
Abstract
This study aimed to detect the phenotypic differences between the brown (BB) and white (WW) feathered quails and their reciprocal crosses (BW and WB) over two successive generations. The WW and cross quails, especially the BW, had the heaviest body weights, throughout the studied period, with significant variations between the two studied generations (P<0.05). Moreover, the WW and BW possessed the largest egg production during the F1, while in the F2, the BB had superiority among the studied quails with a prominent superiority of the F2 over the F1 (P<0.05). However, the F1 had higher egg weights than F2 with superiority of WW quails compared to the others (P<0.05). Also, the WW quails had the lowest lipid contents of the eggs. These phenotypic variations among the studied quails might be preliminarily explained by the results of the analyzed microsatellite markers despite the few markers used. The high variability among the BW and WB quails might be due to the larger number of alleles (NA and Ne) and the lower values of FIS with low heterozygosity levels (HO and He). Moreover, the BW and BB were the closest, while WB and WW were the farthest because of the high and low genetic identities and the high and low genetic distance between them, respectively. So the obtained results might introduce an initial scientific basis for evaluating and employing the genetic properties of BB, WW, BW, and WB quails in further genetic improvement program, and more microsatellite markers are recommended.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Japanese quail (Coturnix japonica) is a small promising poultry species reared mainly for meat and egg production, especially in rural low-resource areas. It is characterized by distinct productive and reproductive characteristics including rapid growth, short generation interval, early sexual maturation, stress and disease resistance, and a high rate of laying eggs (Farajiarough et al. 2018).
As a result of plumage color mutations, there are many well-known varieties of Japanese quails such as gray-, white-, and brown-colored-feathered quails (Bagh et al. 2016; ELSaidy et al. 2021). These varieties have distinguished productive and reproductive characteristics (Ibrahim et al. 2021). Accordingly, quail varieties are divided into meat-producing, egg-producing, or dual-purpose quails (Ibrahim et al. 2021). Because of the close breeding within the same population, the Japanese quail is very susceptible to inbreeding depression, so crossbreeding between the distant quail varieties is effective to restore the genetic diversity in quail populations (Nunome et al. 2020). Consequently, it is crucial to assess the physiological and genetic variations among these varieties to establish efficient breeding strategies to improve their production by providing the needed information about the most effective genotypes (Dávila et al. 2009).
Microsatellite markers or simple sequence repeats (SSRs) are good genetic markers that can be effectively used to study the genetic variance within or among breeds (Nunome et al. 2020). They are characterized by the presence of 1–6 base repeats in the eukaryotic genome including birds (Kayang et al. 2000), abundant polymorphism, codominance inheritance, and easy scoring by the polymerase chain reaction (PCR) (Kayang et al. 2002). In quail, microsatellites can be used to establish quail genetic maps. In this regard, Kayang et al. 2004 (Kayang et al. 2004) established the first generation of microsatellite markers linkage map which contains 72 microsatellite markers. Additionally, Miwa et al. (2005) fixed the position of quail functional genes and QTLs depending on three microsatellite primers: GUJ0071, GUJ0097, and GUJ0061 (Miwa et al. 2005). Moreover, microsatellites can be effectively used to analyze the genetic diversity among different quail populations through polymorphism analysis (Bai et al. 2018).
Despite that there are many studies that have reported the variation in the productive and reproductive traits among different quail varieties, they only documented this at the phenotypic level, and only the genetic parameters have been calculated. Besides, to the best of our knowledge, only one study (Ibrahim et al. 2021) has involved the genetic diversity analysis depending on microsatellite markers, and this was done on different quail varieties. Thus, the current study aimed to illustrate the physiological variations in two varieties of Japanese quails (brown-feathered quail strain (BB), white-feathered quail strain (WW), and their reciprocal crosses (BW×BW and WB×WB). Besides, we tried to provide a preliminary explanation and understanding of the genetic bases of these variations using some microsatellite markers.
Material and methods
Bird management
The current study was conducted on four populations of Japanese quail (Coturnix japonica): brown-feathered (BB) and white-feathered quail (WW) varieties and their reciprocal crosses (BW×BW and WB×WB). The bird management was done following the regulations of the animal care and ethics committee of the Poultry Production Department, Faculty of Agriculture, Kafrelsheikh University, Egypt. The study continued for two generations after establishing the base population. The number of chicks for each population and the number of selected groups during the laying period in the two generations are listed in Table 1.
One-day-old chicks were wing-banded and weighed before being moved to heated brooders where they were reared on the floor in a conventional open-sided house until they reached 6 weeks old. Thereafter, they were individually housed in cages (depth: 15 cm; width: 18 cm; height: 18 cm).
The house was equipped with electric and gas heaters where the temperature was kept at 35 °C through the first 3 days and 32 °C during the next 4 days and step by step decreased by 2 °C per week until it was kept at 24 °C. Birds received a 24-h lighting photoperiod for the first 2 weeks depending on the natural and artificial light, followed by a 16 h:8 h light-to-dark cycle. Food and water were offered ad libitum. Birds were fed a standard diet of 28% crude protein (CP), 3100 kcal metabolizable energy (ME) kg−1 from 0 to 2 weeks, followed by a grower diet (24% CP and 2900 kcal ME kg−1) from 4 to 6 weeks of age. During the laying period, birds received a layer diet of 20% CP and 2700 kcal ME.
Phenotypic traits
Body weight (BW) was recorded weekly from the beginning to 6 weeks old and then at 12 and 15 weeks old. The eggs (EN) laid from the 6th week until 15 weeks old were individually recorded. Egg weight (EW) was also individually recorded to the nearest 0.1 g for 3 days/week for each hen, and the average egg weight was determined per hen.
To assess egg quality, egg weight, albumen height, albumen width, albumen weight, yolk width, yolk height, yolk color, yolk weight, the Haugh unit (HU), and eggshell thickness were recorded individually, where yolk and albumen height were recorded using a tripod micrometer (mm). To evaluate the egg shape index, two axes, transversal and longitudinal of the egg, were measured using the Vernier caliper, and then, the egg shape index was calculated depending on the following formula: \(\textrm{Egg}\ \textrm{shape}\ \textrm{index}=\frac{\textrm{transversal}\ \textrm{axis}}{\textrm{longitudinal}\ \textrm{axis}}\times 100\). Albumen and yolk indices were calculated from their height and width measurements using the following formula:
At 47 days old, through the F1 and F2, 20 males and 20 females from each quail population were randomly selected and euthanized by cervical dislocation after 12 h fasting according to ELSaidy et al. (ELSaidy et al. 2021) to determine carcass traits including weight after slaughtering, carcass weight (empty carcass), and relative organ weights (%), including the liver, gizzard, heart, intestine, abdominal fat, ovary, testicle, and oviduct.
Biochemical analysis
At the end of the second generation (F2), blood samples (n=20/group) were collected from the wing vein into tubes without anticoagulant, allowed to coagulate at room temperature, and then centrifuged at 3000×g for 10 min to separate the serum, which was stored at −20 °C for further analysis. Total cholesterol, creatinine, total antioxidant, lipid peroxide (malondialdehyde), albumin, and total lipid concentrations were colorimetrically determined using an automatic analyzer (ChemWell®, Megazyme, Wicklow, Ireland) and commercial kits (Labtest Diagnóstica S. A, Lagoa Santa, Brazil).
Microsatellite markers isolation and genotyping
The analysis was carried out at the Animal Production Biotechnology Laboratory, Central Lab Network, National Research Center, Dokki, Egypt. A volume of 3 mL of whole blood was collected from the wing veins of 20 individuals from each quail population (BB = B×B, WW = W×W, BW = BW×BW, and WB = WB×WB). Blood samples were collected in sterile tubes containing 0.5 mL EDTA as an anticoagulant and stored at −20 °C until DNA extraction. Genomic DNA was extracted using GeneJET Whole Blood Genomic DNA Purification Mini Kit (Thermo Fisher Scientific) according to the manufacturer’s instructions. The integrity of DNA was electrophoretically evaluated using 1% agarose gel stained with ethidium bromide (0.5 μg/mL). DNA concentration and purity were assessed using the NanoDrop (Eppendorf, Hamburg, Germany) with absorbance at a wavelength of 260 nm and 280 nm.
Three microsatellite markers were used to recognize the variations of phenotypic traits between the four populations of Japanese quail. Full characterization of the analyzed microsatellite markers was presented in a previous study by Kayang et al. (Kayang et al. 2000). One monomorphic (GUJ0096) and two polymorphic markers (GUJ0063 and GUJ0085) were considered in this study. The primer sequences of these three microsatellite markers with their annealing temperature, GenBank accession numbers, and their length of repeats are shown in Table 2. The PCR mixture was carried out in a total volume of 25 μL containing 50 ng genomic DNA, 10 pmol of each primer, 2.5 μL 10X buffer, 1.5 mM MgCl2, 0.2 mM of each dNTP, and 1 U from DreamTaq (Thermo Fisher Scientific). The reaction was accomplished in TM Thermal Cycler (MJ Research PTC-100 thermocycler, USA). The thermal cycling profile included an initial denaturation step at 95 °C for 5 min followed by 35 cycles of 94 °C for 30 s, annealing at 55 °C for 1 min, and extension at 72 °C for 1 min, followed by a final extension step of final extension at 72 °C for 5 min. PCR successful products were identified on 2% agarose gel in 1X TBE buffer and visualized after staining with ethidium bromide (EtBr).
Statistical analysis
GLM procedure of SAS 9.2 (SAS Institute Inc., 2008) was used to analyze the phenotypic data. All the traits during the fattening and laying periods were analyzed depending on the following linear model: Yijk= μ+ Gi +GTj + (G×GT)ij+ eijk, where μ is the general mean, Gi is the effect of generation (2 levels (F1, F2), GTj is the effect of genetic type or quail varieties, (G×GT)ij is the effect of interaction between generation and quail varieties, and eijk is the residual effect. The different levels of each effect included in the models were compared using Duncan’s multiple range test where significance levels were detected as a first-class error at α=0.05. For carcass traits and blood parameters, the previous model was applied but with adding the sex effect (2 levels). For microsatellite data, the number of alleles for each marker was counted using a microsatellite analyzer 4.05 (Dieringer and Schlötterer 2003). Additionally, the mean number of effective alleles (Ne, the number of equally frequent alleles at a marker) and the heterozygosity of the 3 SSRs (observed heterozygosity (HO), expected heterozygosity (He)) in the four quail lines were estimated using GenAlEx version 6.0 (Peakall and Smouse 2006). Polymorphic information content (PIC) was determined using the Cervus software package (CERVUS 3.0.7). The fixation index (FIS) for each marker and the chi-square statistic for the Hardy-Weinberg equilibrium (HWE) were also calculated using the GENEPOP program version 3.4 (Raymond 1995). Also, using the POPULATIONS software (version 1.2.30) (Saitou and Nei 1987), the microsatellite phylogenetic tree of the four studied quail populations was constructed depending on the Nei genetic distance (DA) by using the neighbor-joining (NJ) method.
Results
Effect of quail varieties and generation of body weight measurements
Table 3 displays the effect of quail varieties, generation, and their interaction on body weight during the fattening and laying period. These factors significantly influenced the body weight measurements (P < 0.05). At the first week of age, the WB quail variety in F1 showed the highest body weights (P < 0.05). This effect was reversed in F2, and WB quails along with BW had the lowest body weights among the quail populations (P < 0.05). With increasing birds’ age (at the third and fourth weeks of age), the WW quails displayed the highest weights, followed by BW, compared to the other quail genotypes (P < 0.05).
During egg production, the male’s and female’s weights were recorded (Table 3). At 6 weeks old, the WW and BW males, in the first generation, had significantly higher body weights compared to the other quail varieties (P < 0.05), while WB males had the lowest body weight (P < 0.05). In the second generation, BW and BB males displayed the highest weights, while WW males had the lowest weights (P < 0.05). At 12 weeks old, the highest weights were found in the case of WW, WB, and BW (in a descending manner), but the BB males showed the lowest weights (P < 0.05). In the second generation, the BW males were the heaviest among the other quails’ males (P < 0.05). The latter finding continued until the end of egg production with the BW being the heaviest followed by WB males (P < 0.05). Unlike males, females did not show any changes in their body weights through the reported egg production period (P > 0.05).
The external and internal egg quality characteristics
Differences in egg numbers and weights among different quail populations through two successive generations were also reported (Tables 4 and 5). It seems that the number of eggs produced was significantly influenced by quail varieties and the generation. In this regard, the F1 WW and BW recorded a distinct higher onset of egg production compared to the other quails’ populations (P < 0.05). This higher egg number continued to the seventh week of egg production and then declined. In the second generation (F2), the BB achieved the highest number of eggs produced compared to the other quail varieties (P < 0.05). However, the production fluctuated between increases and decreases throughout the period of egg production. In general, there was a clear superiority of the F2 production compared to the F1 with the BB quail population being the most superior among quail populations. Besides, egg weights were influenced by the quail genotypes and generation (Table 5). The F1 showed higher egg weights compared to the F2 with the superiority being reported in the case of WW quails in F1 compared to others (P < 0.05).
Examining the effect of quail varieties and generation differences on the external and internal egg characteristics revealed an obvious competition among quail varieties through the two generations (Table 6). The values of egg length and width in F2 were higher than those of F1 with the highest values measured in the case of BW for egg length and WB and BW for egg width (P < 0.05). Additionally, yolk and albumen heights demonstrated higher values in F2 compared to F1 with the BB and BW in F1 and WW and WB in F2 having the highest values (P < 0.05). For yolk width, the highest scores were reported in the case of BB in F1 compared to the other quail genotypes in the two generations (P < 0.05). Albumen length and width did not display any changes among different quail genotypes in the two generations (P > 0.05). For shell thickness, the F2 values were significantly higher compared to those of F1 with BB genotypes having superiority in the two generations (P < 0.05). The yolk color additionally exhibited marked changes (P < 0.05). The highest values were found in the case of WB in F1 and BB in F2. For egg shape and yolk indices, they did not show any significant changes among quail populations and between generations (P > 0.05). While the quail population caused a significant effect on the albumen index values, the greatest values were noticed in the case of the WB population in F2, and the lowest values were measured in the case of WW and WB in F1 (P < 0.05).
Carcass trait properties among different quail populations throughout two successive generations
Differences in carcass characteristics among the four Japanese quail varieties and the two generations are presented in (Table 7). There are significant variations in live body weights and weights after slaughtering with the heaviest weights measured in the case of WB in F2 compared with the other varieties (P < 0.05). However, the highest percentages of carcass weights were noticed in the case of BW in F2, and WB showed the lowest % (P < 0.05). The relative weights of internal organs such as the liver, heart, and gizzard also exhibited marked differences among quail varieties and generations (P < 0.05). The highest relative weights of the liver were reported for WB in F1 and BW in F2 (P < 0.05), while the lowest relative weight was found in the case of WB in F2. The relative heart weight displayed the highest % in the case of BB in F2, while the lowest values were found in the case of BB, WW, and WB in F1 and WB in F2 (P < 0.05). For gizzard weights, there was not an obvious variation between F1 and F2, and the lowest % was found in the case of WB quail variety in F2 and WW in F1 (P < 0.05). The relative weight of the intestine was also calculated. All varieties exhibited nearly similar relative weights except WW which showed the lowest value (P < 0.05). BB and WB varieties in F2 showed the highest percentages of abdominal fat compared to the other varieties either in F1 or F2 (P < 0.05). Despite there being no significant variations in the ovary weights either among different quail varieties or generations, the F2 birds in all quail varieties showed higher weights of testicle and oviduct compared to those of F1 (P < 0.05).
The biochemical profile
The biochemical profile of the four quail populations was evaluated at the end of the study (Fig. 1). There were no distinct observed variations among different quail populations for all assessed parameters (P > 0.05). However, the greatest and lowest values of cholesterol were reported for BB and BW, respectively (P > 0.05). The WB quails displayed the highest non-significant concentration of total lipid compared to the others (P > 0.05). For MDA concentration, the WW and WB quails showed the highest levels, but BB and WB showed the lowest levels (P > 0.05). All quail populations displayed nearly the same total antioxidant capacities (P > 0.05). The WB and BB exhibited higher albumen and creatinine concentrations, while WW and WB showed the lowest levels, respectively (P > 0.05).

Biochemical constituents of the four japanese quail vareities (BB, WW, BW, and WB) at the end of the second generation (F2). A Cholestrol level (mg/dL), B total lipid concertation (mg/dL), C lipid peroxide (malondialdehyde), D total antioxidant capacity (μmol/L), and E, F albumin and creatinine concentration, respectively. Results were presented as mean ± SE
Genetic estimation
The amplification of the three studied microsatellite markers (GUJ0063, GUJ0085, and GUJ0096) was successfully done. Then, the genetic diversity of the three microsatellite loci among the four Japanese quail populations was analyzed and presented in Tables 8 and 9. However, due to the small number of markers used, these results are considered an initial clarification of genetic causes for the reported phenotypic variations. Further deep investigations depending on a larger number of markers are recommended.
The total number of the observed and effective alleles (NA and Ne, respectively) varied among quail populations. The NA numbers varied between 2 and 4 for the three studied markers (GUJ0063, GUJ0085, and GUJ0096), with a total average of 2.33±0.26 alleles. The greatest NA number (4) was found for the GUJ0063 marker in the case of the WB population, while the locus GUJ0096 showed a monomorphic pattern across the BB and WW quail populations. The average NA of the three markers within each quail population ranged from 2.00±0.58 in the case of BB and WW to 3.00±0.58 for the WB population. These observed numbers of alleles were associated with a relatively small to moderate effective number of alleles (Ne) ranging from 1.00 for the GUJ0096 in the case of the BB and WW quails to 2.90 for the GUJ0063 marker in the case of WB, with a total average of 1.76±0.17. Besides, the averages of Ne of the studied markers within each quail population were 1.66±0.21, 1.64±0.34, 1.53±0.29, and 2.22±0.49 for the BW, BB, WW, and WB, respectively. The allelic frequencies in the case of GUJ0063 and GUJ0096 ranged from 0.125 to 0.500 in the BW quails and ranged from 0.125 to 0.375 in the case of GUJ0085 in the same population. For BB quails, the allelic frequencies varied from 0.125 to 0.500, 0.167 to 0.500, and 1.00 for the GUJ0063, GUJ0085, and GUJ0096, respectively, while in the case of the WW quails, the allelic frequency ranged from 0.167 to 0.333 for the GUJ0063, 0.125 to 0.375 for the GUJ0085, and 1.00 for the GUJ0096. For the WB quails, the allelic frequencies varied from 0.125 to 0.250, 0.125 to 0.375, and 0.125 to 0.500 for the GUJ0063, GUJ0085, and GUJ0096, respectively.
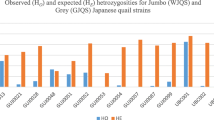
The levels of the observed and expected heterozygosity were also reported. The three microsatellite loci showed variable levels of observed heterozygosity (HO) ranging from 0.00 for the GUJ0085 and GUJ0096 in the case of BB and WW quails to 0.75 for the GUJ0063 in the case of the BB quails, with a total average of 0.28 ±0.07. The levels of the expected heterozygosity similarly ranged from 0.00 for the GUJ0096 in the case of BB and WW quails to 0.75 for the GUJ0063 in the case of WB quails. Moreover, Shannon’s diversity index (I) displayed variations ranging from 0.38 in the case of BW quails for the GUJ0096 locus to 1.21 for the GUJ0063 in the case of WW and WB quails, whereas the GUJ0096 was monomorphic in the case of the BB and WW quails. The fixation coefficient (FIS) ranged from −0.412 for the GUJ0063 in the case of the BB quails to 1.00 for the GUJ0085 in the case of the BB and WW quails, with an overall average of 0.242 ± 0.135. The FIS values of all markers within each population were 0.081±0.16, 0.29±0.58, 0.67±0.27, and 0.084±0.12 for the BW, BB, WW, and WB quails, respectively, while its values ranged from −0.143, 0.048, and 0.588 for the GUJ0096, GUJ0063, and GUJ0085, respectively.
Besides, the GUJ0063 (in the case of the BB, WW, and WB quails), GUJ0085 (in the case of the BW quails), and GUJ0096 (in the case of BW and WB quails) loci were highly informative markers (PIC > 0.50) with the PIC values ranging from 0.513 to 0.791. However, the GUJ0063 (in the case of BW quails) and GUJ0085 (in the case of the WB quails) displayed reasonably informative values (0.50 > PIC > 0.25) with the PIC values of 0.302 and 0.335, respectively. The GUJ0085 marker in the case of the BB and WW quails exhibited low PIC values, 0.145 and 0.165, respectively. Moreover, from the listed values of the chi-square and P values of the HWE in Table 8, we concluded that the HO and He did not differ significantly (P > 0.05) in all quail populations except the GUJ0085 in the case of the BW and WW quails which differed significantly (P >0.05). Table 9 presents the measures of the genetic identity and distance between the four studied quail populations. The lowest Nei genetic identity value (0.791) was recorded between the WB and WW populations, while the highest value (0.952) was found between the BW and BB quails. Regarding the genetic distance, the highest value (0.234) was between the WB and WW quails; thus, those strains were the farthest. However, the lowest value (0.049) was observed between the BW and BB quails; thus, those strains were the closest for the studied markers. These results were supported by the neighbor-joining (NJ) phylogenetic tree (Fig. 2). The tree showed a close relationship between the BW and BB quail populations. Furthermore, the BB and BW quails were clustered together and showed the most probable structure clustering, while the WW and WB populations were clustered independently confirming the high genetic distance.

Neighbor-joining unrooted phylogenetic tree of the four Japanese quail varieties depending on 3 microsatellite markers based on Nei’s genetic distances. POP1 = BW quails, POP2 = BB quails, POP3 = WW quails, and POP4 = WB quails
Discussion
Phenotypic characteristics
The current study represents a physiological comparative evaluation of two different plumage color Japanese quail populations (BB and WW) and their reciprocal crossings (BW and WB) to determine the best one for meat and egg production. In this regard, we depended on several measurements such as body weights, egg number, and weight, egg quality characteristics, and carcass traits, as well as the biochemical constituents which are considered as potentially concrete bases for choosing the appropriate egg-/meat-producing population that suits the breeders’ requirements. The foremost results indicated marked variations among the BB, WW, BW, and WB quails in the aforementioned measurements during the fattening and laying period. Among these variations, the findings of body weight measurements varied among all quail populations, which agreed with Nasr et al.’s, Tavaniello et al.’s, and Maiorano et al.’s findings (Maiorano et al. 2009; Nasr et al. 2017; Tavaniello et al. 2014) who stated significant variations in body weight between different lines of Japanese quails bred for meat (broilers) and egg production (layers). These differences might be attributed to the genetic makeup of quail varieties, age, and/or nutritional management (Tavaniello 2014). Among the studied quails’ populations, WW showed the superiority of body weights which might be explained by the significant correlations between plumage color and body weight (Al-Kafajy et al. 2018; Inci et al. 2015; Nasr et al. 2017). On the other side, our findings disagreed with the findings of Tarhyel et al. (2012) who reported that white-colored quails displayed the lowest body weights. Additionally, the BW and WB came in second and third places after the WW quails, respectively. Such superiority of cross lines perhaps indicates the direct additive genetic effect of sire line on quail body weight (Aboul-Seoud et al. 2021). Besides, the heavy weights were observed in F2 compared to F1 which could be explained by the higher level of heterosis in the F2 generation that again might be associated with the genetic structure of the F2 birds (Tavaniello 2014).
When comparing the carcass characteristics among different quail populations, significant variations were observed with the highest values reported in the case of WW and BW for carcass % and most of the internal organ weights; in addition, the values of F2 were higher than that of F1. These current results are in agreement with Sharaf et al.’s (Sharaf et al. 2006) findings which reported a significant superiority of crossbred quail populations compared to the purebreds as a result of the possible positive heterosis effect, while these findings disagreed with Inci et al.’s (Inci et al. 2015) findings which stated that the white-colored quails, compared to the wild-type quail, had the lowest values of carcass traits suggesting a depressive effect of the recessive white plumage color mutation on carcass traits. Regarding the abdominal fat %, the marked highest values were noticed in the case of the BB and WB quails. This perhaps can be explained by the association of high-fat deposition with brown plumage–colored quails. The differences in the measured carcass traits may be linked with the body weight variations among different quail populations. This is probably because of the positive genetic correlations between body weight and carcass traits (Hussain et al. 2013; Ibrahim et al. 2021). Therefore, body weight is considered the concrete index in evaluating the differences among different animal and poultry species (Ahmad et al. 2018; Hussain et al. 2013; Khaldari et al. 2010).
For the egg number, there were no clear superiorities of any of the studied quail populations through the recorded period of production. However, there were clear advantages of BB at the beginning of egg production, and then, the cross lines showed higher performance. These findings might come in agreement with Bed’hom et al.’s and Inci et al.’s (Bed’hom et al. 2012; Inci et al. 2015) findings who did not report any superiority of any different colored-quail populations. For egg weight, the values of the first hatch (F1) were higher, for all quail populations, than the second one (F2). Our findings were comparable with those of Ashok and Reddy and Lan et al. (Ashok and Reddy 2010; Lan et al. 2021) who stated an association between egg weight and different quail genotypes. This might be explained by the superior nongenetic effects on weights. Upon examining the effect of quail varieties and generation on egg quality characteristics including albumen height, albumen width, albumen weight, yolk width, yolk height, yolk color, yolk weight, the Haugh unit (HU), and eggshell thickness, along with shape, albumin, and yolk indices, significant variations were found. In general, the BW and WB had the highest values, relative to the others, and most of the assessed parameters and values of F2 were higher than that of F1. These findings might be explained with the significant association of feather color with egg quality characteristics in Japanese quail population (Al-Kafajy et al. 2018; Inci et al. 2015; Lan et al. 2021). Besides, for biochemical parameter evaluations, there were no clear variations among the studied quail populations. These findings have come in line with Al-Kafajy et al.’s results who reported non-significant differences among different plumage-colored quails in terms of cholesterol, total lipids, and triglycerides (Al-Kafajy et al. 2018).
The summary of the phenotypic evaluation indicated the presence of contradictory findings on the studied traits, with the previous studies, among the different plumage color quails, whereas some authors have proved no differences, while others established significant differences. This is owing to the complications to compare body weights, carcass, and egg quality results with those reported in other studies, because of many reasons such as the genetic structure, environmental and managerial conditions, considered statistical factors, feeding regime, and age of slaughtering (Vali 2008).
Genetic evaluation
Despite the obtained results, the three studied microsatellite markers could assist in genetically explaining the detected physiological variations in the four quail populations: BB, WW, BW, and WB in terms of body weights, carcass characteristics, and egg production and quality features. However, this explanation may be inadequate, and more markers are recommended for the future explanation since results are varied depending on the number of the studied microsatellite markers.
The genetic variation can be assessed depending on many genetic diversity indices such as the observed and the effective number of alleles (NA and Ne, respectively) as they give an indication on the difference in allele frequency in a population (Abdel-Kafy et al. 2021). In this regard, the average number of alleles ranged from 1 to 4 with a range of Ne form 1.00 to 2.90. This small range of NA probably explains the less genetic variation among the studied quail populations which might be explained by the small sample size of those quail populations that might predispose the genetic drift and inbreeding (Abdel-Kafy et al. 2021). Although there were small NA and Ne ranges which might indicate low genetic variation, the level of HO and He varied from 0.00 to 0.75 without deviation from HWE in most of the studied quail populations indicating that these quail populations were in HWE. However, the WW and BW quails had the lowest level of heterozygosity as confirmed by low values of HO (0.00 and 0.25, respectively) and a significant deviation from the Hardy-Weinberg equilibrium (HWE) in case GUJ0085, which might indicate a possibility of a non-random mating and inbreeding as confirmed by less heterozygosity (Abdel-Kafy et al. 2021) and the high positive values of heterozygosity deficit index (FIS). This high levels of inbreeding in these population might be risky as it could cause genetic diseases and negatively influence bird’s performance (Sharma et al. 2016). Our findings agreed with Ibrahim et al. and Kayang et al. (Ibrahim et al. 2021; Kayang et al. 2002) who reported nearly similar number of alleles, HO and He values. However, our findings disagreed with the previous studies which were conducted on different quail genotypes by Bai et al. (Bai et al. 2013; Bai et al. 2018), Kawahara-Miki et al. (Kawahara-Miki et al. 2013), and Shimma and Tadano (Shimma and Tadano 2019) which reported higher number of alleles. In summary, the incomparable results in most of the diversity estimates from different studies are probably because of using different microsatellite sets as well as the number of the analysed markers (Sharma et al. 2016). Additionally, the differences in allele numbers (NA and Ne) between the current and previous investigations may be linked with the differences in sample size, population structure, number of markers used, population-specific alleles and/or allele scoring bias (null allele or allele drop out), and sampling strategy (Rashid et al. 2020). Thus, more microsatellite markers are recommended for further explanation among these quail populations.
Besides, the values of the polymorphic information content (PIC) suggested that most of the used microsatellite loci were variably polymorphic and informative ( 0.20 > PIC > 0.50) according to the classification by Botstein et al. (Botstein et al. 1980). However, GUJ0096 marker showed monomorphic pattern in BB and WW quails. The high PIC values (> 0.50) for GUJ0063 in case of BB, WW, and WB, those for GUJ0085 in case of BW, and for GUJ0096 in case of BW and WB might suggest a high degree of polymorphism between the studied quail populations and the possibility of a progeny to acquire some allelic markers from its parents (Ibrahim et al. 2021). This finding requires further deep confirmation using more microsatellite and other genetic markers. In addition, the stated high PIC values of the studied markers possibly explain the potential role of these markers for population assignment as well as genome mapping studies in addition to genetic diversity analysis (Sharma et al. 2016). Nevertheless, this is not enough for validating the genetic diversity among the studied quail populations, and further investigations using more markers are recommended. Thus, collectively, from the results of HO, PIC, and Shannon’s information index, we can conclude that the selected 3 microsatellites markers, used in this study, might present a possible assessment of the genetic diversity in the different quail populations (Rashid et al. 2020).
Moreover, the BB and BW quail populations might be closely related based on the reported low values of genetic distance and high genetic identity. This probable low genetic differentiation between these two quail populations might designate a recent gene flow among themselves as well as common ancestors for constructing the populations (Rashid et al. 2020). However, again, this finding demands more detailed explanation using more microsatellite markers.
Conclusion
The current investigation presented the physiological variations and their possible preliminary genetic explanation between BB and WW and their reciprocal crosses, WB and BW, in live body weight, carcass characteristics, egg weights, and quality. In conclusion, the WW quails exhibited the heaviest weights followed by BW, WB, and BB, respectively. Besides, the F2 populations showed higher weights compared to the F1. Additionally, the WW and BW quails possessed higher percentages for the carcass, and most of the internal organ weights and the values of F2 were higher than that of F1. However, the BB and WB had the highest values of abdominal fat %. The obtained results might introduce an initial scientific basis for evaluating and employing the genetic properties of BB, WW, BW, and WB quails in the genetic improvement of the Japanese quail to overcome inbreeding in quail farming and, accordingly, improve its performance. Further deep analysis of the genetic diversity among these quail populations using more microsatellite markers is recommended.
Data availability
The authors acknowledge that the data presented in this study must be deposited and made publicly available in an acceptable repository, prior to publication.
Code availability
Not applicable.
References
Abdel-Kafy, E.-S.M., Ramadan, S.I., Ali, W.H., Youssef, S.F., Shabaan, H.A., El-Deighadi, A., and Inoue-Murayama, M., 2021. Genetic and Phenotypic Characterization of Domestic Geese (Anser anser) in Egypt. Animals, 11, 3106
Aboul-Seoud, D., Aboul-Seoud, D., Shaban, A., and Aboul-Hassan, M., 2021. Effect of crossing two selected lines for early and lately age at sexual maturity on some growth and carcass traits in Japanese quail. Al-Azhar Journal of Agricultural Research, 46, 67-73
Ahmad, S., Mehmood, S., Javed, K., Mahmud, A., Usman, M., Rehman, A., Ishaq, H., Hussain, J., and Ghayas, A., 2018. Different selection strategies for the improvement of the growth performance and carcass traits of Japanese quails. Brazilian Journal of Poultry Science, 20, 497-506
Al-Kafajy, F.R., Al-Shuhaib, M.B.S., Al-Jashami, G.S., and Al-Thuwaini, T.M., 2018. Comparison of three lines of Japanese quails revealed a remarkable role of plumage color in the productivity performance determination. The Journal of World's Poultry Research, 8, 111-119
Ashok, A., and Reddy, P.M., 2010. Evaluation of reproductive traits in three strains of japanese quail. Veterinary World, 3, 169-170
Bagh, J., Panigrahi, B., Panda, N., Pradhan, C., Mallik, B., Majhi, B., and Rout, S., 2016. Body weight, egg production, and egg quality traits of gray, brown, and white varieties of Japanese quail (Coturnix coturnix japonica) in coastal climatic condition of Odisha. Veterinary World, 9, 832
Bai, J., Huang, Y., Zhang, X., Yang, Y., Pang, Y., and Qi, Y., 2018. Polymorphism analysis of 3 quail groups by using microsatellite marker. Life Science Journal, 15,
Bai, J., Pang, Y., Wu, S., Yu, M., Zhang, X., Zhao, S., and Xu, H., 2013. Polymorphism analysis of Chinese yellow quail using microsatellite markers. The Journal of Animal and Plant Sciences, 23, 1072-1076
Bed’hom, B., Vaez, M., Coville, J.-L., Gourichon, D., Chastel, O., Follett, S., Burke, T., and Minvielle, F., 2012. The lavender plumage colour in Japanese quail is associated with a complex mutation in the region of MLPH that is related to differences in growth, feed consumption and body temperature. BMC genomics, 13, 1-10
Botstein, D., White, R.L., Skolnick, M., and Davis, R.W., 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. American journal of human genetics, 32, 314
Dávila, S., Gil, M., Resino-Talaván, P., and Campo, J., 2009. Evaluation of diversity between different Spanish chicken breeds, a tester line, and a White Leghorn population based on microsatellite markers. Poultry Science, 88, 2518-2525
Dieringer, D., and Schlötterer, C., 2003. Microsatellite analyzer (MSA): A platform independent analysis tool for large microsatellite data sets. Molecular Ecology Resources, 3, 167–169. https://doi.org/10.1046/j.1471-8286.2003.00351.x
ELSaidy, N., Kirella, A., El-Kassas, S., Dawood, M.A., and Abouelenien, F., 2021. Reducing the Abundance of Harmful Bacteria of Rooftop Tank–Stored Drinking Water Using Silver Nanoparticles and Acetic Acid and Its Impact on Japanese Quail Growth Performances. Biological Trace Element Research, 199, 3062-3072
Farajiarough, H., Rokouei, M., Maghsoudi, A., and Ghazaghi, M., 2018. Comparative study of growth patterns in seven strains of Japanese quail using nonlinear regression modeling. Turkish journal of veterinary and animal sciences, 42, 441-451
Hussain, J., Akram, M., Sahota, A., Javed, K., Ahmad, H., Mehmood, S., Ahmad, S., Sulaman, R., Rabbani, I., and Jatoi, A., 2013. Selection for higher three week body weight in Japanese quail: 1. Effect on growth performance. JAPS: Journal of Animal & Plant Sciences, 23,
Ibrahim, N.S., El-Sayed, M.A., Assi, H.A.M., Enab, A., and Abdel-Moneim, A.-M.E., 2021. Genetic and physiological variation in two strains of Japanese quail. Journal of Genetic Engineering and Biotechnology, 19, 1-12
Inci, H., Sogut, B., Sengul, T., Sengul, A.Y., and Taysi, M.R., 2015. Comparison of fattening performance, carcass characteristics, and egg quality characteristics of Japanese quails with different feather colors. Revista Brasileira de Zootecnia, 44, 390-396
Kawahara-Miki, R., Sano, S., Nunome, M., Shimmura, T., Kuwayama, T., Takahashi, S., Kawashima, T., Matsuda, Y., Yoshimura, T., and Kono, T., 2013. Next-generation sequencing reveals genomic features in the Japanese quail. Genomics, 101, 345-353
Kayang, B., Inoue-Murayama, M., Nomura, A., Kimura, K., Takahashi, H., Mizutani, M., and Ito, S., 2000. Brief communication. Fifty microsatellite markers for Japanese quail. Journal of Heredity, 91, 502-505
Kayang, B., Vignal, A., Inoue-Murayama, M., Miwa, M., Monvoisin, J.L., Ito, S., and Minvielle, F., 2004. A first-generation microsatellite linkage map of the Japanese quail. Animal genetics, 35, 195-200
Kayang, B.B., Inoue-Murayama, M., Hoshi, T., Matsuo, K., Takahashi, H., Minezawa, M., Mizutani, M., and Ito, S.I., 2002. Microsatellite loci in Japanese quail and cross-species amplification in chicken and guinea fowl. Genetics Selection Evolution, 34, 233-253
Khaldari, M., Pakdel, A., Yegane, H.M., Javaremi, A.N., and Berg, P., 2010. Response to selection and genetic parameters of body and carcass weights in Japanese quail selected for 4-week body weight. Poultry Science, 89, 1834-1841
Kimura, M. and Crow, J.F., 1964. The Number of Alleles That Can Be Maintained in a Finite Population. Genetics, 49, 725-738.
Lan, L., Nhan, N., Hung, L., Diep, T., Xuan, N., Loc, H., and Ngu, N., 2021. Relationship between plumage color and eggshell patterns with egg production and egg quality traits of Japanese quails, Veterinary World, 14 (4): 897-902.
Lewontin RC. 1972. The apportionment of human diversity. In Evolutionary biology (eds Steere WC, Dobzhansky T, Hecht MK), pp. 381-398. New York, NY: Springer.
Maiorano, G., Elminowska-Wenda, G., Mika, A., Rutkowski, A., and Bednarczyk, M., 2009. Effects of selection for yolk cholesterol on growth and meat quality in Japanese quail (Coturnix coturnix japonica). Italian Journal of Animal Science, 8, 457-466
Miwa, M., Inoue-Murayama, M., Kayang, B., Vignal, A., Minvielle, F., Monvoisin, J., Takahashi, H., and Ito, S., 2005. Mapping of plumage colour and blood protein loci on the microsatellite linkage map of the Japanese quail. Animal genetics, 36, 396-400
Nasr, M.A., Ali, E.-S.M., and Hussein, M.A., 2017. Performance, carcass traits, meat quality and amino acid profile of different Japanese quails strains. Journal of food science and technology, 54, 4189-4196
Nunome, M., Yoshioka, R., Shinkai, T., Kino, K., and Matsuda, Y., 2020. Assessment of genetic diversity and genetic relationships of farm and laboratory quail populations in Japan using microsatellite DNA markers. Veterinary Medicine and Science, 6, 1000-1008
Peakall, R., and Smouse, P.E., 2006. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular ecology notes, 6, 288-295
Rashid, M.A., Manjula, P., Faruque, S., Bhuiyan, A.F.H., Seo, D., Alam, J., Lee, J.H., and Bhuiyan, M.S.A., 2020. Genetic diversity and population structure of indigenous chicken of Bangladesh using microsatellite markers. Asian-Australasian Journal of Animal Sciences, 33, 1732
Raymond, M., 1995. GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. Journal of Heredity, 86, 248-249
Saitou, N., and Nei, M., 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular biology and evolution, 4, 406-425
Sharaf, M., Mandour, M., and Taha, A., 2006. Effect of diallel crossing on same growth performance, carcass traits and immune response against new castle disease virus vaccine of Japanese quails. Egyptian Poultry Science, 3, 1451-1470
Sharma, R., Kumar, B., Arora, R., Ahlawat, S., Mishra, A., and Tantia, M., 2016. Genetic diversity estimates point to immediate efforts for conserving the endangered Tibetan sheep of India. Meta gene, 8, 14-20
Shimma, K., and Tadano, R., 2019. Genetic differentiation among commercial lines of laying-type Japanese quail. The Journal of Poultry Science, 56, 12-19
Tarhyel, R., Tanimomo, B., and Hena, S., 2012. Effect of sex, colour and weight group on carcass characteristics of Japanese quail. Scientific Journal of Animal Science, 1, 22-27
Tavaniello, S., 2014. Effect of cross-breed of meat and egg line on productive performance and meat quality in Japanese quail (Coturnix japonica) from different generations. (UNIVERSITY OF MOLISE)
Tavaniello, S., Maiorano, G., Siwek, M., Knaga, S., Witkowski, A., Di Memmo, D., and Bednarczyk, M., 2014. Growth performance, meat quality traits, and genetic mapping of quantitative trait loci in 3 generations of Japanese quail populations (Coturnix japonica). Poultry Science, 93, 2129-2140
Vali, N., 2008. The Japanese quail: A review. International Journal of Poultry Science, 7, 925-931
Acknowledgements
The authors would like to thank Dr. Sherif I. Ramadan, associate professor of Animal and Poultry Production, Department of Animal Wealth Development, Faculty of Veterinary Medicine, Benha University, Toukh, 13736, Egypt, for his help in analyzing the SSR data.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This work was financially supported in the framework of the project “Biological production of nano-selenium spheres and its application in livestock production” by the National Strategy for Genetic Engineering and Biotechnology, Academy of Scientific Research and Technology, Egypt.
Author information
Authors and Affiliations
Contributions
IE, YE, MG, and MR conceived and designed the research. IE, SE-K, MG, and MR conducted the experiments. SE-K contributed new reagents or analytical tools. IE, SE-K, and MA analyzed the data. SE-K, YE, EME-K, and MA drafted the manuscript. YE, SE-K, and MA reviewed and approved the final version before submission. All authors read and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval
The animal study was reviewed and approved, and all experiments in the current study have been done in line with the Local Experimental Animal Care Committee guidelines and approved by the Institutional Ethics Committee of Kafrelsheikh University, Egypt.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Elkhaiat, I., El-Kassas, S., Eid, Y. et al. Assessment of variations in productive performance of two different plumage color varieties of Japanese quail and their reciprocal crosses. Trop Anim Health Prod 55, 195 (2023). https://doi.org/10.1007/s11250-023-03604-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11250-023-03604-5