
Abstract
Galectin-3 is a member of the galectin family, widely expressed in immune cells and plays a role mainly in inflammation, autoimmunity, apoptosis, and chemotaxis. We summarized the roles of Galectin-3 in diabetes and its complications, as well as the underlying mechanisms. Clinical research has determined that the circulating level of Galectin-3 is closely related to diabetes and its complications, thus it is promising to use Galectin-3 as a predictor and biomarker for those diseases. Galectin-3 also may be considered as an ideal therapeutic target, which has broad prospects in the prevention and treatment of diabetes and its complications, especially macrovascular and microvascular complications.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Diabetes mellitus (DM) is a public health problem worldwide. It was reported that the global diabetes prevalence in 2019 was estimated to be 463 million people, rising to 578 million by 2030 and 700 million by 2045 [1,2,3,4]. Meta-inflammation theory suggests that both type 1 diabetes mellitus (T1D) and type 2 diabetes mellitus (T2D) are chronic inflammatory diseases. T1D involves inflammation in pancreatic islet tissue, while inflammation in adipose tissue result in T2D, which are attributed to the imbalance of pro-inflammatory and anti-inflammatory cells and molecules [5, 6].
Galectin-3, encoded by LGALS3 gene, is a member of β-galactoside-binding lectin family subtype of galectin, formerly known as carbohydrate-binding protein-35 (CBP-35) [7,8,9]. Human Galectin-3 is a 26 kDa size lectin, mainly comprises a C-terminal carbohydrate recognition binding domain (CRD) and N-terminal domain [10,11,12]. Galectin-3 is widely distributed in the tissues such as human hematopoietic tissues, thymus, lymph nodes, spleen, and mainly produced by immune cells such as macrophages, mast cells, and eosinophils, et al., and can be secreted extracellularly [13, 14]. It is regarded as a powerful pro-inflammatory signaling factor, which plays an important role in the activation, chemotaxis, and cytokine release of inflammatory cells. Increasing evidence has proven that levels of circulating Galectin-3 are elevated in chronic inflammatory diseases including obesity, diabetes and its complications, suggesting that Galectin-3 is closely related to those disease status [15]. Thus, clarifying the potential pathogenic mechanism of Galectin-3 in diabetes and diabetic complications will provide a basis for finding new biomarkers for precision disease prediction and early-diagnosis, and developing potential therapeutic targets.
Therefore, we reviewed the clinical evidence of Galectin-3 in diabetes and its complications and discussed the potential mechanism of such relevance. Furthermore, further perspectives and potential directions were provided, which can be used as a reference for further clinical research and scientific foundation for new drug development.
2 Galectin-3 in type 2 diabetes
T2D is a chronic inflammatory disease, characterized by obesity-associated insulin resistance [16,17,18,19,20]. Autoimmunity plays a pivotal part in a series of chronic diseases [21,22,23]. Several clinical studies reported that circulating level of Galectin-3 was significantly higher in T2D patients [24,25,26,27,28] (Table 1). Weigert et al. [24] found that serum Galectin-3 level was significantly higher in T2D patients. Interestingly, they reported that Galectin-3 level reduced by metformin treatment, the mechanism of which may be metformin lowered oxidative stress and the formation of advanced glycation end products (AGEs), and then decreased Galectin-3 expression. In 2019, a cross-sectional study conducted by Atalar et al. [25] enrolling healthy control, prediabetes and T2D, found that serum Galectin-3 increased in T2D but not prediabetes and healthy controls, and had a positive correlation with FPG and HbA1c, as well as hs-CRP, whose level was a useful biomarker for early estimating the chronic inflammatory process in diabetes. The findings of this study suggested that Galectin-3 may play a role in the progression of prediabetes stage to diabetes stage. In summary, all current human studies provided strong evidence of Galectin-3 as a marker, even a pathogenic factor, of T2D.
Efforts on mechanism investigation also have been made both in cell studies and animal studies. Li et al. [29] showed that the genetic depletion of Galectin-3 improved both systemic and tissue insulin sensitivity, and glucose tolerance in HFD mice, indicating Galectin-3 induces insulin resistance and deteriorates glucose homeostasis. Mechanically, they found that elevated circulating Galectin-3 from macrophages directly interacted with insulin receptor, impaired the major steps of insulin signaling pathway, inducing cellular insulin resistance in adipose tissue, liver and muscle. Petrovic et al. [30] further demonstrated that pancreatic Galectin-3 overexpression promoted β-cell apoptosis triggered by cytokines and palmitate, and increased NO2-induced oxidative stress in β cells, suggesting that Galectin-3 participants in β-cell damage and insulitis in HFD-induced T2D. Yu et al. [31] reported that the inhibition of Galectin-3 by TD139, an effective inhibitor of the galactoside binding pocket of Galectin-3, improved insulin resistance in HFD-induced mice, suggesting the potential of TD139 in the treatment of T2D.
In conclusion, serum Galectin-3 is elevated in T2D patients, and the elevated Galectin-3 plays a role in pathogenesis of T2D by disturbing insulin signaling pathway in insulin-targeting organs and inducing β-cell inflammation and death in pancreatic islets. Thus, it is promising to use Galectin-3 as a predictive biomarker and therapeutic target for T2D.
3 Galectin-3 in type 1 diabetes
In recent years, the incidence of T1D has increased rapidly at a rate of 3%-5% per year worldwide. T1D is an autoimmune disease caused by the selective destruction of insulin-producing β cells. Damaged β cells release their own antigens and present them to autoreactive T cells in the pancreatic draining lymph nodes (PLN) [32, 33]. These activated autoreactive T cells migrate to the pancreatic islets, thereby promoting inflammation and mediating β-cell apoptosis [34,35,36].
Early studies have shown that Galectin-3 plays a pivotal role in autoimmune diseases including autoimmune encephalomyelitis (EAE) [37], rheumatoid arthritis (RA) [38] and systemic sclerosis [39], et al. Karlsen, A.E. et al. identified six polymorphisms in the Galectin-3 gene (LGALS3) and proved that a haplotype containing three SNPs transmitted to unaffected offspring in 257 T1D families increased significantly. This finding was verified in an independent set of 170 T1D families, suggesting that LGALS3 is a T1D susceptibility gene, providing genetic association between Galectin-3 and T1D [40].
The destruction of pancreatic β-cells in T1D is mainly due to disorders of adaptive immune cells as well as innate immune cells [41]. Macrophage is the first-line cell to invade the pancreatic islets, and acts as an antigen presenting cell and an effector cell, contributing to the pathogenesis and development of autoimmune diabetes [42,43,44,45]. A study in vitro showed that Galectin-3 chemoattracts monocytes and macrophages through the G protein coupling pathway [46]. Galectin-3 involves in the migration of neutrophils to sites of infection or inflammation, activates neutrophils and promotes their adhesion [47, 48]. If the infection persists and is not controlled by neutrophils, Galectin-3 recruits macrophages to the site of infection, strengthening further defenses [49, 50]. Galectin-3 also participates in the innate immune response and is considered to be a regulator of T cell activation [51], thus promoting pro-inflammatory cytokines which up-regulate the apoptotic signal [52, 53]. Cytokines can increase the expression of Galectin-3 mRNA, making it a vicious circle process [54].
In vitro data provided that the expression of Galectin-3 was upregulated in BB-DP islets after IL-1β exposure, however, whether Galectin-3 elevation is a causal factor leading to apoptosis, or a compensatory effect to combat apoptosis is unclear [55]. Saksida et al. [52] proved that genetic deletion or pharmacological inhibition of Galectin-3 preserved β cell function and promoted survival by reducing mitochondrial apoptotic pathway triggered by pro-inflammatory cytokines, indicating the pro-inflammatory property of Galectin-3 in pancreatic β cell apoptosis. The main mechanism is that Galectin-3 downregulates the expression of major genes in this apoptotic pathway and upregulates the expression of anti-apoptotic genes [52, 55]. By analyzing TD139-treated RINm5F cells, TD139 significantly ameliorates cell apoptosis by Galectin-3 under the action of various cytokines [56]. In contrast, an in vitro study showed that overexpression of Galectin-3 in RIN-cell inhibited cytokine mediated apoptosis, suggesting the protective role of Galectin-3 against cytokine toxicity induced β cell apoptosis [40]. Indeed, different location of Galectin-3 exerts various functions, for example, cytoplasmic Galectin-3 has anti-apoptotic activity maintaining mitochondrial integrity, whereas nuclear and extracellular Galectin-3 plays pro-apoptotic role [57], which may explain the controversial results.
Animal studies also conducted to investigate whether Galectin-3 contributes to the pathogenesis of T1D. Mensah-Brown et al. demonstrated that the lack of Galectin-3 protected C57BL/6 mice from MLD-STZ induced diabetes, accompanied with significant less mononuclear infiltration in the pancreatic islets and with retention of higher insulin content when compared with WT mice. Macrophages in the abdominal cavity and pancreatic draining lymph nodes from Galectin-3 knockout mice expressed lower interferon-γ (IFN-γ), TNF-α, IL-17 and inducible nitric oxide synthesis (iNOS) than those from WT mice [58]. On summary, the studies above confirmed that both the deletion of Galectin-3 gene or pharmacological inhibition reduces the apoptosis of pancreatic β-cells. Therefore, Galectin-3 plays a role in the occurrence of β-cell destruction in T1D and is a promising therapeutic target for T1D patients.
4 Galectin-3 in diabetes complications
The most common complication of diabetes is vascular disease, which is also the main cause of morbidity and mortality in diabetic patients [59]. Vascular complications are mainly divided into macrovascular complications including cerebrovascular, cardiovascular and peripheral vascular disease, and microvascular complications including retinopathy, nephropathy, and diabetic foot [60].
4.1 Macrovascular complications
The major vascular complication of diabetes is the accelerated formation of atherosclerosis, which involves important blood vessels in the body, such as coronary arteries, carotid arteries, and peripheral arteries [61]. Galectin-3 plays a pro-inflammatory role in the occurrence and development of atherosclerosis and can be used as a risk factor for atherosclerosis [62].
Ozturk et al. performed coronary CT scans on 158 patients with T2D and reported that Galectin-3 was positively correlated with the number of coronary arteries with atherosclerosis [63]. Saeed et al. followed up a population-based nationwide cohort in Norway, and found that after the adjustment of conventional risk factors, Galectin 3 was significantly associated with the risk of future coronary heart disease (CHD) and may help to predict the occurrence of CHD in patients with T1D [64]. These findings suggest that Galectin-3 could be a predictor for coronary atherosclerosis in patients with T1D or T2D.
4.2 Microvascular complications
4.2.1 Diabetic nephropathy
Diabetic nephropathy is the main cause of end-stage renal disease in diabetic patients [65, 66]. A prospective study involving 1320 cases of T2D with an average follow-up of 9 years showed that the level of serum Galectin-3 was independently associated with the progression of diabetic nephropathy [67]. Compared with patients with baseline serum Galectin-3 levels in the lower quarter, patients with Galectin-3 levels in the upper quarter had a three-fold increase in the risk of renal function decline and a two-fold increase in the risk of massive proteinuria [67]. The results of a single-center prospective clinical study demonstrated that Galectin-3 levels are elevated in diabetic nephropathy and are positively correlated with the urine albumin/creatinine ratio [68]. These findings suggest that increased circulating Galectin-3 may be predictive of poor prognosis in diabetic nephropathy.
4.2.2 Diabetic retinopathy
Approximately two-thirds of diabetic patients have diabetic retinopathy. Studies have found that Galectin-3 is related to the pathogenesis of diabetic retinopathy. Galectin-3 may affect visual function during diabetes by combining with AGEs [69]. An animal study by Bauer et al. on lipopolysaccharide-induced neuroinflammation found that Galectin-3 is expressed in both Müller cells and microglia/macrophages of the normal retina; and demonstrated that increase in Galectin-3 expression was accompanied by significant increased neurotoxicity in the explanted retina [70]. Abreu et al. showed that the damage to optic nerve fibers caused by microglia/macrophages was reduced in Galectin-3 knockout mice [71]. Compared with the microglia of wild-type mice, the microglia of Galectin-3 knockout mice were less pro-inflammatory, resulting in excess white matter [72] and reduced retinal ganglion cell apoptosis [71]. The cause may be that iNOS is less activated in the optic nerve of Galectin-3 knockout mice [73]. Stitt et al. [74] infused preformed AGEs into wild-type and Galectin-3 knockout mice and found that, in oxygen-induced proliferative retinopathy, Galectin-3 knockout mice have reduced retinal ischemia and higher retinal angiogenesis potential. Therefore, Galectin-3 seems to be a pivotal molecule triggering neurodegenerative, oxidative and inflammatory processes preceding vascular modifications in diabetic retinopathy, and its modulation might be a useful tool to prevent diabetic visual complications.
4.2.3 Diabetic foot
Diabetic foot is one of the long-term complications of diabetes, and 50%-60% of diabetic foot patients have some degree of peripheral artery disease [75, 76]. Studies have reported that Galectin-3 is associated with the occurrence of diabetic foot ulcers [75, 77]. Galectin-3 promotes blood vessel formation and new blood vessel formation because Galectin-3 increases the level of VEGF-A [78, 79]; thus, Galectin-3 promotes the healing of diabetic foot ulcers (DFUs). In a prospective study with 30 healthy controls, 30 patients diagnosed with T2D but without DFUs and evident peripheral artery disease (PAD) as the diabetic control group and 31 patients diagnosed with T2D with DFUs at Wagner stage 2–4, serum Galectin-3 levels together with VEGF-A were significantly higher in T2D with DFUs compared with the other two groups, indicating as a defense mechanism against DFUs, thus contributing to wound healing [77]. When chronic skin damage occurs, Galectin-3 expression is inversely correlated with the level of AGEs in diabetic or non-diabetic patients, suggesting that Galectin-3 helps to reduce the accumulation of AGEs in the skin wound [80]. These studies provide an insight about Galectin-3 as an emerging biomarker which may give an indication of prognosis in diabetic foot with special reference to DFUs.
4.3 Diabetic cardiomyopathy
Diabetic cardiomyopathy is the main cause of heart failure in diabetes patients. Jelena et al. enrolled 189 participants with T2D and/or arterial hypertension (HT), and found that levels of galectin-3 were higher in patients with both T2D and HT, and correlated with left ventricular (LV) mass, indicating the potential role of Galectin-3 for early detection of myocardial structural and functional alterations [81]. Bolotskykh et al. found that non-diabetic patients with heart failure are accompanied by insulin resistance, and the levels of Galectin-3, TNF-α and insulin in such heart failure patients are significantly increased [82]. Myocardial inflammation and fibrosis are accompanied by changes in the Galectin-3 concentration [83]. Further studies confirmed that the mechanism of inducing cardiomyocyte changes including hypertrophy, fibrosis, increased stiffness, and impaired relaxation is triggered by the pro-inflammatory environment caused by adipose tissue dysfunction, and Galectin-3 can be used as a biomarker for changes in myocardial function and fibrosis caused by high calories [84, 85]. This finding shows that Galectin-3 is involved in the development of cardiac fibrosis and impaired myocardium remodeling, resulting in heart failure and atrial fibrillation [85]. The role of Galectin-3 as a prognostic marker of heart failure is described, and the tentative use of Galectin-3 inhibition is a potential therapeutic approach to prevent cardiac inflammation and fibrosis.
5 Further perspectives
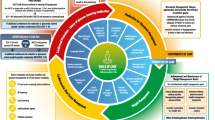
As a powerful cytokine related to inflammation, autoimmunity, apoptosis, and chemotaxis, Galectin-3 is involved in several metabolic abnormalities, mainly in diabetes and diabetic complications (Fig. 1). A large number of clinical studies have shown that Galectin-3 is related to the occurrence of diabetes and its complications. The main conclusion is that the level of serum Galectin-3 is higher in patients with diabetes and its complications.

The role of galectin-3 in diabetes. In type 1 diabetes, macrophage-derived Galectin-3 participates in β cell death by recruiting and activating innate immune cells, including neutrophils and macrophages, to islets; In type 2 diabetes, Galectin-3 directly interacted with insulin receptor, impaired the major steps of insulin signaling pathway, inducing cellular insulin resistance in adipose tissue, liver and muscle
There are also a large number of animal experiments revealing that Galectin-3 promotes the inflammation of pancreatic islet β cells and insulin target organs, leading to pancreatic β cell failure and insulin resistance, which in turn leads to diabetes. According to existing evidence, Galectin-3 inhibitors such as TD139 can reduce the expression of Galectin-3, thereby improving diabetes and its complications. However, whether Galectin-3 can be used in clinical medicine still needs to be resolved.
To fill in the current gaps, the following research including: 1) To further clarify the specific mechanism of Galectin-3 in diabetes and remove obstacles for clinical research. 2) To develop safe and effective Galectin-3 inhibitors for the prevention and treatment of diabetes are warrant.
Abbreviations
- AGEs:
-
Advanced glycation end products
- BMI:
-
Body mass index
- CEA:
-
Carcinoembryonic antigen
- CHD:
-
Coronary heart disease
- CPB-35:
-
Carbohydrate-binding protein-35
- CRD:
-
Carbohydrate recognition binding domain
- CRP:
-
C reactive protein
- CVD:
-
Cardiovascular diseases
- DFUs:
-
Diabetic foot ulcers
- DM:
-
Diabetes mellitus
- DXA:
-
Dual-energy x-ray absorptiometry
- HFD:
-
High fat diet
- IRS1:
-
Insulin receptor substrate 1
- JNK:
-
C-Jun-N-terminal kinase
- NASH:
-
Nonalcoholic steatohepatitis
- NLRP3:
-
NOD-like receptor family pyrin domain protein 3
- LAMP-1:
-
Lysosomal-associated membrane proteins 1
- LAMP-2:
-
Lysosomal-associated membrane proteins 2
- LncRNA:
-
Long non-coding RNA
- LPS:
-
Lipopolysaccharide
- OGTT:
-
Oral glucose tolerance test
- PAD:
-
Peripheral artery disease
- PPAR:
-
Peroxisome proliferator-activated receptor
- T1D:
-
Type 1 diabetes mellitus
- T2D:
-
Type 2 diabetes mellitus
References
Saeedi P, Petersohn I, Salpea P, Malanda B, Karuranga S, Unwin N, et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res Clin Pract. 2019;157: 107843. https://doi.org/10.1016/j.diabres.2019.107843.
Li T, Providencia R, Mu N, Yin Y, Chen M, Wang Y, et al. Association of metformin monotherapy or combined therapy with cardiovascular risks in patients with type 2 diabetes mellitus. Cardiovasc Diabetol. 2021;20:30. https://doi.org/10.1186/s12933-020-01202-5.
Zhang F, Liu L, Zhang C, Ji S, Mei Z, Li T. Association of metabolic syndrome and Its components with risk of stroke recurrence and mortality: a meta-analysis. Neurology. 2021;97:e695–705. https://doi.org/10.1212/WNL.0000000000012415.
Ceriello A, Catrinoiu D, Chandramouli C, Cosentino F, Dombrowsky AC, Itzhak B, et al. Heart failure in type 2 diabetes: current perspectives on screening, diagnosis and management. Cardiovasc Diabetol. 2021;20:218. https://doi.org/10.1186/s12933-021-01408-1.
Wang Y, Xiao Y, Zhong L, Ye D, Zhang J, Tu Y, et al. Increased neutrophil elastase and proteinase 3 and augmented NETosis are closely associated with β-cell autoimmunity in patients with type 1 diabetes. Diabetes. 2014;63:4239–48. https://doi.org/10.2337/db14-0480.
Xiao Y, Xu A, Law LS, Chen C, Li H, Li X, et al. Distinct changes in serum fibroblast growth factor 21 levels in different subtypes of diabetes. J Clin Endocrinol Metab. 2012;97:E54–8. https://doi.org/10.1210/jc.2011-1930.
Maurice B, Amel AE, Dennis B, Marie E, Zoubida K, Martine MJ, et al. Exploring the role of galectin 3 in kidney function: a genetic approach. Glycobiology. 2006;16:36–45.
Roff CF, Wang JL. Endogenous lectins from cultured cells. Isolation and characterization of carbohydrate-binding proteins from 3T3 fibroblasts. J Biol Chem. 1983;258:10657–63.
Hoverfelt A, Sallinen R, Soderlund JM, Forsblom C, Pettersson-Fernholm K, Parkkonen M, et al. DDOST, PRKCSH and LGALS3, which encode AGE-receptors 1, 2 and 3, respectively, are not associated with diabetic nephropathy in type 1 diabetes. Diabetologia. 2010;53:1903–7. https://doi.org/10.1007/s00125-010-1771-3.
Gong HC, Honjo Y, Nangia-Makker P, Hogan V, Mazurak N, Bresalier RS, Raz A. The NH2 terminus of galectin-3 governs cellular compartmentalization and functions in cancer cells. Cancer Res. 1999;59(24):6239–45.
Barondes SH, Cooper DN, Gitt MA, Leffler H. Galectins: Structure and function of a large family of animal lectins. J Biol Chem. 1994;269:20807–10. https://doi.org/10.1016/s0021-9258(17)31891-4.
Johannes L, Jacob R, Leffler H. Galectins at a glance. J Cell Sci. 2018;131. https://doi.org/10.1242/jcs.208884.
Yang RY, Rabinovich GA, Liu FT. Galectins: structure, function and therapeutic potential. Expert Rev Mol Med. 2008;10: e17. https://doi.org/10.1017/S1462399408000719.
Suthahar N, Meijers WC, Sillje HHW, Ho JE, Liu FT, de Boer RA. Galectin-3 Activation and Inhibition in Heart Failure and Cardiovascular Disease: An Update. Theranostics. 2018;8:593–609. https://doi.org/10.7150/thno.22196.
Yang RY, Hsu DK, Liu FT. Expression of galectin-3 modulates T-cell growth and apoptosis. Proc Natl Acad Sci U S A. 1996;93:6737–42.
Bryk-Wiazania AH, Undas A. Hypofibrinolysis in type 2 diabetes and its clinical implications: from mechanisms to pharmacological modulation. Cardiovasc Diabetol. 2021;20:191. https://doi.org/10.1186/s12933-021-01372-w.
Ceriello A, Prattichizzo F. Variability of risk factors and diabetes complications. Cardiovasc Diabetol. 2021;20:101. https://doi.org/10.1186/s12933-021-01289-4.
Mosenzon O, Del Prato S, Schechter M, Leiter LA, Ceriello A, DeFronzo RA, et al. From glucose lowering agents to disease/diabetes modifying drugs: a SIMPLE approach for the treatment of type 2 diabetes. Cardiovasc Diabetol. 2021;20:92. https://doi.org/10.1186/s12933-021-01281-y.
Soltani S, Mansouri K, Parvaneh S, Thakor AS, Pociot F, Yarani R. Diabetes complications and extracellular vesicle therapy. Rev Endocr Metab Disord. 2021. https://doi.org/10.1007/s11154-021-09680-y.
Konkwo C, Perry RJ. Imeglimin: current development and future potential in type 2 diabetes. Drugs. 2021;81:185–90. https://doi.org/10.1007/s40265-020-01434-5.
Xiao C, Nemazee D, Gonzalez-Martin A. MicroRNA control of B cell tolerance, autoimmunity and cancer. Semin Cancer Biol. 2020;64:102–7. https://doi.org/10.1016/j.semcancer.2019.04.004.
Khan S, Gerber DE. Autoimmunity, checkpoint inhibitor therapy and immune-related adverse events: A review. Semin Cancer Biol. 2020;64:93–101. https://doi.org/10.1016/j.semcancer.2019.06.012.
Muller I, Barrett-Lee PJ. The antigenic link between thyroid autoimmunity and breast cancer. Semin Cancer Biol. 2020;64:122–34. https://doi.org/10.1016/j.semcancer.2019.05.013.
Weigert J, Neumeier M, Wanninger J, Bauer S, Farkas S, Scherer MN, et al. Serum galectin-3 is elevated in obesity and negatively correlates with glycosylated hemoglobin in type 2 diabetes. J Clin Endocrinol Metab. 2010;95:1404–11. https://doi.org/10.1210/jc.2009-1619.
Atalar MN, Abusoglu S, Unlu A, Tok O, Ipekci SH, Baldane S, et al. Assessment of serum galectin-3, methylated arginine and Hs-CRP levels in type 2 diabetes and prediabetes. Life Sci. 2019;231: 116577. https://doi.org/10.1016/j.lfs.2019.116577.
Lin D, Hong X, Sun K, Zhang X, Lian H, Wang J, Mao N, Zhang X, Ren M, Yan L, Li F. Galectin-3/adiponectin as a new biological indicator for assessing the risk of type 2 diabetes: a cross-sectional study in a community population. Aging. 2021;13(11):15433.
Ohkura T, Fujioka Y, Nakanishi R, Shiochi H, Sumi K, Yamamoto N, Matsuzawa K, Izawa S, Ohkura H, Ueta E, Kato M. Low serum galectin-3 concentrations are associated with insulin resistance in patients with type 2 diabetes mellitus. Diabetol Metab Syndr. 2014;6(1):1–7.
Vora A, de Lemos JA, Ayers C, Grodin JL, Lingvay I. Association of Galectin-3 With Diabetes Mellitus in the Dallas Heart Study. J Clin Endocrinol Metab. 2019;104:4449–58. https://doi.org/10.1210/jc.2019-00398.
Li P, Liu S, Lu M, Bandyopadhyay G, Oh D, Imamura T, et al. Hematopoietic-Derived Galectin-3 Causes Cellular and Systemic Insulin Resistance. Cell. 2016;167(973–84): e12. https://doi.org/10.1016/j.cell.2016.10.025.
Petrovic I, Pejnovic N, Ljujic B, Pavlovic S, Miletic Kovacevic M, Jeftic I, et al. Overexpression of Galectin 3 in Pancreatic beta Cells Amplifies beta-Cell Apoptosis and Islet Inflammation in Type-2 Diabetes in Mice. Front Endocrinol (Lausanne). 2020;11:30. https://doi.org/10.3389/fendo.2020.00030.
Yu H, Yang F, Zhong W, Jiang X, Zhang F, Ji X, et al. Secretory Galectin-3 promotes hepatic steatosis via regulation of the PPARgamma/CD36 signaling pathway. Cell Signal. 2021;84: 110043. https://doi.org/10.1016/j.cellsig.2021.110043.
Parente EB, Harjutsalo V, Forsblom C, Groop PH, FinnDiane Study G. The impact of central obesity on the risk of hospitalization or death due to heart failure in type 1 diabetes: a 16-year cohort study. Cardiovasc Diabetol. 2021;20:153. https://doi.org/10.1186/s12933-021-01340-4.
Wu N, Bredin SSD, Jamnik VK, Koehle MS, Guan Y, Shellington EM, et al. Association between physical activity level and cardiovascular risk factors in adolescents living with type 1 diabetes mellitus: a cross-sectional study. Cardiovasc Diabetol. 2021;20:62. https://doi.org/10.1186/s12933-021-01255-0.
Wallberg M, Cooke A. Immune mechanisms in type 1 diabetes. Trends Immunol. 2013;34:583–91. https://doi.org/10.1016/j.it.2013.08.005.
Atkinson MA, Eisenbarth GS, Michels AW. Type 1 diabetes. Lancet. 2014;383:69–82. https://doi.org/10.1016/S0140-6736(13)60591-7.
Roep BO, Tree TIM. Immune modulation in humans: implications for type 1 diabetes mellitus. Nat Rev Endocrinol. 2014;10:229–42.
Jiang HR, Al Rasebi Z, Mensah-Brown E, Shahin A, Xu D, Goodyear CS, et al. Galectin-3 deficiency reduces the severity of experimental autoimmune encephalomyelitis. J Immunol. 2009;182:1167–73. https://doi.org/10.4049/jimmunol.182.2.1167.
Neidhart M, Zaucke F, von Knoch R, Jungel A, Michel BA, Gay RE, et al. Galectin-3 is induced in rheumatoid arthritis synovial fibroblasts after adhesion to cartilage oligomeric matrix protein. Ann Rheum Dis. 2005;64:419–24. https://doi.org/10.1136/ard.2004.023135.
Faludi R, Nagy G, Tokes-Fuzesi M, Kovacs K, Czirjak L, Komocsi A. Galectin-3 is an independent predictor of survival in systemic sclerosis. Int J Cardiol. 2017;233:118–24. https://doi.org/10.1016/j.ijcard.2016.12.140.
Karlsen AE, Storling ZM, Sparre T, Larsen MR, Mahmood A, Storling J, et al. Immune-mediated beta-cell destruction in vitro and in vivo-A pivotal role for galectin-3. Biochem Biophys Res Commun. 2006;344:406–15. https://doi.org/10.1016/j.bbrc.2006.03.105.
Donath MY, Storling J, Berchtold LA, Billestrup N, Mandrup-Poulsen T. Cytokines and beta-cell biology: from concept to clinical translation. Endocr Rev. 2008;29:334–50. https://doi.org/10.1210/er.2007-0033.
Mensah-Brown E, Shahin A, Parekh K, Hakim AA, Shamisi MA, Hsu DK, et al. Functional capacity of macrophages determines the induction of type 1 diabetes. Ann N Y Acad Sci. 2006;1084:49–57. https://doi.org/10.1196/annals.1372.014.
Kolb H, Burkart V, Appels B, Hanenberg H, Kantwerk-Funke G, Kiesel U, et al. Essential contribution of macrophages to islet cell destruction in vivo and in vitro. J Autoimmun. 1990;3(Suppl 1):117–20. https://doi.org/10.1016/s0896-8411(09)90020-8.
Lukic ML, Stosic-Grujicic S, Shahin A. Effector mechanisms in low-dose streptozotocin-induced diabetes. Dev Immunol. 1998;6:119–28. https://doi.org/10.1155/1998/92198.
Homo-Delarche F, Drexhage HA. Immune cells, pancreas development, regeneration and type 1 diabetes. Trends Immunol. 2004;25:222–9. https://doi.org/10.1016/j.it.2004.02.012.
Papaspyridonos M, McNeill E, de Bono JP, Smith A, Burnand KG, Channon KM, et al. Galectin-3 is an amplifier of inflammation in atherosclerotic plaque progression through macrophage activation and monocyte chemoattraction. Arterioscler Thromb Vasc Biol. 2008;28:433–40. https://doi.org/10.1161/ATVBAHA.107.159160.
Hsu DK, Yang RY, Pan Z, Yu L, Salomon DR, Fung-Leung WP, et al. Targeted disruption of the galectin-3 gene results in attenuated peritoneal inflammatory responses. Am J Pathol. 2000;156:1073–83. https://doi.org/10.1016/S0002-9440(10)64975-9.
Sato S, Ouellet N, Pelletier I, Simard M, Rancourt A, Bergeron MG. Role of galectin-3 as an adhesion molecule for neutrophil extravasation during streptococcal pneumonia. J Immunol. 2002;168:1813–22. https://doi.org/10.4049/jimmunol.168.4.1813.
Bergeron Y, Ouellet N, Deslauriers AM, Simard M, Olivier M, Bergeron MG. Cytokine kinetics and other host factors in response to pneumococcal pulmonary infection in mice. Infect Immun. 1998;66:912–22. https://doi.org/10.1128/IAI.66.3.912-922.1998.
Fillion I, Ouellet N, Simard M, Bergeron Y, Sato S, Bergeron MG. Role of chemokines and formyl peptides in pneumococcal pneumonia-induced monocyte/macrophage recruitment. J Immunol. 2001;166:7353–61. https://doi.org/10.4049/jimmunol.166.12.7353.
Sato S, Nieminen J. Seeing strangers or announcing danger: galectin-3 in two models of innate immunity. Glycoconj J. 2002;19:583–91. https://doi.org/10.1023/B:GLYC.0000014089.17121.cc.
Saksida T, Nikolic I, Vujicic M, Nilsson UJ, Leffler H, Lukic ML, et al. Galectin-3 deficiency protects pancreatic islet cells from cytokine-triggered apoptosis in vitro. J Cell Physiol. 2013;228:1568–76. https://doi.org/10.1002/jcp.24318.
Donath MY, Ehses JA, Maedler K, Schumann DM, Ellingsgaard H, Eppler E, et al. Mechanisms of beta-cell death in type 2 diabetes. Diabetes. 2005;54(Suppl 2):S108–13. https://doi.org/10.2337/diabetes.54.suppl_2.s108.
Eizirik DL, Darville MI. beta-cell apoptosis and defense mechanisms: lessons from type 1 diabetes. Diabetes. 2001;50(Suppl 1):S64–9. https://doi.org/10.2337/diabetes.50.2007.s64.
Sparre T, Christensen UB, Gotfredsen CF, Larsen PM, Fey SJ, Hjernø K, et al. Changes in expression of IL-1β influenced proteins in transplanted islets during development of diabetes in diabetes-prone BB rats. Diabetologia. 2004;47:892–908.
Van Gassen N, Staels W, Van Overmeire E, De Groef S, Sojoodi M, Heremans Y, et al. Concise review: macrophages: versatile gatekeepers during pancreatic beta-cell development, injury, and regeneration. Stem Cells Transl Med. 2015;4:555–63. https://doi.org/10.5966/sctm.2014-0272.
Radosavljevic G, Volarevic V, Jovanovic I, Milovanovic M, Pejnovic N, Arsenijevic N, et al. The roles of Galectin-3 in autoimmunity and tumor progression. Immunol Res. 2012;52:100–10. https://doi.org/10.1007/s12026-012-8286-6.
Mensah-Brown EP, Al Rabesi Z, Shahin A, Al Shamsi M, Arsenijevic N, Hsu DK, et al. Targeted disruption of the galectin-3 gene results in decreased susceptibility to multiple low dose streptozotocin-induced diabetes in mice. Clin Immunol. 2009;130:83–8. https://doi.org/10.1016/j.clim.2008.08.024.
Zimmet P, Alberti KG, Shaw J. Global and societal implications of the diabetes epidemic. Nature. 2001;414:782–7. https://doi.org/10.1038/414782a.
Schalkwijk CG, Stehouwer CD. Vascular complications in diabetes mellitus: the role of endothelial dysfunction. Clin Sci (Lond). 2005;109:143–59. https://doi.org/10.1042/CS20050025.
Schalkwijk CG, Miyata T. Early- and advanced non-enzymatic glycation in diabetic vascular complications: the search for therapeutics. Amino Acids. 2012;42:1193–204. https://doi.org/10.1007/s00726-010-0779-9.
Madrigal-Matute J, Lindholt JS, Fernandez-Garcia CE, Benito-Martin A, Burillo E, Zalba G et al. Galectin-3, a biomarker linking oxidative stress and inflammation with the clinical outcomes of patients with atherothrombosis. J Am Heart Assoc. 2014;3. https://doi.org/10.1161/JAHA.114.000785.
Ozturk D, Celik O, Satilmis S, Aslan S, Erturk M, Cakmak HA, et al. Association between serum galectin-3 levels and coronary atherosclerosis and plaque burden/structure in patients with type 2 diabetes mellitus. Coron Artery Dis. 2015;26:396–401. https://doi.org/10.1097/MCA.0000000000000252.
Saeed M, Tapia G, Ariansen I, Stene LC, Seljeflot I, Tell GS, et al. Serum Galectin-3 and subsequent risk of coronary heart disease in subjects with childhood-onset type 1 diabetes: a cohort study. Diabetes Care. 2021. https://doi.org/10.2337/dc20-1712.
de Boer IH, Rue TC, Hall YN, Heagerty PJ, Weiss NS, Himmelfarb J. Temporal trends in the prevalence of diabetic kidney disease in the United States. JAMA. 2011;305:2532–9. https://doi.org/10.1001/jama.2011.861.
Tufro A, Veron D. VEGF and podocytes in diabetic nephropathy. Semin Nephrol. 2012;32:385–93. https://doi.org/10.1016/j.semnephrol.2012.06.010.
Tan KCB, Cheung CL, Lee ACH, Lam JKY, Wong Y, Shiu SWM. Galectin-3 is independently associated with progression of nephropathy in type 2 diabetes mellitus. Diabetologia. 2018;61:1212–9. https://doi.org/10.1007/s00125-018-4552-z.
Hodeib H, Hagras MM, Abdelhai D, Watany MM, Selim A, Tawfik MA, et al. Galectin-3 as a prognostic biomarker for diabetic nephropathy. Diabetes Metab Syndr Obes. 2019;12:325–31. https://doi.org/10.2147/DMSO.S194410.
Canning P, Glenn JV, Hsu DK, Liu FT, Gardiner TA, Stitt AW. Inhibition of advanced glycation and absence of galectin-3 prevent blood-retinal barrier dysfunction during short-term diabetes. Exp Diabetes Res. 2007;2007:51837. https://doi.org/10.1155/2007/51837.
Bauer PM, Zalis MC, Abdshill H, Deierborg T, Johansson F, Englund-Johansson U. Inflamed In Vitro retina: cytotoxic neuroinflammation and Galectin-3 expression. PLoS One. 2016;11: e0161723. https://doi.org/10.1371/journal.pone.0161723.
Abreu CA, De Lima SV, Mendonca HR, Goulart CO, Martinez AM. Absence of galectin-3 promotes neuroprotection in retinal ganglion cells after optic nerve injury. Histol Histopathol. 2017;32:253–62. https://doi.org/10.14670/HH-11-788.
Mendonca HR, Carvalho JNA, Abreu CA, Mariano de Souza Aguiar Dos Santos D, Carvalho JR, Marques SA et al. Lack of Galectin-3 attenuates neuroinflammation and protects the retina and optic nerve of diabetic mice. Brain Res. 2018;1700:126–37. https://doi.org/10.1016/j.brainres.2018.07.018.
Mendonca HR, Carpi-Santos R, da Costa CK, Blanco Martinez AM. Neuroinflammation and oxidative stress act in concert to promote neurodegeneration in the diabetic retina and optic nerve: galectin-3 participation. Neural Regen Res. 2020;15:625–35. https://doi.org/10.4103/1673-5374.266910.
Stitt AW, McGoldrick C, Rice-McCaldin A, McCance DR, Glenn JV, Hsu DK, et al. Impaired retinal angiogenesis in diabetes: role of advanced glycation end products and galectin-3. Diabetes. 2005;54:785–94.
Armstrong DG, Cohen K, Courric S, Bharara M, Marston W. Diabetic foot ulcers and vascular insufficiency: our population has changed, but our methods have not. J Diabetes Sci Technol. 2011;5(6):1591–5.
Driver VR, Fabbi M, Lavery LA, Gibbons G. The costs of diabetic foot: the economic case for the limb salvage team. J Am Podiatr Med Assoc. 2010;100:335–41. https://doi.org/10.7547/1000335.
Gunes AE, Eren MA, Koyuncu I, Taskin A, Sabuncu T. Investigation of galectin-3 levels in diabetic foot ulcers. J Wound Care. 2018;27:843–8. https://doi.org/10.12968/jowc.2018.27.12.843.
Wesley UV, Vemuganti R, Ayvaci ER, Dempsey RJ. Galectin-3 enhances angiogenic and migratory potential of microglial cells via modulation of integrin linked kinase signaling. Brain Res. 2013;1496:1–9. https://doi.org/10.1016/j.brainres.2012.12.008.
Markowska AI, Liu FT, Panjwani N. Galectin-3 is an important mediator of VEGF- and bFGF-mediated angiogenic response. J Exp Med. 2010;207:1981–93. https://doi.org/10.1084/jem.20090121.
Pepe D, Elliott CG, Forbes TL, Hamilton DW. Detection of galectin-3 and localization of advanced glycation end products (AGE) in human chronic skin wounds. Histol Histopathol. 2014;29:251–8. https://doi.org/10.14670/HH-29.251.
Seferovic JP, Lalic NM, Floridi F, Tesic M, Seferovic PM, Giga V, et al. Structural myocardial alterations in diabetes and hypertension: the role of galectin-3. Clin Chem Lab Med. 2014;52:1499–505. https://doi.org/10.1515/cclm-2014-0265.
Bolotskykh G, Rudyk I. TNFa and Serum Galectin-3 in Patients with Heart Failure with Preserved Left Ventricular Ejection Fraction, Insulin Resistance and Type 2 Diabetes Mellitus. J Card Fail. 2014;20:S24-S.
Xu GR, Zhang C, Yang HX, Sun JH, Zhang Y, Yao TT, et al. Modified citrus pectin ameliorates myocardial fibrosis and inflammation via suppressing galectin-3 and TLR4/MyD88/NF-kappaB signaling pathway. Biomed Pharmacother. 2020;126: 110071. https://doi.org/10.1016/j.biopha.2020.110071.
Lepojarvi ES, Piira OP, Paakko E, Lammentausta E, Risteli J, Miettinen JA, et al. Serum PINP, PIIINP, galectin-3, and ST2 as surrogates of myocardial fibrosis and echocardiographic left venticular diastolic filling properties. Front Physiol. 2015;6:200. https://doi.org/10.3389/fphys.2015.00200.
Goncalves N, Silva AF, Rodrigues PG, Correia E, Moura C, Eloy C, et al. Early cardiac changes induced by a hypercaloric Western-type diet in subclinical obesity. Am J Physiol Heart Circ Physiol. 2016;310:H655–66. https://doi.org/10.1152/ajpheart.00684.2015.
Saeed M, Tapia G, Ariansen I, Stene LC, Seljeflot I, Tell GS, et al. Serum Galectin-3 and Subsequent Risk of Coronary Heart Disease in Subjects With Childhood-Onset Type 1 Diabetes: A Cohort Study. Diabetes Care. 2021;44:810–6. https://doi.org/10.2337/dc20-1712.
Iacoviello M, Aspromonte N, Leone M, Paradies V, Antoncecchi V, Valle R, et al. Galectin-3 serum levels are independently associated with microalbuminuria in chronic heart failure outpatients. Res Cardiovasc Med. 2016;5: e28952. https://doi.org/10.5812/cardiovascmed.28952.
Acknowledgements
We thank AJE for language editing service, and we thank Ms Hailan Zou for editing the manuscript.
Funding
This work was supported by National Key Research and Development Project (2018YFE0114500 to Y.X.), the National Natural Science Foundation of China (NSFC) grant (81870577 to Y.X.), the National Science Foundation of Hunan Province for Excellent Young Scholars (2020JJ3056 to Y.X.), and the science and technology innovation Program of Hunan Province (2021RC 3033 to Y.X.)
Author information
Authors and Affiliations
Contributions
Writing—original draft preparation, Yanhua Li; writing‑review and editing, Tian Li and Yang Xiao; Idea: Yang Xiao; graphical design, Zhiguang Zhou. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Consent for publication
All authors read and approve the publication.
Competing interests
None.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Facts
➢ Diabetes has become a top healthcare concern worldwide. Chronic complications of diabetes, especially cardiovascular complications, are the primary causes of death and disability in the ageing population.
➢ There is a growing concern that medical advances in reducing cardiovascular disease will be reversed by the epidemic of obesity and diabetes.
➢ Galectin-3 is widely expressed in inflammatory cells and plays a role in the activation, transport, and cytokine release of inflammatory cells.
Open questions
➢ Is circulating Galectin-3 a risk factor for diabetes and diabetes complications?
➢ What are the specific mechanisms of Galectin-3 in the pathophysiology of diabetes and diabetes complications?
➢ Can we get clinical and theranostic opinion for treating diabetes-related cardiovascular diseases regarding the potential of Galectin-3 in diabetes and its complications?
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, Y., Li, T., Zhou, Z. et al. Emerging roles of Galectin-3 in diabetes and diabetes complications: A snapshot. Rev Endocr Metab Disord 23, 569–577 (2022). https://doi.org/10.1007/s11154-021-09704-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11154-021-09704-7