
Abstract
Urbanization has transformed more than 50% of the land area and brought the emergence of a new humanized-based ecosystem. In answer to this, urban systems have looked for become resilient. Thus, urban landscapes must enhance their resilience capacities; robustness, adaptability, and transformability. However, there is a necessity of a well performing and easy gathering indicators. In this paper, we propose that bats are a good indicator of city’s capacity to be resilient. We used bat permanence, species diversity, and activities to analyze Mexico City resilience capacities. We used citizen science to register bat sounds around the city. From these data we identified bat species and two main activities: passes and hunting buzzes. Data showed that the east and center area of the city had the lower species richness and relative abundance, while south was the richest. This diversity patterns were in response to different habitat characteristics, mainly those related to landscape. However, activities and permanence were strongly related with human presence. The bat permanence modeled, indicated that bat community structure could be shaped under changing scenarios. Due to their differential sensitivity bats are good indicators of habitat changes in urban landscapes. Cities seeking resilience should perform planned-habitat management to achieve a bat-friendly status and promote the ecosystem services that bats can provide.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In the last two decades, urbanization has transformed more than 50% of the planet’s land area (Buettner 2015). The environmental and structural changes done on once natural ecosystems, has brought a novel and functionally complex human-constructed urban system (Pataki 2015). This process posits environmental and social changes that are unprecedented in scale, scope, and complexity. In this context, cities are facing multiple challenges in order to move from a vulnerable to a more resilient state (Meerow et al. 2016). Resilience is defined as an ecosystem’s ability to maintain basic functional characteristics in the face of disturbance (Holling 1973). More recently, it has been proposed that ecosystems have multiple stable states and in a constant state of flux, so it can be possible to distinguish between static resilience, referring to a system’s ability to bounce back to its previous state, and dynamic resilience, which focuses on maintaining key functions when perturbed (Holling 1996). Ecologically, Hollings’ definitions allow integrating several resilience issues into the complex network of an urban system. Although theoretical bases are well established, the current necessities are focused in the finding of good indicators of resilience capacities.
Resilience theory emphasizes change, uncertainty, and the capacity of systems to adapt, but the framework to analyze resilience is not always clear (Gunderson 2001). Therefore, a first step could be to establish the resilience capacities that are not only specific strategies for resilience preparedness but are a part of the ecological, social and economic structure of an urban landscape (Norris et al. 2008). In an urban context, capacities could be represented by all those modified variables that lead cities to resilience. Good indicators most have the ability to reliable represent robustness, adaptability and transformability of a landscape (Folke et al. 2002, 2010; Meuwissen et al. 2019; Walker et al. 2004).
Bats are not only taxonomically, but also extremely diverse in ecological terms, occupying many trophic levels (Kunz and Fenton 2005). They provide essential ecosystem services controlling insect populations, dispersing seeds and pollinating several plants (Kunz et al. 2011). Thus, bats are used as indicator for studies on the impact of landscape changes. Habitat loss and fragmentation driven by urbanization frequently result in negative changes in bats (Ramos Pereira et al. 2018; Russo and Ancillotto 2015). For example, although, flight may allow bats to travel through urban landscapes and exploit habitat patches that are otherwise isolated, the effects of urbanization results in declines in diversity and abundance of bats (Russo and Ancillotto 2015). However, responses to urbanization differ among species (Furlonger et al. 1987), and thus the relationship between urbanization and bats is pretty complex. Some species that have sympatric distributions differ in their selection for daytime roost sites or preferred foraging habitat (Aldridge and Rautenbach 1987; Furlonger et al. 1987; Gaisler et al. 1998). For example, Eptesicus fuscus and Myotis lucifugus frequently use buildings or other anthropogenic structures as roosts in contrast to solitary species of Lasiurus, which prefer trees for roosts. Such patterns of habitat use may result in species-specific responses to urban development affecting their permanence in the community (Anderson et al. 1992).
With urbanization, one of the first process affected is bat foraging, even if some species may successfully exploit urban roosting sites (Gehrt and Chelsvig 2003; Neubaum et al. 2007), they could not find suitable foraging areas near their roosts and thus must travel large distances to reach food (Avila-Flores and Fenton 2005; Rydell 2006). Indeed, urbanization leads population declines of insect groups of importance to bat foraging such as moths (Jung and Kalko 2011; Russo and Ancillotto 2015). However, there are reports that street lights may attract insects and improve the insect capture by bats adapted to hunt in artificially illuminated sites as well (Tomassini et al. 2014).
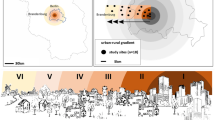
Under this context, it can be expected that urban bats community have differential responses to factors that give heterogeneity to cities such as landscape configuration, roost availability, proximity to adequate foraging sites and water sources, climate and anthropogenic disturbance (Barbar et al. 2015; Mendes et al. 2017; Tews et al. 2004). Thus, we propose use bats as indicators of robustness, adaptability and transformability as the basic resilience capacities of a highly urbanized landscape (Fig. 1). In this work, we tested the hypothesis that urbanization could have a general negative effect on bat species diversity, activities, and permanence. We addressed two objectives. First, we analyzed how bat diversity and their activities vary across the urban landscape. Secondly, we modeled the permanence of bats as indicators of urban resilience capacities.

Bat-based conceptual framework for evaluation of resilience capacities of an urban landscape. A conserved bat community can inhabit cities, when it has the capacities to become more resilient, the confluence of both characteristics will give a resilient designed urban landscape. (a) Bat species richness allows detecting the robustness of an urban landscape when bat species persist, but with changes in composition and abundances due to its capacity to withstand stress. (b) Bat community structure allows to detect adaptability capacities of an urban landscape because non-structural changes favor a richer bat community structuration as city’s adaptability grows. (c) Bat community structure and bat permanence allows to detect transformability capacities of an urban landscape because structural changes would favor, on one hand an increment in bat diversity and, on the other, changes in community composition as the city transforms its landscape to get better condition for bats. During transformation, bat community would decrease due to bat capacities to confront changes, but once new landscape elements were incorporated bat community will restructure. Figure based on Meuwissen et al. (2019)
Methods
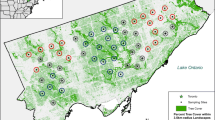
Bat sampling was carried out in Mexico City (CDMX), one of the world’s largest megacities, with an estimated 20 million inhabitants in about 150,000 ha. We regionalized the CDMX into Center, South, North, East and West areas. Although this regionalization was made on geographic bases, it also includes some urbanization characteristics that allow to have different degrees of urbanization (Fig. 2). The city is at 2,240 masl and a temperate weather (17–25 ºC) with extreme variations in summer and winter. Its landscape is composed by a matrix of habitats that cover from protected natural areas to highly industrialized ones. Natural vegetation varies from temperate forests in the southern areas to dry shrubs at north.

Study area showing the bat sampling points and the five regions in which the whole Mexico City was divided
Bat sampling
Bats sampling was performed with Echo Meter Touch 2 (Wildlife Acoustics Inc.), a bat detector that registered ultrasonic vocalizations and recorded in an attached device, this allows the further analysis of calls in order to identify the bat species and their activity. This approach gives information about bats flying, feeding or roosting behavior. Because gathering data could be time consuming, we implemented citizen science tools, which allows the use of extensive bat sampling protocols (Newson et al. 2015).
For data gathering we first contacted people interested in participating in the bat project throughout CDMX. This first approach was through the implementation of an interview that looked for a compilation of knowledge and experiences of city inhabitants about bats (GGG, unpublished data). Once we detected the people willing to participate (about 50 volunteers) in the project; we gave them a basic training about the use of the bat detector, sampling protocol and data gathering. After training, a test of acquired skills was performed in order to avoid bias and improve data quality (Kosmala et al. 2016). This basic training included (1) capacitated volunteers taking recordings and showing images of bat species detected (2) Standardized recordings time and walk velocity, and (3) Although volunteers had the option to choose the better route for the transects, we checked there were no biases when trying to avoid some urban elements such as large factory areas.
Bat data was collected at sunset (18:00 h.) and finished after midnight (about 01:00 h). Sampling was conducted for five consecutive nights out in the summer (June-September 2019), when the availability of insects is higher, and absence of bats was not biased by this factor. Bat sampling followed one of the four cardinal points and consisted in a point-line sampling. Each volunteer walked in a straight line for three kilometers at a constant speed, and each 100 m stopped and register bat activity for three minutes and continued walking to next point. In total were performed between 5 and 8 transects per sampling areas.
Recordings were first registered using the automated bat identification, and were further confirmed with Kaleidoscope software, mainly in the case of Myotis species. Once each recording was assigned to a species, we classified them for type of bat activities. To make this, we reviewed the full spectrum of each bat call to characterized wavelength and amplitude. With these metrics, we determined two main activities (1) Hunting (hereafter named hunting buzz), when a call become shorter and broader in bandwidth indicating that bat localized and approached to a prey and (2) when wave still similar in frequency and length, meaning bat is “passing” from one site to another (Stahlschmidt and Brühl, 2012).
Urban variables
Landscape variables
We used several landscape variables to describe the relationship between bat species and activities with the urbanization in the CDMX. First we performed the analysis of a Sentinel-2 satellite image (S2A_MSIL1C_20190107T170701_T14QMG) downloaded from the European Spatial Agency (accessed 10/02/2019, https://scihub.copernicus.eu/dhus/#/home). Image was preprocessed for atmospheric and geometric correction and further classified into (1) Closed natural vegetation, (2) Open natural vegetation, (3) Urban areas, (4) Farm- and livestock lands (5) Water and (6) Mixed areas which include all those areas with farmlands and secondary vegetation (altogether constitute bat habitat). We used data from the Visible Infrared Imaging Radiometer Suite (VIIRS) Day/Night Band for a light intensity characterization. Image was downloaded from the National Centers for Environmental Information at the National Oceanic and Atmospheric Administration (NOAA; last accesed 01/02/2019 https://ngdc.noaa.gov/eog/viirs/download_dnb_composites.html). Additionally, we used vector data of Natural Protected Areas (NPA) and Water Bodies (WB) (last accessed 14/05/2019, http://www.atlas.cdmx.gob.mx/datosabiertos.php) generated by the CDMX government (see maps in online resource 1).
We extracted the values for each sampling from all digital information sources in order to get specific habitat information. We classified green areas in small (SGA), mainly integrated by urban parks, cemeteries and arboreal street vegetation (< 100 ha); and large green areas (LGA) composed by urban parks ≥ 100 ha. To estimate the proximity of these areas to each sampling area, we measured the distance between them, if they were into a buffer of 2 km, it was classified as near, and if was out of this buffer as far. This distance is the average flying of an insectivorous bat per night (Bergeson et al. 2020; Rainho and Palmeirim 2011),
Additionally, we analyzed the effect of habitat complexity on bats, for this, we constructed an index that considered all variables described above. The most complex habitat was determined when this index was integrated for more than four landscape components. An habitat with medium complexity had 2–3 elements, while those areas with only one element were considered as no complex one.
Environmental variables
We used as environmental variables: temperature and precipitation, both in general influence the distribution, presence and activities of bats. Mean annual data of temperature and precipitation were obtained from worldclim (ver. 2.1) bioclimatic variables (bio 1 and bio 12) with a spatial resolution of 30 s (about 1 km2), an assumed that this represent the 3 km around as rain has low variability at those scales (last accessed 03/02/2020; https://worldclim.org/data/worldclim21.html). We used this historical registers because it comprise the long-term rain variability. In addition to these variables, we used data the air quality index (AQI) that include O3, NO2, SO2, CO, and suspended particles, gained from the automatic atmospheric network of the CDMX that have monitoring stations in barely all alcaldías of the city (last accessed 03/02/2020; http://www.aire.cdmx.gob.mx).
Human variables
Finally, in order to integrate one variable that describes human activity we used population density. Although this variable is static, it allowed us to know the number of inhabitants in the areas where sampling were done. These data were obtained from the National Institute of Statistics, Geography and Informatics (last accessed 03/02/2020; https://www.inegi.org.mx/app/mapa/espacioydatos/). These data are part of the National Population Census, thus there are data for each alcaldía were bat sampling was done.
Data analysis
To determine if there was a difference in the number of passing count calls among CDMX regions we used a general linear model based on a poisson distribution and using loglink function. To evaluate bat as indicator of the resilience capacities, we used use of information theoretic model (ITM) selection based on Akaike’s Information Criterion. (AIC). This method consist in built a complete model (all variables have influence in dependent variable) and then to evaluate all models constructed and get the best-fitted final model (Richards 2005). We made this model to determine which set of variables (described above) better explained the bat species richness, bat diversity represented by the Shannon diversity index, passing call count, and hunting buzz count. Bat habitat was defined as random effect into models. Also, we constructed individual-based accumulation curves that were both rarefied and extrapolated based on Hill numbers (Chao et al. 2014). These bat community metrics allowed us to detect the differential impact of urbanization on bat species numbers.
In order to model bat species permanence probabilities in the CDMX we used a series of Markov chains which allowed, through an stochastic process to model the future development of change progression of a given population based on a discrete inicial state and assigned probabilities (Caswell 2000). To make the Markov models, we build the transition matrices based on the lower number of bat independent recordings for each species as initial number. To stablish the transition probabilities, we used the coefficients of ITM constructed for bat species richness and assumed three scenarios (1) basal scenario (worst ITM, AIC = 3879.12) was constructed under the assumption that urban condition allows presence of only generalist bat species in low densities, (2) Optimal scenario had the conditions that allow bats presence in low densities but with higher diversity than scenario one. This scenario was the last model ranked among those with a AIC weight < 7, meaning a model with good predictors but not the best one (Ferguson-Gow et al. 2014) and (3) Ideal scenario (best ITM AIC = 3737.3) was the one that allows not only bat presences, but have higher diversity than both 1 and 2 scenarios under urban conditions. To know the variation of models we estimated the likelihood of the transition matrix per species. All analyses were performed in R 2022.02 (Team-R 2022) using vegan 2.6 (Dixon 2003) and iNEXT 2.0.2 (Hsieh et al. 2016) and MuMIn (Bartoń 2023) libraries.
Results
We registered 12 species of bats from four taxonomical families. The most frequently detected family was Vespertillionidae and the least frequently detected was the subfamily Glossophaginae. The most abundant bat species was Myotis velifer (Vespertillionidae), and the least abundant was Nyctimomops macrotis (Molossidae). However, the relative abundance varies depending on the area analyzed.
The Center and East areas had the lower bat abundance (F = 7.60, P = 0.00028), and species richness (F = 12.48; P < 0.001) than other areas of CDMX (Fig. 3). In both areas the most abundant bat species were from Myotis genera. In the north Mormoops megalophylla was the most abundant species, while in the west of the city was Eptesicus fuscus. In the south of the city Myotis volans was abundant, while Pteronotus parnellii was the rarest one.

Differences in bat diversity General lineal model for bat species diversity registered in the Mexico City
The individual-based accumulation species curves showed that all areas except the East reached an asymptote with a data sampling effort of about 200 bat calls. Extrapolation within the rarefaction curve shows that South and West areas could increment at least two more species than registered, while East curve extrapolation indicate that up to 11 species could be found (Fig. 4a). The number of individual calls registered range from 200 to 500 in total, but the sample coverage was similar for all areas (Fig. 4b).

Individual-based rarefaction curves (a) and sampling coverage (b) for the five areas analyzed in the Mexico City. Dashed lines show extrapolations
These patterns of species richness and diversity were differentially determined by habitat, environmental and human variables. In the case of species richness, ITM showed that habitat complexity was the most important variable explaining higher bat species richness. Variables such as light and population negatively affect bat richness. (ITM coefficient of 0.78; Table 1). For species diversity, also habitat complexity had a strong positive influence, but population density affects negatively bat diversity (ITM coefficient of 0.60; Table 1). According with the best-fitted model (ITM coefficient of 0.719), bat activity was positively determined by the proximity to NPA and negatively by population density (Table 1). The hunting buzz was positively explained by the proximity to NPA and habitat complexity (ITM coefficient of 0.7821 (Table 1). All models but species richness was negatively explained by contamination.
Markov chain models predicted that under the basal scenario bat populations in the CDMX would persist but at lower abundances. Only M. velifer, M. volans and N. macrotis will have abundances over 50 individuals. In the optimal scenario species such as E. perotis, M. velifer and T. brasiliensis, will be very abundant, other as L. nivalis become rarer. Under the last scenario the persistence of almost all bat species grow above 50%, but with lower abundances. Bats species such as L. nivalis and M. thysanodes, will become more abundant (Fig. 5).

Markov probabilities of bat permanence under three different habitat scenarios in the Mexico City. Upper panel shows de AIC of each information-theoretical model (ITM) in which was based
Discussion
Our hypothesis posited a negative effect of urbanization on bat species richness, diversity and their permanence, which further would affect CDMX resilience capacities. Although our hypothesis was supported by data, we documented a complex relation between bats and the urban ecosystem. We documented 12 species of four taxonomic families. Before, Avila and Fenton (2005), registered five species of bats, with a similar sampling effort and more recently, García (2018), documented 11 species at the south of the city. However, there are reports indicating that in the CDMX could be up to 23 species of bats (Hortelano-Moncada and Cervantes 2011). Species richness in cities can naturally vary but the influence of urban environment is a determinant as well (Krauel and LeBuhn 2016; Li and Wilkins 2014; Russo and Ancillotto 2015). This changes in bat community, are not surprising, as there is an ongoing novel ecosystem emerging (Barbar et al. 2015). The point is whether the city has the capacities to become resilient, under the face of a taxonomic and functional homogenization that affects directly the ecosystem services and ultimately the productivity and goods that bats provide.
Urban robustness and adaptability
Robustness and adaptability reflects the capacity of a city to maintain its natural conditions ready to bounce back by itself (former) or making wider its urban conditions limits (later) but without changes in its structure (Folke et al. 2010; Meuwissen et al. 2019). We detected almost all bat species in the whole area of the CDMX, contrary to that reported in other studies where bat distributed according to their ecological characteristics (Avila-Flores and Fenton 2005; Li and Wilkins 2014). For example, M. velifer was evenly distributed, this species is commonly found in cities (Krauel and LeBuhn 2016), and it has been documented to respond to spatial heterogeneity and to the vegetation and water source availability like other Myotis species (Fabianek et al. 2011; Krauel and LeBuhn 2016; Threlfall et al. 2012). Mormoops megalophylla was another species we commonly registered in the CDMX, it has been documented that this species avoids forested areas within the cities (Rodríguez-Aguilar et al. 2017), and are able to feed insects attracted by artificial lights (Jung and Kalko 2011). Similarly is the case of T. brasiliensis, this species is considered as highly successful bat species in cities (Armstrong 2008; Li and Wilkins 2014). Thus, it is remarkable that this species was not found in some areas at east of the CDMX.
On the other hand, N. macrotis, was the rarest species documented in this work. This species was relatively abundant at the west of the city, a region with large areas of natural vegetation and an active river. It has been documented that this molossid species based its diet in Lepidoptera, which are more easily found in natural forested areas. Accordingly, this bat species was not found in the east and center of the study area, which is known are of the most inhabited and urbanized areas in de CDMX. This species richness variation could be used as a measure of robustness.
In regard of this, we documented that different variables determine different aspects of bats at community level. For example, bat species richness, was negatively determined by light and density population. In the case of light, there are opposite opinions. In one hand, bats could be positive affected by urban illumination, at least in temperate areas where insectivorous bats are the dominant species. At these places, streetlights play the role of insect’s attraction, which further attract bat species that feed around artificial lights and shape the structure of such urban bat community (Rydell 2006; Schoeman 2016). Nonetheless, there are reports that document the artificial lights negatively affect bats (Laforge et al. 2019; Stone et al. 2009, 2015).
Bat diversity was positively influenced by the proximity of the larger green urban areas and protected areas; both involve vegetation (introduced or native). Former, are represented for citizen parks and some cemeteries that have a spotted distribution in the CDMX. While latter are mainly distributed in the surroundings limits of the city (Supplementary material). Typically, both areas are source of food and roosting areas, but also allow bats getting travel corridors (Gallo et al. 2018; Pearce and Walters 2012).
It has been documented that habitat complexity favors high bat species diversity in natural areas (August 1983). In urban landscapes, bat species show similar patterns (Li and Wilkins 2014; Russo and Ancillotto 2015; Russo et al. 2002; Threlfall et al. 2012). We documented that urban habitat complexity positively influence both species richness and feeding buzz. This is significant because as more species can coexist in an urban area more bat diversity with a further better perform of their ecosystem services (Lewanzik and Voigt 2014; Orozco-Lugo et al. 2013; Parker et al. 2019; Williams-Guillén et al. 2016).
Overall, these results suggest both CDMX’s robustness and adaptability capacities are present but are prone to be improved. Bats “see” a city as a whole landscape, avoiding such areas with characteristics that affect their interactions and using those areas which promote that ones (de Araújo and Bernard 2016; Duchamp et al. 2004; Gallo et al. 2018; Krauel and LeBuhn 2016). Therefore, cities ought be designed to promote bat diversity and their activities in order to attain ecosystem functions and other components of diversity that could use bats as conservation umbrella (Baldock et al. 2019).
Human dimension are dynamic and highly heterogeneous, thus only one variable could not describe all socio-economic processes associated with bat ecology (Li and Wilkins 2014). We used population density and air pollution, as proxy of this dynamics (Alberti et al. 2003). In all cases, population density negatively affects bats. Areas highly inhabited used all the space for habitation or economical activities leaving little space for bat roosting or feeding. Competition for a limited number of high-quality sites in the roost or competition for foraging areas could explain the lower local species richness and diversity in the center and east of the CDMX. On the other hand, bats has frequently been used as bioindicators of metal pollution (Jones et al. 2009; Zukal et al. 2015). For example, in Central Mexico, higher concentrations of Cu and Zn were found in Tadarida brasiliensis, this was associated with vehicular traffic (Ramos-H et al. 2020), but rather there are reports of how air pollution affects bats at community level such as we documented.
Our results documented that different bat species use the urban landscape in very distinctive ways. Roosts could be a factor promoting the presence of molossids in cities (García 2018), because buildings and other structures such as bridges offer many crevice like sites that resemble natural molossid roosts (Bell 1980). Theoretically, every site in the city is accessible to molossids, because can travel long distances to reach foraging areas and fly at very high altitudes (Griffin and Thompson 1982; Wilkins 1989). However, we showed that other characteristics, besides the accessibility, influences bats urban distribution. The most highlighting thing that our data shows is that the different variables we took into account determine differentially the characteristics measured on bat community. Only bat diversity was mainly explained by environmental variables, while such associated with bat interactions were associated with human dimensions.
Urban transformability
Transformability is another resilience capacity; with this capacity urban landscapes could have structural changes in order to improve its resilience. The Markov models allow detecting the impact of CDMX conditions on bat relative abundances (based on bat calls registered), under different scenarios. Our basal scenario represents the most adverse conditions and only two Myotis species resists this condition. In the optimal scenario species such as T. brasiliensis would be abundant, this species has been documented as adapted to urban landscapes (Armstrong 2008; Krauel and LeBuhn 2016; Li and Wilkins 2014). At these two scenarios is highlighted the abundance growth of those persisting species, mirrored the absence of interspecific competition, as a result of depletion of other bat species, which could become easier to find roosting areas or food (Bergeson et al. 2020; Lesinski et al. 2000; Russo and Ancillotto 2015). The ideal scenario showed the higher probabilities of bat persistence, but at lower abundance, which would allowed a richer bat community within the urban landscape as answer to better conditions in the city characteristics making it more resilient (Evelyn et al. 2004; Kelly et al. 2016).
The design of a city for resilience means to get the capacity of understanding all stages of resilience (Desouza and Flanery 2013), but above all this, the issue is to acknowledge that fauna, including human, is confronting a novel emerging ecosystem. Thus, as better as the urban landscape will be designated, the managing of properties emerging will enhance social, economic and ecological values. According to our results, cities could develop looking for habitat complexity. This landscape most includes green areas and water bodies avoiding higher population densities and inadequate artificial lights.
We demonstrated that CDMX barely complain its resilience capacity, but is highly prone to be addressed toward a redesign to become an emergent ecosystem, where bat community monitoring will allow registering how to respond to environmental stressors. Resilience in cities generally refers to the ability to absorb, adapt and respond to changes in the urban system. However, it also needs to meet other key contemporary urban goals such as sustainability, governance and economic development. Bat species could look as a biological issue; nonetheless its ecology allows depict a scenario where, cities as human-constructed ecosystem can be designed as a resilient one that include in its development the conjunction of people and wild species.
Data Availability
Datasets could be accessed through a correspondence author request.
References
Alberti M, Marzluff JM, Shulenberger E, Bradley G, Ryan C, Zumbrunnen C (2003) Integrating humans into ecology: opportunities and challenges for studying urban ecosystems. Bioscience 53:1169–1179
Aldridge HDJN, Rautenbach IL (1987) Morphology, echolocation and resource partitioning in insectivorous bats. J Anim Ecol 56:763–778
Anderson HM, Hutson V, Law R (1992) On the conditions for permanence of species in ecological communities. Am Nat 139:663–668
Armstrong K (2008) Brazilian free-tailed bat (Tadarida brasiliensis). Mammalian Species 4:1–6
August PV (1983) The role of habitat complexity and heterogeneity in structuring tropical mammal communities. Ecology 64:1495–1507
Avila-Flores R, Fenton MB (2005) Use of spatial features by foraging insectivorous bats in a large urban landscape. J Mammal 86:1193–1204
Baldock KCR, Goddard MA, Hicks DM, Kunin WE, Mitschunas N, Morse H, Osgathorpe LM, Potts SG, Robertson KM, Scott AV (2019) A systems approach reveals urban pollinator hotspots and conservation opportunities. Nat Ecol Evol 3:363–373
Barbar F, Werenkraut V, Morales JM, Lambertucci SA (2015) Emerging ecosystems change the spatial distribution of top carnivores even in poorly populated areas. PLoS ONE 10, e0118851
Bartoń K (2023) MuMIn: Multi-Model Inference v 1.47.5
Bell GP (1980) Habitat use and response to patches of prey by desert insectivorous bats. Can J Zool 58:1876–1883
Bergeson SM, Holmes JB, O’Keefe JM (2020) Indiana bat roosting behavior differs between urban and rural landscapes. Urban Ecosyst 23:79–91
Buettner T (2015) Urban estimates and projections at the United Nations: the strengths, weaknesses, and underpinnings of the world urbanization prospects. Spat Demography 3:91–108
Caswell H (2000) Matrix population models. Sinauer Sunderland, MA, USA
Chao A, Gotelli NJ, Hsieh TC, Sander EL, Ma KH, Colwell RK, Ellison AM (2014) Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84:45–67
de Araújo MLVS, Bernard E (2016) Green remnants are hotspots for bat activity in a large Brazilian urban area. Urban Ecosyst 19:287–296
Desouza KC, Flanery TH (2013) Designing, planning, and managing resilient cities: a conceptual framework. Cities 35:89–99
Dixon P (2003) VEGAN, a package of R functions for community ecology. J Veg Sci 14:927–930
Duchamp JE, Sparks DW, Whitaker JJO (2004) Foraging-habitat selection by bats at an urban–rural interface: comparison between a successful and a less successful species. Can J Zool 82:1157–1164
Evelyn MJ, Stiles DA, Young RA (2004) Conservation of bats in suburban landscapes: roost selection by Myotis yumanensis in a residential area in California. Biol Conserv 115:463–473
Fabianek F, Gagnon D, Delorme M (2011) Bat distribution and activity in Montréal Island green spaces: responses to multi-scale habitat effects in a densely urbanized area. Ecoscience 18:9–17
Ferguson-Gow H, Sumner S, Bourke AFG, Jones KE (2014) Colony size predicts division of labour in attine ants. Proc Royal Soc B: Biol Sci 281:20141411
Folke C, Carpenter S, Elmqvist T, Gunderson L, Holling CS, Walker B (2002) Resilience and sustainable development: building adaptive capacity in a world of transformations. AMBIO: A Journal of the Human Environment 31:437–440
Folke C, Carpenter SR, Walker B, Scheffer M, Chapin T, Rockström J (2010) Resilience thinking: integrating resilience, adaptability and transformability. Ecol Soc 15
Furlonger CL, Dewar HJ, Fenton MB (1987) Habitat use by foraging insectivorous bats. Can J Zool 65:284–288
Gallo T, Lehrer EW, Fidino M, Kilgour RJ, Wolff PJ, Magle SB (2018) Need for multiscale planning for conservation of urban bats. Conserv Biol 32:638–647
Gaisler J, Zukal J, Rehak Z, Homolka M (1998) Habitat preference and flight activity of bats in a city. J Zool 244:439–445
García Y (2018) Localización Y descripción De Los refugios urbanos utilizados por murciélagos residentes y su relación con las áreas verdes, en la zona centro-sur de la ciudad de México, México. Facultad De Ciencias. UNAM, p 95
Gehrt SD, Chelsvig JE (2003) BAT ACTIVITY IN AN URBAN LANDSCAPE: PATTERNS AT THE LANDSCAPE AND MICROHABITAT SCALE. Ecol Appl 13:939–950
Griffin DR, Thompson D (1982) High altitude echolocation of insects by bats. Behav Ecol Sociobiol 10:303–306
Gunderson LH (2001) Panarchy: understanding transformations in human and natural systems. Island press
Holling CS (1973) Resilience and stability of ecological systems. Annu Rev Ecol Syst 4:1–23
Holling CS (1996) Engineering resilience versus ecological resilience. Eng within Ecol Constraints 31:32
Hortelano-Moncada Y, Cervantes FA (2011) Diversity of wild mammals in a megalopolis: Mexico City, Mexico. Changing diversity in changing Environment. IntechOpen
Hsieh TC, Ma KH, Chao A (2016) iNEXT: an R package for rarefaction and extrapolation of species diversity (H ill numbers). Methods Ecol Evol 7:1451–1456
Jones G, Jacobs DS, Kunz TH, Willig MR, Racey PA (2009) Carpe noctem: the importance of bats as bioindicators. Endanger Species Res 8:93–115
Jung K, Kalko EKV (2011) Adaptability and vulnerability of high flying neotropical aerial insectivorous bats to urbanization. Divers Distrib 17:262–274
Kelly RM, Kitzes J, Wilson H, Merenlender A (2016) Habitat diversity promotes bat activity in a vineyard landscape. Agric Ecosyst Environ 223:175–181
Kosmala M, Wiggins A, Swanson A, Simmons B (2016) Assessing data quality in citizen science. Front Ecol Environ 14:551–560
Krauel JJ, LeBuhn G (2016) Patterns of bat distribution and foraging activity in a highly urbanized temperate environment. PLoS ONE 11:e0168927
Kunz TH, Fenton MB (2005) Bat ecology. University of Chicago Press
Kunz TH, Braun de Torrez E, Bauer D, Lobova T, Fleming TH (2011) Ecosystem services provided by bats. Ann N Y Acad Sci 1223:1–38
Laforge A, Pauwels J, Faure B, Bas Y, Kerbiriou C, Fonderflick J, Besnard A (2019) Reducing light pollution improves connectivity for bats in urban landscapes. Landscape Ecol 34:793–809
Lesinski G, Fuszara E, Kowalski M (2000) Foraging areas and relative density of bats (Chiroptera) in differently human transformed landscapes. Z fur Saugetierkunde 65:129–137
Lewanzik D, Voigt CC (2014) Artificial light puts ecosystem services of frugivorous bats at risk. J Appl Ecol 51:388–394
Li H, Wilkins KT (2014) Patch or mosaic: bat activity responds to fine-scale urban heterogeneity in a medium-sized city in the United States. Urban Ecosyst 17:1013–1031
Meerow S, Newell JP, Stults M (2016) Defining urban resilience: a review. Landsc Urban Plann 147:38–49
Mendes ES, Fonseca C, Marques SF, Maia D, Ramos Pereira MJ (2017) Bat richness and activity in heterogeneous landscapes: guild-specific and scale-dependent? Landscape Ecol 32:295–311
Meuwissen MPM, Feindt PH, Spiegel A, Termeer CJAM, Mathijs E, de Mey Y, Finger R, Balmann A, Wauters E, Urquhart J (2019) A framework to assess the resilience of farming systems. Agric Syst 176:102656
Neubaum DJ, Wilson KR, O’Shea TJ (2007) Urban maternity-roost selection by big brown bats in Colorado. J Wildl Manag 71:728–736
Newson SE, Evans HE, Gillings S (2015) A novel citizen science approach for large-scale standardised monitoring of bat activity and distribution, evaluated in eastern England. Biol Conserv 191:38–49
Norris FH, Stevens SP, Pfefferbaum B, Wyche KF, Pfefferbaum RL (2008) Community resilience as a metaphor, theory, set of capacities, and strategy for Disaster readiness. Am J Community Psychol 41:127–150
Orozco-Lugo L, Guillén-Servent A, Valenzuela-Galván D, Arita HT (2013) Descripción De Los pulsos de ecolocalización de once especies de murciélagos insectívoros aéreos de una selva baja caducifolia en Morelos. México Therya 4:33–46
Parker KA, Springall BT, Garshong RA, Malachi AN, Dorn LE, Costa-Terryll A, Mathis RA, Lewis AN, MacCheyne CL, Davis TT (2019) Rapid increases in bat activity and diversity after wetland construction in an urban ecosystem. Wetlands 39:717–727
Pataki DE (2015) Grand challenges in urban ecology. Front Ecol Evol 3:57
Pearce H, Walters CL (2012) Do green roofs provide habitat for bats in urban areas? Acta Chiropterologica 14:469–478
Rainho A, Palmeirim JM (2011) The importance of distance to resources in the spatial modelling of bat foraging habitat. PLoS ONE 6, e19227
Ramos Pereira MJ, Fonseca C, Aguiar LMS (2018) Loss of multiple dimensions of bat diversity under land-use intensification in the Brazilian Cerrado. Hystrix the Italian Journal of Mammalogy 29:25–32
Ramos-H D, Medellín RA, Morton-Bermea O (2020) Insectivorous bats as biomonitor of metal exposure in the megalopolis of Mexico and rural environments in Central Mexico. Environ Res 185:109293
Richards SA (2005) Testing ecological theory using the information-theoretic approach: examples and cautionary results. Ecology 86:2805–2814
Rodríguez-Aguilar G, Orozco-Lugo CL, Vleut I, Vazquez L-B (2017) Influence of urbanization on the occurrence and activity of aerial insectivorous bats. Urban Ecosyst 20:477–488
Russo D, Ancillotto L (2015) Sensitivity of bats to urbanization: a review. Mammalian Biology 80:205–212
Russo D, Jones G, Migliozzi A (2002) Habitat selection by the Mediterranean horseshoe bat, Rhinolophus euryale (Chiroptera: Rhinolophidae) in a rural area of southern Italy and implications for conservation. Biol Conserv 107:71–81
Rydell J (2006) Bats and their insect prey at streetlights. Ecol Consequences Artif Night Lighting 2:43–60
Schoeman MC (2016) Light pollution at stadiums favors urban exploiter bats. Anim Conserv 19:120–130
Stahlschmidt P, Brühl CA (2012) Bats as bioindicators–the need of a standardized method for acoustic bat activity surveys. Methods Ecol Evol 3:503–508
Stone EL, Jones G, Harris S (2009) Street lighting disturbs commuting bats. Curr Biol 19:1123–1127
Stone EL, Harris S, Jones G (2015) Impacts of artificial lighting on bats: a review of challenges and solutions. Mammalian Biology 80:213–219
Team-R (2022) R: A language and environment for statistical computing
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, Jeltsch F (2004) Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr 31:79–92
Threlfall CG, Law B, Banks PB (2012) Sensitivity of insectivorous bats to urbanization: implications for suburban conservation planning. Biol Conserv 146:41–52
Tomassini A, Colangelo P, Agnelli P, Jones G, Russo D (2014) Cranial size has increased over 133 years in a common bat, Pipistrellus kuhlii: a response to changing climate or urbanization? J Biogeogr 41:944–953
Walker B, Holling CS, Carpenter SR, Kinzig A (2004) Resilience, adaptability and transformability in social–ecological systems. Ecol Soc 9
Wilkins KT (1989) Tadarida brasiliensis. Mamm Species 331:1–10
Williams-Guillén K, Olimpi E, Maas B, Taylor PJ, Arlettaz R (2016) Bats in the anthropogenic matrix: challenges and opportunities for the conservation of Chiroptera and their ecosystem services in agricultural landscapes. Bats in the Anthropocene: conservation of bats in a changing world. Springer, Cham, pp 151–186
Zukal J, Pikula J, Bandouchova H (2015) Bats as bioindicators of heavy metal pollution: history and prospect. Mammalian Biology 80:220–227
Acknowledgements
We thank all citizen scientists that support our bat sampling. Many biology students from Facultad de Estudios Superiores Zaragoza help in different parts of the project. Thanks to all of them. We thank observations of two anonymous reviewers, all comments significantly improve the paper. GGG was supported by National Autonomous University of Mexico through the projects PAPIIT IA206018 and PAPIME PE216219.
Funding
GGG was supported by UNAM through projects PAPIIT IA206018 and PAPIME PE216219.
Author information
Authors and Affiliations
Contributions
Authors equally contributed to all sections of paper.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Competing interests
The authors declare they have no competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Conflicts of interest
The authors declare they have no conflict of interest with any person or institution that could influence the work reported in this paper.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gutiérrez-Granados, G., Rodríguez-Zúñiga, M.T. Bats as indicators of ecological resilience in a megacity. Urban Ecosyst 27, 479–489 (2024). https://doi.org/10.1007/s11252-023-01462-5
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01462-5