
Abstract
Intensification in land-use and farming practices has had largely negative effects on bats, leading to population declines and concomitant losses of ecosystem services. Current trends in land-use change suggest that agricultural areas will further expand, while production systems may either experience further intensification (particularly in developing nations) or become more environmentally friendly (especially in Europe). In this chapter, we review the existing literature on how agricultural management affects the bat assemblages and the behavior of individual bat species, as well as the literature on provision of ecosystem services by bats (pest insect suppression and pollination) in agricultural systems. Bats show highly variable responses to habitat conversion, with no significant change in species richness or measures of activity or abundance. In contrast, intensification within agricultural systems (i.e., increased agrochemical inputs, reduction of natural structuring elements such as hedges, woods, and marshes) had more consistently negative effects on abundance and species richness. Agroforestry systems appear to mitigate negative consequences of habitat conversion and intensification, often having higher abundances and activity levels than natural areas. Across biomes, bats play key roles in limiting populations of arthropods by consuming various agricultural pests. In tropical areas, bats are key pollinators of several commercial fruit species. However, these substantial benefits may go unrecognized by farmers, who sometimes associate bats with ecosystem disservices such as crop raiding. Given the importance of bats for global food production, future agricultural management should focus on “wildlife-friendly” farming practices that allow more bats to exploit and persist in the anthropogenic matrix so as to enhance provision of ecosystem services. Pressing research topics include (1) a better understanding of how local-level versus landscape-level management practices interact to structure bat assemblages, (2) the effects of new pesticide classes and GM crops on bat populations, and (3) how increased documentation and valuation of the ecosystem services provided by bats could improve attitudes of producers toward their conservation.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Ecosystem Service
- Agroforestry System
- Coffee Berry
- Agricultural Intensification
- Valuable Ecosystem Service
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Agricultural areas cover approximately 40 % of our planet’s terrestrial ecosystems (FAOSTAT 2011), with the 5 billion ha of land under farming and grazing now surpassing the extent of the world’s forested areas (Robertson and Swinton 2005; Power 2010). Agricultural areas are expected to continue to expand with increasing human population growth and resultant resource use: Low- and middle-income countries will experience a 100 % increase in demand for agricultural products by 2050 (Defries et al. 2010; FAO 2011). In the face of increasing pressure on natural resources, the conservation of remaining natural areas is critical for the survival of multitudes of species. However, the ubiquity of agriculture means that farmland cannot be ignored in the context of landscape-level approaches to biodiversity conservation (Vandermeer and Perfecto 2007; Loos et al. 2014).
A growing body of research demonstrates that not only do some agricultural systems harbor high levels of biodiversity and provide a variety of ecosystem services (Tilman 1999; Foley et al. 2005; Tscharntke et al. 2005), but also that characteristics of these agricultural systems may have profound effects upon remaining natural areas (Perfecto and Vandermeer 2010). Agricultural matrices can vary drastically in their quality and permeability, impacting dispersal rates, and hence, long-term population stability of organisms found in less disturbed areas (Ricketts 2001; Laurance 2008; Perfecto and Vandermeer 2010; Tscharntke et al. 2012). On a local scale, different agricultural management approaches often coexist. Some rely on varying chemical inputs (pesticides, fertilizer), or novel plant types (e.g., genetically modified crops incorporating genes for characteristics such as insecticide functions), resulting in environmental contamination, pollution, and dissemination of toxins that could negatively impact biodiversity across multiple spatial scales (Nelson et al. 2009; Power 2010). As a consequence, agricultural management has effects not only on biodiversity, but also on human health and economies.
In the tropics, the expansion of export-oriented agriculture results from population growth and shifts in consumption patterns of developing nations, and is carried out mostly to the detriment of old growth forests and extensively managed grasslands such as pastures (Defries et al. 2010; Lambin and Meyfroidt 2011). As a consequence, croplands are still expanding dramatically, and agricultural practices are likely to further intensify in the near future (more chemical and mechanical inputs, reliance on genetically modified plants with novel manufactured traits). Short-term increases in yield will come at the cost of reduced structural and taxonomic diversity within agricultural systems (Loos et al. 2014) and concomitant loss of crucial ecosystem services.
An additional factor affecting agriculture in the Anthropocene is climate change and the need to adapt cultures to novel environmental conditions: Many areas may become unsuitable for cultivation of their current dominant crops, while extreme weather events may result in reduced yields. Resulting declines in calorie availability, particularly in the developing world (Nelson et al. 2009), will increase the need for agricultural practices that meet both productivity and sustainability goals (Tilman et al. 2002; McShane et al. 2011; Tscharntke et al. 2012). These trends portend major shifts in land-use patterns (Lambin and Meyfroidt 2011) and hence biodiversity, with agricultural intensification , forest and tree roost loss anticipated to have particularly negative effects on bat species richness, abundance, and functional diversity (Fischer et al. 2009, 2010; Jones et al. 2009).
These emerging trends pose major threats to farmland bat assemblages and populations (Jones et al. 2009; Kunz et al. 2011) and could negatively impact human populations by altering the ecosystem services that bats provide. Thus, there is a critical need to assess how agricultural management affects bat populations, and how affected bat populations will in turn affect agricultural production. In this chapter, we review the effects of agricultural land use and management on bat assemblages and the behavior and ecology of individual bat species at field, farm, and landscape scales (Vickery and Arlettaz 2012). We also review the developing literature on ecosystem services—and disservices—provided by bats in agricultural areas. Finally, we synthesize this information to suggest key management recommendations necessary to maintain bat populations in agricultural landscapes and highlight critical knowledge gaps that must be resolved in order to conserve bat diversity and ecosystem functions in a planet increasingly dominated by food production.
2 Methods
We used the Web of Knowledge, Google Scholar, and PubMed search engines to locate publications with the keywords “bats” AND “agriculture,” “agroforestry,” “farm,” and “farmland.” Given the potential importance of bats in provisioning ecosystem services in agricultural areas, we also searched for “bats” AND “ecosystem services,” “pollination,” “pest consumption,” “pest control,” and “pest limitation.” The majority of sources stemmed from peer-reviewed publications, although we also included Master’s and Ph.D. theses and published reports if results from the study in question were not available as journal articles. We also inspected the bibliographies of relevant publications. Each co-author focused on a specific geographic area (RA, assisted by Olivier Roth: Europe; BM: Australia and tropical Asia; EO: temperate North America; PT: sub-Saharan Africa; KWG: tropical Americas). Our searches were limited to publications with English language text or summaries. We focused on agriculture and animal husbandry for the production of calories for human or animal consumption, excluding forestry systems dedicated to timber or fiber production (see Law et al., Chap. 4), studies in which fallows or abandoned fields were the only agricultural systems investigated, as well as investigations that focused on fragmentation without explicit consideration of the effect of agricultural matrix (see Meyer et al., Chap. 3).
We divided results from the literature search into two broad categories of investigations: (1) How agricultural practices affect bat assemblages, ecology, behavior, and/or physiology; and (2) how bats affect agriculture through the provision of ecosystem services such as pollination and pest suppression. Within the first category, most studies addressed effects of land conversion and agricultural management on bat assemblage structure, abundance, activity levels, and behavior. We further subdivided results to consider habitat conversion to agriculture and agricultural intensification. We define agricultural intensification as consisting of at least one of the following: decreased structural complexity of native vegetation (natural and seminatural elements structuring the landscapes such as woodland patches and hedges), increased application of agrochemicals (pesticides, fertilizer), increased crop plant density, increased mechanization, or increased reliance on GM plants. We reviewed results from searches to locate studies which contrasted aspects of bat assemblage structure, abundance, activity, ranging behavior, or diet in either natural and agricultural habitat, or different agricultural systems of contrasting management.
To better quantify the responses of bats to habitat conversion and agricultural intensification across multiple disparate studies, we conducted a meta-analysis . We emphasize that this meta-analysis is based on correlational studies, rather than from controlled experiments; because assignment of treatment locations is not randomized in the majority of these studies, confounding factors could result in spurious effect sizes (Egger et al. 1998). We thus view our meta-analysis as a tool for exploring trends across a diverse suite of studies, with limited conclusive power. We reviewed studies for the inclusion of mean values for at least one response variable in both natural and agricultural areas, or two or more agricultural areas of differing intensification; 32 studies using mist netting, harp trapping, acoustic monitoring, or a combination of these methods included appropriate data. We classify the response variable metrics into two separate categories for analyses, measures of species richness and measures of relative activity or abundance (i.e., pass rates from acoustic monitoring or capture rates from mist netting). We also consider habitat conversion and intensification responses separately.
For each pairwise comparison (natural–agricultural, or agricultural–agricultural), we calculated the effect size as the log odds ratio of the mean value from the lower intensity system divided by the mean from the higher intensity system. Thus, a positive effect size indicates higher species richness or activity/abundance in natural versus agricultural areas or lower intensity versus higher intensity agriculture. We followed García-Morales et al. (2013) and considered mean effect sizes with 95 % confidence intervals that did not include 0 as indicative of a significant effect. In the case of studies comparing multiple natural or agricultural habitats or presenting means for multiple species or species groups (i.e., producing multiple pairwise comparisons for any given combination of metric and response type), we averaged the odds ratio to avoid pseudo replication. Due to the diverse nature of the studies and a lack of clarity about numbers of replicates in some studies, we did not weight studies by sample size or replicates. For our analysis, we thus considered each study as an equally weighted case for the final model. We conducted analyses in R Version 3.0.2 (R Development Core Team 2013) using the packages lme4 and lmerTest. This diverse set of studies includes different methods (e.g., acoustic monitoring versus mist netting) from different regions with ecologically and taxonomically characteristic bat assemblages. To account for some of this variation, we included study method and continent as random effects. Fixed factors included latitudinal zone (temperate, subtropical, and tropical) and whether or not the high-intensity system comprised an agroforestry system (including monocultural orchards).
We also located several studies on ecotoxicology and demography, focusing on the effects of pesticide and GMOs use on bats. A complete review of the effects of pesticides on bats is beyond the scope of this chapter, particularly since bats and contaminants have received recent reviews (O’Shea and Johnston 2009; Bayat et al. 2014). We therefore focus on studies that explicitly link bat agrochemical exposure to changes in bat populations. Similarly, although fertilizers comprise a large portion of the chemical inputs to agriculture, their impacts on bats are indirect.
In considering the benefits of bats for agricultural production (i.e., crop yield), we focus on the provision of two ecosystem services: agricultural pest limitation by insectivorous bats and pollination by tropical bats. We did not consider their role as seed dispersers since human management of farmland vegetation limits the effect and value of bat seed dispersal. Similarly, although bat pollination is key for the unmanaged reproduction of several economically important crops, such as bananas and agaves (Kunz et al. 2011), we did not consider these particular crops because they are mostly propagated vegetatively in such plantations. We instead focus on crops that are almost exclusively reliant on bat pollination under standard cultivation practices. Multiple investigations have characterized the diets of insectivorous bats at the order level, claiming potential consumption of pest insects. To more confidently assess consumption of insects damaging crops, we focused on studies in which known (species level identity) or probable (family level identity) agricultural pests were identified from feces of bats foraging in farms or areas dominated by agriculture. We exclude dietary studies that have sampled exclusively from natural habitats or do not describe the agricultural systems within which bats may have been foraging. We also briefly contrast these with ecosystem disservices of bats in agricultural areas. Bats are associated with costs to agriculturalists, particularly in the subtropics and tropics where frugivorous bats raid crops and sanguivorous bats attack domestic livestock. As with other sections, we focus on direct impacts on productive systems and do not consider the impacts of bat transmission of disease except where it directly impacts agriculture.
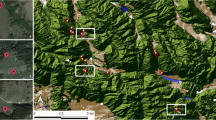
The majority of the nearly 140 investigations reviewed in this chapter have been conducted in temperate North America and Europe (Fig. 6.1). The bulk of studies documenting how habitat conversion or agricultural intensification affects bats has been conducted in Europe and the Neotropics (Fig. 6.1, Table 6.1). Within temperate zones, studies have focused mainly on annual cultivars and pasture, while research in tropical areas is dominated by studies on agroforestry systems, particularly coffee and cacao. Results on ecotoxicology of farmland bats come primarily from North America. Studies demonstrating the consumption of agricultural pests also derive primarily from North America, whereas studies of other ecosystem services provided by bats are limited to the tropics.

Locations of studies on effects of habitat conversion or agricultural intensification (red diamonds) on bats, pesticide contamination (pink triangles) on bats, and ecosystem services (green squares) provided by bats in agriculture
3 Effects of Agricultural Intensity on Bat Assemblage Structure, Behavior, and Ecology
We found 70 studies addressing the effects of habitat conversion or management on the assemblage structure, behavior, or ecology of bats. Fifty-two studies assessed bats in both natural and agricultural areas. Twenty-two studies (42 %) demonstrated negative effects of habitat conversion, twelve (23 %) showed variable responses (e.g., only some species or ensembles declined, different agricultural systems were associated with different effects), twelve (23 %) showed increased richness, activity, or abundance in agricultural areas, and six (12 %) showed little or no difference between agricultural and natural areas. Forty-five studies addressed some aspect of agricultural intensification, with 38 of these (84 %) documenting a negative effect of intensification on bats, four showing variable or neutral (9 %) responses, while three studies (7 %) documented increases in bat richness, abundance, or activity in more intensive systems.
Response variables differ in response to habitat conversion and agricultural intensification (Fig. 6.2, Table 6.2), with measures of species richness showing no significant change between treatments. In contrast, measures of relative activity and abundance show stronger responses (Fig. 6.2). Agroforestry systems are more structurally similar to the original non-anthropogenic land uses, making them less intensive than annual crops dominated by one plant species or pasture systems lacking structural complexity. This relationship presumably explains why agricultural systems that incorporate trees and other large woody perennials on farms and throughout the agricultural landscape have little effect on bat activity and abundance (Fig. 6.2). Agroforestry systems appear to mitigate negative effects on bat assemblages in cases of both habitat conversion and agricultural intensification (Table 6.2).

Mean effect size (log odds ratio, circles) ±95 % CI of relative abundance and activity (left) and species richness (right) of habitat conversion versus agricultural intensification (top row), and of contrasts (both habitat conversion and agricultural intensification) with and without agroforestry systems (bottom row). Positive effect sizes indicate reductions in relative abundance and activity or species richness in response to habitat conversion and intensification
Several studies have considered the effects of agricultural management at landscape scales versus focusing exclusively on farm-level management practices (Estrada et al. 1993; Ekman and de Jong 1996; Verboom and Huitema 1997; Numa et al. 2005; Faria et al. 2006, 2007; Faria and Baumgarten 2007; Fuentes-Montemayor et al. 2011; Boughey et al. 2011; Maas et al. 2013). Within agricultural areas, bat activity increases with proximity to natural areas (Estrada et al. 1993; Verboom and Huitema 1997; Boughey et al. 2011) and in less fragmented landscapes (Fuentes-Montemayor et al. 2011; Frey-Ehrenbold et al. 2013) or in landscapes with more natural elements such as hedgerows and woodlots (Verboom and Huitema 1997).
Agricultural areas also serve as matrix habitat connecting fragmented non-anthropogenic habitats. Although one study has suggested that landscapes dominated by crops and open fields have a stronger negative influence on bats than water (Ekman and de Jong 1996), a recent analysis of bat responses to isolation on islands versus in forest fragments embedded in agricultural matrix suggests that the anthropogenic matrix is more permeable than water matrix (Mendenhall et al. 2014). Thus, agricultural intensification at the landscape level should make the matrix less permeable due to the reduction of natural resources and structural elements such as trees, affecting not only the persistence of bats in fragmented landscapes, but also the degree to which bat assemblages show a negative response to agriculture. A few investigations have confirmed such interactions between farm- and landscape-level intensification: Intensification in cacao matrices in Brazil (Faria et al. 2006, 2007; Faria and Baumgarten 2007) and coffee matrices in Colombia (Numa et al. 2005) resulted in reductions in the species richness and abundance of bats in diverse shade agroforests relative to forest fragments. In Europe, effects of landscape management on bat assemblage structure and ecology in temperate landscapes dedicated to the production of annual crops remain largely unexplored compared to the extensive information available at the field and farm scales.
4 Pesticide Impacts on Bat Populations
Agricultural intensification may remove potential habitat for bats and their prey; the effects of increased agrochemical inputs, such as increased exposure and changes in prey availability, may put resident bats under further pressure. Bats may directly consume pesticides by feeding on fruits, flowers, and arthropods exposed to chemical application. Even bats foraging outside of agricultural areas can be exposed to pesticides via biomagnification as residues are incorporated into the tissues of organisms at higher trophic levels (Bayat et al. 2014).
Investigations of exposure of bats to pesticides and its effects on physiology and mortality first appeared in the 1970s, amid a wave of growing concern regarding the effects of organochlorine pesticides (e.g., DDT, DDE, dieldrin, lindane, endosulfan, aldrin) on ecosystems and observations of declining bat populations at high-profile sites such as the Carlsbad Caverns in New Mexico, USA (Clark 1988, 2001). In some cases, DDT and other organochlorines were even applied directly to bat roosts in efforts to exterminate “vermin” (Kunz et al. 1977), and declines in high-profile bat colonies were linked to organochlorine use (Clark et al. 1978; Clark 2001). Even sublethal exposure to pesticides can have negative consequences for bats, resulting in increased metabolic rates (Swanepoel et al. 1998), and ingestion of pesticide residues on arthropods may poses a potential reproductive risk to certain bat species (Stahlschmidt and Brühl 2012).
Organochlorine residues have been documented in bats in a wide variety of both agricultural and non-agricultural landscapes, although several studies have found increased contaminant loads in bats sampled near agricultural areas (Clark and Prouty 1976; White and Krynitsky 1986) or near sites of pesticide manufacture (O’Shea et al. 2001). In some cases, temporal changes in levels of different contaminants reflect shifts in local agricultural practice as farmers adopt new pesticide regimes (Miura et al. 1978; Clark et al. 1980). Organochlorines are notorious for their persistence in ecosystems, and a variety of studies demonstrate that bats continue to harbor these contaminants in their tissues 20–30 years after the use of these pesticides was banned in sampling areas (Clawson and Clark 1989; Guillén et al. 1994; Schmidt et al. 2000; Sasse 2005). In some cases, persistence may reflect the continued use of these pesticides in lower income nations, as may be the case for the migratory Tadarida brasiliensis (Thies and Thies 1997; Bennett and Thies 2007). Investigations in India (Senthilkumar et al. 2001) and Benin (Stechert et al. 2014) have detected levels or metabolites of organochlorines in bat samples indicative of continued recent use in these regions, especially to fight against malaria. Furthermore, pesticide standards vary between different countries, application often appears to occur non-selectively, and farmers with limited training (especially in developing countries, where agricultural expansion is greatest) are likely to be unaware of the multitude of negative nontargeted environmental impacts affecting human health and biodiversity (Tilman et al. 2001; Yadav 2010).
Despite the clear negative impacts of organochlorines on bats, the effects of agrochemical classes such as pyrethroids and neonicotinoids remain largely unknown (O’Shea and Johnston 2009; Quarles 2013; Bayat et al. 2014), although recent research demonstrates a negative impact on birds (Hallmann et al. 2014). In North America, pesticide contamination has been implicated in bat mortality associated with the fungal pathogen causing white-nose syndrome (WNS), since pesticide load can lead to immunosuppression and endocrine disruption that could make bats more vulnerable to infection (Kannan et al. 2010). “Back of the envelope” calculations suggest declines in bat populations attributed to WNS could translate into an additional 1320 metric tons of insects escaping predation each year (Quarles 2013). The trickle-down impacts on agricultural production could be substantial, although quantitative evidence is lacking. The effects of GM crops incorporating insecticidal traits have been investigated largely in the context of the provisioning of predation services (Federico et al. 2008; Lopez-Hoffman et al. 2014; see next section); however, declines in pest numbers associated with the use of these crops could result in population declines of insectivorous bats (Lopez-Hoffman et al. 2014).
5 Ecosystem Services Provided by Bats in Agricultural Systems
5.1 Insectivorous Bats and Pest Limitation
Of the potential ecosystem services provided by bats, their role in consuming insect pests has received the most attention within agricultural systems. Insectivorous bats have a global distribution and have long been identified as key suppressors of arthropod pests in agricultural systems (Kunz et al. 2011). However, surprisingly little evidence exists quantifying the impact of their predation on arthropod populations, plant damage, or its economic value (Boyles et al. 2013; Maas et al. 2013). Several studies have characterized diets of insectivorous bats (reviewed by Kunz et al. 2011), and the recent development of DNA-based methods for dietary analysis provides an unprecedented amount of detail on the composition of bat diets and allows for the identification of individual pest species. Although few studies have documented direct impacts of bat predation on agricultural pests, an increasing body of evidence documents pest consumption, impacts on arthropods, and estimates of direct economic impacts.
We review 15 studies documenting the consumption of known or probable crop pests by insectivorous bats (Table 6.3). The diets of temperate North American insectivores have received particular attention. Many bat species consume lepidopterans, and studies in North America demonstrate bat predation on devastating pests such as corn earworm (Helicoverpa zea) and fall armyworm (Spodoptera frugiperda) moths (Lee and McCracken 2005; McCracken et al. 2012). Bat species across the world feed on folivorous beetles from a variety of damaging families and species, particularly weevils, leaf beetles, and scarab beetles. Bats may also be underappreciated predators of hemipteran pests, with many studies demonstrating consumption of leafhoppers, froghoppers, spittle bugs, and stink bugs. We emphasize that direct consumption alone is not sufficient to prove that bats are limiting insect pests: Damaging insects may comprise a small proportion of the diet, and nearly every study summarized in Table 6.3 also demonstrated consumption of the predatory arthropods that comprise part of the assemblage of natural enemies. Such intraguild predation could counteract the pest-limiting effects of bat insectivory (Brashares et al. 2010), although herbivores generally comprise the majority of diet by volume in investigations using fecal pellet dissections (Kunz et al. 2011). That the relative abundance, diets, and movements of bats may track populations of agricultural pests (Lee and McCracken 2005; McCracken et al. 2012; Taylor et al. 2013b) suggests that many species are indeed preying heavily on herbivorous insects. This has been assessed in mouse-eared bats, Myotis spp., that track cyclic, massive local aggregations of cockchafers known since centuries for the damages they cause to fruit trees in Central Europe (Arlettaz 1996; Arlettaz et al. 2001).
During lactation, small bat species consume 75 % to over 100 % of their body weight each night (Kurta et al. 1989; Kunz et al. 1995, 2011), and a single maternity colony of 1 million Brazilian free-tailed bats is capable of consuming over 8 tons of insects per night (Kunz et al. 2011). These numbers suggest the staggering potential for bat predation to limit pest insect populations and provide a valuable ecosystem service for agricultural production. Until recently, surprisingly little work had quantified the impact of bat predation on insect biomass (Maas et al. 2015). Exclosure studies have long been a mainstay for studying the impacts of bird predation; however, it was widely assumed that such methods would not be suitable to measure the impact of bat insectivory, due to the misconception that all insect eating bats take highly mobile, flying prey. However, bats capable of gleaning insect prey from substrates exist throughout the world, and their impacts could be monitored via exclosure studies and disentangled from those of birds. This approach has been used fruitfully in the past five years, demonstrating significant increases in arthropod density when bats are absent, in agroecosystems (Williams-Guillén et al. 2008; Maas et al. 2013), reforestation (Morrison and Lindell 2012), and natural forests (Kalka et al. 2008). In Mexican polycultural shade coffee, arthropod densities on coffee plants during the rainy season nearly doubled in the absence of bats, with marked increases in densities of hoppers, katydids, cockroaches, and beetles (Williams-Guillén et al. 2008). However, no effects on plant damage were observed in that study, perhaps as a result of the short duration of the study or release of spiders and other arthropod predators. In Indonesian shade cacao, excluding bats resulted in a 29 % increase in arthropod numbers (Maas et al. 2013). Although herbivory did not differ significantly between cacao plantations with different levels of shade or proximities to primary habitats within the landscape, exclosure of bats resulted in a significant decrease in yields, with the effects of bird and bat predation together valued at an astonishing US $730 per ha and year (bat predation was valued at US $520 per ha and year). However, the effects of bat predation on crop pests are not universal: An exclosure study in Costa Rican coffee found that excluding bats alone had virtually no effect on the density or damage caused to beans by the devastating coffee berry borer (Karp et al. 2013).
Exclosure studies are not suitable to measure the impact of high-flying insectivores, such as molossids. However, careful extrapolations taking into account bat feeding rates, population sizes, pest reproduction, and survivorship, and the costs of inputs allow for estimation of the economic impact of predation for other bats, particularly molossids forming large colonies. Cleveland et al. (2006) estimate that Mexican free-tailed bats (T. brasiliensis) feeding on the cotton bollworm moth in Texas provide pest limitation services worth roughly US $183 per ha and year to cotton growers. Extending these estimates to agricultural areas throughout the USA suggests that bat predation could have a value of nearly US $23 billion annually (Boyles et al. 2011). These benefits hold for both conventional and transgenic cotton (Federico et al. 2008), although the introduction of Bt cotton (a genetically modified organism whose tissues produce an insecticide derived from the bacterium Bacillus thuringiensis), coupled with reduced area in cotton cultivation, has led to a decline in the overall value of this pest limitation service (Lopez-Hoffman et al. 2014).
Valuation of bat-mediated pest suppression is limited for staple crops and for sites outside the southern USA. In northern Mexico, the impact of T. brasiliensis predation on avoided agricultural costs across a variety of staple and commodity crops was estimated at a far more modest $19 per ha and year (Gándara Fierro et al. 2006). In Thailand, the value of wrinkle-lipped bat (Tadarida plicata) predation on a major rice pest, the white-back planthopper (Sogatella furcifera), was estimated to have a monetary value of $1.2 million annually (Wanger et al. 2014). This estimate results in a seemingly paltry $0.13 per ha and year value considered against Thailand’s 8.7 million ha (Redfern et al. 2012) of rice paddies, but in this case an economic approach obscures the true value of the service: This single bat species prevents the loss of nearly 2900 metric tons of rice per year, enough to feed Thailand’s entire population of 66.8 million people for a week. Such investigations underscore the potentially grave consequences for human food security should global bat populations continue declining (Kunz et al. 2011).
5.2 Nectarivorous Bats and Pollination Services
Pollination services to crops by bats are poorly documented. Bats are key pollinators of wild Agave and Musa spp. (Kunz et al. 2011). Although these plants are propagated vegetatively under cultivation, bat pollination plays a critical role in sustaining genetic diversity in the wild relatives of these domestic species, a key aspect of maintaining future food security (Hopkins and Maxted 2011). Within the Americas, several bat pollinated cacti are commercially important fruit species (Kunz et al. 2011). Several species of the hemiepiphytic cactus Hylocereus (pitahaya, dragonfruit) endemic to the Neotropics are now cultivated worldwide. In Mexico, visitation of Hylocereus undatus fruits by bats resulted in significantly higher fruit set than did visitation by diurnal pollinators (Valiente-Banuet et al. 2007). Although H. undatus is self-compatible, other species such as H. costaricensis (an important fruit crop in southern Mesoamerica) apparently rely on pollination by bats and sphingid moths (Weiss et al. 1994; Le Bellec et al. 2006). Nectarivorous bats, particularly the cave nectar bat (Eonycteris spelaea) feed on the flowers of tree beans or petai (Parkia spp.) (Bumrungsri et al. 2008a, b, 2013) and durian (Durio zibethinus) (Bumrungsri et al. 2008b), pollinating these plants in the process. The economic value of this pollination has been estimated at over US $13 million annually in three provinces of Thailand (Petchmunee 2008).
6 The Issue of Ecosystem Disservices of Bats to Agricultural Production
Unfortunately, while the ecosystem services provided by bats are largely invisible, their disservices are obvious. In the Paleotropics, crop raiding by frugivorous pteropodids can cause substantial losses of commercial fruits (see Aziz et al., Chap. 12). For example, in Indian vineyards, Cynopterus sphinx damages up to 90 % of the crop along peripheries of plantations and may cause revenue losses of up to US $590 per ha and year (Srinivasulu and Srinivasulu 2002). In the Neotropics, sanguivorous vampire bats can cause substantial economic damage: Estimates for 1968 placed losses at $47.5 million USD for over 512,000 rabies-related cattle deaths in Latin America (Arellano-Sota 1988). Harassment by vampire bats can put cattle off their feed, resulting in annual weight losses estimated at roughly 40 kg/head and milk production loss of 261 L/head (Schmidt and Badger 1979). These estimates fail to take into account the effects of vampire bats on the medium and small domestic animals (e.g., chickens, pigs, goats) that provide critical sources of animal protein for millions of smallholder farmers across the region.
Not surprisingly, farmers with first-hand experiences of economic losses engendered by bats are more likely to have negative attitudes or report a willingness to destroy bat roosts (Reid 2013). Failure to explicitly address the negative impacts of some bat species likely reduces the efficacy of conservation messages; meanwhile, practical measures to reduce these disservices could benefit multiple bat species by reducing indiscriminate persecution. Different functional groups provide most of the ecosystem services (insectivores, nectarivores) and disservices (frugivores, sanguivores). However, local farmers may not distinguish between these groups. For example, farmers and agricultural technicians in Latin America often attempt to cull vampire bat populations by destroying bat roosts; unfortunately, the widespread belief that all bats are “vampiros” frequently results in the destruction of colonies of beneficial bat species (Mayen 2003; Aguiar et al. 2010). If local people perceive the ecosystem services of one bat group as offsetting the damages of another, then an ecosystem service approach could provide a framework for bat conservation more broadly. Unfortunately, the extent to which knowledge of ecosystem services changes attitudes toward bats in developing countries remains unknown.
7 Discussion
Our review suggests that in all biogeographic regions investigated, at least some bat species persist in and exploit agricultural areas. In many agricultural systems (e.g., tropical agroforestry or historical landscapes of Europe), bat assemblages maintain richness and may even exceed abundances observed in unmanaged areas. Nevertheless, agricultural intensification has a generally negative effect on bats and thus presumably on the ecosystem services they provide. Our analysis did not address differences between bat taxa in their sensitivity to habitat change and intensification. However, evidence from speciose assemblages suggests that forest-adapted insectivorous species are particularly sensitive to habitat conversion (Medellin et al. 2000; Faria and Baumgarten 2007; Williams-Guillén and Perfecto 2010), implying that in some regions, this valuable ecosystem service could be particularly vulnerable to loss in the face of habitat loss.
Although few investigations have considered the scale of intensification, limited information suggests that less managed systems embedded in regions dominated by intensive agriculture may show depauperate bat faunas (Numa et al. 2005; Faria et al. 2007). Declines in bat populations in agricultural regions are concerning not only from the point of view of biodiversity conservation but also regarding human well-being and food security , especially in many tropical areas where smallholder farming systems are dominant. Ongoing losses of these generalist vertebrate predators could have major impacts on insect pest limitation for a wide variety of staple and commodity crops. However, the smallholder farmers in developing nations who most depend on the ecosystem services provided by bats (due to limited access to manufactured inputs or cultivation of bat pollinated crops) may have highly negative attitudes toward these mammals as a result of visible damages caused to crops and livestock (López del Toro et al. 2009; Reid 2013), whereas beneficial impacts on crop yield productivity and the value of biodiversity (i.e., increased ecosystem resilience) are often unknown or unappreciated (Williams-Guillén, unpublished data). These results suggest a pressing need to reassess common approaches to conservation and agricultural management in the Anthropocene.
7.1 Sparing, Sharing, and the Devaluation of Manufactured Capital
Given the anticipated need to nearly double global food production in the twenty-first century, a vigorous debate has emerged with respect to the most viable path to increase production without degrading ecosystem services or reducing biodiversity: land sparing , which posits that increased intensification and yields will reduce pressure to convert non-agricultural lands, versus land sharing , in which agricultural areas are less intensively farmed in order to increase associated biodiversity and habitat permeability (Fischer et al. 2008). Given the vagility and critical role of bats in agricultural production, land sharing approaches might be preferable with respect to the provision of bat-dependent ecosystem services. Many sensitive bat ensembles and species (e.g., many forest-adapted and insectivorous species, e.g., from Phyllostomidae or Vespertilionidae) will require well-structured farmland, i.e., cultivated landscapes including patches of natural and seminatural features for their long-term existence. However, not only do many bat species thrive in diverse agricultural landscapes, but also their loss could affect the provision of pest suppression and pollination services and result in reduced crop productivity. Given the many disadvantages of chemical control of pests, managing agricultural landscapes to maximize the abundance and diversity of bats and other natural enemies must form a key aspect of sustainable agricultural production. However, the design and management of such systems to maximize bat diversity, activity, and ecosystem services is largely unknown, although European conservationists are at the forefront with their strategies to promote biodiversity-friendly farming.
Chemical and mechanical inputs are not the only tools of agricultural intensification. Within recent decades, genetic modification of crops (e.g., Bt corn and cotton) has become increasingly prevalent (James 2011). In the short term, adoption of such varieties does reduce the need to rely on bats and other predators for pest limitation (Lopez-Hoffman et al. 2014), resulting in a “devaluation” of the natural capital provided by bats, and undermines arguments for bat conservation that are based exclusively on provision of ecosystem services. However, as is the case with pesticides, insects are rapidly evolving resistance to Bt crops across the world, resulting in a rapid devaluation of manufactured capital (Lopez-Hoffman et al. 2014). While the value of bats’ natural capital may fluctuate, it likely devalues far less slowly: Bats and insects are engaged in an evolutionary arms race dating back millions of years (Conner and Corcoran 2012). Without bats to buffer the inevitable loss of efficacy of chemical inputs and GM crops, the technological advances that make agricultural intensification possible leave production vulnerable to potentially catastrophic failures to limit pest damage.
8 Research Priorities
8.1 Filling in Biogeographical Knowledge Gaps
Although the effects of habitat conversion and management have been well investigated in Europe and the Neotropics, the extent to which these processes may differ in other regions of the world remains unknown. We highlight a particular lack of knowledge from Africa and Asia; we did not find any studies from East Asia, although we suspect information exists in the Chinese language literature. Understanding the types and magnitudes of ecosystem services provided by bats in a variety of agricultural systems and regions is particularly important.
8.2 Linking Farm Management, Ecosystem Services, and Landscape-Level Processes
The effects of farm-level management on biodiversity and ecosystem services cannot be adequately considered without taking account of landscape -level processes (Tscharntke et al. 2005; Vickery and Arlettaz 2012). Nevertheless, the extent to which local- and landscape-level management interact to shape pest suppression or pollination services is largely uninvestigated. The effect of bats in limiting arthropod pests in agricultural areas is still poorly documented. However, the limited data that exist can demonstrate a vexing degree of divergence in results. For example, bats in Mexican shade coffee have substantial effects on herbivorous insects (Williams-Guillén et al. 2008), while bats in Costa Rican shade coffee had no significant effect on herbivores (Karp et al. 2013). In Indonesian cacao agroforestry systems, insectivorous bats strongly contribute to the suppression of many different pest insect groups and crop yield productivity across gradients of local shade-tree management and forest proximity within the agricultural landscape (Maas et al. 2013). In general, the study sites differ in landscape structure and land use, local farm history and management, habitat dynamics and conversion, intensity of farming practices, and vertebrate insectivore assemblage structure. Elucidating the factors of bat ecosystem service provision is key to managing agricultural areas to sustain bat populations and enhance food production (Maas et al. 2015).
8.3 Pest Suppression in the Face of Climate Change , Pesticides, and GM Crops
Not only will warming climates lead to shifts in the areas suitable for agricultural production, but it will also likely lead to range expansions of tropical pests, increases in pest numbers and damage, with a parallel risk of a drop in the efficacy of pest suppression by natural enemies that might be negatively affected by climate change (Thomson et al. 2010; Bebber et al. 2013). Such changes will make the ecosystem services provided by generalist predators like insectivorous bats more valuable than ever before. However, if agricultural adaptation to climate change relies on landscape-level intensification as a strategy, bats are likely to decline further, reducing their provision of pest suppression services. Despite the myriad negative effects of pesticides (i.e., affecting livelihoods, food security, environment, and health; reviewed by Yadav 2010), farmers across the world might turn to agrochemicals as a first response to increases in pest damage (Wilson and Tisdell 2001), with the Old World’s rapid development of more environmentally friendly farming practices appearing as an exception in this general move. As reviewed in this chapter, older pesticide classes such as organochlorines have particularly detrimental effects on bat populations. However, the degree to which newer pesticide classes affect bats is largely unknown. The neonicotinoids, once touted for their low toxicity, have now been linked to major declines in bees (Van der Sluijs et al. 2013) and more recently in several species of passerines as a result of insect resource depletion (Hallmann et al. 2014). The extent to which use of next-generation pesticides and GM crops is driving and interacting with bat declines and resultant increases in pest damage is a critical research area.
8.4 Quantifying Impact and Value Across Crops and Biomes
Additional valuation of bats’ ecosystem services could provide both guidance for bat management priorities in agricultural areas and compelling rationales for conservation. However, valuation efforts have focused almost exclusively on commodity crops quantified along the single dimension of monetary value. Most of the world’s smallholder farmers focus on staple crop cultivation and may not have the means to substitute the manufactured capital of pesticides and GM crops for bat predation. As Wanger et al. (2014) demonstrate, valuation based on dollars of damage prevented misses many of the criteria most important to subsistence farmers seeking food security . There is an urgent need to better understand the importance of bat ecosystem services across a variety of crop types, regions, and management approaches. Research also highlights the importance of better quantifying the fluctuations in bat service provision across years and seasons, in relation to population fluctuations, reproductive phenology, and agricultural management (Lopez-Hoffman et al. 2014; Wanger et al. 2014; Maas et al. 2015). This level of local, nuanced knowledge is key to managing pest suppression services in such a way that they are actively used as alternatives to agrochemical inputs and GM crops, and to contribute to more biodiversity-friendly and sustainable land-use practices (Tilman et al. 2002; Maas et al. 2015).
8.5 Changing Attitudes and Behaviors Toward Bats in the Developing World
Although the conservation of tropical biodiversity is highly beneficial to global society (Rands et al. 2010), ultimately it is the attitudes and beliefs of farmers and other rural populations that will determine its fate (Brechin et al. 2002; Tscharntke et al. 2012). Throughout the world, bats are subject to misconceptions and poor public perceptions (see Kingston and Barlow, this volume Chap. 17). However, exposure to environmental education can significantly decrease negative attitudes toward bats (López del Toro et al. 2009; Prokop et al. 2009; Reid 2013). These results suggest that reducing bat disservices, conducting environmental education , and building local valuation of beneficial bats could work in concert to improve conservation outcomes. As much as there is a critical need to manage agricultural landscapes to conserve bats, there is a parallel need to understand the local drivers of attitudes toward bats and to develop culturally appropriate, evidence-based interventions that encourage farmers to sustainably manage bat populations and other biodiversity associated with ecosystem services and ecosystem resilience.
References
Aguiar LMS, Brito D, Machado RB (2010) Do current vampire bat (Desmodus rotundus) population control practices pose a threat to Dekeysers nectar bats (Lonchophylla dekeyseri) long-term persistence in the Cerrado? Acta Chiropterol 12:275–282
Arellano-Sota C (1988) Vampire bat-transmitted rabies in cattle. Rev Infect Dis 10:707–709
Arlettaz R (1996) Feeding behaviour and foraging strategy of free-living mouse-eared bats, Myotis myotis and Myotis blythii. Anim Behav 51:1–11
Arlettaz R (1999) Habitat selection as a major resource partitioning mechanism between the two sympatric sibling bat species Myotis myotis and Myotis blythii. J Anim Ecol 68:460–471
Arlettaz R, Perrin N (1995) The trophic niches of sympatric sibling Myotis myotis and Myotis blythii: do mouse-eared bats select prey? Symp Zool Soc Lond vol 67, London: The Society 1960–1999, pp 361–376
Arlettaz R, Perrin N, Hausser J (1997) Trophic resource partitioning and competition between the two sibling bat species. J Anim Ecol 66:897–911
Arlettaz R, Christe P, Lugon A et al (2001) Food availability dictates the timing of parturition in insectivorous mouse-eared bats. Oikos 95:105–111
Avila-Cabadilla LD, Stoner KE, Henry M, Alvarez-Añorve MY (2009) Composition, structure and diversity of phyllostomid bat assemblages in different successional stages of a tropical dry forest. For Ecol Manage 1–11
Aziz SA, Olival KJ, Bumrungsri S, Richards GC, Racey PA (2016) The conflict between pteropodid bats and fruit growers: species, legislation and mitigation. In: Voight CC, Kingston T (eds) Bats in the Anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp. 377–420
Bayat S, Geiser F, Kristiansen P, Wilson SC (2014) Organic contaminants in bats: trends and new issues. Environ Int 63:40–52
Bebber DP, Ramotowski MAT, Gurr SJ (2013) Crop pests and pathogens move polewards in a warming world. Nat Clim Change 3:985–988
Bennett BS, Thies ML (2007) Organochlorine pesticide residues in guano of Brazilian free-tailed bats, Tadarida brasiliensis Saint-Hilaire, from east Texas. Bull Environ Contam Toxicol 78:191–194
Bohmann K, Monadjem A, Noer CL et al (2011) Molecular diet analysis of two African free-tailed bats (Molossidae) using high throughput sequencing. PLoS ONE 6:e21441
Bontadina F, Schofield H, Naef-Daenzer B (2002) Radio-tracking reveals that lesser horseshoe bats (Rhinolophus hipposideros) forage in woodland. J Zool 258:281–290
Boughey KL, Lake IR, Haysom KA, Dolman PM (2011) Improving the biodiversity benefits of hedgerows: how physical characteristics and the proximity of foraging habitat affect the use of linear features by bats. Biol Conserv 144:1790–1798
Boyles JG, Cryan PM, McCracken GF, Kunz TH (2011) Economic importance of bats in agriculture. Science 332:41–42
Boyles JG, Sole CL, Cryan PM, McCracken GF (2013) On estimating the economic value of insectivorous bats: prospects and priorities for biologists. In: Adams RA, Pedersen SC (eds) Bat evolution, ecology, and conservation. Springer, New York, pp 501–515
Brashares JS, Epps CW, Stoner CJ (2010) Ecological and conservation implications of mesopredator release. In: Terborgh J, Estes J (eds) Trophic cascades. Island Press
Braun de Torrez E (2014) Bats, insects and pecans: habitat use and ecosystem services of insectivorous bats in a pecan agroecosystem in central Texas. Boston University, Boston
Brechin SR, Wilshusen P, Fortwangler CL, West PC (2002) Beyond the square wheel: toward a more comprehensive understanding of biodiversity conservation as a social and political process. Soc Nat Resour 15:41–64
Bumrungsri S, Harbit A, Benzie C et al (2008a) The pollination ecology of two species of Parkia (Mimosaceae) in southern Thailand. J Trop Ecol 24:467–475
Bumrungsri S, Sripaoraya E, Chongsiri T et al (2008b) The pollination ecology of durian (Durio zibethinus, Bombacaceae) in southern Thailand. J Trop Ecol 25:85
Bumrungsri S, Lang D, Harrower C et al (2013) The dawn bat, Eonycteris spelaea Dobson (Chiroptera: Pteropodidae) feeds mainly on pollen of economically important food plants in Thailand. Acta Chiropterol 15:95–104
Castro-Luna AA, Galindo-González J (2012) Enriching agroecosystems with fruit-producing tree species favors the abundance and richness of frugivorous and nectarivorous bats in Veracruz, Mexico. Mamm Biol 77:32–40
Clare EL, Barber BR, Sweeney BW et al (2011) Eating local: influences of habitat on the diet of little brown bats (Myotis lucifugus). Mol Ecol 20:1772–1780
Clark DR (1988) How sensitive are bats to insecticides? Wildl Soc B 16:399–403
Clark DR (2001) DDT and the decline of free-tailed bats (Tadarida brasiliensis) at Carlsbad Cavern, New Mexico. Arch Environ Contam Toxicol 40:537–543
Clark DR, Prouty RM (1976) Organochlorine residues in three bat species from four localities in Maryland and West Virginia, 1973. Pestic Monit J 10:44–53
Clark DR, LaVal RK, Swineford DM (1978) Dieldrin-induced mortality in an endangered species, the gray bat (Myotis grisescens). Science 199:1357–1359
Clark DR, LaVal RK, Krynitsky AJ (1980) Dieldrin and heptachlor residues in dead gray bats, Franklin County, Missouri–1976 versus 1977. Pestic Monit J 13:137–140
Clawson RL, Clark DR (1989) Pesticide contamination of endangered gray bats and their food base in Boone County, Missouri, 1982. Bull Environ Contam Toxicol 42:431–437
Cleveland C, Betke M, Federico P, Frank J (2006) Economic value of the pest control service provided by Brazilian free-tailed bats in south-central Texas. Front Ecol Environ 4:238–243
Conner WE, Corcoran AJ (2012) Sound strategies: the 65-million-year-old battle between bats and insects. Annu Rev Entomol 57:21–39
Davy CM, Russo D, Fenton MB (2007) Use of native woodlands and traditional olive groves by foraging bats on a Mediterranean island: consequences for conservation. J Zool 273:397–405
de Jong J (1995) Habitat use and species richness of bats in a patchy landscape. Acta Theriol 40:237–248
Defries RS, Rudel T, Uriarte M, Hansen M (2010) Deforestation driven by urban population growth and agricultural trade in the twenty-first century. Nat Geosci 3:1–4
Dietz M, Pir JB, Hillen J (2013) Does the survival of greater horseshoe bats and Geoffroy’s bats in Western Europe depend on traditional cultural landscapes? Biodivers Conserv 23:3007–3025
Downs NC, Racey PA (2006) The use by bats of habitat features in mixed farmland in Scotland. Acta Chiropterol 8:169–185
Drescher C (2004) Radiotracking of Myotis myotis (Chiroptera, Vespertilionidae) in South Tyrol and implications for its conservation. Mammalia 68:387–395
Egger M, Schneider M, Davey Smith G (1998) Spurious precision? Meta-analysis of observational studies. BMJ 316:140–144
Ekman M, de Jong J (1996) Local patterns of distribution and resource utilization of four bat species (Myotis brandti, Eptesicus nilssoni, Plecotus auritus and Pipistrellus pipistrellus) in patchy and continuous environments. J Zool 238:571–580
Estrada A, Coates-Estrada R (2001) Bat species richness in live fences and in corridors of residual rain forest vegetation at Los Tuxtlas, Mexico. Ecography 24:94–102
Estrada A, Coates-Estrada R (2002) Bats in continuous forest, forest fragments and in an agricultural mosaic habitat-island at Los Tuxtlas, Mexico. Biol Conserv 103:237–245
Estrada A, Coates-Estrada R, Meritt D (1993) Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas, Mexico. Ecography 16:309–318
Estrada A, Jimenez C, Rivera A, Fuentes E (2004) General bat activity measured with an ultrasound detector in a fragmented tropical landscape in Los Tuxtlas, Mexico. Anim Biodivers Conserv 27:1–9
FAO (2011) The state of the world’s land and water resources for food and agriculture: managing systems at risk, pp 1–308
FAOSTAT (2011) Food and Agriculture Organization of the United Nations. faostat. fao3. org. Accessed 13 Aug 2013
Faria D (2006) Phyllostomid bats of a fragmented landscape in the north-eastern Atlantic forest, Brazil. J Trop Ecol 22:531–542
Faria D, Baumgarten J (2007) Shade cacao plantations (Theobroma cacao) and bat conservation in southern Bahia, Brazil. Biodivers Conserv 16:291–312
Faria D, Laps RR, Baumgarten J, Cetra M (2006) Bat and bird assemblages from forests and shade cacao plantations in two contrasting landscapes in the Atlantic forest of southern Bahia, Brazil. Biodivers Conserv 15:587–612
Faria D, Paciencia MLB, Dixo M et al (2007) Ferns, frogs, lizards, birds and bats in forest fragments and shade cacao plantations in two contrasting landscapes in the Atlantic forest, Brazil. Biodivers Conserv 16:2335–2357
Farrow LJ, Broders HG (2011) Loss of forest cover impacts the distribution of the forest-dwelling tri-colored bat (Perimyotis subflavus). Mamm Biol 76:172–179
Federico P, Hallam TG, McCracken GF et al (2008) Brazilian free-tailed bats as insect pest regulators in transgenic and conventional cotton crops. Ecol Appl 18:826–837
Fischer J, Brosi B, Daily GC et al (2008) Should agricultural policies encourage land sparing or wildlife-friendly farming? Front Ecol Environ 6:380–385
Fischer J, Stott J, Zerger A et al (2009) Reversing a tree regeneration crisis in an endangered ecoregion. Proc Natl Acad Sci USA 106:10386–10391
Fischer J, Zerger A, Gibbons P et al (2010) Tree decline and the future of Australian farmland biodiversity. Proc Natl Acad Sci USA 107:19597–19602
Flaquer C, Puig-Montserrat X, Burgas A, Russo D (2008) Habitat selection by Geoffroy’s bats (Myotis emarginatus) in a rural Mediterranean landscape: implications for conservation. Acta Chiropterol 10:61–67
Foley JA, DeFries R, Asner GP et al (2005) Global consequences of land use. Science 309:570–574
Frey-Ehrenbold A, Bontadina F, Arlettaz R, Obrist MK (2013) Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. J Appl Ecol 50:252–261
Fuentes-Montemayor E, Goulson D, Park KJ (2011) Pipistrelle bats and their prey do not benefit from four widely applied agri-environment management prescriptions. Biol Conserv 144:2233–2246
Fukuda D, Tisen OB, Momose K (2009) Bat diversity in the vegetation mosaic around a lowland dipterocarp forest of Borneo. Raffles Bull Zool 57:213–221
Fuller J, Norton LR, Feber RE et al (2005) Benefits of organic farming to biodiversity vary among taxa. Biol Lett 1:431–434
Furey NM, Mackie IJ, Racey PA (2010) Bat diversity in Vietnamese limestone karst areas and the implications of forest degradation. Biodivers Conserv 19:1821–1838
Gándara Fierro G, Correa Sandoval AN, Hernández Cienfuegos CA (2006) Valoración económica de los servicios ecológicos que prestan los murciélagos Tadarida brasiliensis como controladores de plagas en el norte de México, pp 1–18
García Estrada C, Damon A, Hernandez C et al (2006) Bat diversity in montane rainforest and shaded coffee under different management regimes in southeastern Chiapas, Mexico. Biol Conserv 132:351–361
García-Estrada C, Damon A, Sánchez Hernández C et al (2012) Diets of frugivorous bats in montane rain forest and coffee plantations in southeastern Chiapas, Mexico. Biotropica 44:394–401
García-Morales R, Badano EI, Moreno CE (2013) Response of Neotropical bat assemblages to human land use. Conserv Biol 27:1096–1106
Gehrt SD, Chelsvig JE (2003) Bat activity in an urban landscape: patterns at the landscape and microhabitat scale. Ecol Appl 13:939–950
Graf S (2010) Diversity and habitat use of understorey bats in forest and agroforestry systems at the Margin of Lore Lindu National Park (Central Sulawesi, Indonesia). University of Vienna, Vienna
Guillén A, Ibáñez C, Pérez JL et al (1994) Organochlorine residues in Spanish common pipistrelle bats (Pipistrellus pipistrellus). Bull Environ Contam Toxicol 52:231–237
Hallmann CA, Foppen RPB, van Turnhout CAM et al (2014) Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 511:341–343
Hanspach J, Fischer J, Ikin K et al (2012) Using trait-based filtering as a predictive framework for conservation: a case study of bats on farms in southeastern Australia. J Appl Ecol 49:842–850
Harvey CA, González Villalobos JA (2007) Agroforestry systems conserve species-rich but modified assemblages of tropical birds and bats. Biodivers Conserv 16:2257–2292
Harvey CA, Medina A, Sánchez DM et al (2006) Patterns of animal diversity in different forms of tree cover in agricultural landscapes. Ecol Appl 16:1986–1999
Henderson LE, Broders HG (2008) Movements and resource selection of the northern long-eared Myotis (Myotis septentrionalis) in a forest–agriculture landscape. J Mammal 89:952–963
Hopkins J, Maxted N (2011) Crop wild relatives: plant conservation for food security, pp 1–58
James C (2011) Global status of commercialized biotech/GM crops: 2011. International service for the acquisition of agri-biotech applications (ISAAA), Ithaca
Jones G, Morton M (1992) Radio-tracking studies on habitat use by greater horseshoe bats (Rhinolophus ferrumequinum). In: 4th European conference on wildlife telemetry: Remote monitoring and tracking of animals, pp 521–537
Jones G, Jacobs DS, Kunz TH et al (2009) Carpe noctem: the importance of bats as bioindicators. Endanger Species Res 8:93–115
Kalka MB, Smith AR, Kalko EKV (2008) Bats limit arthropods and herbivory in a tropical forest. Science 320:71
Kannan K, Yun SH, Rudd RJ, Behr M (2010) High concentrations of persistent organic pollutants including PCBs, DDT, PBDEs and PFOS in little brown bats with white-nose syndrome in New York, USA. Chemosphere 80:613–618
Karp DS, Mendenhall CD, Sandí RF et al (2013) Forest bolsters bird abundance, pest control, and coffee yield. Ecol Lett 16:1339–1347
Kingston T (2016) Cute, Creepy, or Crispy—How values, attitudes, and norms shape human behavior toward bats. In: Voight CC, Kingston T (eds) Bats in the Anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp. 571–588
Kunz TH, Anthony EL, Rumage WT III (1977) Mortality of little brown bats following multiple pesticide applications. J Wildl Manage 476–483
Kunz TH, Whitaker JO Jr, Wadanoli MD (1995) Dietary energetics of the insectivorous Mexican free-tailed bat (Tadarida brasiliensis) during pregnancy and lactation. Oecologia 101:407–415
Kunz TH, Braun de Torrez E, Bauer D et al (2011) Ecosystem services provided by bats. Ann NY Acad Sci 1223:1–38
Kurta A, Bell GP, Nagy KA, Kunz TH (1989) Energetics of pregnancy and lactation in free ranging little brown bats (Myotis lucifugus). Physiol Zool 62:804–818
Lambin EF, Meyfroidt P (2011) Global land use change, economic globalization, and the looming land scarcity. Proc Natl Acad Sci USA 108:3465–3472
Laurance WF (2008) Theory meets reality: how habitat fragmentation research has transcended island biogeographic theory. Biol Conserv 141:1731–1744
Law et al. (2016) Insectivorous bats and silviculture: balancing timber production and bat conservation. In: Voight CC, Kingston T (eds) Bats in the Anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp. 105–141
Le Bellec F, Vaillant F, Imbert E (2006) Pitahaya (Hylocereus spp.): a new fruit crop, a market with a future. Fruits 61:237–250
Lee Y, McCracken G (2005) Dietary variation of Brazilian free-tailed bats links to migratory populations of pest insects. J Mammal 81:67–76
Leelapaibul W, Bumrungsri S, Pattanawiboon A (2005) Diet of wrinkle-lipped free-tailed bat (Tadarida plicata Buchannan, 1800) in central Thailand: insectivorous bats potentially act as biological pest control agents. Acta Chiropterol 7:111–119
Lentini PE, Gibbons P, Fischer J et al (2012) Bats in a farming landscape benefit from linear remnants and unimproved pastures. PLoS ONE 7:e48201
Lesiński G, Durka A, Rembiałkowska E (2013) Flight activity of serotine Eptesicus serotinus on organic and conventional farms in central Poland. Zool Ecol 23:93–96
Lisón F, Calvo JF (2011) The significance of water infrastructures for the conservation of bats in a semiarid Mediterranean landscape. Anim Conserv 14:533–541
Lisón F, Calvo JF (2013) Ecological niche modeling of three pipistrelle bat species in semiarid Mediterranean landscapes. Acta Oecol 47:68–73
Loos J, Abson DJ, Chappell MJ et al (2014) Putting meaning back into sustainable intensification. Front Ecol Environ 12:356–361
López del Toro P, Andresen E, Barraza L, Estrada A (2009) Attitudes and knowledge of shade-coffee farmers towards vertebrates and their ecological functions. Trop Conserv Sci 2:299–318
Lopez-Hoffman L, Wiederholt R, Sansone C et al (2014) Market forces and technological substitutes cause fluctuations in the value of bat pest-control services for cotton. PLoS ONE 9:e87912
Lumsden LF, Bennett AF (2005) Scattered trees in rural landscapes: foraging habitat for insectivorous bats in south-eastern Australia. Biol Conserv 122:205–222
Lumsden LF, Bennett AF, Silins JE (2002) Location of roosts of the lesser long-eared bat Nyctophilus geoffroyi and Gould’s wattled bat Chalinolobus gouldii in a fragmented landscape in south-eastern Australia. Biol Conserv 106:237–249
Lundy M, Montgomery I (2010) Summer habitat associations of bats between riparian landscapes and within riparian areas. Eur J Wildl Res 56:385–394
Maas B, Clough Y, Tscharntke T (2013) Bats and birds increase crop yield in tropical agroforestry landscapes. Ecol Lett 16:1480–1487
Maas B, Karp DS, Bumrungsri S, et al., (2015) Bird and bat predation services in tropical forests and agroforestry landscapes. Biol Rev Jul 23. doi:10.1111/brv.12211
Mayen F (2003) Haematophagous bats in Brazil, their role in rabies transmission, impact on public health, livestock industry and alternatives to an indiscriminate reduction of bat population. J Vet Med B Infect Dis Vet Public Health 50:469–472
McCracken GF, Westbrook JK, Brown VA et al (2012) Bats track and exploit changes in insect pest populations. PLoS ONE 7:e43839
McShane TO, Hirsch PD, Trung TC et al (2011) Hard choices: making trade-offs between biodiversity conservation and human well-being. Biol Conserv 144:966–972
Medellin R, Equihua M, Amin M (2000) Bat diversity and abundance as indicators of disturbance in Neotropical rainforests. Conserv Biol 14:1666–1675
Medina A, Harvey C, Merlo D et al (2007) Bat diversity and movement in an agricultural landscape in Matiguas, Nicaragua. Biotropica 39:120–128
Mendenhall CD, Karp DS, Meyer CFJ et al (2014) Predicting biodiversity change and averting collapse in agricultural landscapes. Nature 509:213–217
Meyer C et al. (2016) Responses of tropical bats to habitat fragmentation, logging and deforestation. In: Voight CC, Kingston T (eds) Bats in the Anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp. 63–94
Mildenstein TL, Stier SC, Nuevo-Diego CE (2005) Habitat selection of endangered and endemic large flying-foxes in Subic Bay, Philippines. Biol Conserv 126:93–102
Miura T, Koyama T, Nakamura I (1978) Mercury content in museum and recent specimens of chiroptera in Japan. Bull Environ Contam Toxicol 20:696–701
Morrison EB, Lindell CA (2012) Birds and bats reduce insect biomass and leaf damage in tropical forest restoration sites. Ecol Appl 22:1526–1534
Nelson GC, Rosegrant MW, Koo J et al (2009) Climate change: impact on agriculture and costs of adaptation, pp 1–30
Noer CL, Dabelsteen T, Bohmann K, Monadjem A (2012) Molossid bats in an African agro-ecosystem select sugarcane fields as foraging habitat. Afr Zoo 47:1–11
Numa C, Verdú JR, Sánchez-Palomino P (2005) Phyllostomid bat diversity in a variegated coffee landscape. Biol Conserv 122:151–158
O’Shea TJ, Johnston JJ (2009) Environmental contaminants and bats: investigating exposure and effects. In: Kunz TH, Parsons S (eds) Ecological and behavioral methods for the study of bats. The Johns Hopkins University Press, Baltimore, pp 500–528
Obrist MK, Rathey E, Bontadina F et al (2011) Response of bat species to sylvo-pastoral abandonment. For Ecol Manage 261:789–798
O’Shea TJ, Everette AL, Ellison LE (2001) Cyclodiene insecticide, DDE, DDT, arsenic, and mercury contamination of big brown bats (Eptesicus fuscus) foraging at a Colorado Superfund site. Arch Environ Contam Toxicol 40:112–120
Perfecto I, Vandermeer J (2010) The agroecological matrix as alternative to the land-sparing/agriculture intensification model. Proc Natl Acad Sci USA 107:5786–5791
Petchmunee K (2008) Economic valuation and learning process construction: a case study of the cave Nectarivorous bat (Eonycteris spelaea Dobson), pp 1–278
Pineda E, Moreno C, Escobar F, Halffter G (2005) Frog, bat, and dung beetle diversity in the cloud forest and coffee agroecosystems of Veracruz, Mexico. Conserv Biol 19:400–410
Pocock MJO, Jennings N (2007) Testing biotic indicator taxa: the sensitivity of insectivorous mammals and their prey to the intensification of lowland agriculture. J Appl Ecol 45:151–160
Power AG (2010) Ecosystem services and agriculture: tradeoffs and synergies. Philos Trans R Soc B 365:2959–2971
Prokop P, Fančovičová J, Kubiatko M (2009) Vampires are still alive: Slovakian students’ attitudes toward bats. Anthrozoos 22:19–30
Quarles W (2013) Bats, pesticides and white nose syndrome. IPM Pract 33:1–20
Rainho A (2007) Summer foraging habitats of bats in a Mediterranean region of the Iberian Peninsula. Acta Chiropterol 9:171–181
Rambaldini DA, Brigham RM (2011) Pallid bat (Antrozous pallidus) foraging over native and vineyard habitats in British Columbia, Canada. Can J Zool 89:816–822
Randrianandrianina F, Andriafidison D, Kofoky AF et al (2006) Habitat use and conservation of bats in rainforest and adjacent human-modified habitats in eastern Madagascar. Acta Chiropterol 8:429–437
Redfern SK, Azzu N, Binamira JS (2012) Rice in Southeast Asia: facing risks and vulnerabilities to respond to climate change. In: Building resilience for adaptation to climate change in the agriculture sector: proceedings of a Joint FAO/OECD Workshop. FAO, Rome, pp 295–314
Reid JL (2013) Restoring birds, bats, and dispersal mutualisms in a tropical agricultural landscape. University of California, Santa Cruz
Ricketts TH (2001) The matrix matters: effective isolation in fragmented landscapes. Am Nat 158:87–99
Robertson G, Swinton S (2005) Reconciling agricultural productivity and environmental integrity: a grand challenge for agriculture. Front Ecol Environ 3:38–46
Russ J, Montgomery W (2002) Habitat associations of bats in Northern Ireland: implications for conservation. Biol Conserv 107:49–58
Russo D, Jones G (2003) Use of foraging habitats by bats in a Mediterranean area determined by acoustic surveys: conservation implications. Ecography 26:197–209
Russo D, Jones G, Migliozzi A (2002) Habitat selection by the Mediterranean horseshoe bat, Rhinolophus euryale (Chiroptera: Rhinolophidae) in a rural area of southern Italy and implications for conservation. Biol Conserv 107:71–81
Saldaña Vázquez RA, Castro-Luna AA, Sandoval Ruiz CA et al (2013) Population composition and ectoparasite prevalence on bats (Sturnira ludovici; Phyllostomidae) in forest fragments and coffee plantations of central Veracruz, Mexico. Biotropica 45:351–356
Sasse DB (2005) Pesticide residues in guano of gray bats. J Ark Acad Sci 59:214–217
Schmidt KM, Badger DD (1979) Some social and economic aspects in controlling vampire bats. Proc Okla Acad Sci 59:112–114
Schmidt A, Brack V Jr, Rommé R et al (2000) Bioaccumulation of pesticides in bats from Missouri. In: Johnston JJ (ed) ACS symposium series. American Chemical Society, Washington, DC, pp 8–21
Sedlock JL, Weyandt SE, Cororan L et al (2008) Bat diversity in tropical forest and agro-pastoral habitats within a protected area in the Philippines. Acta Chiropterol 10:349–358
Senthilkumar K, Kannan K, Subramanian A, Tanabe S (2001) Accumulation of organochlorine pesticides and polychlorinated biphenyls in sediments, aquatic organisms, birds, bird eggs and bat collected from south India. Environ Sci Pollut Res 8:35–47
Sirami C, Jacobs DS, Cumming GS (2013) Artificial wetlands and surrounding habitats provide important foraging habitat for bats in agricultural landscapes in the Western Cape, South Africa. Biol Conserv 164:30–38
Sosa VJ, Hernández-Salazar E, Hernández-Conrique D, Castro-Luna AA (2008) Murciélagos. In: Manson RH, Hernández-Ortiz V, Gallina S, Mehltreter K (eds) Agroecosistemas Cafetaleros de Veracruz: Biodiverdiad, Manejo, y Conservación. INECOL & INE-SEMARNAT, México D.F, pp 181–192
Srinivasulu C, Srinivasulu B (2002) Greater short-nosed fruit bat (Cynopterus sphinx) foraging and damage in vineyards in India. Acta Chiropterol 4:167–171
Stahlschmidt P, Brühl CA (2012) Bats at risk? Bat activity and insecticide residue analysis of food items in an apple orchard. Environ Toxicol Chem 31:1556–1563
Stahlschmidt P, Pätzold A, Ressl L et al (2012) Constructed wetlands support bats in agricultural landscapes. Basic Appl Ecol 13:196–203
Stechert C, Kolb M, Bahadir M et al (2014) Insecticide residues in bats along a land use-gradient dominated by cotton cultivation in northern Benin. Environ Sci Pollut Res, West Africa. doi:10.1007/s11356-014-2817-8
Storm JJ, Whitaker JO Jr (2008) Prey selection of big brown bats (Eptesicus fuscus) during an emergence of 17-year cicadas (Magicicada spp.). Am Midl Nat 160:350–357
Swanepoel RE, Racey PA, Shore RF, Speakman JR (1998) Energetic effects of sublethal exposure to lindane on pipistrelle bats (Pipistrellus pipistrellus). Environ Pollut 104:169–177
Taylor PJ, Nemudivhiso P, Mphethe V et al (2012) Bats as biocontrol agents in macadamia orchards, Levubu Valley, Limpopo Province: results of a pilot project and future prospects. South Afr Macademia Grow Assoc Yearb 20:41–51
Taylor PJ, Bohmann K, Steyn JN et al (2013a) Bats eat pest green vegetable stink bugs (Nezara viridula): diet analyses of seven insectivorous species of bats roosting and foraging in macadamia orchards. South Afr Macademia Grow Assoc Yearb 21:37–43
Taylor PJ, Monadjem A, Steyn JN (2013b) Seasonal patterns of habitat use by insectivorous bats in a subtropical African agro-ecosystem dominated by macadamia orchards. Afr J Ecol 51:552–561
Thies ML, Thies KM (1997) Organochlorine residues in bats from Eckert James River Cave, Texas. Bull Environ Contam Toxicol 58:673–680
Thomson LJ, Macfadyen S, Hoffmann AA (2010) Predicting the effects of climate change on natural enemies of agricultural pests. Biol Control 52:296–306
Tilman D (1999) Global environmental impacts of agricultural expansion: the need for sustainable and efficient practices. Proc Natl Acad Sci USA 96:5995–6000
Tilman D, Fargione J, Wolff B et al (2001) Forecasting agriculturally driven global environmental change. Science 292:281–284
Tilman D, Cassman KG, Matson PA et al (2002) Agricultural sustainability and intensive production practices. Nature 418:671–677
Tscharntke T, Klein AM, Kruess A et al (2005) Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecol Lett 8:857–874
Tscharntke T, Clough Y, Wanger TC et al (2012) Global food security, biodiversity conservation and the future of agricultural intensification. Biol Conserv 151:53–59
Tuttle SR, Chambers CL, Theimer TC (2006) Potential effects of livestock water-trough modifications on bats in northern Arizona. Wildl Soc B 34:602–608
Valiente-Banuet A, Santos Gally R, Arizmendi MC, Casas A (2007) Pollination biology of the hemiepiphytic cactus Hylocereus undatus in the Tehuacán Valley, Mexico. J Arid Environ 68:1–8
Van der Sluijs JP, Simon-Delso N, Goulson D et al (2013) Neonicotinoids, bee disorders and the sustainability of pollinator services. Curr Opin Environ Sustain 5:293–305
Van Weerd M, Snelder DJ (2008) Human-altered tree-based habitats and their value in conserving bird and bat diversity in northeast Luzon, Philippines. In: Snelder DJ, Lasco RD (eds) Smallholder tree growing for rural development and environmental services. Springer, Dordrecht, pp 347–377
Vandermeer J, Perfecto I (2007) The agricultural matrix and a future paradigm for conservation. Conserv Biol 21:274–277
Vargas Espinoza A, Aguirre LF, Galarza MI, Gareca E (2008) Ensamble de murciélagos en sitios con diferente grado de perturbación en un bosque montano del Parque Nacional Carrasco, Bolivia. Mastozool Neotrop 15:297–308
Verboom B, Huitema H (1997) The importance of linear landscape elements for the pipistrelle Pipistrellus pipistrellus and the serotine bat Eptesicus serotinus. Landsc Ecol 12:117–125
Vickery J, Arlettaz R (2012) The importance of habitat heterogeneity at multiple scales for birds in European agricultural landscapes. In: FR J (ed) Birds and habitat: relationships in changing landscapes. Cambridge University Press, Cambridge, pp 177–204
Walsh AL, Harris S (1996a) Factors determining the abundance of vespertilionid bats in Britain: geographical, land class and local habitat relationships. J Appl Ecol 33:519–529
Walsh AL, Harris S (1996b) Foraging habitat preferences of vespertilionid bats in Britain. J Appl Ecol 33:508–518
Wanger TC, Darras K, Bumrungsri S et al (2014) Bat pest control contributes to food security in Thailand. Biol Conserv 171:220–223
Weiss J, Nerd A, Mizrahi Y (1994) Flowering behavior and pollination requirements in climbing cacti with fruit crop potential. HortScience 29:1487–14982
Whitaker JO Jr (1995) Food of the big brown bat Eptesicus fuscus from materinity colonies in Indiana and Illinois. Am Midl Nat 134:346–360
White DH, Krynitsky AJ (1986) Wildlife in some areas of New Mexico and Texas accumulate elevated DDE residues, 1983. Arch Environ Contam Toxicol 15:149–157
Wickramasinghe L, Harris S, Jones G, Vaughn N (2003) Bat activity and species richness on organic and conventional farms: impact of agricultural intensification. J Appl Ecol 40:984–993
Wickramasinghe L, Harris S, Jones G, Vaughn Jenings N (2004) Abundance and species richness of nocturnal insects on organic and conventional farms: effects of agricultural intensification on bat foraging. Conserv Biol 18:1283–1292
Williams-Guillén K, Perfecto I (2010) Effects of agricultural intensification on the assemblage of leaf-nosed bats (Phyllostomidae) in a coffee landscape in Chiapas, Mexico. Biotropica 42:605–613
Williams-Guillén K, Perfecto I (2011) Ensemble composition and activity levels of insectivorous bats in response to management intensification in coffee agroforestry systems. PLoS ONE 6:e16502
Williams-Guillén K, Perfecto I, Vandermeer J (2008) Bats limit insects in a Neotropical agroforestry system. Science 320:70
Wilson C, Tisdell C (2001) Why farmers continue to use pesticides despite environmental, health and sustainability costs. Ecol Econ 39:449–462
Yadav SK (2010) Pesticide applications: threat to ecosystems. J Hum Ecol 32:37–45
Acknowledgments
We would like to thank the researchers who made their results available to us, particularly those who shared work in preparation or press. We are very grateful to Olivier Roth who carried out a thorough literature search on European farmland bats and Jean-Yves Humbert for feedback on the meta-analysis approach. Justin Boyle, Pia Lentin, Tigga Kingston, and an anonymous reviewer provided detailed feedback that greatly improved this manuscript. Finally, we thank the editors of the current volume for invitation to review this topic and for their patience.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2016 The Author(s)
About this chapter
Cite this chapter
Williams-Guillén, K., Olimpi, E., Maas, B., Taylor, P.J., Arlettaz, R. (2016). Bats in the Anthropogenic Matrix: Challenges and Opportunities for the Conservation of Chiroptera and Their Ecosystem Services in Agricultural Landscapes. In: Voigt, C., Kingston, T. (eds) Bats in the Anthropocene: Conservation of Bats in a Changing World. Springer, Cham. https://doi.org/10.1007/978-3-319-25220-9_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-25220-9_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-25218-6
Online ISBN: 978-3-319-25220-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)