
Abstract
Green roof retrofits offer a promising avenue to increase greenspace and thus biodiversity in the city. The successful colonisation and establishment of plants and animals on green roofs is limited by the location and context of the green roof. Here we use a before, after, control, impact (BACI) design to monitor the colonisation of a new retrofit roof in Melbourne CBD. We find that colonisation for some taxa occurs rapidly, with honeybees (Apis mellifera) arriving four days after flowers had been planted. Other insect taxa, such as native bees, did not colonise the impact roof but were present on the green roof reference site, which was lower in height and planted with native plants. Invertebrate abundance increased for all sites after the retrofit was built, however this sampling period was in late spring when more insects are expected to be active. Bird richness and abundance didn’t change in response to roof greening, but invertebrate richness was significantly higher on reference sites and increased after the green roof retrofit was built for the impact site. There was an apparent ‘spill over’ effect onto the nearby green roof control, which also increased in invertebrate diversity. Overall, invertebrate composition across roof types was driven by floral density. Floral density and richness increased on the impact roof as vegetation matured, subsequently correlating with higher abundance and richness of invertebrates. We use these findings to discuss colonisation of green roofs and argue for strategic placement of new green roof retrofits to maximise their biodiversity potential.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Urban green spaces form increasingly important habitats for animals in an increasingly urbanised world (Prendergast et. al 2022a). Yet, in densely urbanised cities, there are limited options for increasing urban greenspace due to the contested nature of urban land use (Beninde et al. 2015). Incorporating vegetation into buildings through green roofs and green walls can somewhat alleviate these conflicts and allow for habitat provision even in densely urban areas by promoting multifunctional use of urban space (Joshi et al. 2020). Green roof retrofits allow for greening of existing buildings and strategic placement of new greenspaces in places that would be most beneficial (e.g., increasing habitat in already dense urban spaces, Williams et al. 2010), or most effective (e.g., acting as corridors for wildlife, Mayrand and Clergeau 2018).
While a promising approach to increasing greenspace in dense urban areas, the process of adding vegetation to existing roofs comes with several challenges. Structural and weight-loading limitations (Cascone et al. 2018) often mean that retrofits must be shallower, or limited in scope (e.g., extensive rooftops, or planter boxes installed on top of the roof) compared those designed into new buildings. Shading from other buildings can also restrict the success of installed plantings (Wilkinson and Reed 2009). High initial installation costs may be a disincentive to uptake, particularly in buildings with high tenancy turnover, because the recuperation of costs via the economic benefits of green roofs (e.g., increased longevity of waterproof membranes and thermal insulation reducing heating and cooling costs) can take several years to manifest (Manso et al. 2021). Green roofs are also more cost effective at a city scale (Manso et al. 2021), particularly for intensive designs with deeper soil and greater biodiversity benefits (Köhler and Ksiazek-Mikenas 2018), emphasising the importance of government incentives for promoting uptake (Burszta-Adamiak and Fiałkiewicz 2019; Williams et al. 2021). These physical and fiscal limitations can restrict the number of roofs suitable for green roof retrofits (Wilkinson and Reed 2009), as well as the quality of habitat available (Mayrand and Clergeau 2018).
Similar to ground-level greenspaces, the effectiveness of green roofs as habitat is limited by green roof design and location (Gedge and Kadas 2005; Dunnett 2006; Mayrand and Clergeau 2018). Increased exposure to wind, rain, and radiation due to high elevation, and relatively shallow soils (Oberndorfer et al. 2007) will reduce the survival of plants (Lundholm 2006) and the quality of habitat (Mayrand and Clergeau 2018). The vegetation available on green roofs, in turn, influences the fauna species that can successfully utilise roof habitats (Lundholm 2006; Hunter and Hunter 2008; Lundholm and Richardson 2010; Faeth et al. 2011; Nipperess et al. 2012). For example, host-specific invertebrates will only colonise a roof if their host plant, (or host prey) species is present (Southwood 1988; Tonietto et al. 2011; Prendergastt et al. 2022b). In general, species richness of invertebrates is higher on roofs with high plant diversity (Cook-Patton and Bauerle 2012; Braaker et al. 2014), high plant biomass (Madre et al. 2013), greater structural complexity (Lundholm and Richardson 2010; Madre et al. 2013), or large roof area (Madre et al. 2013), as diversification in available food sources increases the likelihood of providing for the resource needs of different invertebrate taxa. Similarly, soil invertebrates are richer where there is a deeper soil layer (Brenneisen 2006; Schrader and Böning 2006), and variation in substrate influences species composition (Jones 2002).
As in other areas of greenspace design, there is increasing interest in the use of locally indigenous plant species to provide better habitats on green roofs (Butler et al. 2012). The suitability of native plants is a contentious issue considering the drastically different conditions on top of roofs which often do not match regional conditions and may be unsuitable for local biota (reviewed in Li & Yeung, and Butler et al. 2012). As a result, green roofs can harbour distinct assemblages of invertebrates suited to more xeric environments (Madre et al. 2013). However, in Australia, many native plant species are suited to these kinds of dry climates and have been successfully planted on green roofs (reviewed in Williams et al. 2021). Studies of faunal use of green roofs in Australia are scarce with only two published examples (Berthon et al. 2015; Dromgold et al. 2020). Berthon et al. (2015) investigated green roofs in Sydney and found that roof area, connectivity and building height were driving changes in invertebrate community richness and composition but did not study vegetation characteristics. Dromgold et al. (2020) studied green roofs in Melbourne and found that invertebrate assemblages were similar between native and non-native planted rooftops. Instead, other factors such as age and roof height were more important in driving community differences. There is a similar lack of information on the biodiversity benefits of green roofs globally (Williams et al. 2014).
Regardless of the quality of habitat provided, the characteristics of the surrounding landscape and the dispersal ability of organisms affects the ability of species to access and successfully utilise roof resources (Tonietto et al. 2011; Ksiazek et al. 2012; Braaker et al. 2014). Green roofs are isolated in three dimensions and act like ‘islands in the sky’ (Berthon 2015; Blank et al. 2017), and this isolation affects the ability of invertebrates to colonise and form viable populations. For example, invertebrate richness and composition is negatively correlated with increased building height (MacIvor 2016) and proximity to surrounding habitat patches (Madre et al. 2013; Braaker et al. 2014). Importantly, the impact of height and landscape context is dependent on the mobility of the species, with low mobility species being less likely to appear on rooftops (Madre et al. 2013; Braaker et al. 2014; Dromgold et al. 2020). However, Berthon (2015) found that green roofs contained more low mobility, and immature individuals than bare roofs in Sydney, suggesting that green roofs could sustain viable populations of reproducing individuals.
The total faunal biodiversity on a rooftop is therefore the result of two balancing processes: immigration of individuals (affected by isolation) and retention of individuals (affected by resource provision). Biodiversity tends to be higher on older rooftops (Schrader and Böning 2006; Wang et al. 2017; Dromgold et al. 2020), as a result of continuous recruitment of both transient and resident individuals. Increasing maturity of vegetation over time provides additional resources that may attract more individuals and allow for larger populations (Schindler et al. 2011; Ksiazek-Mikenas et al. 2018). However, the trajectory of colonisation for every green roof is a highly stochastic process and varies depending on landscape context and vegetation changes on the roof (Ksiazek-Mikenas et al. 2018).
There are four main pathways for invertebrates to colonise rooftops (MacIvor and Ksiazek 2015): i) Active movement of invertebrates onto the roof by flight or crawling; ii) incidental movement by invertebrates onto the roof through air currents; iii) accidental human-mediated transport to the roof through planting; or iv) purposeful translocation of species onto rooftops. Species with high mobility and active search behaviours may be recruited to the rooftop of their own accord, potentially attracted to the resources on the rooftop. Alternatively, individuals may arrive spontaneously to the rooftop through undirected wind currents produced by city structures (Joimel et al. 2018). These wind tunnels may have the capacity to move invertebrates up to very tall rooftops that they might otherwise have not actively flown to. Invertebrates might also be introduced to the roof through human-mediated processes either during the construction from transport with plants and soil, or purposeful translocation. For example, McKinney et al. (2019) found that snails were likely being transported onto green roofs during installation and are commonly transported this way throughout the horticultural industry. Similarly, ladybeetle larvae have been translocated to a rooftop garden in Sydney to combat an aphid problem (Pers comm. Junglefy). Outside rooftop garden contexts, there is a growing industry in selling beneficial insects for the purpose of pest control, but most companies (e.g. https://www.arbico-organics.com/) sell live adults, which have limited benefits (e.g. they fly away) (Skwarecki 2017; Rondoni et al. 2021) and are usually exotic to the area to which they are being introduced, thus posing their own biocontrol risks from non-target effects (Rondoni et al. 2021).
The relative importance of these movement pathways is uncertain, and colonisation of rooftops has rarely been monitored. Often rooftops are seen as ‘stepping stones’ that may allow for movement of invertebrates throughout the landscape, but the connectivity of rooftops to each other and to ground environments is uncertain (Mayrand and Clergeau 2018). High similarity between roof and nearby ground sites has suggested the ability of invertebrates to move vertically onto rooftops (Braaker et al. 2014; Dromgold et al. 2020), but limitations in mobility may restrict how high invertebrates can go (Madre et al. 2013; Berthon et al. 2015; MacIvor 2016). For example, MacIvor (2016) found decreased abundance and lower breeding success of solitary bees on rooftops higher than five stories.
Here, we provide a case study for the colonisation of a newly installed rooftop retrofit in Melbourne Australia. Using observational surveys, we monitored the biodiversity (including invertebrates, birds, and plants), of a carpark roof before and after construction of a green roof retrofit to build a time series of colonisation of fauna and patterns of plant use on the rooftop in relation to changes in floral diversity. We also monitor two nearby rooftops and a ground site to compare the relative change in biodiversity before and after the roof is built to explore the impact of addition of elevated habitat in the landscape. Finally, we have monitored a large, long established green roof planted with native plants and connected to surrounding landscapes as a ‘reference’ site that represents a ‘best case scenario’ of green roof design. We discuss our findings regarding the influence of green roof design on biodiversity, and comment on which plants are being preferentially utilised by invertebrates, particularly native pollinators.
Methods
Study design and study sites
We monitored the impact of installation of a single green roof retrofit on top of an 8-story carpark (hereafter, the “impact” roof). The impact roof was designed as a community garden and contains a mix of productive plantings and flowers, including some Australian native plant species. A soft launch of the roof was announced in December 2021, and the roof will be open to the public after construction has been completed in 2023.
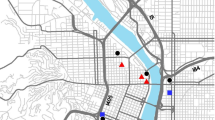
We used a before, after, control, impact study design (BACI, Smith et al. 1993) to assess biodiversity differences between rooftops over time. We included two nearby rooftops (<160 m distance, as the bird flies) as retrofit controls and to monitor the potential movement of invertebrates between rooftops: a residential green roof on an adjacent apartment complex (hereafter, ‘green roof control’), and a bare rooftop on a commercial building (hereafter, ‘bare roof control’). These rooftops were one story lower than the retrofit roof, on the 7th floor of their respective buildings. A nearby ground site, Docklands Park, was included as a ground reference site (~600 m from the impact roof). Since the two adjacent rooftops were much smaller than the proposed green roof development, a larger green roof planted with native plant species and well connected to the surrounding landscape (i.e., adjacent to the large parklands in the Treasury and Fitzroy Gardens) was chosen to be monitored as a ‘best case’ scenario green roof reference site. The green roof reference was much further from the installation roof than other sites (~2.3 km, Fig. 1) and was only one story in height. While the control rooftops are physically closer than the green roof reference, the 3D topographical complexity of urban areas means that animals may not easily be able to traverse between buildings (Kirk et al. 2023), and we therefore consider these to be distinct sampling units. All study sites are located within or adjacent to the highly urbanised central business district of the City of Melbourne, Australia. Further details of the sites are provided in Table 1. Building heights were recorded as the number of stories, and the Australian building code standards were used to estimate roof height in metres, as approximately 3 m per story (Australian Building Codes Board 1996).

Location of field sites within the City of Melbourne. The inset shows the location of the city of Melbourne within Australia
All sites were monitored for insects and bird activity, and plant phenology on two to four occasions between January – March 2020, before the impact green roof was installed. The green roof was installed in June 2021. The impact roof was sampled once immediately (4 days) after plants were installed; however, COVID-19 lockdown restrictions prevented sampling of other sites at this time. After restrictions eased, all control and ground sites were monitored on three occasions in October – November 2021, approximately three months after initial installation of plants on the impact site. Unfortunately, access to the green roof reference was unable to be organised during this period due to COVID-19 restrictions, and the site was not sampled post-construction. There are therefore no ‘after’ surveys for the green roof reference. Overall, COVID-19 restrictions made access to rooftops particularly difficult and delayed construction such that the bare roof control site had undergone renovations and lease changes between the before and after sampling periods. Sections of the impact roof and an adjacent carpark roof was still under construction during most of the surveys after plants had been installed, but these works were all located several metres from monitored garden beds, so the construction works were unlikely to impact on surveys.
Bird and insect surveys
We performed observational surveys using timed transect walks to record invertebrates and birds utilising the site and recorded their abundance, behaviour, and the elements of the roof they were interacting with (plant species, substrate, or artificial structures). Behaviours recorded included interaction with flowering parts (i.e., collecting pollen or nectar), resting on plant tissues or artificial surfaces, nesting in artificial structures (mostly for spiders), or “flying through” where the organism was sighted but did not land or interact with any elements of the roof. Invertebrate ‘presence’ was also noted in cases where organisms were not seen, but traces were left such as webs, or leaf damage, but no abundance was recorded in these instances. Transect walks included active searching of cavities and vegetation for invertebrate presence, however with a focus on floral visitors. Surveys were conducted by two observers, who visited independently on separate sampling occasions. Each observer visited each site at least once before the green roof was installed, but only one observer was available to conduct the surveys after the installation.
There were large differences in site size, and total sampling time was scaled to allow for more consistent search effort across sites. For larger roof sites (>1000 m2) such as the impact site, and the green roof reference site we used a defined transect walk of approximately 100 m for 45 min. The ground site (Docklands Park) was surveyed initially as part of another study (Berthon 2022) with a similar 100 m transect surveyed for 30 min. Control sites were smaller (~300 m2) and a transect of approximately 60 m was lain and surveyed for 30 min of active search time. Invertebrates were identified to order level during surveys and photographed to enable further classification to order and family level, using online resources and taxonomic expertise of the researchers involved. Invertebrates were only recorded if they were seen actively interacting with plant or artificial substrates, and where invertebrate identification was not possible, the observation was noted as an ‘Unknown’ species and added to total abundance counts, but not used in the calculations of taxonomic richness. Bird species encountered were recorded in a similar fashion but able to be identified on site.
Plant surveys
We used information from site managers and previous surveys to generate plant species lists for the green roof and ground sites, but spontaneous plant species occurred frequently in later surveys. Plant lists contained species and varieties (where information was available). For comparing plant richness across sites, plants were reduced to genus level to avoid discrepancies with identifying hybrids between observers. Plant origin was recorded as native to Australia, or exotic (originating from outside Australia). A total list of plant taxa on each site is given in Supplementary Material S1.
On each sampling occasion we recorded the proportion of plant genera that were flowering and the total floral abundance. Floral abundance was measured as the total number of flowering units within the transect area was calculated on each sampling occasion. Flower units were counted as the smallest attractive unit of an inflorescence, e.g., single flowers for plants with simple flowers (e.g., Viola (Violaceae) flowers) or plants with inflorescences where pedicels are distinct e.g., Plectranthus (Lamiaceae) flowers, the entire composite flower of an Asteraceae sp., or the floral spike of a Banksia (Proteaceae) or Callistemon (Myrtaceae). To account for differences in the sampling area of each roof, floral abundance was converted to floral density i.e., the average number of floral units per m2.
Statistical analyses
Invertebrates were split into 14 taxonomic groups that we expect to vary in their plant preferences, and ability to colonise rooftops: ants (Formicidae); beetles (Coleoptera); bugs (Hemiptera); butterflies and moths (Lepidoptera); exotic honeybees (Apis mellifera), native bees (primarily Halictidae and Colletidae); dragonflies (Odonata); hoverflies (Syrphidae and Bombylidae families); other flies (primarily Sarcophagidae); mantids (Mantodea); spiders (Araneae); wasps (Vespidae); thrips (Thysanoptera); and silverfish (Lepismatidae). Native insects were those that have originated in Australia, and exotics are those that have been introduced, typically post-European-colonisation. For most groups in Australia, origin is relatively unknown, and we were only able to distinguish origin reliably for the Hymenoptera. The richness of taxonomic groups and total abundance of invertebrates was calculated per sampling period. Richness estimates only included observations that were able to be identified to at least ordinal-level, and included invertebrates observed ‘flying through’ the site. Spiders were counted in richness estimates where webs were present, but these observations were not included in abundance estimates. Leaf damage observations were unable to assigned to a particular species and were also not included in richness or abundance estimates. Birds were analysed as separate species or genera given their ease of identification. All statistical analyses below were run in R statistical computing software (R Core Team 2022).
Comparing invertebrate diversity and abundance before and after the roof was built
The richness and abundance of invertebrates and birds on rooftops before and after the green roof retrofit was constructed was compared using generalised linear regression models including roof type and time period (‘before’ or ‘after’ the green roof installation) as explanatory variables. Interaction effects were used since the impact roof was expected to change the most between time periods, and observer effects were included using observer as an additional fixed effect. The full model formula is given below:
The abundance data was modelled using the lme4 package (Bates et al. 2015) with a Poisson distribution as is common for count data (Sellers et al. 2012), and a Gaussian distribution for the richness data. We built models for birds and invertebrates separately, given differences in taxonomic resolution. The Poisson models were evaluated using the z-distribution for p-value testing, and the gaussian model were evaluated with a t-distribution, both with 16 degrees of freedom. To visualise the interactions, a complimentary regression tree analysis was used that iteratively splits the response variable based on the groups present in the independent variables. Final groupings are decided based on splits that minimise the variation of the response variable within groups and maximises the variation in the response variable between groups. Splits are performed progressively for each independent variable, and if subgroups do not meet this rule, or all options are exhausted then no further splits are made. Regression trees were implemented and visualised using the rpart (Therneau & Atkinson 2022) and rpart.plot (Milborrow 2022) packages.
Comparing compositional differences
The composition of invertebrates and birds was compared across roof types using NMDS ordination techniques from the vegan package (Oksanen et al. 2022). Invertebrate abundance data were reduced to presence/absence to account for biases in detection of organisms among rooftops. These biases were particularly a problem for spiders, which were sometimes identified as present based on web observations without accurate abundance counts. Therefore, relative abundances between rooftops were not used in compositional analyses. To visualise composition differences the impact roof was split into two distinct categories since the composition of invertebrates is expected to change after plants were installed and the composition was analysed per sampling visit rather than compiling across visits for a single site. A dissimilarity matrix was calculated using Bray–Curtis dissimilarity and the differences were visualised by importing the output of the NMDS into the ggplot2 plotting framework (Wickham 2016).
Abundance and diversity of invertebrates through time
A time series of colonisation was visualised for the impact roof based on the presence of different taxa during each sampling visit across the study period. Since the effect of time period (before/after greening) on abundance was previously analysed, and there was a large gap in between sample periods, temporal trends are descriptive. However, linear regression models were used to test whether invertebrate richness and abundance (with a Poisson distribution) changed as a result of floral density and richness of flowering roof plants, which increased as the roof matured. Similar time series were built for the other green roofs and for birds. Regression models were run for the impact site alone, and all sites combined.
Plant use by invertebrates and birds on rooftops
Where plants were present, the total proportion of plants that were interacted with by at least one insect was calculated. A species interaction matrix was built for each roof to describe the interaction of invertebrates with plants and artificial surfaces on rooftops. The impact site was split into separate before and after webs considering changes in availability of substrates. Animals that were recorded on site as ‘flying through’ but not interacting with the site elements were removed from this analysis. Primarily this was to understand the plant use and success of plantings on the roof when compared to our green roof reference site and to begin to collate data for informing green roof plant choices. Using the bipartite package (Dormann et al. 2009) we analysed the network to find particularly useful plant species (plant ‘hubs’) using the species strength metric (Bersier et al. 2002) which describes the relative dependency of the invertebrate groups on each plant species. Networks were built for birds (species) and invertebrates (groups) separately considering they had different levels of taxonomic resolution. Since birds were rare and highly mobile, a single network was made across all sites to determine preferences.
Results
Plant density and diversity
Plant diversity and floral density differed between sites (Table 2), with the newly established retrofit increasing in floral diversity and abundance over time (see below). The green roof reference had the highest average floral density per sampling period, and, after greening, the impact site had higher floral density than the ground site. Some spontaneous plant species were also found to be growing on the bare roof control site, however most had died by the end of the study, likely due to high exposure and lack of water on the roof.
Timed survey results
A total of 388 observations of invertebrates and 37 of birds were recorded as well as their interaction with roof elements, across 26 sampling occasions. Some of the invertebrate observations were of insect-related leaf damage, but without visual identification of the perpetrator, or small flying insects that were unable to be classified to order level (4% of observations). These were classed as ‘Unidentified’ and removed from further analyses. An additional 7 observations were made of dead animals present on the rooftops, but these were not included in the observation count since, in most cases, it is uncertain whether they had died on the roof or been carried in by the wind. Two of these insects were caught in spider webs and it is assumed they had died on the roof, but it is possible they had been blown into the webs, so they were excluded from analysis. For the two moth taxa that were found dead on the bare roof control, their cause of death is likely exposure since both were sheltering in either roof structures or a cocoon.
One observation was also made of a fungal disease growing on the newly constructed green roof, and anecdotes of aphids and ladybeetle predation were also given to the authors from construction workers while the roof was inaccessible. These are not used in analyses but are useful notes for the understanding of green roof systems. There is likely a wealth of this kind of tacit knowledge present in green roof organisations and maintenance teams, and this could be the topic of future work.
Changes in richness and abundance of fauna
Linear models and regression trees showed differences in invertebrate richness across site types and time periods (i.e., before and after construction) (Supplementary Material B). Observer effects were negligible (p > 0.6) and did not appear as splits in the regression trees. Before the impact roof was built, the reference green roof had higher richness (t = 2.39, p = 0.03, Fig. 2a) and abundance (z = 10.11, p < 0.01, Fig. 2b) of invertebrate groups than all other rooftops. The ground reference also showed higher richness (t = 1.67, p = 0.12) and abundance (z = 6.3, p < 0.001) than control and impact roof sites (Fig. 3). After the green roof was installed, the impact site increased in richness (t = 1.63, p = 0.12, Fig. 3) and abundance (z = 1.99, p < 0.001) of invertebrates and became more similar to the ground site (Fig. 2). The control green roof site also showed a trend towards increased invertebrate richness (t = 1.34, p = 0.2, Figs. 2a and 3), and overall, sites increased in abundance of invertebrates (z = 1.18, p = 0.04, Fig. 2b). Conversely, the bare roof control and ground sites showed a trend towards decreasing invertebrate richness and there was much more variability between visits for the ground site after the retrofit roof was installed (Fig. 2). Unfortunately, the reference site was unable to be sampled after the green roof was built due to COVID-19 restrictions.

Comparison of the a richness and b abundance of invertebrates, as well as c richness and d abundance of birds before (light grey) and after (dark grey) the roof was constructed for each roof type

Regression tree for partitioning variation in richness of invertebrate groups between site types across each sampling period. Terminal branches are shaded based on invertebrate richness, with higher richness indicated by darker hues. Number of observations in each terminal node are also indicated. Regression trees for the other models are given in Supplementary Material B
Bird richness and abundance was not related to either roof type or time period (Supplementary Material B, Fig. 2c, d). There was a trend for a lower abundance of birds on the ground site and on the green roof control. The bare roof control site had a breeding colony of silver gulls (Chroicocephalus novaehollandiae) on an adjacent rooftop, explaining the high abundance of birds that were recorded. Multiple birds were observed using the impact roof before the green roof was built, however it is likely that ongoing disturbance from construction had impacted on their use of the roof. Overall, the Impact site had the highest number of bird species (Table 3), but different species appeared before and after the green roof retrofit was built.
Compositional changes
NMDS analysis (stress = 0.1556) shows that invertebrate composition was primarily driven by differences in floral density (r = 0.2, p = 0.1) between sites, with clustering of the reference sites, as distinct from the control rooftops (Fig. 4). Before the green roof was built, the invertebrate composition of the impact roof was similar to the control rooftops and contained no flowers. After the green roof retrofit was built, the impact roof composition became closer in composition to the reference sites. Dragonflies and native bees were only found on reference sites, whereas thrips were only seen on the Impact site after it had been planted. A single praying mantis was found while searching the green roof reference site.

Ordination plot comparing the composition of invertebrate groups across sites. Each point represents a single sampling visit at a site and sites that are closer together are more similar in composition. Invertebrate composition is driven by floral density as well as gradients in temperature and humidity (blue arrows). The impact site has been highlighted in red and separated into before the green roof was built on the impact site ("Impact"), and after (“Retrofit”)
The composition of the retrofit roof immediately after planting when the plants were young and there was low floral density, is an outlier to all other sampling periods for this site (see highlights in Fig. 4). The green roof control site also had low floral density, with only one or two flowering plant species (Aeonium, Strelitzia, or Oxalis) present per sampling visit after the green roof was built.
Bird composition was different across sites and multiple species were only observed once, or only on one site. As a result, we could not build a reliable ordination for bird composition, but the birds observed are listed in Table 3. Honeyeaters are known to exist on the ground site (Berthon 2022) but were only seen foraging on Larkspur (Delphinium sp.) on the impact site after it was greened. Similarly, Noisy Miners (Manorina melanocephala) were only observed once on the ground site.
Colonisation over time for the impact roof
Immediately after the green roof was built, there was low richness and abundance of invertebrates, and the latter was lower than the baseline surveys (Fig. 5). There was an exponential increase in abundance of invertebrates over time on the impact roof, mirrored by an increase in floral abundance (Fig. 5A). Floral diversity and invertebrate diversity also increased, but at a slower rate (Fig. 5B). For the impact site alone, linear regression models showed that invertebrate abundance across both time periods was strongly correlated to floral density (z = 12.13, p < 0.0001, df = 5) and floral richness (z = 7.14, p < 0.0001, df = 6). The same trends were present for invertebrate richness (p = 0.04 and 0.03, respectively, Fig. 5B). There was no relationship between bird richness or abundance and floral density on the impact roof (p > 0.3, Fig. 5C, D). There was a slight decrease in bird abundance as floral richness increased (p = 0.035, df = 6), but the effect size was very small, and likely not biologically relevant (Fig. 5C). Other sites showed fluctuations in invertebrate abundance through time, but with no consistent trend, and invertebrate richness was relatively stable across time (Supplementary Material C).

The abundance (A, C) and richness (B, D) of invertebrate groups (A, B) and birds (C, D) related to the richness and density of floral resources on the impact roof over time before and after the green roof retrofit was built. Red lines show changes in animal richness or abundance, while blue lines show parallel changes in richness and abundance of floral resources. Trends through time for other sites are provided in Supplementary Material C
Across all rooftops, invertebrate abundance increased with increasing floral density (z = 15.22, p < 2e-16, df = 24), and richness (z = 11.52, p < 2e-16, df = 25). Across all rooftops, bird abundance (p > 0.9) and richness (p > 0.13) was not related to floral density or richness. All model outputs and graphs related to floral abundance and richness are provided in Supplementary Material C.
Plant use on rooftops
Only 53% of plants were observed to have interactions with invertebrates in newly retrofitted rooftops, 90% on the control green roof, 55% on the green roof reference, and 94% on the ground site (Supplementary Material A, Table 1). The proportion of flowering plants that were utilised was between 56 – 100% and varied between roof types. On green roofs and ground sites, the plant species with the highest species strength also had a high floral density (Table 4). When bare, invertebrates on the impact site largely used artificial surfaces, but when greened, large flowering Asteraceae species had the highest species strength and supported the most invertebrate groups (Fig. 6). Native bees were only found on sites planted with native plant species, where they foraged from species such as Dianella sp., Scaveola sp. and Micromyrtus sp. Artificial structures were utilised by birds, spiders, and flies, and these made up the majority of interactions on bare rooftops but were also utilised on green roofs and ground sites. An interaction matrix of all plant species and their associated plant partners is given in Supplementary Material A, and all network diagrams are given in Supplementary Material D.

Network diagram for impact before (A) and after (B) the green roof retrofit was built. The richness of organisms and the diversity of interactions increase after the retrofit installation. Plant genus names are provided, but full species lists are in Supplementary Material A. “Artificial” refers to artificial structures. Network diagrams for each site are given in Supplementary Material D
Discussion
In an increasingly urbanised world, green roof retrofits may hold the key to providing additional habitat for fauna in cities. This study quantifies the additional biodiversity value provided by a single new green roof retrofit and extends a body of literature arguing that the placement, design, and connectivity of these rooftops will affect their ability to be actively used, particularly by low mobility invertebrates (Blank et al. 2017; Mayrand and Clergeau 2018; Dromgold et al. 2020). We show an increase in invertebrate richness and abundance across roof types and across time for the retrofit roof, concurrent with increase in floral density and diversity. Conversely, the visitation of birds, which are highly mobile, was not different across roof types, time periods or in response to floral density and richness. Instead, birds have been observed to only visit rooftops where their preferred forage is present (Fernandez-Canero and Gonzalez-Redondo 2010) and respond positively to shrub richness (Wang et al. 2017). Similarly, we find that primarily nectivorous birds (Anthochaera sp.) were only present on sites with flowers. Therefore, the effectiveness of green roof design will depend on the movement and colonisation of invertebrates and birds, as well as their ability to utilise plant resources. Throughout this discussion we use ‘colonisation’ to mean appearance of fauna on the roof, which includes both transient visitors as well as residents.
The colonisation of the retrofit rooftop was surprisingly rapid for some invertebrates however there were also some invertebrates that never appeared on the rooftop during our study. For example, honeybees (Apis mellifera) were present and foraging on flowers four days after the plants were installed, whereas native bees were not found at all on the retrofit rooftop. Conversely, native bees and honeybees were both found to be foraging on the green roof reference site. Very low mobility organisms such as mantids were also present on the green roof reference site and not elsewhere. Previous studies have shown that a similar disparity between colonisation of low and high mobility invertebrates (Madre et al. 2013; Braaker et al. 2014; Berthon 2015; Dromgold et al. 2020). For example, Madre et al. (2013) found that 86% of species found on green roofs were classified as “good” dispersers. Similarly, Berthon (2015) found that compositional differences between high and low roofs (defined as < 7.5 stories high) was driven by presence of low mobility taxa on lower rooftops.
Differences in colonisation and composition of invertebrates may be due to differences between rooftops and reference sites in age, connectivity with surrounding habitat patches, and plant composition. The green roof reference and ground site are established sites (> 2 years), and it is possible that native bees may disperse to rooftops later in the colonisation sequence, beyond the sampling period of our study. Several studies show differences in invertebrate composition and richness (Jones 2002; Schrader and Böning 2006; Braaker et al. 2014; Dromgold et al. 2020), bird diversity (Wang et al. 2017) and plant composition (Ksiazek-Mikenas et al. 2018) with increased with time since establishment of the green roof. Despite the short time frame of our study, we show a sharp increase in invertebrate richness within the first three months, and a rapid shift in invertebrate composition with the retrofit becoming compositionally more similar to reference sites after the green roof was installed. This is encouraging, and lends some weight to the often-criticised sentiment: ‘if we built it, they will come’ (Williams et al. 2014; Ksiazek-Mikenas et al. 2018). However, our results represent only a small snapshot of the opportunity for colonisation and longer-term studies of roof dynamics using space-for-time substitution show linear increases in species richness even after two years post-establishment (Ksiazek-Mikenas et al. 2018).
The initial composition of the retrofit roof is expected to be a combination of organisms that have arrived on the roof through human-mediated mechanisms and those that have actively migrated to the roof (Berthon et al. 2015; MacIvor and Ksiazek 2015). Surprisingly, we did not find much evidence of accidental human-mediated transport on plants or in soil given a decrease in richness of invertebrates immediately post construction. A few fungal gnats were present in the soil and likely were from existing eggs, and there was evidence of leaf damage but no sign of perpetrators. This is perhaps not surprising given that nursery plants and soil are often sterilised before transport onto green roofs (MacIvor and Ksiazek 2015), and accidental human-mediated transport has only been recorded for land snails (McKinney et al. 2019). In addition, detection of soil-dwelling organisms is limited in our study by lack of sampling of the soil, which can harbour eggs and other soil organisms. In either case, this suggests that most invertebrates arrived after the roof was constructed.
There are two primary pathways for invertebrates to colonise the roof over time, either active migration or accidental transport through wind currents (Joimel et al. 2018; Ksiazek-Mikenas et al. 2018). For example, wind dispersal is considered a likely mechanism for low mobility collembola to reach green roofs (Joimel et al. 2018). Increasing age and maturity of plant species may increase resource provision and attract further invertebrates to the roof as demonstrated by the strong correlation of invertebrate abundance and floral abundance in our study. Plant resources are also necessary for plant-dependent species to establish resident populations, and nesting resources are often a limiting factor in urban environments (Prendergast et al. 2022a, b). Alternatively, there may be increased opportunities for invertebrates to arrive to the roof by chance the older the roof becomes. In both cases, the chance of active or accidental colonisation of a green roof will increase with increased connectivity at the landscape scale (Berthon 2015; Ksiazek-Mikenas et al. 2018).
Green roofs are isolated, both vertical and horizontally, such that the height of the building and the distance from nearby ground sites are important factors influencing colonisation potential of invertebrates (reviewed in Blank et al. 2017) and birds (Wang et al. 2017). For example, Dromgold et al. (2020) found that the location of the roof near ground greenspace was more important than roof age, height, and size for determining the invertebrate community that colonised. Similarly, Wang et al. (2017) found a lower diversity of both birds and butterflies on taller rooftops, across all guilds. Though we did not specifically test for the influence of connectivity in our study, the green roof reference site is very low and well connected, which may have allowed for colonisation by low mobility organisms like mantids and native bees.
Movement of invertebrates between rooftops is largely unknown (Mayrand and Clergeau 2018), but here we find some evidence for a ‘spill over’ effect, where invertebrate richness of the control rooftop increased after the installation of the green roof retrofit. This could be because of a cumulative effect for adjacent rooftops providing a greater signal for attracting actively dispersing invertebrates. Alternatively, the highly attractive retrofit may cause invertebrates to ‘spill over’ onto adjacent rooftops. A similar spill over effect was not present for the bare roof control, but this roof had undergone construction in the intervening period and did not have a direct line of sight to the retrofit roof like the green roof control did.
Finally, differences in the plant composition and richness between sites, and the association of invertebrate and bird taxa with different plant species, indicate the importance of plant choice for enhancing resource provision on green roofs. The green roof reference and ground sites were planted entirely with native plant species, whereas the green roof retrofit was built primarily as an agricultural rooftop with a diversity of both native and ornamental flowering plants. Use of native plants on green roofs has been shown have little effect on the diversity of beetles on green roofs in Melbourne (Dromgold et al. 2020) but has been suggested to enhance biodiversity elsewhere (reviewed in Berthon et al. 2021a, b, but see Prendergast et al 2022b, Prendergast 2022). Use of native plants may also be beneficial for plant conservation (reviewed in Butler et al. 2012; Li and Yeung 2014) provided the conditions are suitable. In our study, native bees were only present and utilised native plants exclusively on the green roof reference and ground sites but have been known to utilise ornamental plants in ground settings (Berthon 2022; Prendergast and Ollerton 2021; Prendergast 2022). Studies in ground sites have also shown native bees to be more prevalent in areas with native vegetation rather than ornamental plants (Prendergast et al. 2022b). Conversely, native plants that are known to be utilised by native bees (e.g., Brachyscome, Supplementary Material A) were present on the impact roof but were rarely utilised by invertebrates.
Provision of flowering plants was important for increasing the abundance and richness of invertebrates and drove differences in invertebrate composition across sites. For example, the green roof control site had very low flowering richness and had the lowest corresponding richness of invertebrates across all green sites. Compositionally, this low diversity resulted in an invertebrate community not dissimilar from that of the bare roof control and dominated by primarily web building spiders. Previous studies have investigated associations between invertebrates and vegetation type (Dromgold et al. 2020; Gonsalves et al. 2022), vegetation cover (Kyrö et al. 2018; Wooster et al. 2022) or structure and diversity (Madre et al. 2013; Ksiazek-Mikenas et al. 2018), but have rarely investigated floral resources (Tonietto et al. 2011; Benvenuti 2014). For example, Tonietto et al. (2011) found that floral resources were lower on green roofs compared to other greenspace types and showed differences in the bee community composition. Benvenuti (2014) measured pollinator visitation in relation to blooming season and found that pollinator visitation was related to climatic conditions on the roof rather than attractiveness of individual flowers. In ground environments, increasing floral diversity has been shown to increase the diversity and abundance of invertebrates (e.g. Plascencia and Philpott 2017), and cause changes in dietary preferences (Berthon 2022) particularly for pollinators. In our study, the plants most used by invertebrates and birds (plant ‘hubs’ or ‘highly attractive’ plants sensu, Lowenstein et al. 2019) were also flowering species such as Chrysanthemum or Scaevola.
Recommendations for green roof design
Due to the short time frame and low sample size in our study, we are unable to distinguish between the effect of roof age, composition, and connectivity on the appearance of invertebrates on green roofs. However, our results suggest that there is a substantial potential for colonisation of green roof retrofits with a diversity of invertebrates and that diversity of flowering plants is key for enhancing available plant resources. While inclusion of native plants appears to be crucial for supporting some native pollinators, the landscape context of the placement of green roof retrofits, and roof height may inhibit the potential of these spaces to be colonised by low mobility groups. The presence of birds, a high mobility group, is not as limited by rooftop isolation but will depend on the plant resources provided as well as their ability to attract invertebrates.
Where funds are limited, strategic placement of green roofs near large ground habitats, or on low rooftops should be prioritised. Green walls may increase the connectivity of roof and ground sites, and adjacency of additional green roofs may allow for roof-roof movement, but this remains to be tested (Mayrand and Clergeau 2018). Balcony vegetation is another promising avenue for increasing quantity and connectivity of habitat but has not so far been considered at a landscape scale. Increasing the total number of green roofs in the urban environment will also have cumulative benefits for other ecosystem services such as cooling and stormwater regulation (Manso et al. 2021).
Few studies have attempted to assess thresholds of height, area, and connectivity of green roofs for prioritisation of green roof placement. For example, Berthon (2015) found that isolation in one dimension (i.e., height or horizontal connectivity) could potentially be compensated for by reducing isolation in the other. For example, tall rooftops nearby ground sites had similar invertebrate diversity to low rooftops further from ground habitat patches. Similarly, Wang et al. (2017) suggests that rooftops lower than 50 m and with green cover > 1100 m2 are optimal for greening. Data on native bees from America found that nests on rooftops below 5 stories were more successful (MacIvor and Ksiazek 2015). While these thresholds are useful thought experiments, it is worth noting that there is still huge potential for even small spaces to attract invertebrates in the city (Mata et al. 2019; Berthon et al. 2021a, b) and there are cumulative benefits from greening adjacent residential properties in a co-ordinated way (Goddard 2010). Similarly, vertical connectivity may be provided somewhat by flowering trees to connect ground habitats to balconies, but the foraging behaviour of bees in trees remains understudied.
There may be cases where habitat can be created but connectivity is severely limited, and the location of the roof is desirable for its other environmental benefits. In these cases, managers may consider translocating fauna or floral onto rooftops for conservation. This may be of particular benefit for locally threatened and wind pollinated plants since they do not rely on local pollinators and would be able to disperse seed at greater distances, taking advantage of elevation and urban wind currents (Li and Yeung 2014). For faunal translocations, care must be taken to ensure that the green roof provides all necessary habitat requirements (Fernandez-Canero and Gonzalez-Redondo 2010) and hosts conditions viable for growth (Baumann 2006), as well as take into account the potential for genetic drift (Gillespie 2001). Successful translocations are already apparent in integrated pest management applications and could be applied to the urban conservation context, particularly for small invertebrates that may make good use of small green roof habitats.
Finally, buildings provide structural habitat, even without the inclusion of plants, for a range of birds (Mainwaring 2015), bats (Johnson et al. 2019), spiders (Mammola et al. 2018) and bees (Dollin 2021). For example, several bird species have been noted to successfully breed on unvegetated rooftops or other unintended artificial structures such as Peregrine Falcons on a building in Melbourne, Australia (Mirvac n.d.), Ospreys on a light pole in Florida, USA (University of Florida n.d.). Here we found silver gulls (Chroicocephalus novaehollandiae) nesting on an adjacent bare rooftop to our bare roof control but breeding success has not been monitored here. These artificial habitats can be enhanced through careful design that considers dual use of built structures for humans and nature. This can include design specifically for inclusion of animals into the structure of the building (e.g., “bird bricks”, https://www.birdbrickhouses.co.uk/), as well as green infrastructure retrofits.
Data Availability
Data is available on request.
References
Australian Bulding Codes Board (1996) Building Code of Australia. Volume 1. Commonwealth and States and Territories of Australia. Accessed 17 Aug 23 https://ncc.abcb.gov.au/system/files/ncc/BCA_96_Volume_One_Amdt_11.pdf
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48. https://doi.org/10.18637/jss.v067.i01
Baumann N (2006) Ground-nesting birds on green roofs in Switzerland: Preliminary observations. Urban Habitats 4(1):37–50
Beninde J, Veith M, Hochkirch A (2015) Biodiversity in cities needs space: a meta-analysis of factors determining intra-urban biodiversity variation. Ecol Lett 18(6):581–592
Benvenuti S (2014) Wildflower green roofs for urban landscaping, ecological sustainability and biodiversity. Landsc Urban Plan 124:151–161
Bersier L-F, Banašek-Richter C, Cattin M-F (2002) Quantitative descriptors of food-web matrices. Ecology 83(9):2394–2407
Berthon (2022) Plant-insect interactions in the city. Royal Melbourne University of Technology, Melbourne (PhD Thesis)
Berthon K (2015) Invertebrates on green roofs in Sydney. Masters Thesis, Macquarie University
Berthon K, Davies P, Nipperess D, Bulbert M (2015) Confirmed at Last: Green Roofs Add Invertebrate Diversity. State of Australian Cities 2015, QT, Gold Coast, QLD, Urban Research Program at Griffith University on behalf of the Australian Cities Research Network - http://soacconference.com.au/soac-conference-proceedings/#sthash.lgvjE33F.dpuf
Berthon K, Meyer ST, Thomas F, Frank A, Weisser WW, Bekessy S (2021a) Small-scale habitat conditions are more important than site context for influencing pollinator visitation. Front Ecol Evol 544
Berthon K, Thomas F, Bekessy S (2021b) The role of ‘nativeness’ in urban greening to support animal biodiversity. Landsc Urban Plan 205:103959
Blank L, Vasl A, Schindler BY, Kadas GJ, Blaustein L (2017) Horizontal and vertical island biogeography of arthropods on green roofs: a review. Urban Ecosyst 20(4):911–917
Braaker S, Ghazoul J, Obrist MK, Moretti M (2014) Habitat connectivity shapes urban arthropod communities: the key role of green roofs. Ecology 95(4):1010–1021
Brenneisen S (2006) Space for urban wildlife: designing green roofs as habitats in Switzerland. Urban Habitats 4(1):27–36
Burszta-Adamiak E, Fiałkiewicz W (2019) A review of green roof incentives as motivators for the expansion of green infrastructure in European cities. Przegląd Naukowy Inżynieria i Kształtowanie Środowiska 28
Butler C, Butler E, Orians CM (2012) Native plant enthusiasm reaches new heights Perceptions, evidence, and the future of green roofs. Urban For Urban Green 11(1):1–10
Cascone S, Catania F, Gagliano A, Sciuto G (2018) A comprehensive study on green roof performance for retrofitting existing buildings. Build Environ 136:227–239
Cook-Patton SC, Bauerle TL (2012) Potential benefits of plant diversity on vegetated roofs: A literature review. J Environ Manag 106:85–92
Dollin A (2021) Are blue banded bees building in your house? Retrieved 22 Mar 2022, from https://www.aussiebee.com.au/bluebandedbees-in-house.html
Dormann CF, Fruend J, Bluethgen N, Gruber B (2009) Indices, graphs and null models: analyzing bipartite ecological networks. Open Ecol J 2:7–24
Dromgold JR, Threlfall CG, Norton BA, Williams NSG (2020) Green roof and ground-level invertebrate communities are similar and are driven by building height and landscape context. J Urban Ecol 6(1)
Dunnett N (2006) Green roofs for biodiversity: Reconciling aesthetics with ecology. Ann Green Rooftops Sustain Commun Conf, Boston, Boston
Faeth SH, Bang C, Saari S (2011) Urban biodiversity: patterns and mechanisms. Ann N Y Acad Sci 1223(1):69–81
Fernandez-Canero R, Gonzalez-Redondo P (2010) Green roofs as a habitat for birds: a review. J Anim Vet Adv 9(15):2041–2052
Gedge D, Kadas G (2005) Green roofs and biodiversity. Biologist 52(3):161–169
Gillespie JH (2001) Is the population size of a species relevant to its evolution? Evolution 55:2161–2169
Goddard MA (2010) Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol Evol 25(2):90–98
Gonsalves S, Starry O, Szallies A, Brenneisen S (2022) The effect of urban green roof design on beetle biodiversity. Urban Ecosyst 25(1):205–219
Hunter MR, Hunter MD (2008) Designing for conservation of insects in the built environment. Insect Conserv Divers 1(4):189–196
Johnson JS, Treanor JJ, Slusher AC, Lacki MJ (2019) Buildings provide vital habitat for little brown myotis (Myotis lucifugus) in a high-elevation landscape. Ecosphere 10(11):e02925
Joimel S, Grard B, Auclerc A, Hedde M, Le Doaré N, Salmon S, Chenu C (2018) Are Collembola “flying” onto green roofs? Ecol Eng 111:117–124
Jones RA (2002) Tecticolous invertebrates: A preliminary investigation of the invertebrate fauna on green roofs in urban London. English Nature, London
Joshi M, Selmi W, Binard M, Nys G-A, Teller J (2020) Potential for urban greening with green roofs: a way towards smart cities. Conference Proceedings of the International Conference of Smart Data Smart Cities. https://doi.org/10.5194/isprs-annals-VI-4-W2-2020-87-2020
Kirk H, Soanes K, Amati M, Bekessy S, Harrison L, Parris K, Ramalho C (2023) Ecological connectivity as a planning tool for the conservation of wildlife in cities. MethodsX 101989. https://doi.org/10.1016/j.mex.2022.101989
Köhler M, Ksiazek-Mikenas K (2018) Green roofs as habitats for biodiversity. Nature based strategies for urban and building sustainability. Butterworth-Heinemann, pp 239–249
Ksiazek-Mikenas K, Herrmann J, Menke SB, Köhler M (2018) If you build it, will they come? Plant and arthropod diversity on urban green roofs over time. Urban Nat 1:52–72
Ksiazek K, Fant J, Skogen K (2012) An assessment of pollen limitation on Chicago green roofs. Landsc Urban Plan 107(4):401–408
Kyrö K, Brenneisen S, Kotze DJ, Szallies A, Gerner M, Lehvävirta S (2018) Local habitat characteristics have a stronger effect than the surrounding urban landscape on beetle communities on green roofs. Urban For Urban Green 29:122–130
Li WC, Yeung KKA (2014) A comprehensive study of green roof performance from environmental perspective. Int J Sustain Built Environ 3(1):127–134
Lowenstein DM, Matteson KC, Minor ES (2019) Evaluating the dependence of urban pollinators on ornamental, non-native, and ‘weedy’floral resources. Urban Ecosyst 22:293–302
Lundholm JT (2006) Green roofs and facades: a habitat template approach. Urban Habitats 4(1):87–101
Lundholm JT, Richardson PJ (2010) MINI-REVIEW: Habitat analogues for reconciliation ecology in urban and industrial environments. J Appl Ecol 47(5):966–975
MacIvor JS, Ksiazek K (2015) Invertebrates on green roofs. Green roof ecosystems. Springer, pp 333–355
MacIvor JS (2016) Building height matters nesting activity of bees and wasps on vegetated roofs. Israel J Ecol Evol 62(1–2):88–96
Madre F, Vergnes A, Machon N, Clergeau P (2013) A comparison of 3 types of green roof as habitats for arthropods. Ecol Eng 57:109–117
Mainwaring MC (2015) The use of man-made structures as nesting sites by birds: A review of the costs and benefits. J Nat Conserv 25:17–22
Mammola S, Isaia M, Demonte D, Triolo P, Nervo M (2018) Artificial lighting triggers the presence of urban spiders and their webs on historical buildings. Landsc Urban Plan 180:187–194
Manso M, Teotónio I, Silva CM, Cruz COJR, Reviews SE (2021) Green roof and green wall benefits and costs: A review of the quantitative evidence. Renew Sustain Energy Rev 135:110111
Mata L, Garrard GE, Fidler F, Ives CD, Maller C, Wilson J, Thomas F, Bekessy SA (2019) Punching above their weight: the ecological and social benefits of pop-up parks. Front Ecol Environ 17(6):341–347
Mayrand F, Clergeau PJS (2018) Green roofs and green walls for biodiversity conservation: a contribution to urban connectivity? Sustainability 10(4):985
McKinney ML, Gladstone NS, Lentz JG, Jackson F (2019) Land snail dispersal, abundance and diversity on green roofs. PLoS ONE 14(11):e0221135
Milborrow S (2022) rpart.plot: Plot 'rpart' Models: An Enhanced Version of 'plot.rpart'_. R package version 3.1.1. https://CRAN.R-project.org/package=rpart.plot
Mirvac (n.d.) Peregrine Falcons at 367. Accessed: 28 Mar 2022. https://367collins.mirvac.com/workplace/building-overview/falcons-at-367-collins
Nipperess DA, Beattie AJ, Faith DP, Ginn SG, Kitching RL, Reid CA, Russell T, Hughes L (2012) Plant phylogeny as a surrogate for turnover in beetle assemblages. Biodivers Conserv 21(2):323–342
Oberndorfer E, Lundholm J et al (2007) Green roofs as urban ecosystems: ecological structures, functions, and services. Bioscience 57(10):823–833
Oksanen J, Simpson G, Blanchet F, Kindt R, Legendre P, Minchin P, O'Hara R, Solymos P, Stevens M, Szoecs E, Wagner H, Barbour M, Bedward M, Bolker B, Borcard D, Carvalho G, Chirico M, De Caceres M, Durand S, Evangelista H, FitzJohn R, Friendly M, Furneaux B, Hannigan G, Hill M, Lahti L, McGlinn D, Ouellette M, Ribeiro Cunha E, Smith T, Stier A, Ter Braak C, Weedon J (2022) _vegan: Community Ecology Package_. R package version 2.6-4. https://CRAN.R-project.org/package=vegan
Plascencia M, Philpott S (2017) Floral abundance, richness, and spatial distribution drive urban garden bee communities. Bull Entomol Res 107(5):658–667
Prendergast KS, Ollerton J (2021) Plant-pollinator networks in Australian urban bushland remnants are not structurally equivalent to those in residential gardens. Urban Ecosyst 24(5):973–987
Prendergast K (2022) The influence of plant species, origin and color of garden nursery flowers on the number and composition of pollinating insect visitors. J Agric Urban Entomol 38:21–40
Prendergast KS, Dixon KW, Bateman PW (2022a) A global review of determinants of native bee assemblages in urbanised landscapes. Insect Conserv Divers 15:385–405
Prendergast KS, Tomlinson S, Dixon KW, Bateman PW, Menz MHM (2022b) Urban native vegetation remnants support more diverse native bee communities than residential gardens in Australia’s southwest biodiversity hotspot. Biol Cons 265:109408
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rondoni G, Borges I, Collatz J, Conti E, Costamagna AC, Dumont F, ... Cock MJ (2021) Exotic ladybirds for biological control of herbivorous insects–a review. Entomol Exp Appl 169(1):6–27
Schindler BY, Griffith AB, Jones KN (2011) Factors influencing arthropod diversity on green roofs. Cities Environ (CATE) 4(1):5
Schrader S, Böning M (2006) Soil formation on green roofs and its contribution to urban biodiversity with emphasis on Collembolans. Pedobiologia 50(4):347–356
Sellers KF, Borle S, Shmueli G (2012) The COM-Poisson model for count data: a survey of methods and applications. Appl Stoch Models Bus Ind 28(2):104–116
Skwarecki B (2017) Don’t buy ladybugs to control aphids, and other gardening myths debunked. Accessed 28 Mar 2022. https://www.lifehacker.com.au/2017/04/dont-buy-ladybugs-to-control-aphids-and-other-gardening-myths-debunked/
Smith EP, Orvos DR, Cairns J Jr (1993) Impact assessment using the before-after-control-impact (BACI) model: concerns and comments. Can J Fish Aquat Sci 50(3):627–637
Southwood T (1988) Tactics, strategies and templets. Oikos: 3–18
Therneau T, Atkinson B (2022) _rpart: Recursive Partitioning and Regression Trees_. R package version 4.1.19. https://CRAN.R-project.org/package=rpart
Tonietto R, Fant J, Ascher J, Ellis K, Larkin D (2011) A comparison of bee communities of Chicago green roofs, parks and prairies. Landsc Urban Plan 103(1):102–108
University of Florida (n.d.) A short history of the OspreyCam at the University of Florida. Accessed: 28 Mar 2022. https://wec.ifas.ufl.edu/extension/ospreycam/history.php
Wang JW, Poh CH, Tan CYT, Lee VN, Jain A, Webb EL (2017) Building biodiversity: drivers of bird and butterfly diversity on tropical urban roof gardens. Ecosphere 8(9):e01905
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag, New York
Wilkinson SJ, Reed R (2009) Green roof retrofit potential in the central business district. Prop Manag 27(5):284–301
Williams NS, Rayner JP, Raynor KJ (2010) Green roofs for a wide brown land: Opportunities and barriers for rooftop greening in Australia. Urban For Urban Green 9(3):245–251
Williams NS, Lundholm J, Scott MacIvor J (2014) Do green roofs help urban biodiversity conservation? J Appl Ecol 51(6):1643–1649
Williams NSG, Bathgate RS, Farrell C, Lee KE, Szota C, Bush J, Johnson KA, Miller RE, Pianella A, Sargent LD, Schiller J, Williams KJH, Rayner JP (2021) Ten years of greening a wide brown land: A synthesis of Australian green roof research and roadmap forward. Urban For Urban Green 62:127179
Wooster E, Fleck R, Torpy F, Ramp D, Irga PJB (2022) Urban green roofs promote metropolitan biodiversity: A comparative case study. Build Environ 207:108458
Acknowledgements
The authors would like to thank the numerous building managers, local residents, and the grounds team at Parliament house, who each assisted us with access to sites, and site-specific plant identification. We would also like to thank Dr. Georgia Garrard for her support in the development of this project, and two anonymous reviewers for their generous comments that greatly improved the final manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This project has been funded by the Victorian Government as a joint initiative between the Department of Environment, Land, Water and Planning and the City of Melbourne. SB was funded by ARC Linkage Grant LP160100324, ARC Discovery Grant DP210103787 and the European Union’s Horizon 2020 research and innovation programme under grant agreement No. 730426 (Urban GreenUP).
Author information
Authors and Affiliations
Contributions
KB and SB designed the research and were awarded the funding. JB and KB planned and collected the data, with assistance from RW in the latter stages of the project. KB analysed the data with help from FE and FT. All authors have contributed their ideas and revisions to the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Berthon, K., Thomas, F., Baumann, J. et al. Floral resources encourage colonisation and use of green roofs by invertebrates. Urban Ecosyst 26, 1517–1534 (2023). https://doi.org/10.1007/s11252-023-01392-2
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01392-2