
Abstract
The formation of an extracellular matrix surface network (ECMSN), and associated changes in the distribution of arabinogalactan-protein and pectin epitopes, have been studied during somatic embryogenesis (SE) and callogenesis of Trifolium nigrescens Viv. Scanning electron microscopy observations revealed the occurrence of an ECMSN on the surface of cotyledonary-staged somatic embryos as well as on the peripheral, non-regenerating callus cells. The occurrence of six AGP (JIM4, JIM8, JIM13, JIM16, LM2, MAC207) and four pectin (JIM5, JIM7, LM5, LM6) epitopes was analysed during early stages of SE, in cotyledonary-staged somatic embryos and in non-embryogenic callus using monoclonal antibodies. The JIM5 low methyl-esterified homogalacturonan (HG) epitope localized to ECMSN on the callus surface but none of the epitopes studied were found to localize to ECMSN over mature somatic embryos. The LM2 AGP epitope was detected during the development of somatic embryos and was also observed in the cell walls of meristematic cells from which SE was initiated. The pectic epitopes JIM5, JIM7, LM5 and LM6 were temporally regulated during SE. The LM6 arabinan epitope, carried by side chains of rhamnogalacturonan-I (RG-I), was detected predominantly in cells of embryogenic swellings, whilst the LM5 galactan epitope of RG-I was uniformly distributed throughout the ground tissue of cotyledonary-staged embryoids but not detected at the early stages of SE. Differences in the distribution patterns of low and high methyl-esterified HG were detected: low ester HG (JIM5 epitope) was most abundant during the early steps of embryo formation and highly methyl-esterified form of HG (JIM7 epitope) became prevalent during embryoid maturation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Plant cell walls together with the cytoskeleton determine polarity and control plant morphogenesis (Baluška et al. 2003). Changes in cell wall composition play crucial roles during plant growth and development, pathogen interactions as well as in responses to environmental signals. Precise cell divisions and cell expansion during somatic embryogenesis (SE) are influenced by cell wall components and properties. Remodeling of cell walls during plant development requires deposition, modification and degradation of the existing set of macromolecules but also continued synthesis and deposition of new polymers (for review see Cosgrove 2005; Seifert and Blaukopf 2010).
Arabinogalactan-proteins (AGPs) are plant cell surface macromolecules belonging to the subfamily of hydroxyproline-rich glycoproteins. They can act as signalling molecules and are involved in vegetative, reproductive and cellular growth and development (for review see Seifert and Roberts 2007; Ellis et al. 2010). AGPs are associated with plasma membranes, cell walls and secretions (Showalter 2001). Studies with β-glucosyl Yariv reagent, which can precipitate AGPs, have indicated their involvement in cell proliferation in cell suspension cultures of Rosa sp. (Serpe and Nothnagel 1994) in cell elongation in carrot cell suspension cultures (Willats and Knox 1996) and in programmed cell death in cell cultures of Arabidopsis (Gao and Showalter 1999). It has been also demonstrated that AGPs can regulate SE. Application of different fractions of exogenous AGPs can either promote or inhibit somatic embryo formation in carrot callus culture (Kreuger and van Holst, 1995; Toonen et al. 1997). Studies with AGP antibodies have demonstrated that the expression of some AGP epitopes (JIM4, JIM13, JIM16 and LM2) are developmentally regulated during SE in Daucus (Stacey et al. 1990), Zea (Šamaj et al. 1999a) and Cichorium (Chapman et al. 2000a) and can serve as specific developmental markers of embryogenic cells or/and somatic embryos. The exact mechanisms underlying AGP functions are unclear. However, roles for AGPs in cellular adhesion, signalling and recognition during SE have been proposed (Šamaj et al. 1999a, 2006).
Pectins are major components of primary cell walls of all land plants. These acidic polysaccharides are characterized by structural complexity and heterogeneity (for review see Verhertbruggen and Knox 2006; Harholt et al. 2010). Pectins contribute to the mechanical properties of cell walls (porosity, elasticity, adhesion, binding of ions). As multifunctional macromolecules pectins are involved in plant growth, development and morphogenesis (Jarvis et al. 2003) and can also serve as a source of oligosaccharide fragments which act as signalling molecules for development and defense (Dumville and Fry 2000). The use of monoclonal antibodies has shown that the distribution of different pectic epitopes in cell walls is developmentally regulated and can also be tissue, organ and species specific (Bush and McCann 1999; Willats et al. 1999; McCartney et al. 2000). In the embryogenic cultures of carrot (Kikuchi et al. 1995) and Medicago arborea (Endress et al. 2009) differences were evident in the pectic polysaccharide content between embryogenic and non-embryogenic callus cells. Data relating to the distribution of specific pectic epitopes during SE are scarce and indicate differences in the methyl-esterification of homogalacturonan (HG) during the early stages of SE (Verdeil et al. 2001; Šamaj et al. 2006).
Induction of morphogenesis in several plant species cultured in vitro has been reported to be accompanied by the presence of a continuous fibrillar layer on the outer cell surface. This structure called an extracellular matrix surface network (ECMSN) was observed during SE in monocotyledons (maize, Šamaj et al. 1995; peach palm, Steinmacher et al. 2012), dicotyledons (coffee, Sondahl et al. 1979; Cichorium hybrid, Dubois et al. 1990) as well as in gymnosperms (black pine, Jasik et al. 1995). The ECMSN has been specifically related to induction and early stages of SE (Šamaj et al. 1995; Rumyantseva et al. 2003; Bobák et al. 2004). This structure was also reported on the root surface of maize (Šamaj et al. 1999b) during androgenesis in wheat anther cultures (Konieczny et al. 2005) and in organogenesis induced in kiwifriut endosperm-derived callus (Popielarska-Konieczna et al. 2008). Analysis of the chemical composition of the ECMSN with the use of different solvents and enzymes revealed differences between species and identified both proteins (Dubois et al. 1992) and lipophilic substances (Konieczny et al. 2005) as components. Other authors using specific antibodies identified AGP and pectin epitopes in the ECMSN: the JIM4 AGP epitope was detected in an embryogenic culture of maize (Šamaj et al. 1999a) the JIM5 pectic HG epitope in embryogenic coconut calli (Verdeil et al. 2001). The role of ECMSN during morphogenesis is still uncertain, but cell adhesion, a protective function and co-ordinating developmental stages have been proposed (Verdeil et al. 2001).
Recently, we reported that immature zygotic embryos of Trifolium nigrescens Viv. can produce somatic embryos on media containing 1-naphtaleneacetic acid (NAA) and N6-[2-isopentenyl]-adenine (2iP) (Konieczny et al. 2010). In this study using immunolabeling techniques we report the changes in the distribution of selected AGP and pectic epitopes in relation to somatic embryo development and ECMSN formation.
Materials and methods
Plant material and culture conditions
Seeds of T. nigrescens Viv. ssp. nigrescens were kindly provided by the Institute of Plant Genetics and Crop Plant Research, Gatersleben, Germany. The procedure for donor plant growth, explant preparation and culture conditions for SE induction were as described previously (Konieczny et al. 2010). Briefly, somatic embryos were induced from 3 cm in length immature zygotic embryos at the cotyledonary stage (CsZE) on Murashige and Skoog (1962) basal medium supplemented with 30 g l−1 (w/v) sucrose, 8 g l−1 agar (Difco, USA), 0.5 mg l−1 NAA and 2.0 mg l−1 2iP, pH 5.7. Cultures were maintained in a growth chamber for 15 days at 25 ± 2 °C under diffuse cool white fluorescent light (80 μmol m−2 s−1) with a 16 h photoperiod.
Scanning electron microscopy
Explants after 0, 7 and 15 days of culture were fixed in 5 % (v/v) glutaraldehyde in 0.1 M phosphate buffer (pH 7.2) for 2 h at room temperature. After dehydration through graded ethanol series ethanol was replaced by acetone prior to drying in a critical-point drying system (LADD critical point dryer). Samples were then coated with gold (Jeol JFC-1100 E ion-sputtering system) and observed with a scanning electron microscope (Hitachi S-4700) at 20 kV.
Conventional light and fluorescence microscopy
For microscopy observations (conventional light and fluorescence) samples were collected after 7, 10 and 15 days of culture. For conventional light microscopy material was fixed in 5 % (v/v) glutaraldehyde in 0.1 M phosphate buffer (pH 7.2), dehydrated in a graded series of ethanol and embedded in Technovit 7100 (Heraeus Kulzer, Germany) according to manufacturer’s instructions. Sections (5 μm thickness) were routinely stained with 0.1 % (w/v) aquatic solution of toluidine blue or with periodic acid-Schiff’s reagent (PAS) for starch visualization.
For immunolabeling the explants were fixed for 2 h at room temperature in 4 % (v/v) formaldehyde in PEM buffer (50 mM PIPES, 5 mM EGTA, 5 mM MgSO4, pH 6.9). After fixation samples were washed in phosphate-buffered saline (PBS, pH 7.2) and dehydrated at 4 °C in an ethanol series (30, 50, 70, 90, 96 %). Then the samples were infiltrated in Steedman’s wax (polyethylene glycol 400 distearate and 1-hexadecanol in proportion 9:1). Sections (7 μm thickness) were collected on poly-l-lysine coated microscope slides (VWR International, USA) and de-waxed in 96 % ethanol, rehydrated in an ethanol series and rinsed in PBS. Specimens were blocked in PBS containing 5 % milk protein (Marvel, Premier Beverages, UK) for 30 min. Slides were incubated in a 5-fold dilution of primary monoclonal antibodies (Plant Probes, UK) for 1.5 h in order to detect different pectin and arabinogalactan epitopes (Supplementary Table 1). After washes in PBS, samples were incubated with anti-rat antibody coupled with fluorescein isothiocyanate (FITC, Sigma-Aldrich, USA; dilution 1:100) for 1 h in darkness. After labeling the slides were rinsed extensively in PBS. The sections were mounted in anti-fade medium Citifluor AF1 (Agar Scientific, UK) and examined under an Olympus BX61 microscope equipped with epifluorescence irradiation.
Micrographs of embryogenic swellings and globular embryoids (early-staged somatic embryos referred here as EsSE), cotyledonary-staged somatic embryos (CsSE) and callus were taken using a Hamamatsu ORCA285 camera (Hamamastu, http://www.hamamatsu.com) and Volocity software (Perkin Elmer).
Results
Identification of ECMSN on the surface of cotyledonary-staged somatic embryos and parenchymatous cells of callus
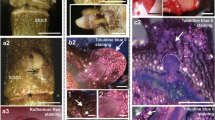
Scanning electron microscopy (SEM) images revealed that protodermal cells of CsZE of T. nigrescens were tightly packed with a smooth surface devoid of fibrillar or granular protrusions (Fig. 1a, b). On the 7th day of culture numerous globular- and heart-stage somatic embryos were formed directly from the explant, on the swollen hypocotyl region of CsZE, just below the cotyledonary nodes (Fig. 1c–e). The surface of somatic embryos during early stages of development as well as the protodermis of cultured explants were smooth and devoid any secretion (Fig. 1d, e). SE was asynchronous and after 15 days numerous somatic embryoids were observed at different stages of development on the explant (Fig. 1f). CsSE appeared as bipolar structures with two clearly visible cotyledons and an elongated embryo axis (Fig. 1f, g). After 15 days of culture direct SE was accompanied by the intense proliferation of callus originating from explant hypocotyl (Fig. 1f). In contrast to the early stages of somatic embryo development, CsSE were covered with strands of a compact layer (Fig. 1g, h) or were almost entirely coated with continuous ECMSN (Fig. 1i) with a net-like appearance at its edge (Fig. 1j). The peripheral part of the callus was composed of large, oval or oblong, loosely attached cells of parenchymatous nature, most of which were covered by a distinct ECMSN formed either as a discontinuous fibrillar network and/or membrane-like coat covering a relatively large area of tissue (Fig. 1k, l). With continued culture this outer part of callus reminded non-regenerative and no changes in appearance of callus-produced ECMSN were observed.

SEM micrographs of T. nigrescens CsZE after 0 (a, b), 7 (c–e) and 15 days of culture (f–l). a Hypocotyl of CsZE (explant); b magnification of hypocotyl surface cells from a. c Somatic embryos emerging directly from hypocotyl of CsZE (arrowhead-globular, arrows-heart-stage). d, e Early stages of somatic embryo development: globular (d) and heart-stage (e) somatic embryo with smooth surface; inset showing detailed view of embryo surface. f Explant with torpedo stage and mature somatic embryos developed directly on explant hypocotyl and callus cells (arrowhead) emerging from CsZE axis. g–j Cotyledonary stage embryoids coated partly (g) or almost entirely (i) with continuous ECMSN (arrowhead in g, i); h detailed view of protodermis and ECMSN from g; j magnification from i. k, l Peripheral, non-regenerating callus cells. k Fibrillar net-like material on the callus and between the cells (arrowhead) and continuous ECMSN covering callus cells (arrow); l surface view of heterogeneous layer from k. Bar = 200 μm (c, f); 100 μm (a, g, i, k); 50 μm (e); 30 μm (b, d, j); 10 μm (h, l)
The LM2 AGP epitope and LM6 RG-I epitope were expressed in cells from which somatic embryos develop
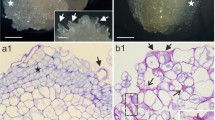
After 7 days of culture both protodermis and the subprotodermal regions of CsZE hypocotyls became more meristematic in appearance followed by production of numerous embryogenic swellings which protruded over the explant surfaces (Fig. 2a). During this period of culture the cells of the inner cortex of CsZE enlarged markedly when compared to the control ones (data not shown) and their nuclei became faintly stained or even invisible after toluidine blue staining. SE was asynchronous and after 10 days of culture embryogenic swellings and globular embryoids (Fig. 2b) could be observed concomitantly on the same explant. Cell walls of embryogenic swellings were not labeled by most of anti-AGP antibodies tested: JIM4, JIM8, JIM13, JIM16 and MAC207 (data not shown) although JIM8 and JIM13 epitopes were detected in the outer cells of explant. However, strong fluorescence due to LM2 binding was identified in cell walls of meristematic cells at the explant periphery and in cells of embryogenic swellings (Fig. 2c).

Indirect immunofluorescence detection of AGP and pectin epitopes in EsSE and CsSE of T. nigrescens. a Sections of embryogenic protuberance on the explant after 7 days of culture stained with toluidine blue. Explants after 10 (b–g) and 15 (h–k) days of culture. b Section stained with PAS reaction showing globular embryoids. c Section showing LM2 epitope occurrence (arrowhead) in the cell walls of embryogenic swellings. d Section showing JIM5 weak binding to the cells of EsSE (arrowhead); JIM5 epitope expression in the explant tissues mainly restricted to the cells lining extracellular spaces (inset, arrowhead). e Weak fluorescence signal in the embryogenic swelling due to JIM7 (arrowhead). f LM5 binding not detected in embryogenic protrusion. g Strong signal of LM6 binding to some cells of embryogenic swelling (arrowhead). h PAS reaction-stained section showing anatomy of the embryo. i Section showing only weak binding of JIM5 to cortex of CsSE (arrowheads). j Moderate fluorescence signal of JIM7 binding to cortex and protodermis (arrowhead). k Section showing strong signal due to LM5 epitope in cortex but no signal in the protodermal cells; magnification of cotyledon (inset). Bar = 100 μm
Pectic epitopes were visible in the explant and EsSE cell walls. The low methyl-esterified HG epitope (JIM5) was especially abundant in the cell walls of the tricellular junctions in the explant cortex, whilst cell walls of embryogenic swellings showed a moderate fluorescence signal (Fig. 2d). The high methyl-esterified HG epitope (JIM7) was detected in cell walls of almost all cells of the explant as well as in the cells of emerging embryos, but detection within embryogenic protrusions was much weaker compared to the other cells of explant (Fig. 2e). The LM5 galactan epitope, carried by side chains of RG-I, was expressed by some cells of the inner cortex of the explant and was not found in the somatic embryos during early stages of development (Fig. 2f). The expression of the LM6 arabinan epitope of side chains of RG-I was present in the cells of embryogenic swellings as well as in the protodermis and subprotodermal region of the explant, but detected in only some cells of the inner cortex of the CsZE hypocotyl (Fig. 2g).
No AGP epitopes were detected in cotyledonary-staged somatic embryos, whilst pectic HG and RG-I epitopes were expressed in a tissue dependent manner
Histological observation revealed that CsSE had distinct root and shoot meristems and two cotyledons (Fig. 2h). The cells of CsSE were not labeled by any of the anti-AGPs antibodies (data not shown). Out of four anti-pectin antibodies used here, three (JIM5, JIM7, LM5) bound to the cell walls of CsSE (Fig. 2i–k). Weak detection of the JIM5 epitope was observed in the cells of cotyledons, hypocotyl and root pole (Fig. 2i). Significantly, the JIM5 epitope was not found in the cell walls of protodermis. The whole somatic embryo, including protodermal cells, showed moderate immunofluorescence due to JIM7 binding (Fig. 2j). The LM5 galactan epitope was strongly detected in cells of the cortex, but the protodermis remained unlabeled by this probe (Fig. 2k). The LM6 arabainan epitope was not detected in cell walls of CsSE. Notably, none of the anti-AGP and anti-pectin antibodies tested bound to the surface of mature somatic embryos (data not shown).
Abundant expression of the JIM5 HG epitope overlaps with ECMSN on parenchyma cells of callus
After 15 days of culture, callus was composed of loosely attached parenchymatous, non-regenerative cells in the outer parts of tissue and much smaller, compact cells in its inner region. Positive staining with the PAS reaction revealed starch grains in most callus cells and also polysaccharide-containing secretions between loosely attached peripheral cells of tissue (Fig. 3a). The AGP epitopes JIM4 and JIM16 were not found in the callus (data not shown). The AGP epitopes JIM8, JIM13 and LM2 were detected in the parenchymatic peripheral cells but not in the inner region of the callus (Fig. 3b–d). Conversely, both outer and inner cells of callus showed a weak fluorescence signal due to MAC207 (Fig. 3e).

Indirect immunofluorescence detection of AGP and pectin epitopes in non-regenerative callus tissue of T. nigrescens after 15 days of culture. a Section stained with PAS reaction showing general view: loosely attached, large parenchymatic cells in the outer part of tissue and PAS positive secretions between callus cells (arrowhead). Immunofluorescence (arrow) of loosely attached callus cells due to JIM8 (b) JIM13 (c) and LM2 (d). e Weak fluorescence signal of MAC207 binding to detaching (arrow) and inner callus cells. f, g Sections showing JIM5 binding to the outer and inner callus cells (arrows in f, g) and between the cells (arrowhead in f). h Weak immunofluorescence of callus cells due to JIM7 (arrow). LM5 binding to both, inner and detaching callus cells (i). j LM6 epitope immunolocalized in some outer callus cells (arrow). Bar = 100 μm
Immunolocalization of the JIM5 HG epitope was observed in the intercellular spaces between peripheral callus cells (Fig. 3f) coinciding with ECMSN visible in the SEM micrographs. JIM5 and JIM7 epitopes were localized in cell walls of callus cells: a low level of the JIM7 epitope was present in both outer parenchyma and more meristematic inner regions of tissue whilst the JIM5 epitope was abundant in cell walls of detaching parenchyma and only weak fluorescence due to JIM5 binding was observed in the cells localized in the central part of callus (Fig. 3g, h). All callus cells were moderately marked by the LM5 antibody (Fig. 3i) but the LM6 arabinan epitope was only weakly detected in occasional surface callus cells (Fig. 3j).
Discussion
Structure of the ECMSN
When maintained on medium containing NAA and 2iP, CsZE of T. nigrescens produced somatic embryos followed by induction of callus from the hypocotyl of explants. SEM observations indicated that both SE and callogenesis were accompanied by the occurrence of a conspicuous ECMSN as a continuous layer or fibrillar network covering the neighboring cells of mature embryoids or calli. Similar extracellular structures have been considered as a structural marker of embryogenic development in vitro in several monocots and dicots (Šamaj et al. 2006). In T. nigrescens ECMSN of similar appearance to that found on somatic embryos was also observed on large, external, non-regenerative cells of callus. This observation is consistent with previous studies on wheat (Pilarska et al. 2007) and banana (Xu et al. 2011) and confirm that in certain species and/or under specific in vitro conditions the ability to produce ECMSN is not a unique feature of cells capable of regeneration.
Formation of an ECMSN was shown to be developmentally regulated and restricted to early stages of SE (e.g. Šamaj et al. 1995, 1999a; Blehová et al. 2010; Lai et al. 2011). In contrast to above-mentioned reports, the ECMSN of T. nigrescens occurred during late stages of embryo regeneration only, i.e. on cotyledonary-staged embryoids, but was never observed either on SE-competent cells of explants or on embryoids at globular, heart or torpedo stages. Šamaj et al. (1995) and Bobák et al. (2003/4) suggested that the formation of an ECMSN can be a stress response of the explants imposed by specific in vitro conditions. Previously, we found that the incidence of both morphological abnormalities and genetic instability in SE of T. nigrescens increased with time of culture (Konieczny et al. 2012). It may not be ruled out, that occurrence of ECMSN at relatively late stages of culture (around 15 days after explantation), could be a result of prolonged exposure to unfavorable culture conditions. On the other hand, the ability to produce an ECMSN in T. nigrescens seems to be a specific feature of tissues induced in vitro: throughout the culture no extracellular secretion was observed on the surface of original explants, that is, cotyledonary-staged zygotic embryos.
In order to gain insight into the chemical composition of the ECMSN in T. nigrescens we used immumnocytochemistry with cell wall directed probes. No AGP or pectin epitopes were detected on the external cell walls of surface cells of somatic embryos. However, strong detection of the JIM5 HG epitope was observed in the cell walls and within intercellular spaces of outer parenchyma of non-regenerative callus. The abundance of polysaccharides between peripheral cells of T. nigrescens callus was also confirmed by positive PAS reaction. Pectin polymers have been already identified in the ECMSN of embryogenic cultures of carrot (Iwai et al. 1999), chicory (Chapman et al. 2000b), coconut (Verdeil et al. 2001), and banana (Xu et al. 2011). In addition, conspicuous differences in pectic composition of the ECMSN coating embryogenic/organogenic and non-regenerative callus domains were reported (Iwai et al. 1999; Konieczny et al. 2007; Pilarska et al. 2007; Xu et al. 2011). The results obtained in our study also indicate a tissue-specific composition of the ECMSN: JIM5-reactive HG was detected in the ECMSN over callus surface but not on the surface of somatic embryos. In tissues cultured in vitro, Popielarska-Konieczna et al. (2008) and Xu et al. (2011) localized the JIM5 epitope to the ECMSN coating non-dividing and relatively large parenchyma cells of callus. Noticeably, the cells of T. nigrescens that carried the JIM5 epitope were loosely attached which suggests a relationship between location of low methyl-esterified HG and cell separation in cultured callus. Indeed, abundant accumulation of low ester HG has been considered as a common mechanism of withstanding stress that tends to tear cells from neighbours (Jarvis et al. 2003). Šamaj et al. (1999a, 2006) reported expression of the AGP epitope JIM4 together with the pectic epitope JIM7 in the mucilage layer on the surface of Zea mays embryogenic callus. The AGPs recognized by the LM2, JIM13 and JIM16 antibodies as well as JIM5- and JIM7-reactive pectins were found in the ECMSN coating globular proembryos of Cichorium (Chapman et al. 2000a). In T. nigescens only the JIM5 epitope localized to extracellular secretions, whilst the expression of other identified epitopes (JIM7, LM5, LM6, MAC207, LM2, JIM8, JIM13) was restricted exclusively to the cell walls of callus cells. This would allow us to suggest that low methyl-esterified HG recognized by the JIM5 antibody might be the main component of strands, fibrils or continuous coat of the ECMSN over callus surface. Although a possible involvement of other pectic substances (recognized by JIM7, LM5, LM6) and some AGPs (recognized by MAC207, LM2, JIM8, JIM13) in the formation of ECMSN in T. nigescens callus should not be excluded.
Localization of pectin and AGP epitopes in somatic embryos and callus tissue
In T. nigrescens the LM2 epitope was the only AGP epitope detected during the development of somatic embryos. Strong detection of the LM2 epitope was observed in the meristematic cells from which somatic embryos were produced, but no signal was detected in mature embryoids. Previously, Pan et al. (2011) reported in banana a diversity of immunolabeled AGPs that decreased with somatic embryo maturation. Saare-Surminski et al. (2000) suggested a crucial role of AGPs bearing the LM2 epitope for induction of early embryogenic stages in Euphorbia pulcherimma. In T. nigrescens, LM2 binding was observed not only in cell walls of meristematic cells but also loosely attached and non-dividing cells of callus parenchyma. This may indicate the dual role for LM2-reactive AGPs in the tissues studied: associated with induction of SE and, in case of callus, related to cell maturation/separation. Indeed, the epitope recognized by a single antibody may occur on different AGPs with different protein backbones and functions (Schultz et al. 2000).
Use of antibodies raised against arabinan (LM6) and galactan (LM5) side chains of RG-I revealed that these polymers were temporally regulated during the development of somatic embryos of T. nigrescens. Our observations are consistent with previous studies on the sequential appearance of galactose-rich domains of RG-I in differentiating cells and maturating somatic embryos (Willats et al. 1999; McCartney et al. 2003; Xu et al. 2011), a phenomenon which may lead to cell wall stiffening (McCartney et al. 2000). Conversely, actively proliferating cells of callus or embryogenic clumps have been shown to contain large amounts of arabinose side chains of RG-I and only poor amount of LM6-reactive pectins (Kikuchi et al. 1995; Willats et al. 1999; Majewska-Sawka and Münster 2003; Xu et al. 2011) and our results seem to confirm this. Aside from differences in the expression of RG-I eptitopes, our experiments revealed differences in the distribution pattern of low and high methyl-esterified HG during somatic embryo development. Similarly to the studies on citrus (Ramirez et al. 2003) and banana (Xu et al. 2011) the JIM5 epitope was the most abundant form of HG at early stages of embryo formation, but decreased with embryoid maturation as the highly methyl-esterified form became prevalent. However, whilst the JIM7 epitope was evenly distributed throughout the embryo body, the signal resulting from JIM5 binding was not detected in the protodermis of CsSE. This observation contrasts with reports of Bárány et al. (2010) and Dobrowolska et al. (2012) where the JIM7, but not the JIM5 epitope was absent from the protodermis of mature embryos. These discrepancies in the respective results are difficult to explain. Nevertheless, as was shown by Dobrowolska et al. (2012), specific culture conditions may have a strong impact on pectin composition and distribution in developing somatic embryos.
Abbreviations
- 2iP:
-
N6-[2-isopentenyl]-adenine
- AGP:
-
Arabinogalactan-protein
- BSA:
-
Bovine serum albumin
- EsSE:
-
Early-staged somatic embryos
- CsSE:
-
Cotyledonary stage somatic embryos
- CsZE:
-
Cotyledonary stage zygotic embryos
- ECMSN:
-
Extracellular matrix surface network
- EGTA:
-
Ethylene glycol-bis(β-aminoethyl ether)-N,N,N′N′-tetraacetic acid
- FITC:
-
Fluorescein isothiocyanate
- HG:
-
Homogalacturonan
- NAA:
-
1-Naphtaleneacetic acid
- PAS:
-
Periodic acid-Schiff’s reagent
- PBS:
-
Phosphate-buffered saline
- PIPES:
-
Piperazine-N,N-bis(2-ethanesulfonic acid)
- RG-I:
-
Rhamnogalacturonan-I
- SE:
-
Somatic embryogenesis
- SEM:
-
Scanning electron microscopy
References
Baluška F, Šamaj J, Wojtaszek P, Volkman D, Menzel D (2003) Cytoskeleton-plasma membrane-cell wall continuum in plants Emerging links revisited. Plant Physiol 133:482–491
Bárány I, Fadon B, Risueno MC, Testilano PS (2010) Cell wall components and pectin esterification levels as markers of proliferation and differentiation events during pollen development and pollen embryogenesis in Capsicum annuum L. J Exp Bot 61:1169–1175
Blehová A, Bobák M, Šamaj K, Hlinková E (2010) Changes in the formation of an extracellular matrix surface network during early stages of indirect somatic embyogenesis in Drosera spathulata. Acta Bot Hung 52:23–33
Bobák M, Šamaj J, Hlinková E, Hlavačka A, Ovečka M (2003/4) Extracellular matrix in early stages of direct somatic embryogenesis in leaves of Drosera spathulata. Biol Plant 47:161-166
Bobák M, Šamaj J, Pretová A, Blehová A, Hlinková E, Ovečka M, Kutarnová Z, Hlavačka A (2004) The histological analysis of indirect somatic embryogenesis on Drosera spathulata Labill. Acta Physiol Plant 26:353–361
Bush MS, McCann MC (1999) Pectic epitopes are differentially distributed in the cell walls of potato (Solanum tuberosum) tubers. Physiol Plant 107:201–213
Chapman A, Blervacq AS, Vasseur J, Hilbert JL (2000a) Arabinogalactan-proteins in Cichorium somatic embryogenesis: effect of β-glucosyl Yariv reagent and epitope localization during embryo development. Planta 211:305–314
Chapman A, Blervacq AS, Hendriks T, Slomianny C, Vasseur J, Hilbert JL (2000b) Cell wall differentiation during early somatic embryogenesis in plants. II. Ultrastructural study and pectin immunilocalization on chicory embryos. Can J Bot 78:824–831
Clausen MH, Willats WGT, Knox JP (2003) Synthetic methyl hexagalacturonate hapten inhibitors of anti-homogalacturonan monoclonal antibodies LM7, JIM5 and JIM7. Carbohydr Res 338:1797–1800
Cosgrove DJ (2005) Growth of the plant cell wall. Nat Rev Mol Cell Biol 6:850–861
Dobrowolska I, Majchrzak O, Baldwin T, Kurczynska EU (2012) Differences in protodermal cell wall structure in zygotic and somatic embryos of Daucus carota (L.) cultured on solid and in liquid media. Protoplasma 249:117–129
Dubois T, Guedira M, Dubois J, Vasseur J (1990) Direct somaticembryogenesis in roots of Cichorium: is callose an early marker? Ann Bot 65:539–545
Dubois T, Dubois J, Gudeira M, Diop A, Vasseur J (1992) SEM characterization of an extracellular matrix around somatic proembryos in roots of Cichorium. Ann Bot 70:119–124
Dumville JC, Fry SC (2000) Uronic acid-containing oligosaccharins: their biosynthesis, degradation and signalling roles in non-diseased plant tissues. Plant Physiol Biochem 38:125–140
Ellis M, Egelund J, Schultz CJ, Bacic A (2010) Arabinogalactan-proteins: key regulators at the cell surface? Plant Physiol 153:403–419
Endress V, Barriuso J, Ruperez P, Martin JP, Blazquez A, Villalobos N, Guerra H, Martin L (2009) Differences in cell wall polysaccharide composition between embryogenic and non-embryogenic calli of Medicago arborea L. Plant Cell Tissue Org Cult 97:323–329
Gao M, Showalter AM (1999) Yariv reagent treatment induces programmed cell death in Arabidopsis cell cultures and implicates arabinogalactan protein involvement. Plant J 19:321–331
Harholt J, Suttangkakul A, Scheller HV (2010) Biosynthesis of Pectin. Plant Physiol 153:384–395
Iwai H, Kikuchi A, Kobayashi T, Kamada H, Satoh S (1999) High levels of non-methylesterified pectins and low levels of peripherally located pectins in loosely attached non-embriogenic callus of carrot. Plant Cell Rep 18:561–566
Jarvis MC, Briggs SPH, Knox JP (2003) Intercellular adhesion and cell separation in plants. Plant Cell Environ 26:977–989
Jasik J, Salajova T, Salaj J (1995) Developmental anatomy and ultrastructure of early somatic embryos in European black pine (Pinus nigra Arn.). Protoplasma 185:205–211
Jones L, Seymour GB, Knox JP (1997) Localization of pectin galactan in tomato cell walls using a monoclonal antibody specific to (1→4)-β-D galactan. Plant Physiol 113:1405–1412
Kikuchi A, Satoh S, Nakamura N, Fujii T (1995) Differences in pectic polysaccharides between carrot embryogenic and nonembryogenic calli. Plant Cell Rep 14:279–284
Konieczny R, Bohdanowicz J, Czaplicki AZ, Przywara L (2005) Extracellular matrix surface network during plant regeneration in wheat anther culture. Plant Cell Tissue Org Cult 83:201–208
Konieczny R, Świerczyńska J, Czaplicki AZ, Bohdanowicz J (2007) Distribution of pectin and arabinogalactan protein epitopes during organogenesis from androgenic callus of wheat. Plant Cell Rep 26:355–363
Konieczny R, Pilarska M, Tuleja M, Salaj T, Ilnicki T (2010) Somatic embryogenesis and plant regeneration in zygotic embryos of Trifolium nigrescens (Viv.). Plant Cell Tissue Org Cult 100:123–130
Konieczny R, Sliwinska E, Pilarska M, Tuleja M (2012) Morphohistological and flow cytometric analyses of somatic embryogenesis in Trifolium nigrescens Viv. Plant Cell Tissue Organ Cult 109:131–141
Kreuger M, van Holst GJ (1995) Arabinogalactan-protein epitopes in somatic embryogenesis of Daucus carota L. Planta 127:135–141
Lai KS, Yusoff K, Maziah M (2011) Extracellular matrix as the early structural marker for Centella asiatica embryogenic tissues. Biol Plant 55:549–553
Majewska-Sawka A, Münster A (2003) Cell-wall antigens in mesophyll cells and mesophyll-derived protoplasts of sugar beet: possible implication in protoplast recalcitrance? Plant Cell Rep 21:946–954
McCartney L, Ormerod AP, Gidley MJ, Knox JP (2000) Temporal and spatial regulation of pectic (1→4)-β-D galactan in cell walls of developing pea cotyledons: implication for mechanical properties. Plant J 22:105–113
McCartney L, Steele-King CG, Jordan E, Knox JP (2003) Cell wall pectic (1→4)-β-D-galactan marks the acceleration of cell elongation in the Arabidopsis seedling root meristem. Plant J 33:447–454
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiol Plant 15:473–497
Pan X, Yang X, Lin G, Zou R, Chen H, Šamaj J, Xu C (2011) Ultrastructural changes and the distribution of arabinogalactan proteins during somatic embryogenesis of banana (Musa spp. AAA cv Yueyoukang 1). Physiol Plant 142:372–389
Pennell RI, Janniche L, Kjellbom P, Scofield GN, Peart JM, Roberts K (1991) Developmental regulation of a plasma membrane arabinogalactan protein epitope in oilseed rape flowers. Plant Cell 3:1317–1326
Pilarska M, Czaplicki AZ, Konieczny R (2007) Patterns of pectin epitope expression during shoot and root regeneration in androgenic cultures of two wheat cultivars. Acta Biol Cracov Ser Bot 49:69–72
Popielarska-Konieczna M, Kozieradzka-Kiszkurno M, Świerczyńska J, Góralski G, Ślesak H, Bohdanowicz J (2008) Ultrastructure and histochemical analysis of extracellular matrix surface network in kiwifruit endosperm-derived callus culture. Plant Cell Rep 27:1137–1145
Ramirez C, Chiancone B, Testilano PS, Garcia-Fojeda B, Germana MA, Risueno MC (2003) First embryogenic stages of Citrus microspore-derived embryos. Acta Biol Crac Ser Bot 45:53–58
Rumyantseva NI, Šamaj J, Ensikat HJ, Salnikov VV, Kostyukova YA, Baluška F, Volkmann D (2003) Changes in the extracellular matrix surface network during cyclic reproduction of proembryonic cell complexes in the Fagopyrum tataricum (L.) gaertn callus. Dokl Biol Sci 391:375–378
Saare-Surminski K, Preil W, Knox JP, Lieberei R (2000) Arabinogalactan proteins in embryogenic and non-embryogenic callus cultures of Euphorbia pulcherrima. Physiol Plant 108:180–187
Šamaj J, Bobák M, Blehová A, Krińtin J, Auxtova- Šamajova O (1995) Developmental SEM observations of an extracellular matrix in embryogenic calli of Drosera rotundifolia and Zea mays. Protoplasma 186:45–49
Šamaj J, Baluška F, Bobák M, Volkman D (1999a) Extracellular matrix surface network of embryogenic units of friable maize callus contains arabinogalactan-proteins recognized by monoclonal antibody JIM4. Plant Cell Rep 18:369–374
Šamaj J, Ensikat HJ, Baluška F, Knox JP, Barthlott W, Volkmann D (1999b) Immunogold localization of plant surface arabinogalactan-proteins using glycerol liquid substitution and scanning electron microscopy. J Microsc 193:150–157
Šamaj J, Bobák M, Blehová A, Pretová A (2006) Importance of cytoskeleton and cell wall in somatic embryogenesis. In: Mujib A, Šamaj J (eds) Somatic embryogenesis in plants: new approaches Plant Cell Monographs. Springer, Heidelberg, pp 35–50
Schultz CJ, Johnson KL, Currie G, Bacic A (2000) The classical arabinogalactan protein gene family of Arabidopsis. Plant Cell 12:1751–1767
Seifert GJ, Blaukopf C (2010) Irritable walls: the plant extracellular matrix and signaling. Plant Physiol 153:467–478
Seifert GJ, Roberts K (2007) The biology of arabinogalactan proteins. Ann Rev of Plant Biol 58:137–161
Serpe MD, Nothnagel EA (1994) Effect of Yariv phenylglykosides on Rosa cell-suspensions: evidence for the involvement of arabinogalactan-proteins in cell proliferation. Planta 193:542–550
Showalter AM (2001) Arabinogalactan-proteins: structure, expression and function. Cell Mol Life Sci 58:1399–1417
Sondahl MR, Salisbury JL, Sharp WR (1979) SEM characterization of embryogenic tissue and globular embryos during high-frequency somatic embryogenesis in coffee callus cells. Z Pflanzenphysiol 94:185–188
Stacey NJ, Roberts K, Knox JP (1990) Patterns of expression of the JIM4 arabinogalactan-protein epitope in cell cultures and during somatic embryogenesis in Daucus carota L. Planta 180:285–292
Steinmacher DA, Saare-Surminski K, Lieberei R (2012) Arabinogalactan proteins and the extracellular matrix surface network during peach palm somatic embryogenesis. Physiol Plant 146:336–349
Toonen MAJ, Schmidt EDL, Hendriks T, van Kammen A, de Vries SC (1997) Promotive and inhibitory effects of diverse arabinogalactan proteins on Daucus carota L. somatic embryogenesis. Planta 203:188–195
Verdeil JL, Hocher V, Huet C, Grosdemange F, Escoute J, Ferriere N, Nicole M (2001) Ultrastructural changes in coconut calli associated with the acquisition of embryogenic competence. Ann Bot 88:9–18
Verhertbruggen Y, Knox JP (2006) Pectic polysaccharides and expanding cell walls. In: Verleben JP, Vissenberg K (eds) The expanding cell. Springer–Berlin pp 139-158
Willats WGT, Knox JP (1996) A role for arabinogalactan-proteins in plant cell expansion: evidence from studies on the interaction of β-glucosyl Yariv reagent with seedlings of Arabidopsis thaliana. Plant J 9:919–925
Willats WGT, Marcus SE, Knox JP (1998) Generation of a monoclonal antibody specific to (1→5)-a-L-arabinan. Carbohyd Res 308:149–152
Willats WGT, Steele-King CG, Marcus SE, Knox JP (1999) Side chains of pectic polysaccharides are regulated in relation to cell proliferation and cell differentiation. Plant J 20:619–628
Xu C, Zhao L, Pan X, Šamaj J (2011) Developmental localization and methylesterification of pectin epitopes during somatic embryogenesis of banana (Musa ssp. AAA). PLoS ONE 6:e22992
Yates EA, Valdor JF, Haslam SM, Morris HR, Dell A, Mackie W, Knox JP (1996) Characterization of carbohydrate structural features recognized by anti-arabinogalactan-protein monoclonal antibodies. Glycobiology 6:131–139
Acknowledgments
The Institute of Genetics and Crop Plant Research (Gatersleben, Germany) is acknowledged for providing T. nigrescens seeds. The SEM images were made in the Laboratory of Field Emission Scanning Electron Microscopy and Microanalysis at the Institute of Geological Sciences of the Jagiellonian University. This project was supported by Polish Ministry of Higher Education (N N303 410236). MP was funded by British-Polish Young Scientists Programme (YSP) of British Council and Polish Ministry of Science and Higher Education (WAR/342/127).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Pilarska, M., Knox, J.P. & Konieczny, R. Arabinogalactan-protein and pectin epitopes in relation to an extracellular matrix surface network and somatic embryogenesis and callogenesis in Trifolium nigrescens Viv.. Plant Cell Tiss Organ Cult 115, 35–44 (2013). https://doi.org/10.1007/s11240-013-0337-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11240-013-0337-8