wind in ferns and lycophytes can vary greatly between taxa but many species are known to have remarkable long-distance dispersal abilities (Barrington 1993; Wolf et al. 2001) and thus are often able to overcome geographical barriers more easily than other taxonomic groups (Tyron 1970). However, our understanding of how fern and lycophyte lineages evolved to occupy their current climatic niches is still incipient (Bystriakova et al. 2011; Hernandez-Rojas et al. 2021). Previous evidence suggests that epiphyte and terrestrial species richness respond differently to environmental gradients (Aros-Mualin et al. 2021), indicating that they might show important differences in their climatic niches, yet the extent of those differences is poorly understood (Nitta et al. 2020). The more restricted growth of epiphytic fern and lycophyte species, for example, makes them more susceptible to changes in atmospheric conditions (Williams et al. 2020; Hernandez-Rojas et al. 2021) and it has been shown that they are also more likely to be at higher risk under future changes in climate (Reyes et al. 2021b). For example, climate variables that are believed to be particularly important to epiphyte diversity are high temperature, water availability (precipitation/humidity), constant cloud cover, and low seasonality (Kromer 2013; Hernandez-Rojas et al. 2020).
In this study we use a large dataset of carefully curated occurrence records of ferns and lycophytes of Honduras to understand the relationship between their climatic niches and their available niche space. Our goals were (1) to use high-resolution climatic layers of temperature and precipitation variables to describe the structure of the available climatic conditions across the entire country; (2) to determine the extent to which the available climatic space is effectively occupied by different fern and lycophyte species, both with respect to their taxonomy and growth habit; and (3) to assess which regions of the available climatic space will be most affected according to scenarios of climate change.
Methods
Occurrence data
Fern and lycophyte data from Honduras were extracted from Batke et al. (2022). Their database held a total of 22,194 herbarium records from the years 1870 to 2018. Following the removal of duplicate entries and entries that could not be assigned a spatial reference, we retained 17,539 records. We taxonomically updated all records by cross-referencing all 17,539 records manually using Reyes et al. (2021a) updated checklist of Honduran ferns and lycophytes. The taxonomic validation resulted in 356 taxonomic changes (e.g., including moving species into different families or replacing species due to synonymy). A further 445 records were identified as invalid and were excluded (e.g., species that do not have a valid name). Moreover, we omitted any records with missing epithet information. Following the exclusion of all of these records, we retained a total of 11,791 (53% of the original database) for further analysis.
Growth form classification
We used Zotz et al. (2021) new database of vascular epiphytes and cross-referenced it against our database to separate epiphytes from terrestrial species in our analysis. We manually checked each record to ensure that species were assigned the correct growth form classification, based on published data (e.g., Batke et al. 2016; Reyes et al. 2021b) and field experience by the authors. Epiphytes were defined following Zotz (2013) and Zotz et al. (2021).
Climatic variables
We obtained climate data from Racines et al. (2018a), which include gridded climate surfaces for Honduras (30-year average) calculated from weather station observations from different official sources (local, national, and regional institutions) for the period between 1981 and 2010 at a spatial resolution of 30 s (~ 1 km2). The entire dataset includes 85 variables, including monthly precipitation (prec), monthly minimum temperature (tmin), maximum temperature (tmax), mean temperature (tmean), and diurnal temperature range (dtr), as well as seasonal and annual rasters derived based on the monthly datasets. However, to reduce collinearity in the dataset, we only included annual and seasonal means for those variables. We then carried out a Principal Component Analysis based on a covariance matrix of the remaining 25 bioclimatic variables, which were scaled and centered (i.e., divided and subtracted by the corresponding means and variances, respectively) prior to the analysis. The number of PCs retained for further analyses was based on a broken-stick criterion, as implemented in ‘vegan’ 2.5–7 (Oksanen et al. 2020). We then extracted bioclimatic data from fern and lycophyte occurrence records using the extract function in ‘raster’ 3.4–13 (Hijmans 2021) and projected them onto the principal components calculated previously. To further explore the extent of occupation of different species across the available climate space, we compared the variance in the scores for different ordination quadrants using Levene’s test, as implemented in ‘car’ 3.0–11 (Fox and Weisberg 2019).
Finally, we explored how expected shifts due to climate change could affect the structure of the available climatic space using the predictions by Racines et al. (2018b). We used RCP2.6 and RCP8.5 IPCC projections, as these represent the best case and worst-case climate forcing scenarios. RCP2.6 projects forecasts under the assumption that atmospheric CO2 concentration will stabilize by the year 2020 and reduce by the year 2100, keeping global temperature rise below 2 °C by 2100. RCP8.5 has been referred to as the worst-case scenario and assumes that emissions continue to rise throughout the twenty-first century. We obtained the values on each grid cell of the same set of climatic variables obtained above and projected them based on the PCA built from the current climate data. We compared the extent of the corresponding climatic spaces by estimating concave hulls, as implemented in ‘concaveman’ 1.1.0 (Gombin et al. 2020). This approach provides a convenient way to assess which regions of climatic space will be most affected by climate change. All analyses were carried out in R 4.1.1 (R Core Team 2021).
Results
The PCA was efficient in summarizing the variation in climatic conditions across Honduras, with the first two PCs explaining over 91% of the variance in the dataset. The loadings of the PCA are shown in Table 1. PC1 mostly reflected variation in temperature, especially lower temperatures, whereas PC2 loadings described a trade-off between maximum temperature and diurnal temperature range versus precipitation (particularly during autumn and winter). In other words, PC1 describes overall variation in temperature, whereas PC2 can be interpreted as describing relative aridity, with high PC2 scores indicating regions with high maximum temperatures and low precipitation (Fig. 1).

Principal component analysis of climatic conditions found in Honduras. All climatic conditions are shown in gray circles. Occurrence of different fern and lycophyte species across the available climate space for A the most species-rich families and B divided into epiphyte and terrestrial life forms. Closed circles represent species means, whereas black open circles indicate individual records. The arrows on A describe the general climate space (see text for details)
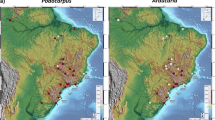
In general, species were distributed throughout most of the available climatic space, indicating that the climatic conditions found throughout Honduras in general tend to be suitable to fern and lycophyte species (Fig. 1). Furthermore, there were several regions within the available climate space where ferns and lycophytes have not been recorded from. For example, regions that experience high temperature and low levels of precipitation have comparatively fewer fern records. Figure 2 shows a geographical projection of the PC scores, clearly indicating that the Departments of Nacaome, Choluteca, and the Rio Ulua in Cortes are comparatively arid. Similarly, the Sierra Madre mountains are defined by lower precipitation and high temperatures, whereas the La Mosquitia region in the northeast are defined by high precipitation and high temperatures (Figs. 1 and 2). In quadrants with negative PC1 scores, species tended to be less frequent, as well as less dense closer to the edges of the available niche space. For example, variance was over four times higher across the available niche space in cooler conditions than in warmer conditions (F = 45.31, p = 3.60e-11). It is important to note that this difference was not a by-product of the variances of the available climatic space when they compared in a similar manner (7.1 vs. 7.2, respectively). We found similar differences when comparing regions with high diurnal temperature and low annual precipitation (i.e., positive PC2 scores), with areas of low diurnal temperature and high annual precipitation (i.e., positive PC1) (F = 53.55, p = 7.18e-13). However, in the second comparison, the difference could not simply be explained by differences in the available climatic niche space (8.9 vs. 23.3, respectively).

Spatial distribution of principal component scores of the climatic conditions across Honduras. The climatic conditions associated with each grid cell were subject to a principal component analysis and the resulting scores were mapped back to each corresponding cell for visualization. PC1 mostly reflected variation in temperature, especially lower temperatures (A), whereas PC2 loadings described a trade-off between maximum temperature and diurnal temperature range versus precipitation (particularly during autumn and winter) (B). See text for details
Interestingly, there were no obvious differences in the pattern of occupation among the most species-rich families (Fig. 1A). When comparing the entire available climatic space, there were significant differences between life forms (i.e., epiphytes vs. terrestrial ferns) in their pattern of occupation (PC1, F = 7.35, p = 0.0069; PC2, F = 9.19, p = 0.0025; Table 2, Fig. 1B), with epiphytes showing smaller variance in their climatic niches than terrestrial species (13.38 vs. 21.48 and 8.52 vs. 15.22, for PC1 and PC2, respectively).
Climate projections for the years 2020 to 2099 for RCP2.6 and RCP8.5 suggest how climate space in Honduras will shift in the future (Fig. 3). In general, the climate space will shift to hotter and drier conditions, with RCP8.5 projections showing the strongest shift (Fig. 4). Between 18.9 and 57.8% the analyzed species will have their current average climatic niche outside future climatic spaces (Table 3). This result is not due to a reduction in available climatic space, but rather an overall mismatch between present and future climatic spaces (Table 3 and Fig. 4). Interestingly, the proportion of impacted species is larger for epiphytic than for terrestrial species, except for the most severe shifts (i.e., RCP8.5 for the year 2069 and RCP8.5 for the year 2099), in which this tendency is reversed (Table 3). It is important to note that there are 23 species that are indicated as outside the present climatic space. Those were species living near the boundaries of the present climatic space but were categorized to be outside because of the way we computed the concave hull. They were left to ensure that all comparisons were carried out using the same framework.

Principal component analysis of climatic conditions found in Honduras, including climate space projections under RCP2.6 (A) and RCP8.5 (B) for different years. Panel (C) shows the outline of all the projections. Red empty circles represent the species means

Predicted consequences of climate space shifts according to different climate change scenarios (A–F). The white area represents the current climate space, yellow the future-predicted climate space, and orange shows the overlap between current and predicted (see Table 3 for details)
Discussion
Although fern and lycophyte species were widely distributed throughout the available climatic space in Honduras, we found evidence that particular climate conditions were occupied more frequently. In relatively colder conditions (positive PC1 scores), species tended to be distributed throughout the entire available climatic space, whereas in warmer conditions (negative PC1 scores), their distribution became more limited, with precipitation and diurnal temperature being the limiting factors. In addition, terrestrial species showed more variation in their occupied climatic niches compared to epiphytes. This is not surprising, as epiphytes are believed to be more sensitive to variable and extreme climate conditions during different stages of their life (Benzing 1998; Klinghardt and Zotz 2021). To the best of our knowledge, our study is the first to explicitly demonstrate this pattern for ferns and lycophytes; however, the mechanisms underlying the difference between epiphytes and terrestrial species are not easily determined. We suggest two non-exclusive mechanistic explanations. Firstly, the rate of climatic niche evolution between these two life forms could differ (Hernandez-Rojas et al. 2021). For instance, it has been argued that life history might affect rates of climatic niche evolution in plants, with woody lineages accumulating fewer changes per million years in climatic niche space than related herbaceous lineages (Smith and Beaulieu 2009). A more rigorous test of this hypothesis would be required for our ferns (O'Meara et al. 2006), but more precise phylogenetic and trait data are currently lacking for many of our study species (Kessler et al. 2016). Alternatively, epiphytes are likely to be less well buffered from changes in climate conditions compared to terrestrial species due to their dependence on the forest canopy, often living in less shaded conditions than their conspecific terrestrial taxa and thus being less well buffered higher in the canopy (de Frenne et al. 2019). Assessing the relative importance of these mechanisms might provide valuable insight to understanding how life history traits drive the occupation of the fern climatic niche space in Honduras. It is also worth noting that the higher variance in epiphyte niche occupation could also be partially explained by the lack of host-specific habitat in areas that have high levels of seasonality and low rainfall, such as coastal regions.
There are some important caveats of the approach used in the present study. Firstly, for practical reasons of working with over 700 species, we did not explicitly analyze intraspecific variation in climatic niches. However, it is becoming increasingly clear, particularly in plant species that intraspecific variation in functional traits might lead to marked consequences for community-level dynamics (e.g., Lajoie and Vellend 2015; Niu et al. 2020). Secondly, microhabitats might be more important than general climatic conditions to determine the individual distribution of fern and lycophyte species (Jones et al. 2011). Finally, the data used in our study were based on Batke et al. (2022) database of fern and lycophyte records from Honduras. This dataset was based on herbarium records from the years 1850 to 2018, which can be limiting due to known recorder biases (Daru et al. 2017). However, databases using collated herbarium information are often the only and most comprehensive resource to assess the effect of climate change on entire species groups (Loiselle et al. 2007).
Climate change is expected to have a profound influence on fern and lycophyte communities (Gasper et al. 2021), especially epiphytes (Hsu et al. 2011; Hsu et al. 2014; Reyes et al. 2021b). Our study showed that some regions of the available climatic space will be shifting in Honduras under RCP2.6 and RCP8.5. Particularly species that occurred close to the vicinity of the current climate space or species that have a more restricted climate niche are more likely to experience shifts or losses in the future. For example, areas of moderate precipitation and low temperatures are likely to be decreasing (Fig. 3). Range shifts have frequently been reported for plant and animal species (Magrin et al. 2014) and are believed to be particularly problematic for species that already live close to their climate limit or have a narrow climate range. A recent study by Reyes et al. (2021b) showed that 63 ferns in Celaque National Park, the tallest mountain in Honduras, are likely to shift their range fully or partially above the maximum altitude of the mountain under a RCP2.6 scenario for the year 2050. They also showed that 65% of these species were epiphytes. This is especially concerning, as epiphytic ferns and lycophytes often show exceptional species richness in mid and higher altitudes (Suissa et al. 2021). Celaque is known as one of the most species-rich areas for ferns in Honduras (Rojas-Alvarado 2012) and is believed to have one of the highest levels of endemism in the country. Our climate space analysis showed similar trends as discussed in Reyes et al. (2021b), in that species in high-elevation areas, such as Celaque, are likely to lose their currently occupied climate space (Fig. 3). This is because these high-elevation areas are characterized by low temperature and high precipitation, which is a rare climate niche in Honduras and will be lost as a result of rising temperatures and a decrease in precipitation (Fig. 2).
We found that terrestrial species are at higher risk in the future, as their climate niche is in closer proximity to their current climate space. Our forecast predictions showed that proportionally more terrestrial species will fall outside the available climate space in the future (Table 3). For example, under a worse-case scenario (RCP8.5) for the year 2099, we estimate that 391 species might shift their average climate niche outside the available climate space. Of the 391 species 17.1% are epiphytes and 82.9% are terrestrial species. For the same projection, we also found that the overlap of climate space between the current and future projection is as little as 47.1%. This is a worrying shift and could result in a large loss of species, in cases where species cannot adopt or have a narrow climate niche (e.g., highly specialized species).
It needs to be noted that we calculated the average niche positions of species in the predicted available climatic space for Honduras. This means that although the average niche positions might fall outside the predicted available climatic space for some taxa, it does not necessarily imply that these species will completely disappear. This is because the shift in climate space might still be suitable for survival but might result in those species being closer to their niche limit. Having a narrower available climate space would make these species more susceptible to sudden perturbations in their environment (e.g., higher predicted extreme events, such as drought, Magrin et al. 2014). Species that are more dynamically capable of physiological response to sudden changes in their environment (e.g., by closing their stomata) and/or are morphologically capable to adapt (e.g., by changing their stomatal density) are less likely to be affected. For example, it has been shown that the predicted increase in atmospheric CO2 conditions under RCP8.5 for the year 2050 (approximately to 560 ppm) is likely to increase plant water use efficiency as plants close their stomata more (Purcell et al. 2018). Experimental work has demonstrated that this is particularly pronounced in ferns that receive more light (Batke et al. 2018), as would be the case of many epiphytic ferns and lycophytes when compared to lower understory terrestrial species. It is therefore possible that the combined effect of elevated CO2, increases in temperature (to a certain point, see Purcell et al. 2018) and decreases in precipitation, might only negatively affect species that are less physiologically and morphologically adaptable. This is likely to include ferns and lycophytes that require higher availability of water to stay physiologically active to maintain their water to carbon balance, which would include water loving ferns, such as species in the Hymenophyllaceae (i.e., filmy ferns). Currently, there are little detailed data available on the morphological and physiological responses of ferns to changes in climate, making a systematic species comparison difficult (Cheng et al. 2010; Franks et al. 2016).
In summary, our study found that the climate space across Honduras is changing and that some fern and lycophyte species are going to be at higher risk compared to others. Specifically, climate space availability will likely change in areas that experience lower precipitation and temperatures, resulting in unsuitable average niche conditions for up to 57.8% of Honduran fern and lycophyte species. In general, Honduras is expected to become drier in the near future, with more extreme events of high drought (Magrin et al. 2014). It is important to note that, given that we focused on species means, we have not explored intraspecific variation in climatic niche occupation. However, the species near the boundaries of the Honduran climatic niche space are particularly relevant for further studied using ecological niche modeling (Syfert et al. 2018). Nevertheless, the implications of our findings are particularly relevant to conservation management, as our data highlight areas at higher risk for fern and lycophyte species loss and therefore can be used to directly inform policy planning. Ferns and lycophytes constitute the most systematically organized group of Honduran plants (Reyes et al. 2021a) and are ideal for studying Honduran climatic niche variation. Considering that ferns and lycophytes are particularly sensitive to changes in climate, if tested in the future, they could be used as a proxy for other Honduran plant taxa. To test this, more taxonomic work would be required to update the current list of other vascular plant species and to assemble a similar inventory as done by Reyes et al. (2021a).
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Aros-Mualin D, Noben S, Karger DN, Carvajal-Hernández CI, Salazar L, Hernández-Rojas A, Kluge J, Sundue MA, Lehnert M, Quandt D, Kessler M (2021) Functional diversity in ferns is driven by species richness rather than by environmental constraints. Front Plant Sci 11:615723. https://doi.org/10.3389/fpls.2020.615723
Atwater DZ, Ervine C, Barney JN (2018) Climatic niche shifts are common in introduced plants. Nat Ecol Evol 2(1):34–43. https://doi.org/10.1038/s41559-017-0396-z
Batke S, Cascante-Marin A, Kelly D (2016) Epiphytes in Honduras: a geographical analysis of the vascular epiphyte flora and its floristic affinities to other Central American countries. Trop Ecol 57(4):663–675
Batke S, Holohan A, Hayden R, Fricke W, Porter AS, Evans-Fitz Gerald CM (2018) The pressure is on–epiphyte water-relations altered under elevated CO2. Front Plant Sci 9:1758. https://doi.org/10.3389/fpls.2018.01758
Batke S, Dallimore D, Reyes-Chavez J, Maradiaga RFD, Jones I, Atkinson W, Acosta FL, Reid G (2022) The largest digital database of fern and lycopod records from Honduras – spatial, temporal and collector biases. Bot J Linnean Soc. https://doi.org/10.1093/botlinnean/boac019
Barrington DS (1993) Ecological and historical factors in fern biogeography. J Biogeogr 20:275–279. https://doi.org/10.2307/2845635
Benzing DH (1998) Vulnerabilities of tropical forests to climate change: the significance of resident epiphytes. In: Markham A (ed) Potential Impacts of Climate Change on Tropical Forest Ecosystems. Springer, Dordrecht, pp 379–400
Brown JL, Hill DJ, Dolan AM, Carnaval AC, Haywood AM (2018) PaleoClim, high spatial resolution paleoclimate surfaces for global land areas. Sci Data 5(1):180254. https://doi.org/10.1038/sdata.2018.254
Buckley LB, Davies TJ, Ackerly DD et al (2010) Phylogeny, niche conservatism and the latitudinal diversity gradient in mammals. Proc R Soc b 277(1691):2131–2138. https://doi.org/10.1098/rspb.2010.0179
Bystriakova N, Schneider H, Coomes D (2011) Evolution of the climatic niche in scaly tree ferns (Cyatheaceae, Polypodiopsida). Bot J Linn Soc 165(1):1–19. https://doi.org/10.1111/j.1095-8339.2010.01092.x
Cheng W, Sakai H, Matsushima M, Yagi K, Hasegawa T (2010) Response of the floating aquatic fern Azolla filiculoides to elevated CO2, temperature, and phosphorus levels. Hydrobiologia 656:5–14. https://doi.org/10.1007/s10750-010-0441-2
Daru BH, Park DS, Primack RB et al (2018) Widespread sampling biases in herbaria revealed from large-scale digitization. New Phytol 217(2):939–955. https://doi.org/10.1111/nph.14855
De Frenne P, Zellweger F, Rodríguez-Sánchez F et al (2019) Global buffering of temperatures under forest canopies. Nat Ecol Evol 3:744–749. https://doi.org/10.1038/s41559-019-0842-1
Duran A, Pie MR (2015) Tempo and mode of climatic niche evolution in Primates. Evolution 69(9):2496–2506. https://doi.org/10.1111/evo.12730
Fox J, Weisberg S (2019) An R companion to applied regression, 3rd edn. Sage, Thousand Oaks
Franks PJ, Britton-Harper ZJ (2016) No evidence of general CO2 insensitivity in ferns: one stomatal control mechanism for all land plants? New Phytol 211:819–827. https://doi.org/10.1111/nph.14020
Gasper AL, Grittz GS, Russi CH, Schwartz CE, Rodrigues AV (2021) Expected impacts of climate change on tree ferns distribution and diversity patterns in subtropical Atlantic Forest. Perspect Ecol Conserv 19:369–378. https://doi.org/10.1016/j.pecon.2021.03.007
Gombin J, Vaidyanathan R, Agafonkin V (2020) Concaveman: A very fast 2D concave hull algorithm. R package version 1.1.0. https://CRAN.R-project.org/package=concaveman
Hernández-Rojas AC, Kluge J, Krömer T et al (2020) Latitudinal patterns of species richness and range size of ferns along elevational gradients at the transition from tropics to subtropics. J Biogeogr 47(6):1383–1397. https://doi.org/10.1111/jbi.13841
Hernandez-Rojas AC, Kluge J, Noben S, Reyes CJD, Krömer T, Carvajal-Hernández CI, Salazar L, Kessler M (2021) Phylogenetic diversity of ferns reveals different patterns of niche conservatism and habitat filtering between epiphytic and terrestrial assemblages. Front Biogeogr 13(3):e50023. https://doi.org/10.21425/F5FBG50023
Herrera JM, Ploquin EF, Rasmont P, Obeso JR (2018) Climatic niche breadth determines the response of bumblebees (Bombus spp.) to climate warming in mountain areas of the Northern Iberian Peninsula. J Insect Conserv 22(5):771–779. https://doi.org/10.1007/s10841-018-0100-x
Hijmans RJ (2021) Raster: geographic data analysis and modeling. R package version 3.4–13. https://CRAN.R-project.org/package=raster
Hsu RCC, Oostermeijer JGB, Wolf JHD (2014) Adaptation of a widespread epiphytic fern to simulated climate change conditions. Plant Ecol 215:889–897. https://doi.org/10.1007/s11258-014-0340-0
Hsu RCC, Tamis WLM, Raes N et al (2012) Simulating climate change impacts on forests and associated vascular epiphytes in a subtropical island of East Asia. Divers Distrib 18:334–347. https://doi.org/10.1111/j.1472-4642.2011.00819.x
Hutchinson GE (1957) Concluding remarks. Cold Spring Harbour Symp Quant Biol 22:415–427
Jezkova T (2016) Rates of change in climatic niches in plant and animal populations are much slower than projected climate change. Proc Royal Soc B: Biol Sci 283:1-9. https://doi.org/10.1098/rspb.2016.2104.
Jones MM, Szyska B, Kessler M (2011) Microhabitat partitioning promotes plant diversity in a tropical montane forest. Glob Ecol Biogeogr 20(4):558–569. https://doi.org/10.1111/j.1466-8238.2010.00627.x
Kamilar JM, Muldoon KM (2010) The climatic niche diversity of Malagasy primates: a phylogenetic perspective. PLoS ONE 5(6):e11073. https://doi.org/10.1371/journal.pone.0011073
Karger DN, Conrad O, Böhner J et al (2017) Climatologies at high resolution for the earth’s land surface areas. Sci Data 4(1). https://doi.org/10.1038/sdata.2017.122
Kerkhoff AJ, Moriarty PE, Weiser MD (2014) The latitudinal species richness gradient in New World woody angiosperms is consistent with the tropical conservatism hypothesis. Proc Natl Acad Sci 111(22):8125–8130. https://doi.org/10.1073/pnas.1308932111
Kessler M, Karger DN, Kluge J (2016) Elevational diversity patterns as an example for evolutionary and ecological dynamics in ferns and lycophytes. J Syst Evol 54(6):617–625. https://doi.org/10.1111/jse.12218
Khaliq I, Fritz SA, Prinzinger R, Pfenninger M, Böhning-Gaese K, Hof C (2015) Global variation in thermal physiology of birds and mammals: evidence for phylogenetic niche conservatism only in the tropics. J Biogeogr 42(11):2187–2196. https://doi.org/10.1111/jbi.12573
Klinghardt M, Zotz G (2021) Abundance and seasonal growth of epiphytic ferns at three sites along a rainfall gradient in Western Europe. Flora 274:151749. https://doi.org/10.1016/j.flora.2020.151749
Krömer T, Acebey A, Kluge J, Kessler M (2013) Effects of altitude and climate in determining elevational plant species richness patterns: a case study from Los Tuxtlas, Mexico. Flora-Morphol Distrib Funct Ecol Plants 208(3):197–210. https://doi.org/10.1016/j.flora.2013.03.003
Lajoie G, Vellend M (2015) Understanding context dependence in the contribution of intraspecific variation to community trait–environment matching. Ecology 96(11):2912–2922. https://doi.org/10.1890/15-0156.1
Linares-Palomino R, Cardona V, Hennig EI et al (2008) Non-woody life-form contribution to vascular plant species richness in a tropical American forest. In: Van der Valk AG (ed) Forest ecology. Springer, Dordrecht, pp 87–99
Linares-Palomino R, Cardona V, Hennig E, Hensen I, Hoffmann D, Lendzion J, Soto D, Herzog S, Kessler M (2009) Non-woody life-form contribution to vascular plant species richness in a tropical american forest. Plant Ecol 201:87–99
Loiselle BA, Jørgensen PM, Consiglio T, Jiménez I, Blake JG, Lohmann LG, Montiel OM (2007) Predicting species distributions from herbarium collections: does climate bias in collection sampling influence model outcomes? J Biogeogr 35:105–116. https://doi.org/10.1111/j.1365-2699.2007.01779.x
Magrin GO, Marengo JA, Boulanger J-P et al (2014) Central and South America. In: Barros, VR, Field CB, Dokken DJ, et al. eds. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press: Cambridge. pp 1499–1566
Meyer ALS, Pie MR (2018) Environmental prevalence and the distribution of species richness across climatic niche space. J Biogeogr 45:2348–2360. https://doi.org/10.1111/jbi.13419
Meyer ASL, Pie MR (2022) Climate change estimates surpass rates of climatic niche evolution in Primates. Int J Primatol 43:40–56
Moran RC, Riba R (1995) Psilotaceae a Salviniaceae. In. Davidse G, Sousa MS, Knapp S eds. Flora Mesoamericana.Vol. 1. Universidad Nacional Autónoma de México: México, 470
Racines NCE, Herrera LL, Monserrate F (2018a) 30-seconds (1 km2) monthly, seasonal and annual gridded historical climate surfaces for Honduras. Harvard Dataverse V1: downloaded on 18 October 2021
Racines NCE, Herrera LL, Monserrate F (2018b) 30-seconds (1 km2) gridded future climate change scenarios for Honduras. Harvard Dataverse V1: downloaded on 21 October 2021
Syfert MM, Brummitt NA, Coomes DA et al (2018) Inferring diversity patterns along an elevation gradient from stacked SDMs: a case study on Mesoamerican ferns. Glob Ecol Conserv 16:e00433. https://doi.org/10.1016/j.gecco.2018.e00433
Nitta JH, JrJE W, Davis CC (2020) Life in the canopy: community trait assessments reveal substantial functional diversity among fern epiphytes. New Phytol 227(6):1885–1899. https://doi.org/10.1111/nph.16607
Niu K, Zhang S, Lechowicz MJ (2020) Harsh environmental regimes increase the functional significance of intraspecific variation in plant communities. Funct Ecol 34(8):1666–1677. https://doi.org/10.1111/1365-2435.13582
Oksanen J, Blanchet G, Friendly M et al (2020) Vegan: Community Ecology Package. R package version 2.5–7. https://CRAN.R-project.org/package=vegan
O’Meara BC, Ané C, Sanderson MJ, Wainwright PC (2006) Testing for different rates of continuous trait evolution using likelihood. Evolution 60(5):922–933. https://doi.org/10.1111/j.0014-3820.2006.tb01171.x
Petitpierre B, Kueffer C, Broennimann O, Randin C, Daehler C, Guisan A (2012) Climatic niche shifts are rare among terrestrial plant invaders. Science 335(6074):1344–1348. https://doi.org/10.1126/science.1215933
Pie MR, Campos LL, Meyer AL (2017) The evolution of climatic niches in squamate reptiles. Proc R Soc B 284(1858):20170268. https://doi.org/10.1098/rspb.2017.0268
Pie MR, Divieso R, Caron FS (2021) The evolution of climatic niche breadth in terrestrial vertebrates. J Zool Syst Evol Res 59(6):1155–1166. https://doi.org/10.1111/jzs.12508
PPGI (2016) A community derived classification for extant lycophytes and ferns. J Syst Evol 54(6):563–603. https://doi.org/10.1111/jse.12229
Purcell C, Batke SP, Yiotis C et al (2018) Increasing stomatal conductance in response to rising atmospheric CO2. Ann Bot 121(6):1137–1149. https://doi.org/10.1093/aob/mcx208
Quintero I, Wiens JJ (2013) Rates of projected climate change dramatically exceed past rates of climatic niche evolution among vertebrate species. Ecol Lett 16(8):1095–1103. https://doi.org/10.1111/ele.12144
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing: Vienna, Austria. https://www.R-project.org/
Ramírez-Barahona S, Barrera-Redondo J, Eguiarte LE (2016) Rates of ecological divergence and body size evolution are correlated with species diversification in scaly tree ferns. Proc R Soc b 283:20161098. https://doi.org/10.1098/rspb.2016.1098
Reyes-Chávez J, Tarvin S, Batke SP (2021a) Ferns and Lycophytes of Honduras: a new annotated checklist. Phytotaxa 506(1):1–113. https://doi.org/10.11646/phytotaxa.506.1.1
Reyes-Chávez J, Quail M, Tarvin S, Kessler M, Batke SP (2021b) Nowhere to escape–diversity and community composition of ferns and lycophytes on the highest mountain in Honduras. J Trop Ecol 37(2):72–81. https://doi.org/10.1017/S0266467421000122
Rojas-Alvarado A (2012) New records of club mosses (Lycopodiophyta and ferns (Pteridophyta) from Honduras and Parque Nacional Montanas de Celaque. Rev Biodivers Neotrop 2(2):83–92
Smith SA, Beaulieu JM (2009) Life history influences rates of climatic niche evolution in flowering plants. Proc R Soc b 276(1677):4345–4352. https://doi.org/10.1098/rspb.2009.1176
Suissa JS, Sundue MA, Testo WL (2021) Mountains, climate and niche heterogeneity explain global patterns of fern diversity. J Biogeogr 48:1296–1308. https://doi.org/10.1111/jbi.14076
Sugiyama M, Shiogama H, Emori S (2010) Precipitation extreme changes exceeding moisture content increases in MIROC and IPCC climate models. Proc Natl Acad Sci 107(2):571–575. https://doi.org/10.1073/pnas.0903186107
Tryon R (1970) Development and evolution of fern floras of oceanic islands. Biotropica 2:76–84. https://doi.org/10.2307/2989765
Ulloa CU, Acevedo-Rodríguez P et al (2017) An integrated assessment of the vascular plant species of the Americas. Science 358(6370):1614–1617. https://doi.org/10.1126/science.aao0398
Wasof S, Lenoir J, Aarrestad PA et al (2015) Disjunct populations of European vascular plant species keep the same climatic niches. Glob Ecol Biogeogr 24(12):1401–1412. https://doi.org/10.1111/geb.12375
Wiens JJ, Donoghue MJ (2004) Historical biogeography, ecology and species richness. Trends Ecol Evol 19(12):639–644. https://doi.org/10.1016/j.tree.2004.09.011
Williams CB, Murray JG, Glunk A, Dawson TE, Nadkarni NM, Gotsch SG (2020) Vascular epiphytes show low physiological resistance and high recovery capacity to episodic, short-term drought in Monteverde, Costa Rica. Funct Ecol 34(8):1537–1550. https://doi.org/10.1111/1365-2435.13613
Wild M, Gagnon D (2005) Does lack of available suitable habitat explain the patchy distributions of rare calcicole fern species? Ecography 28(2):191–196. https://doi.org/10.1111/j.0906-7590.2005.04113.x
Wolf PG, Schneider H, Ranker TA (2001) Geographic distributions of homosporous ferns: does dispersal obscure evidence of vicariance? J Biogeogr 28(2):263–270
Zotz G (2013) The systematic distribution of vascular epiphytes–a critical update. Bot J Linn Soc 171(3):453–481. https://doi.org/10.1111/boj.12010
Zotz G, Weigelt P, Kessler M, Kreft H, Taylor A (2021) EpiList 1.0: a global checklist of vascular epiphytes. Ecology 102:e03326. https://doi.org/10.1002/ecy.3326
Acknowledgements
This work was supported by Edge Hill University Research Investment Fund and Chevening Scholarships, the UK government global scholarship program, funded by the Foreign, Commonwealth and Development Office (FCDO), and partner organizations.
Funding
This work was supported by Edge Hill University Research Investment Fund and Chevening Scholarships, the UK government global scholarship program, funded by the Foreign, Commonwealth and Development Office (FCDO), and partner organizations.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Data were gathered by SB, TD, and JR. The analysis was carried out by MP. The manuscript was written by all authors. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by Dafeng Hui.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visithttp://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pie, M.R., Batke, S.P., Reyes-Chávez, J. et al. Fern and lycophyte niche displacement under predicted climate change in Honduras. Plant Ecol 223, 613–625 (2022). https://doi.org/10.1007/s11258-022-01235-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-022-01235-8