
Abstract
Human settlements, including cities, may provide wildlife with new ecological niches, in terms of habitat types and food availability, thus requiring plasticity for adaptation. The crested porcupine Hystrix cristata is a habitat-generalist, large-sized rodent, also recorded in some suburban areas, but no information is available on its habitat use in metropolitan landscapes. Here, we assessed the land-use factors influencing the presence of crested porcupines in a metropolitan area of Central Italy. We collected data on the occurrence of crested porcupines from the metropolitan area of Rome, following an observer-oriented approach to record occurrences and retreive pseudo-absences. We then related the presence/absence of H. cristata to landscape composition. Occupancy models showed that cultivations and scrubland were positively related to porcupine presence, most likely as they provide food resources and shelter sites, respectively. Although the crested porcupine has been confirmed as a “generalist” species in terms of habitat selection, a strong preference for areas limiting the risk of being killed and providing enough food and shelter was observed. We therefore suggest that the crested porcupine may adapt to deeply modified landscapes such as large cities by selecting specific favourable land-use types.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Urbanisation is one of the main land-use modifications occurring at large scales globally, as human settlements are encroaching into rural areas and natural habitats (Ditchkoff et al. 2006; McKinney 2006). Behavioural flexibility and generalist ecological niche has helped mammal species to thrive and form self-sustaining populations in urban areas, with a process called synurbisation (Santini et al. 2019). Besides species that only occasionally cross cities (“urban-visitors” and “urban-explorers”), several taxa, usually defined as “urban-dwellers” or “urban-adapters”, thrive and successfully exploit urban environments (Baker et al. 2003; Grimm et al. 2008; Bateman and Fleming 2012; Balestrieri et al. 2016; Uchida et al. 2021). Synurbisation may pose a challenge to wildlife managers, as there is a strong need to find a trade-off between limiting human-wildlife conflicts and movements of animal-right groups (Don Carlos et al. 2009; La Morgia et al. 2017; Honda et al. 2018). Moreover, synurbic species may be relatively rare or legally protected taxa, i.e. representing a further management issue. Thus, analysing the spatial ecology of species of conservation concern and species generating human-wildlife conflict in urban ecosystems is crucial for their long-term management (Gehrt et al. 2009; Ancillotto et al. 2016; Cronk and Pillay 2021).
A pool of native and introduced species has benefited by the expansion of human settlements, which may provide increased food availability and shelter sites, as well as decreased predation risk (Sever and Mendelssohn 1989; Contesse et al. 2004; Marks and Bloomfield 2006). Wild mammals living in or around urban areas may exhibit different traits as their rural counterparts because of a different predation pressure or adaptation to human-induced stresses (Ditchkoff et al. 2006; Santini et al. 2019). As to mammals, about 3.5% known species worldwide is regularly recorded in urban areas, with a peak of species in Southern and Central Europe, i.e. areas with a long history of deep landscape modifications. Carnivores and rodents are the most represented mammalian orders in urban environments (Santini et al. 2019). The crested porcupine Hystrix cristata is one of the largest rodents inhabiting urban areas, and has been recorded in at least 10 cities within its range of introduction (Grano 2016; Lovari et al. 2017; Santini et al. 2019; Manenti et al. 2020). This species is generally described as a “habitat generalist” as it can exploit a wide range of habitat types from woodland to farmlands, despite being linked to covered habitats (e.g. woodland and scrubland) for denning (Mohr 1965; Sonnino 1998; Monetti et al. 2005; Mori et al. 2014a; Lovari et al. 2017). Porcupines are widely poached in Central and Southern Italy, mainly because they are considered as crop raiders, even though it has been shown that most damage only occurs in small unprotected vegetable gardens (Ghigi 1917; Laurenzi et al. 2016; Lovari et al. 2017). In suburban areas, where poaching pressure might be the highest (Lovari et al. 2017), the crested porcupine mostly select thorny thickets for denning and feed mostly on fruits (Lovari et al. 2017). In the suburbs, farmlands and fallows are avoided by porcupines, whereas no data are available on the ecology of this large rodent in metropolitan areas.
The aim of this study is assessing the spatial ecology of the crested porcupine population in Rome (Italy), which currently is the only European metropolitan area hosting a self-sustaining population of crested porcupine (Grano 2016; Santini et al. 2019). Specifically, we ran occupancy models to evaluate how landscape composition affects the presence of the crested porcupine within the urban area, using occurrence data collected over a 16 years time window, i.e. since the first available records of this species in Rome. We predicted that porcupines, as being generalist rodents, would prefer areas providing the best trade-off between food availability and predation risk, thus selecting urban environments with cultivated (i.e. exploited for foraging) and vegetation-covered areas (exploited for shelter). Our results also provide information on how limiting human-porcupine conflict (cf. Cerri et al. 2017; Lovari et al. 2017), by showing the environmental preferences of this urban-dwelling generalist species in a metropolitan area, where contacts between this species and humans are the highest.
Materials and methods
Study area
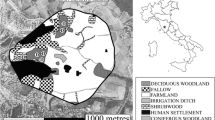
We focused on the area included in the Rome beltway (“Grande Raccordo Anulare” ring highway, hereafter GRA), i.e. within the closed ring highway that embraces the metropolitan area of Rome without discontinuity, separating the city from countryside and periphery (average diameter: 21 km; total length: 68 km). The GRA entails an area of 46,000 ha within the municipal territory of Rome. Rome represents the Italian metropolis with the highest density of green public areas (urban parks and cemeteries), i.e. the 5.1% of the study area. Rome is also the third Italian municipality for the surface of agricultural land (after Florence and Bari), with cultivations (orchards, vineyards, olive groves, arable lands, and horticultural crops) covering the 29.9% of our study area (www.istat.it, accessed on 01.02.2021). Woodlands and scrublands cover respectively 3.8% and 2.5% of the study area. The remaining part of the study area is covered with human settlements (58.4%) and archaeological areas (0.3%) (Fig. 1).

Study area, land-cover typologies, and occurrence records used for analyses. Outside the frame we report the UTM 32 coordinates of the study area; the grid used for analyses (1 km) is also shown.
Rome is a rich city in animal biodiversity (e.g. Zapparoli 1997; Capotorti et al. 2019), hosting at least 39 mammal species, including medium to large-sized ones such as the wild boar Sus scrofa, the red fox Vulpes vulpes, the stone marten Martes foina and the crested porcupine (Amori et al. 2009; Todini and Crosti 2020). The latter is known to occur in the study area at least since the early XX century (Lepri 1911; Miller 1912), and it was confirmed continuously since the early 2000s (Gippoliti and Amori 2006; Grano 2016), thus suggesting that the population within the study area is relatively stable.
Data collection and validation
Occurrence data of the crested porcupine used in this study were collected between April 2005 and December 2020, and uploaded on Ornitho and iNaturalist citizen science platforms by one of the authors (LA). We used this time span, as the stable presence of the crested porcupine within the Rome mentropolitan area was only confirmed in the early 2000s, despite occasional records since the early XX Century (Gippoliti and Amori 2006; Amori et al. 2009). All data were collected and/or verified year by year by LA during the 16 year period during extensive field-work throughout the metropolitan area to describe and monitor the local mammal diversity. Occurrences were then uploaded by the same author after 2017 for the Italian Mammal Atlas project (https://www.inaturalist.org/observations?project_id=mammiferi-d-italia). Data were validated through an expert-based process, as described in the regulation of this platform. Most observations were represented by quills, dead animals, or footprints. Also, faecal pellets of the crested porcupine are easily identifiable, showing an unmistakable oblong and curved-olive shape (Mori et al. 2021). All data were georeferenced with a < 10 m accuracy, and entered into a database.
Occupancy models
The study area was divided into 460 cells of 1 km² each using QGIS (ver. 3.16.1: QGIS Development Team 2019). The average home range size of the crested porcupine in Mediterranean environments is ~ 0.5 km² (Lovari et al. 2013; Mori et al. 2014a); thus the cell size we adopted limits the risk of pseudoreplication (i.e. the same individual detected in multiple cells).
We used a static (single-season) occupancy model as most of the sites were monitored in one single sampling year. Following Milanesi et al. (2020), pseudo-absences were assigned to records of all the species detected by the same observer, excluding porcupine records. For cells monitored during more than one year, the year with the largest number of records of any species was selected for analyses. We considered the percentage of cover of seven habitat typologies describing land-use cover as covariates of occupancy: human settlements, scrubland, archaeological areas, woodland, urban green areas, cultivations and fallows. These habitat typologies were obtained by reclassifying the land-use map of the Latium Region of the year 2016 (original land-use map available at https://dati.lazio.it/). For each study site (1 km cell), we extracted the percentage cover of each habitat typology.
Statistical analyses were performed in the software R 3.5.1 (R Core Team 2013). We used the unmarked and MuMIN packages respectively to conduct occupancy models and to select the best models (Fiske and Chandler 2011; Bartoń 2018). Animal species are rarely observed with perfect accuracy, particularly when nocturnal and elusive as the crested porcupine (Corsini et al. 1995; Lovari et al. 2017). Occupancy models allow to estimate species distribution, and to evaluate relationships between occupancy and environmental features, taking into account possibility that the target species remained undetected during sampling (MacKenzie et al. 2003), and are thus particularly important for the analysis of data collected through citizen science campaigns (Altwegg and Nichols 2019; Marta et al. 2019). Occupancy models allow estimating the detection probability of species based on a series of detection / non-detection data at fixed sites (Kéry et al. 2013). This approach thus requires information on non-detections, i.e. on surveys during which the target species was not detected. These data are not easy to obtain from citizen science datasets (MacKenzie et al. 2003; Altwegg and Nichols 2019). In our study pseudo-absences were estimated through an “observer-oriented” approach (Milanesi et al. 2020), i.e. by considering records of species other than the target one. Specifically, we assumed that a cell was surveyed in a specific date if the database included at least one record of a species, recorded by an observer that has detected target species (Milanesi et al. 2020) as a valuable method to inform species distribution models (Milanesi et al. 2020). Multiple records collected having the same date and the same cell were considered to be one single observation.
We related detection probability to the date of the survey (day of the year), also considering a quadratic term. Pearson’s correlation coefficient was used to test for independence between pairs of covariates: pairs of covariates with r > |0.70| were considered as strongly correlated; thus, only one covariate (i.e. the most important one for the porcupine: Mori et al. 2014a) of the pair was included in the model. The cover of human settlements was excluded from the models as being significantly and negatively correlated with cultivations (r = -0.80), which is an important habitat type for the crested porcupine (Mori et al. 2014a). We ran a total of 256 competing models, considering each possible combination of independent variables on detection and occupancy. Models were ranked through the Akaike’s Information Criterion corrected for small samples (AICc): the best model was the one with the lowest AICc (Burnham and Anderson 2004). Models with an AICc difference < 2 were considered equally supported. We estimated the significance of the variables included in the best model through a likelihood ratio test, considering as significant variables when p < 0.05.
Results
Between April 2005 and December 2020, our survey provided records from 129 cells. The presence of the crested porcupine was recorded in 65 out of 129 sampled cells, with a total of 100 detections of crested porcupine (Fig. 1). The best-occupancy model suggested that the occupancy of the crested porcupine was positively and significantly related to the cover of scrubland and cultivations (Tables 1 and 2). The second best model included only the cover of cultivations and had a difference of AICc of 2.04 (Table 1), hence it was not considered. None of the other covariates was included in a model with high support, on the basis of AICc values. The detection probability of the porcupine was unrelated to the date of survey. The average detection probability per survey was 0.89 (95% confidence interval: 0.74–0.96).
Discussion
In our work, we showed that the probability of occurrence of the crested porcupine in our urban ecosystem was positively correlated to coverage of cultivations and scrublands. The crested porcupine is a monogamous species who pair for life and show a sedentary behaviour after dispersal and settlement in a territory (Mori et al. 2016); therefore, the movements of adults are mostly determined by food search and not by mate search (Lovari et al. 2013; Mazzamuto et al. 2019). This may provide support to the importance of cultivations within urban environments, which provide porcupines with clumped and abundant food resources (Lovari et al. 2017), even within human settlements. Scrublands are mostly selected as den sites, particularly where human pressure (i.e. poaching risk) is highest, which may support the use of this habitat types in dense human settlements (Tinelli and Tinelli 1980; Monetti et al. 2005; Lovari et al. 2017). Accordingly, within the Rome urban area, the few dens of crested porcupines whose location is known, occur in one archaeological area (“Catacombe di Priscilla”, n = 1; Grano 2016), as well as in a semi-natural scrubland area within a protected natural reserve (n = 2), and in a densely vegetated area of a large recreational park (n = 1; Ancillotto L. pers. obs.). Therefore, results support the prediction that the crested porcupine in a metropolitan area would select habitats providing a favourable trade-off between food abundance and shelter availability.
In natural habitats, the crested porcupine mainly feeds on underground storage organs of plants (e.g. bulbs, tubers and rhizomes), but can also eat fruits and vegetables (Bruno and Riccardi 1995; Mori et al. 2020). Crested porcupines may exploit nearby cultivated areas which provide easy access to food resources (e.g. figs and pumpkins) and do not require time-consuming active excavation which may limit vigilance (Lovari et al. 2017). Most likely, the positive correlation between cultivations and occupancy in the urban environment could also be explained by limited food resources in some areas of the city (e.g. recreational parks characterised by deeply modified floras), thus leading porcupine individuals to expose themselves to open and, therefore, risky habitats. Accordingly, few data occurred in recreational areas, possibly underused to limit encounters with humans and potential natural predators, e.g. red foxes and domestic dogs, which are often abundant in urban parks in Rome (Amori et al. 2009). Local high densities of wild boar (Todini and Crosti 2020) may also limit the occupancy by the crested porcupines in some areas (Mazzamuto et al. 2019). Furthermore, artificial lights at night may limit the use of these areas by crested porcupines, which are known to avoid bright areas and bright moonlight nights (Mori et al. 2014b). In fact, it is much more likely that these areas, as well as archaeological sites, wetlands, and human settlements, are avoided as not providing sufficient food resources. Therefore, all these habitats are avoided by porcupines also in natural contexts, yet some individuals, especially sub-adults, may visit them occasionally and create temporary or seasonal burrows (Pigozzi and Patterson 1990; Börger 2002; Mori and Assandri 2019).
Noise pollution and vehicular traffic have also been reported to alter the spatial behaviour of the porcupine (Mori et al. 2013; Mori 2017), and long-distance roads are known to hinder wildlife movements (Forman and Alexander 1998; Seidler et al. 2015). Accordingly, records of crested porcupines in Italian urban areas increase when vehicular traffic (and related human pressure) is the lowest (e.g. during the lockdown following the SARS-CoV 2 pandemic outbreak: Manenti et al. 2020). In this context, the ring highway in Rome may represent a barrier to the transit of porcupines, as green corridors between green areas inside and outside the city centre are few.
Crop damages by crested porcupines may occur in areas covered by cultivations, including vegetable gardens, potentially triggering conflict with humans (Sforzi et al. 1999; Laurenzi et al. 2016). No conflict between humans and porcupines has been reported in Rome to date though; strictly nocturnal habits and local protection of cultivations through fences may have in fact promoted coexistence between porcupines and humans in urban and suburban areas (Lovari et al. 2017), as evidenced in the closely related Indian crested porcupine Hystrix indica in Israel (Sever and Mendelssohn 1989). However, the intense illumination of the highway between Haifa and Tel-Aviv (National Road 2, Israel) did not prevent Indian porcupines from foraging in the nearby of the roadside, contrary to what expected from a nocturnal species that avoids brightest nights (Sever and Mendelssohn 1989).
Our results highlight that a large generalist rodent such as H. cristata may become an actual urban-dwelling species (cf. Santini et al. 2019), by occupying spots of suitable habitats, namely represented by patches of natural or agricultural areas, eventually persisting in one of the largest metropolitan areas in southern Europe. The behavioural and physiological mechanisms that allow such persistence without eliciting conflicts, as well as whether urban populations exhibit gene flow with nearby non-urban ones, is still to be cleared. Thus, the urban population of H. cristata in Rome provides a suitable study system to furtherly shed light on mechanisms and consequences of synurbization in mammals.
Data Availability
All data are available on iNaturalist platform (www.inaturalist.org).
References
Altwegg R, Nichols JD (2019) Occupancy models for citizen-science data. Methods Ecol Evol 10:8–21
Amori G, Battisti C, De Felici S (2009) I Mammiferi della provincia di Roma – dallo stato delle conoscenze alla gestione e conservazione delle specie. Provincia di Roma – Assessorato alle Politiche dell’Agricoltura, Stilgrafica Editions, Rome, Italy
Ancillotto L, Santini L, Ranc N, Maiorano L, Russo D (2016) Extraordinary range expansion in a common bat: the potential roles of climate change and urbanization. Sci Nat 103:15
Baker PJ, Ansell RJ, Dodds PAA, Webber CE, Harris S (2003) Factors affecting the distribution of small mammals in an urban area. Mammal Rev 33:95–100
Balestrieri A, Bogliani G, Boano G, Ruiz-González A, Saino N, Costa S, Milanesi P (2016) Modelling the distribution of forest-dependent species in human-dominated landscapes: patterns for the pine marten in intensively cultivated lowlands. PLoS ONE 11:e0158203
Bartoń K (2018) MuMIn: Multi-Model Inference. R Package version 1.42.1. http://r-forge.r-project.org/projects/mumin/. Accessed on 31.01.2021
Bateman PW, Fleming PA (2012) Big city life: carnivores in urban environments. J Zool 287:1–23
Börger L (2002) Uso dello spazio nell’istrice (Hystrix cristata L., 1758) in un’area costiera Mediterraneo. MSc Dissertation in Scienze Biologiche, Università degli Studi di Pisa, Pisa, Italy-
Bruno E, Riccardi C (1995) The diet of the crested porcupine Hystrix cristata L., 1758 in a Mediterranean area. Zeit SäugetierK 60:226–236
Burnham KP, Anderson DR (2004) Multimodel inference: understanding AIC and BIC in model selection. Sociol Meth Res 33:261–304
Capotorti G, Ortí MMA, Copiz R, Fusaro L, Mollo B, Salvatori E, Zavattero L (2019) Biodiversity and ecosystem services in urban green infrastructure planning: A case study from the metropolitan area of Rome (Italy). Urban For Urban Greening 37:87–96
Cerri J, Mori E, Vivarelli M, Zaccaroni M (2017) Are wildlife value orientations useful tools to explain tolerance and illegal killing of wildlife by farmers in response to crop damage? Eur J Wildl Res 63:70
Contesse P, Hegglin D, Gloor S, Bontadina F, Deplazes P (2004) The diet of urban foxes (Vulpes vulpes) and the availability of anthropogenic food in the city of Zurich, Switzerland. Mammal Biol 69:81–95
Corsini MT, Lovari S, Sonnino S (1995) Temporal activity patterns of crested porcupines. Hystrix cristata J Zool (Lond) 236:43–54
Cronk NE, Pillay N (2021) Home range and use of residential gardens by yellow mongoose Cynictis penicillata in an urban environment. Urban Ecosyst 24:127–139
Ditchkoff SS, Saalfeld ST, Gibson CJ (2006) Animal behavior in urban ecosystems: modifications due to human-induced stress. Urban Ecosyst 9:5–12
Don Carlos AW, Bright AD, Teel TL, Vaske JJ (2009) Human–black bear conflict in urban areas: an integrated approach to management response. Hum Dim Wildl 14:174–184
Fiske IJ, Chandler RB (2011) Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J Stat Soft 43:1–23
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29:207–231
Gehrt SD, Anchor C, White LA (2009) Home range and landscape use of coyotes in a metropolitan landscape: conflict or coexistence? J Mammal 90:1045–1057
Ghigi A (1917) I mammiferi d’Italia considerati nei loro rapporto con l’agricoltura. Natura Milano 8:85–137
Gippoliti S, Amori G (2006) Historical data on non-volant mammals in Rome: What do they say about urban environment. Aldrovandia 2:69–72
Grano M (2016) An unusual urban refuge for the crested porcupine, Hystrix cristata, (Linnaeus, 1758) (Mammalia Rodentia): the ancient Catacombs of Priscilla in Rome (Italy). Biodiv J 7: 345–346
Grimm NB, Faeth DH, Golubiewski NE, Redman CL, Wu J, Bai X, Briggs JM (2008) Global change and the ecology of cities. Science 319:756–760
Honda T, Iijima H, Tsuboi J, Uchida K (2018) A review of urban wildlife management from the animal personality perspective: the case of urban deer. Sci Tot Environm 644:576–582
Kéry M, Guillera-Arroita G, Lahoz-Monfort JJ (2013) Analysing and mapping species range dynamics using occupancy models. J Biogeogr 40:1463–1474
La Morgia V, Paoloni D, Genovesi P (2017) Eradicating the grey squirrel Sciurus carolinensis from urban areas: an innovative decision-making approach based on lessons learnt in Italy. Pest Manage Sci 73:354–363
Laurenzi A, Bodino N, Mori E (2016) Much ado about nothing: assessing the impact of a problematic rodent on agriculture and native trees. Mammal Res 61:65–72
Lepri G (1911) Aggiunte alle ricerche faunistiche e sistematiche sui Mammiferi d’Italia che formano oggetto di caccia. Boll Soc Zool Ital 12:241–250
Lovari S, Sforzi A, Mori E (2013) Habitat richness affects home range size in a monogamous large rodent. Behav Processes 99:42–46
Lovari S, Corsini MT, Guazzini B, Romeo G, Mori E (2017) Suburban ecology of the crested porcupine in a heavily poached area: a global approach. Eur J Wildl Res 63:10
MacKenzie DI, Nichols JD, Hines JE, Knutson MG, Franklin AB (2003) Estimating site occupancy, colonization and local extinction when a species is detected imperfectly. Ecol 84:2200–2207
Marks CA, Bloomfield TE (2006) Home-range size and selection of natal den and diurnal shelter sites by urban red foxes (Vulpes vulpes) in Melbourne. Wildl Res 33:339–347
Manenti R, Mori E, Di Canio V, Mercurio S, Picone M, Caffi M, Brambilla M, Ficetola GF, Rubolini D (2020) The good, the bad and the ugly of COVID-19 lockdown effects on wildlife conservation: insights from the first European locked down country. Biol Conserv 249:108728
Marta S, Lacasella F, Romano A, Ficetola GF (2019) Cost-effective spatial sampling designs for field surveys of species distribution. Biodiv Conserv 28:2891–2908
Mazzamuto MV, Lo Valvo M, Anile S (2019) The value of by-catch data: how species-specific surveys can serve non-target species. Eur J Wildl Res 65:1–9
McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biol Cons 127:247–260
Milanesi P, Mori E, Menchetti M (2020) Observer-oriented approach improves species distribution models from citizen science data. Ecol Evol 10:12104–12114
Miller GS (1912) Catalogue of the Mammals of Western Europe (Europe exclusive of Russia) in the collection of the British Museum. British Museum, London, p 1019
Mohr E (1965) Altweltliche Stachelschweine. Ziemsen Verlag Publisher (Eds.), Wittenburg Lutherstadt, Germany
Monetti L, Massolo A, Sforzi A, Lovari S (2005) Site selection and fidelity by crested porcupines for denning. Ethol Ecol Evol 17:149–159
Mori E, Assandri G (2019) Coming back home: recolonisation of abandoned dens by crested porcupine Hystrix cristata and European badgers Meles meles after wood-cutting and riparian vegetation mowing events. Hystrix 30:39–43
Mori E, Corsini B, Mazza G, Menchetti M (2013) From the shores to the cities: road mortality of the crested porcupine in Southern Tuscany. In: Bertolino S, Capizzi D, Mori E, Colangelo P, Scaravelli D (2013). Secondo convegno Italiano sui Piccoli Mammiferi – Libro dei Riassunti: 41
Mori E (2017) Porcupines in the landscape of fear: effect of hunting with dogs on the behaviour of a non-target species. Mammal Res 62:251–258
Mori E, Lovari S, Sforzi A, Romeo G, Pisani C, Massolo A, Fattorini L (2014a) Patterns of spatial overlap in a monogamous large rodent, the crested porcupine. Behav Proc 107:112–118
Mori E, Nourisson DH, Lovari S, Romeo G, Sforzi A (2014b) Self-defence may not be enough: moonlight avoidance in a large, spiny rodent. J Zool (Lond) 294:31–40
Mori E, Menchetti M, Lucherini M, Sforzi A, Lovari S (2016) Timing of reproduction and paternal cares in the crested porcupine. Mammal Biol 81:345–349
Mori E, Di Gregorio M, Mazza G, Ficetola GF (2020) Seasonal consumption of insects by the crested porcupine in Central Italy. Mammalia 85:231–235
Pigozzi G, Patterson IJ (1990) Movements and diet of crested porcupines in the Maremma National Park, central Italy. Acta Theriol 35:173–180
QGIS Development Team (2019) QGIS Geographic Information System. Open Source Geospatial Foundation Project. http://qgis.osgeo.org. Accessed on 01.02.2021
R Core Team (2013) R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing ISBN 3-900051-07-0. http://www.Rproject.org/ Accessed on 28.01.2021
Santini L, Gonzàlez-Suàrez M, Russo D, Gonzalez-Voyer A, von Hardenberg A, Ancillotto L (2019) One strategy does not fit at all: determinants of urban adaptations in mammals. Ecol Lett 22:365–376
Seidler RG, Long RA, Berger J (2015) Identifying impediments to long-distance mammal migrations. Conserv Biol 29:99–109
Sever Z, Mendelssohn H (1989) Porcupines on the edge of town. Isr Land Nat 13:112–115
Sforzi A, Massolo A, Bozzi R, Lovari S(1999) L’istrice in provincia di Grosseto: aspetti biologici e gestionali. Technical Report to the Provincial Council of Grosseto, Grosseto, Italy
Sonnino S (1998) Spatial activity and habitat use of crested porcupine, Hystrix cristata L., 1758 (Rodentia, Hystricidae) in central Italy. Mammalia 62:175–189
Tinelli A, Tinelli P(1980) Le tane di istrice e di tasso. Censimento e densità delle tane nella Riserva Presidenziale di Castelporziano per la conservazione dell’istrice e del tasso. In: Segretariato Generale della Presidenza della Repubblica. Technical Report, Tenuta di Castelporziano, Roma, Italy
Todini A, Crosti R (2020) Il cinghiale (Sus scrofa) come determinante di cambiamenti di vegetazione in una foresta urbana mediterranea: impatto sulla biodiversità di un’area protetta. Forest@ 17:71
Uchida K, Yamazaki T, Ohkubo Y, Yanagawa H (2021) Do green park characteristics influence human-wildlife distance in arboreal squirrels? Urban For Urban Greening 58:126952
Zapparoli M (1997) Urban development and insect biodiversity of the Rome area, Italy. Landsc Urb Planning 38:77–86
Acknowledgements
Authors would like to thank Flavio Rocchi and Mattia Menchetti who provided some information (verified by Leonardo Ancillotto) on the presence of the crested porcupine in Rome. We are grateful to Charles Nilon for useful comments to the manuscript.
Funding
The study was supported by no external fund.
Author information
Authors and Affiliations
Contributions
EM and LA conceived the research, LA collected data on the field, MF, RM and GFF ran the analyses; EM wrote the first draft of the manuscript. All authors contributed to and approved the final draft.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study did not involve human subjects or animal manipulation or maintenance in captivity or laboratory.
Consent for publication
All authors agree and consent with the publication of the study.
Competing interests
Authors declare that they have no competing interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Mori, E., Molteni, R., Ancillotto, L. et al. Spatial ecology of crested porcupine in a metropolitan landscape. Urban Ecosyst 25, 1797–1803 (2022). https://doi.org/10.1007/s11252-022-01264-1
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-022-01264-1