
Abstract
Corvids are known for their urban exploitation, often using artificial infrastructure to roost and nest. In many areas of its distribution range, the pied crow (Corvus albus) is often common in areas of anthropogenic land use, often breeding and foraging near human settlements. In this study, we monitored pied crow use of cellular telecommunication towers as nesting sites in Pietermaritzburg, South Africa, during three breeding seasons. We found that of the 36 towers situated across the urban mosaic landscape, pied crows used 17 (47.2%) as nesting sites while 19 were not used. Many towers were used in subsequent years as nesting sites. Overall, 41 pied crow nests on towers in urban mosaic landscapes were counted by the end of the study. Towers had a mean (± SE) distance of 1595.2 ± 151.0 m between them and were located mainly in the suburbs of Edendale and Northdale. As shown by the hotspot analysis and spatial distribution of used and unused towers, it was unclear why some towers were not being used as nesting sites. We assessed the relationship between pied crow nesting sites and land cover features. We found that tower nesting was best explained by cropland (wi = 0.58), forest (wi = 0.94), grassland (wi = 0.66), schools (wi = 0.56) and relief (wi = 0.56). The continued persistence of pied crows in urban mosaic landscapes with natural and /or managed green spaces, the availability of anthropogenic infrastructure and food in these, and their behavioural plasticity will likely increase their numbers in such landscapes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
One of the biggest threats to biodiversity in developing countries is the loss of natural habitat because of anthropogenic land transformation (Meyer and Turner 1992; Tilman et al. 1994, 2017; Alexander et al. 2019a,b,c). Urban development generally replaces large portions of land with artificial surfaces of cement and asphalt that make up the roads and buildings (McKinney 2002). As more and more land is being transformed, generally, the habitats and ecological processes that animals depend on become threatened (Shochat et al. 2006). However, some urban mosaic landscapes have natural and/ or managed green spaces interspersed with built areas where various species persist (Downs et al. 2021). Some species, especially those with flexible behaviour, thrive in urban mosaic landscapes (Widdows and Downs 2015, 2016, 2018; Widdows et al. 2015; Singh and Downs 2016a, b; Patterson et al. 2018, 2019; Thabethe et al. 2018; Thatcher et al. 2019a,b).
Corvids are among some of the well-known avian urban exploiters (Altevogt and Davis 1979; Marzluff et al. 1994; Gorenzel and Salmon 1995; Withey and Marzluff 2009; Jokimäki et al. 2017ér et al. 2019; Benmazouz et al. 2021). Their opportunistic and flexible behaviour enables them to use foraging and breeding opportunities in urban landscapes, facilitating the prevalence of urban scavenger corvids (Walcott 1974; Withey and Marzluff 2009; Campbell 2009; Jokimäki et al. 2017ér et al. 2019; Benmazouz et al. 2021). The European hooded crow (Corvus corone) exemplifies the success of some corvid species in urban environments with its spread and success across Europe (Kövér et al. 2015). The occurrence of the common raven (Corvus corax) in Alaska had a positive seasonal correlation with commercial and residential zones (Baltensperger et al. 2013). Many corvid species use artificial nest sites in urban areas, including exotic tree species and anthropogenic infrastructure like utility poles and buildings (roofs and attics) (Jurčević Agić 2006; McIvor et al. 2012ér et al. 2015; Jokimäki et al. 2017; Benmazouz et al. 2021).
Pied crows (Corvus albus) occur in urban areas of Africa, where they are opportunistic exploiters and scavengers (Lamm 1958; Mundy and Cook 1977; Campbell 2009; Kopij 2019; Craig et al. 2021; Nyatanga et al. 2021). With their numbers increasing in South Africa, the presence of pied crows in urban environments has increased (Johnson and Murn 2019; SABAP2 2022). An increase in the pied crow population is considered undesirable because of their threat to other taxa (Dean and Milton 2000; Johnson and Murn 2019). Where they occur in large numbers, they likely exert pressure on other species as most corvids, including pied crows, rob nests of other birds species lowering their breeding productivity and prey on a range of species (Wanless and Jupiter 2002; Dean 2005; Fincham and Lambrechts 2014). In South Africa, their increasing population and range expansion have been attributed to increased anthropogenic infrastructure, including dwellings, roads, and utility lines, resulting in a proliferation of alternate nesting sites, among other factors (Dean and Milton 2003; Cunningham et al. 2016). In particular, anthropogenic structures, including windmills, telephone and electrical poles, and pylons, are central to their present breeding success (Dean and Milton 2000; Cunningham et al. 2016). Pied crows have a history of using artificial structures for nesting, although trees are still prominent in wild areas as nest sites (Malan 1994; Dean et al. 2006). However, their use of anthropogenic infrastructure for breeding purposes in urban areas remains poorly studied. Mundy and Cook (1977) described pied crow nest sites in urban areas of Sokoto, Nigeria, in 1970 and 1971. One of these was on a telecommunication tower (Mundy and Cook 1977).
Although pied crows’ use of roadside utility poles and pylons for nesting has been documented, their use of cellular telecommunication towers as nesting sites is undocumented in South Africa (Malan 1994; Dean and Milton 2000; Dean 2005). Pied crows’ behavioural plasticity and the availability of anthropogenic infrastructure as alternate nesting sites in urban mosaic landscapes could alter their nest site preferences and/or indicate an increase in breeding pairs (Malan 1994; Miller and Cale 2000). Some studies have shown this has resulted in high corvid concentrations (McIvor et al. 2012; Kopij 2019; Craig et al. 2021; Nyatanga et al. 2021). More information is needed to understand better the extent of pied crows’ infrastructure use for nesting in urban areas. Consequently, our study determined the extent of use of cellular telecommunication towers (towers hereafter) as nesting sites by pied crows in Pietermaritzburg, KwaZulu-Natal, South Africa, and how land-use features could influence this use. We predicted that the pied crows would use towers for nesting and that various land use features would further influence this. To our knowledge, this is the first study to report on the nesting of pied crows on towers in urban mosaic landscapes in South Africa.
Methods
Study area
The city of Pietermaritzburg is the main economic hub of the uMsunduzi Municipality (area = ~ 63,511 km2) and located ~ 80 km inland from Durban in KwaZulu-Natal Province, South Africa. It is one of the seven local municipalities found in the uMgungundlovu District Municipality (Fig. 1). Henceforth, we refer to our study area as ‘Pietermaritzburg’ to avoid confusion. The population of Pietermaritzburg is estimated at over 670 000 people (Statistics South Africa 2017) and growing. The Pietermaritzburg central business area is situated in the lower basin of the uMsunduzi River and its tributaries (Fig. 1). The hills surrounding the city separate the urban from the rural on the peripheries. Much of the remaining fertile agricultural land has been developed for housing and road infrastructure to meet growing population demands. It forms a landscape mosaic of urban built interspersed with managed and natural green spaces, subsistence farming and exotic tree plantations. Average annual temperatures differ between 16℃ and 17℃ (http://weathersa.co.za/home/recentclimate). The climate of Pietermaritzburg is strongly influenced by topography. Seasonal changes are typically characterised by austral summer rainfall and dry winters. Mean annual rainfall is usually between 748 and 1017 mm p.a. Higher lying northern and western areas tend to receive more rainfall and thus are relatively cooler than central parts.

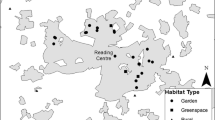
Distribution of pied crow (Corvus albus) nests on cellular telecommunication towers in the study area of Pietermaritzburg, KwaZulu-Natal, Province, South Africa
Nesting sites
During the breeding season, pied crows are monogamous and territorial, building bowl nests of sticks and twigs, sometimes including anthropogenic material like wire (Mundy and Cook 1977; Dean 2005). They generally breed in the austral spring and summer (depending on latitude). Clutch size varies from 1 to 7 eggs with incubation 18–19 days, and the young generally fledge after 35–43 days (Dean 2005). At the start of our study, we had little knowledge of previous pied crow nesting activity on towers and relied on old nest material and personal observations for confirmation. Thus, in November and December of 2016, 2017 and 2018, we monitored known towers (n = 36; Fig. 1) in and around Pietermaritzburg for nesting activity of pied crows. At the start of the surveys, we acquired the geographical location coordinates for each tower using a global positioning system (GPS, Garmin etrex, USA). In each year of surveys, towers were monitored monthly at random to check for nesting activity of pied crows. Breeding was confirmed by observing adult birds incubating on the nest or making frequent flights to the nest on a tower. We monitored towers without nests for 1 h on each visit. For these towers, five more days of monitoring were made for confirmation of no use. Unattended nest material could indicate previous use, but if unattended for extended periods, this was generally destroyed by the weather or removed by cellular telecommunication company technicians. We found no old nesting material visible on non-used towers to suggest previous use.
Spatial analyses
We conducted all landscape analyses of pied crow nest sites on towers using ArcGIS 10.3.1 (ESRI, Redlands, CA, USA). We obtained geospatial data from the Geography Department in the School of Agriculture, Earth and Environmental Sciences, University of KwaZulu-Natal, South Africa. We used the proximity of the Msunduzi shapefile (area = 635.11 km2) to define the study boundary. Towers that fell outside the study boundary were not included in the analyses and therefore reduced the total to 36. We identified key land-use and physiographic features that were likely to influence the breeding of pied crows in this urban mosaic landscape using classified land cover maps (Geoterra Image 2010). We extracted six main features within a 1 km radius (area = 3.14 km2) buffers of each tower (Table 1). Nests of pied crows located on roadside telephone poles were observed separated by 1–2 km (Malan 1994). The spatial distribution of pied crow nests in Pietermaritzburg was examined using the spatial statistic method in ArcGIS 10.3.1 (ESRI, Redlands, CA, USA) (Fig. 1). Point pattern was analysed using Average Nearest Neighbour Analysis distances and Hot Spot Analysis (Cover and Hart 1967) to determine spatial clustering of used towers. Using the frequency of nesting for each used tower, we used the Getis-Ord Gi* z-value (GiZscore) statistic to assess the spatial clustering of nested towers across the Pietermaritzburg study area (Fig. 2). The Inverse Distance Weighing (IDW) tool in ArcGis Spatial Analyst was used to improve the display of the GiZscores for the nested towers by smoothing of the neighbouring points across the extent of the study area. Furthermore, the average distance between towers was calculated using a Near Analysis tool in ArcMap (ESRI, Redlands, CA, USA).

Tower nesting hotspots of pied crow (Corvus albus) in the study area of Pietermaritzburg, KwaZulu-Natal, Province, South Africa
Statistical analyses
We performed all statistical analyses with R 3.6.1 (R Development Core Team, Vienna, Austria). We tested whether there was any correlation amongst predictor variables by visual inspection of the scatterplot matrix. Highly correlated predictor variables were not preferred as they could cause unreliable and unstable estimates (Graham 2003). Thus, we further tested the multicollinearity of predictor variables statistically by calculating variance inflation factors (VIF) (Stine 1995). Housing density, road density and altitude were subsequently removed from the analyses because of an inflated variance factor of greater than five. We performed binary Generalised Logistic Modelling (GLM) to test the effect of land-use and physiographic features on the nesting of pied crows (i.e. nesting = 1, no nesting = 0) on towers in the Pietermaritzburg study area. The best model was chosen based on the lowest (< 1 delta) Akaike information criterion (AIC) values (Burnham et al. 2011).
Results
Nesting sites
Of the 36 towers that we monitored, 17 (47.2%) were positively identified as nesting sites for pied crows, while there was no nesting observed on 19 by the end of the study (Table 1). From the 17 that were used, nine (52.9%) were used for three consecutive breeding seasons, two were used only once, and six were used twice. Overall, used towers accounted for a total of 41 nests by the end of the study. Nesting had reduced on three towers between 2016 and 2017 and one between 2017 and 2018. The observed mean distance (± SE) between towers was 1595.2 + 151.0 m, and the expected distance was 2100.12 m.
We found that the number of towers and associated pied crow nests varied among the suburbs of Pietermaritzburg. Most towers (n = 13) and associated pied crow nests (n = 16) were in Edendale, followed by Northdale (n = 5 towers; n = 6 nests) (Table 1; Figs. 1, 2 and 3). No nests were found in The Grange (n = 1 tower, n = 0 nests), Mkondeni (n = 2 towers, n = 0 nests) and Sweetwaters (n = 3 towers, n = 0 nests) (Figs. 2 and 3).

Summary of suburbs where cellular telecommunication towers were situated in the study area and the total number of pied crow nests found in the present study
Cluster analyses
Overall, the towers used by pied crows for nesting were not statistically clustered across the study area (634 km2) (NN index = 0.759, z = 0.466, P = 0.510), with nesting being more in the centre of the study area. Tower distribution in the study area was more centralised and towards the east (Fig. 2). Hence, when overlaying the used and unused towers layers together, we found that they had similar spatial distribution patterns (Fig. 2). It was unclear why some towers were not used based on tower distribution and nesting hotspots.
Land cover features
When we related nesting site use of towers by pied crows with land cover features (Table 2), the best model was chosen with the lowest (< 1 delta) AIC values (Table 3). The best candidate model and its relative variable importance were cropland (wi = 0.58), forest (wi = 0.94), grassland (wi = 0.66), schools (wi = 0.56), and relief (wi = 0.56) as the main predictors of nesting of pied crows on towers (Tables 2 and 3; Fig. 4). Overall, the GLM mixed model revealed that there was a significant and positive influence on tower nesting by the grassland (β = 13.14 ± 4.73, P < 0.05) and distance to plantations (β = 14.12 ± 6.72, P < 0.05). The number of schools within 1 km2 radius (β = -13.29 ± 1.55, P < 0.001), distance to cropland (β = -8.57 ± 1.58, P < 0.001), and relief (β = -0.67 ± 0.07, P < 0.001) all had a negative and significant influence on nesting site use of towers by pied crows in the study area. Furthermore, nesting by pied crows on towers was influenced negatively but not significantly by distance to forest (P = 0.43) and sugarcane (P = 0.25). Distance to subsistence farming (P = 0.25) and water (P = 0.17) (Table 2) had a positive influence on nesting on towers but not significantly.

Probability of predicted relationship between nesting of pied crows on cellular telecommunication towers and predictor variables in the present study: (a) nearest distance to cropland and (b) forest, (c) number of schools within a km radius buffer of each nesting sites, (d) area of grassland, and (e) relief
Discussion
Pied crow use of artificial structures for nesting purposes in southern Africa has been documented previously (Malan 1994; Dean et al. 2006). Records of pied crows using anthropogenic structures as nesting sites date from the 1960s (Clancey 1964). Our findings showed that of the towers monitored in the urban mosaic landscape, almost half (47.2%, n = 17) were used by pied crows as nest sites. Of these towers, 52.9% (n = 9) were used for three consecutive years of the study. All in all, towers used for nesting accounted for 41 pied crow nests. In particular, some suburbs like Edendale and Northdale had more towers and pied crow nests than other suburbs in the study area. Nesting site use declined on four towers during our study. This may have been because of nest removal by cellular telecommunication company technicians. This reduction showed the inconsistency of nesting on some towers.
The use of artificial structures for nesting is fairly common among corvid species (McIvor et al. 2012; Benmazouz et al. 2021; Noreen and Sultan 2021). As predicted, we found frequent use of towers as nesting sites by pied crows in our study area. Mundy and Cook (1977) first documented pied crows using a telecommunication tower in Africa as a nest site. Urban breeding and alternative use of artificial infrastructure by pied crows could be because of various factors, one of which may be the scarcity of suitable nesting sites. Due to their height, towers could also be chosen based on safety (McGowan 2001).
Nineteen towers were not used as nesting sites by pied crows in the study. Notably, towers in certain suburbs had no nesting during our study. There was a possibility of nests being removed, although we made no such observations during our surveys. One of the landowners reported that a pied crow nest was removed from the tower in the past (pers. comm.). Corvid nests on anthropogenic structures, especially utility poles or pylons, are sometimes removed by maintenance staff to combat electrical faults caused by nests (McIvor et al. 2012). For example, nests of hooded crows are often removed from power poles in Scotland; however, nest removal was not a permanent solution in deterring them from nesting on power poles, as they would often nest on the same poles again (McIvor et al. 2012). Removal of pied crow nests from towers because of their habit of using wire in their nest material is likely (Vosloo 2009; Joseph et al. 2017; NDS pers. obs). As in the case of the hooded crows, this may not deter them permanently, and they soon could nest on towers again.
Like most birds, the breeding activity of pied crows starts at the onset of spring (Chittenden et al. 2016). Our surveys were performed at the height of the breeding season, so we missed most of the courtship, site inspection, and other nesting activities that occurred before (Mundy and Cook 1977). Starting surveys with the onset of breeding activities may increase understanding of tower usage. When we conducted our surveys, we found no breeding pairs activity at any unused towers to suggest potential nesting, so breeding activity was observed only at the used towers.
We found nested towers were not statistically clustered in the study area, with a mean distance of 1595 m between them. Interestingly, based on the distribution of nested towers, unused towers shared close to similar zones as those shown by nested towers. Thus, these results could suggest that nesting on towers may have little to do with the distribution of towers in the study area, with every available tower likely to be used as a nesting site. Mundy and Cook (1977) reported a minimum distance of 200 m between pied crow nests in an urban area of northern Nigeria, in a 3-ha nesting area. So, the spacing between the towers was adequate and could not have been detrimental to the pied crows’ choice of nesting sites when other factors were excluded. In the wild, the pied crows typically nest on isolated trees, giving the impression that they could be territorial (Goodwin et al. 1976). Some corvid species are known to be territorial (Erikstad et al. 1982). When using artificial infrastructure for nesting, pied crows can nest near white-backed vultures (Gyps africanus) and greater kestrels (Falco rupicoloides) (Malan 1994; Johnson and Murn 2019). When nesting on telephone poles and trees next to roads, it has been found that the sequential distribution of pied crow nests can be separated by 1–2 km distance (Malan 1994). In urban settings, the availability of favourable nesting sites and food resources could be key in promoting the persistence of pied crows (Lamm 1958; Mundy and Cook 1977; Campbell 2009; Kopij 2019; Craig et al. 2021; Nyatanga et al. 2021) as is typical for other corvids in urban areas (McIvor et al. 2012; Greggor et al. 2016).
We found that the likelihood of pied crows nesting on towers in the study area was affected by land-use features; it increased with the grassland area but decreased with the availability of schools in a 1 km radius. There was a negative relationship between nesting and foresty and sugarcane. Pied crows are known to avoid forested areas but not their clearings (Jenkins and Underhill 1997). Therefore, the use of towers near forests and sugarcane fields for nesting in the study area may be less but cannot be ruled out. Croplands and subsistence farming were key in determining the probability of nesting. As anticipated, water bodies and subsistence farming positively influenced nesting. Schools, recreational parks, and marketplaces tend to accumulate litter which attracts scavenging birds, especially corvids (Waite 1983). During surveys, pied crows were seen foraging on school playgrounds and in rubbish bins of recreational parks (NDS pers. obs). We anticipated nesting sites to be positively influenced by schools, with the probability of nesting likely to increase with the number of schools within a 1 km radius of towers. However, most towers were not located close to school premises and fell outside the 1 km radius chosen. The wide distribution range of pied crows in Africa could also suggest that the species occupies and breeds in variable altitudes. Our findings indicate that towers located in the lower-lying areas might be preferred for nesting.
Urban roosting and breeding are common in corvid species (Jollie 1976; Houston and Schmutz 1999; McGowan 2001; Benmazouz et al. 2021). In the case of pied crows, roosting sites are relatively difficult to find in urban areas where their numbers can be counted. In this study, one roosting site was discovered near a landfill site (CTD pers. obs.). Pied crow roost sites are usually found in periurban or natural and managed green spaces in urban landscape mosaics (Dean and Milton 2000; Senoge unpublished data). The combined effects of climate change and anthropogenic land-use changes are likely to determine the continued persistence of pied crows in urban mosaic landscapes with natural and /or managed greenspaces and the availability of anthropogenic infrastructure and food (Hof et al. 2011; Downs et al. 2021). Their behavioural plasticity will likely increase their numbers in such landscapes (Downs et al. 2021). The pied crows’ urban presence and persistence could be seen as slow compared with other corvids (Kövér et al. 2015); however, it is likely more because of little documentation (Lamm 1958; Mundy and Cook 1977; Campbell 2009; Kopij 2019; Craig et al. 2021; Nyatanga et al. 2021).
Our findings showed the use of towers as nesting sites by pied crows in the urban mosaic landscape of Pietermaritzburg, with almost half of the towers monitored used for nesting. Although nested towers were not significantly clustered, their spatial distribution was similar to that of unused towers. Our results show that topography of nesting towers, cropland, and forestry could influence the nesting of pied crows. As with recreational parks and refuse collection sites, schools are likely to attract pied crows and potentially influence their breeding behaviour in urban mosaic landscapes. Our results also show that towers located in less built-up areas, such as those characterised by open grasslands, may be preferred for nesting. The height of towers was not considered in this study but allowed for more accessible observations and identification of pied crow nests. Use of exotic trees (e.g., Eucalyptus spp. and coniferous trees) as nest sites by pied crows may be common in urban mosaic landscapes and needs to be investigated. Engagement with locals could also improve locating active nesting sites on trees. Nests were found in greater numbers in some suburbs because of the high number of towers available. Although we found no nesting activity on towers in some suburbs, it was unclear if any previous nesting took place on towers in these locations. Starting in August, regularly monitoring towers may provide more information regarding the use of towers by pied crows in urban mosaic landscapes. Furthermore, it is important to extend the monitoring of breeding pairs starting in early spring until late summer to understand pied crows’ breeding behaviour and movements.
Data Availability
Data for this study are available on reasonable request from the authors but belong to the University of KwaZulu-Natal.
Code Availability
(software application or custom code)
Not applicable.
References
Alexander J, Ehlers Smith DA, Ehlers Smith YC, Downs CT (2019a) Drivers of fine-scale avian functional diversity: an assessment of the effects of eco-estate housing development and management. Landsc Ecol 34:537–549. https://doi.org/10.1007/s10980-019-00786-y
Alexander J, Ehlers Smith DA, Ehlers Smith YC, Downs CT (2019b) Eco-estates, diversity hotspots or isolated developments? Connectivity of eco-estates in the Indian Ocean Coastal Belt, KwaZulu-Natal, South Africa. Ecol Indic 103:425–433. https://doi.org/10.1016/j.ecolind.2019.04.004
Alexander J, Ehlers Smith DA, Ehlers Smith YC, Downs CT (2019c) A multi-taxa functional diversity assessment of the effects of eco-estate development in the mixed land-use mosaic of the KwaZulu-Natal North Coast, South Africa. Landsc Urban Plan 192:103650. https://doi.org/10.1016/j.landurbplan.2019.103650
Altevogt R, Davis TA (1979) Urbanization in nest building of indian house crows Corvus splendens. J Bombay Nat Hist Soc 76:283–290. https://www.biodiversitylibrary.org/part/151544
Baltensperger AP, Mullet TC, Schmid MS, Humphries GRW, Kövér L, Huettmann F (2013) Seasonal observations and machine-learning-based spatial model predictions for the common raven (Corvus corax) in the urban, sub-arctic environment of Fairbanks, Alaska. Polar Biol 36:1587–1599. https://doi.org/10.1007/s00300-013-1376-7
Benmazouz I, Jokimäki J, Lengyel S, Juhász L, Kaisanlahti-Jokimäki M-L, Kardos G, Paládi P, Kövér L (2021) Corvids in urban environments: a systematic global literature review. Animals 11:3226. https://doi.org/10.3390/ani11113226
Burnham KP, Anderson DR, Huyvaert KP (2011) AIC model selection and multimodel inference in behavioural ecology: some background, observations, and comparisons. Behav Ecol Sociobiol 65:23–35
Campbell M (2009) Factors for the presence of avian scavengers in Accra and Kumasi, Ghana. Area 41:341–349
Chittenden H, Davies G, Weiersbye I (2016) Roberts bird guide: illustrating over 1000 bird species in southern Africa (2nd ed). John Voelcker Bird Book Fund, Cape Town
Clancey PA (1964) The birds of Natal and Zululand. Oliver & Boyd, Edinbugh, Scottland
Cover T, Hart P (1967) Nearest neighbor pattern classification. IEEE Trans Inform Theory 13:21–27
Craig AJFK, Hulley PE, Mullins RLG (2021) Urban birds in the Eastern Cape: local observations from Makhanda (Grahamstown) and future questions. Ostrich 92:70–82. https://doi.org/10.2989/00306525.2020.1816585
Cunningham S, Madden C, Barnard P, Amar A (2016) Electric crows: powerlines, climate change and the emergence of a native invader. Divers Distrib 22:17–29
Dean WRJ (2005) Pied crow Corvus albus. In: Hockey PAR, Dean WRJ, Ryan PG (eds) Roberts Birds of Southern Africa. John Voelcker Bird Book Fund, Cape Town
Dean WRJ, Milton S (2000) Directed dispersal of Opuntia species in the Karoo, South Africa: are crows the responsible agents? J Arid Environ 45:305–314. https://doi.org/10.1006/jare.2000.0652
Dean WRJ, Milton S (2003) The importance of roads and road verges for raptors and crows in the succulent and Nama-Karoo, South Africa. Ostrich 74:181–186. https://doi.org/10.2989/00306520309485391
Dean WRJ, Milton S, Anderson M (2006) Use of road kills and roadside vegetation by Pied and Cape crows in semi-arid South Africa. Ostrich 77:102–104. https://doi.org/10.2989/00306520609485516
Downs CT, Alexander J, Brown M, ChibesaM, Ehlers Smith YC, Gumede ST, Hart LA, Kalle R, Maphalala M, Maseko M, McPherson S, Ngcobo SP, Patterson L, Pillay K, Price C, Raji IA, Ramesh T, Schmidt W, Senoge ND, Shivambu TC, Shivambu N, Singh N, Singh P, Streicher J, Thabethe V, Thatcher H, Widdows C, Wilson AL, Zungu MM, Ehlers Smith D (2021) Modification of the third phase in the framework for vertebrate species persistence in connected urban environments based on a review of case studies from KwaZulu-Natal Province, South Africa. Ambio 50:1866–1878. https://doi.org/10.1007/s13280-021-01501-5
Erikstad KE, Blom R, Myrberget S (1982) Territorial hooded crows as predators on willow ptarmigan nests. Wildl Manage 46:109–114. https://www.jstor.org/stable/3808413
Fincham JE, Lambrechts N (2014) How many tortoises do a pair of pied crows Corvus albus need to kill to feed their chicks? Biodivers Observ 5:135–138
Goodwin D, Gillmor R, Museum B (1976) Crows of the world. Cornell University Press, Ithaca, New York
Gorenzel WP, Salmon TP (1995) Characteristics of american crow urban roosts in California. J Wildl Manage 59:638–645. https://doi.org/10.2307/3801939
Graham MH (2003) Confronting multicollinearity in ecological multiple regression. Ecology 84:2809–2815
Greggor AL, Clayton NS, Fulford AJ, Thornton A (2016) Street smart: faster approach towards litter in urban areas by highly neophobic corvids and less fearful birds. Anim Behav 117:123–133. https://doi.org/10.1016/j.anbehav.2016.03.029
Hof C, Levinsky I, Araújo MB, Rahbek C (2011) Rethinking species’ ability to cope with rapid climate change. Glob Change Biol 17:2987–2990. https://doi.org/10.1111/j.1365-2486.2011.02418.x
Houston CS, Schmutz JK (1999) Changes in bird populations on canadian grasslands. Stud Avian Biol 19:87–94
Jenkins A, Underhill L (1997) Pied Crow. In: Harrison JA, Allan DG, Underhill LG, Herremans M, Tree AJ, Parker V, Brown CJ (eds) The atlas of southern african birds, vol 2. Passerines, Johannesburg: BirdLife South Africa, p 104
Jokimäki J, Suhonen J, Vuorisalo T, Kövér L, Kaisanlahti-Jokimäki M-L (2017) Urbanization and nest-site selection of the black-billed magpie (Pica pica) populations in two finnish cities: from a persecuted species to an urban exploiter. Landsc Urban Plan 157:577–585. https://doi.org/10.1016/j.landurbplan.2016.08.001
Johnson TF, Murn C (2019) Interactions between pied crows Corvus albus and breeding whitebacked vultures Gyps africanus. Ethol Ecol Evol 31:240–248. https://doi.org/10.1080/03949370.2018.1561523
Jollie M (1976) Species interrelationships of three corvids. Biologist 58:89–111
Joseph GS, Seymour CL, Foord SH (2017) The effect of infrastructure on the invasion of a generalist predator: Pied crows in southern Africa as a case-study. Biol Conserv 205:11–15. https://doi.org/10.1016/j.biocon.2016.11.026
Jurčević Agić I (2006) Ravens, Corvus corax (L. 1758), nesting on high-voltage transmission line pylons in Croatia. Belg Zool 136:167–171. https://biblio.naturalsciences.be/associated_publications/bjz/bibliographic-references/ISI_000243430400007
Kopij G (2019) Population density and structure of birds breeding in an urban habitat dominated by large baobabs (Adansonia digitata), Northern Namibia. Biosystems Divers 27:354–360. https://doi.org/10.15421/011947
Kövér L, Gyüre P, Balogh P, Huettmann F, Lengyel S, Juhász L (2015) Recent colonization and nest site selection of the hooded crow (Corvus corone cornix L.) in an urban environment. Landsc Urban Plan 133:78–86. https://doi.org/10.1016/j.landurbplan.2014.09.008
Kövér L, Lengyel S, Takenaka M, Kirchmeir A, Uhl F, Miller R, Schwab C (2019) Why do zoos attract crows? A comparative study from Europe and Asia. Ecol Evol 9:14465–14475. https://doi.org/10.1002/ece3.5881
Lamm DW (1958) A nesting study of the pied crow at Accra, Ghana. Ostrich 29:59–70. https://doi.org/10.1080/00306525.1958.9639069
Malan G (1994) Distribution of nests of the pied crow Corvus albus on telephone poles in the upper Karoo. Ostrich 65:44–45. https://doi.org/10.1080/00306525.1994.9632680
Marzluff JM, Boone RB, Cox GW (1994) Historical changes in populations and perceptions of native pest bird species in the West. Stud Avian Biol 15:202–220
Meyer WB, Turner BL. 1992. Human population growth and global land-use/cover change. Ann Rev Ecol System 23:39–61
McGowan KJ (2001) Demographic and behavioral comparisons of suburban and rural american crows. In: Marzluff J, Bowman R, Donnelly R (eds) Avian ecology and conservation in an urbanizing world. Springer, Boston, MA, pp 365–381
McIvor GE, Rowe C, Healy SD (2012) Deterring hooded crows from re-nesting on power poles. Wildl Soc Bull 36:729–734. https://www.jstor.org/stable/wildsocibull2011.36.4.72
McKinney ML (2002) Urbanization, biodiversity, and conversation. Bioscience 52:883–890
Miller JR, Cale P (2000) Behavioral mechanisms and habitat use by birds in a fragmented agricultural landscape. Ecol Appl 10:1732–1748
Mundy P, Cook A (1977) Observations on the breeding of the pied crow and great spotted cuckoo in northern Nigeria. Ostrich 48:72–84. https://doi.org/10.1080/00306525.1977.9634084
Noreen Z, Sultan H (2021) Trees to towers shift: power lines emerging as a novel habitat for birds in Gujranwala, Pakistan. Pure Appl Biol 11:616–628. https://doi.org/10.19045/bspab.2022.110063
Nyatanga T, Ndaimani H, Gaza T (2021) Local variations in the response of birds to human presence in urban areas. Ostrich 92:270–279. https://doi.org/10.2989/00306525.2021.1976858
Patterson L, Kalle R, Downs CT (2018) Factors affecting presence of vervet monkey troops in a suburban matrix in KwaZulu-Natal, South Africa. Landsc Urban Plan 169:220228. https://doi.org/10.1016/j.landurbplan.2017.09.016
Patterson L, Kalle R, Downs CT (2019) Living in the suburbs: space use by vervet monkeys (Chlorocebus pygerythrus) in an eco-estate, South Africa. Afr J Ecol 57:539–551. https://doi.org/10.1111/aje.12629
SABAP2 (2022) http://sabap2.adu.org.za/species
Shochat E, Warren PS, Faeth SH, McIntyre NE, Hope D (2006) From patterns to emerging processes in mechanistic urban ecology. TREE 21:186–191. https://doi.org/10.1016/j.tree.2005.11.019
Singh P, Downs CT (2016a) Hadedas in the hood: Hadeda ibis activity in suburban neighbourhoods of Pietermaritzburg, KwaZulu-Natal, South Africa. Urban Ecosys 19:1283–1129. https://doi.org/10.1007/s11252-016-0540-6
Singh P, Downs CT (2016b) Hadeda ibis (Bostrychia hagedash) urban nesting and roosting sites. Urban Ecosys 19:1295–1305. https://doi.org/10.1007/s11252-016-0541-5
Stine RA (1995) Graphical interpretation of variance inflation factors. Am Stat 49:53–56. https://doi.org/10.1080/00031305.1995.10476113
Thabethe V, Downs CT (2018) Citizen science reveals widespread supplementary feeding of african woolly-necked storks in suburban areas of KwaZulu-Natal, South Africa. Urban Ecosys 21:965–973. https://doi.org/10.1007/s11252-018-0774-6
Thatcher H, Downs CT, Koyama NF (2019a) Anthropogenic influences on the time budgets of urban vervet monkeys. Landsc Urban Plan 181:38–44. https://doi.org/10.1016/j.landurbplan.2018.09.014
Thatcher H, Downs CT, Koyama NF (2019b) Positive and negative interactions with humans concurrently affect vervet monkey (Chlorocebus pygerythrus) ranging behavior. Int J Primatol 40:496–510. https://doi.org/10.1007/s10764-019-00099
Tilman D, May RM, Lehman CL, Nowak MA (1994) Habitat destruction and the extinction debt. Nature 371:65–66. https://doi.org/10.1038/371065a0
Tilman D, Clark M, Williams DR, Kimmel K, Polasky S, Packer C (2017) Future threats to biodiversity and pathways to their prevention. Nature 546:73–81. https://doi.org/10.1038/nature22900
Vosloo H (2009) Bird nesting guidelines. Internal Report for Eskom Transmission, Sunninghill, South Africa
Waite RK (1983) Issues in corvid behaviour and ecology. PhD thesis, Keele University, UK
Walcott CF (1974) Changes in bird life in Cambridge, Massachusetts from 1860 to 1964. Auk 91:151–160. https://www.jstor.org/stable/4084670
Wanless RM, Jupiter TA (2002) Predation on doves by pied crows, Corvus albus, on Aldabra Atoll. Ostrich 73:73. https://doi.org/10.2989/00306520209485361
Widdows C, Downs CT (2015) A genet drive-through: are large spotted genets using urban areas for “fast food”? A dietary analysis. Urban Ecosys 18:907–920. https://doi.org/10.1007/s11252-015-0438-8
Widdows C, Downs CT (2016) Urban roost temperatures of large-spotted-genets: the effect of anthropogenic structures. J Therm Biol 57:66–71. https://doi.org/10.1016/j.jtherbio.2016.02.004
Widdows C, Downs CT (2018) Genets in the city: community observations and perceptions of large spotted genets in an urban environment. Urban Ecosys 21:357–367. https://doi.org/10.1007/s11252-017-0722-x
Widdows C, Ramesh T, Downs CT (2015) Factors affecting the distribution of large spotted genets (Genetta tigrina) in an urban environment in South Africa. Urban Ecosys 18:1401–1413. https://doi.org/10.1007/s11252-015-0449-5
Withey JC, Marzluff JM (2009) Multi-scale use of lands providing anthropogenic resources by american crows in an urbanizing landscape. Landsc Ecol 24:281. https://doi.org/10.1007/s10980-008-9305-9
Acknowledgements
We thank Kerushka Pillay, Manqoba Zungu and Vuyisile Thabethe for their support and contributions. We are most grateful to the reviewers for their comments that improved our manuscript.
Funding
Open access funding provided by University of KwaZulu-Natal. We are thankful to the University of KwaZulu-Natal (ZA) and the National Research Foundation (ZA, Grant 98404) for funding and making this study possible. We thank the Ford Wildlife Foundation (ZA) for vehicle support.
Author information
Authors and Affiliations
Contributions
NDS and CTD conceptualised the study. CTD sought funding. NDS collected and analysed data. NDS drafted the manuscript. CTD edited the draft. All authors approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval
No ethics approval was required.
Conflict of Interest
The authors declare no conflict of interest nor competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Senoge, N.D., Downs, C.T. The use of cellular telecommunication towers as nesting sites by pied crows (Corvus albus) in an urban mosaic landscape. Urban Ecosyst 26, 881–892 (2023). https://doi.org/10.1007/s11252-023-01342-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01342-y