
Abstract
Background and aims
Soil organic carbon (SOC) content dominates the organic carbon pools in grasslands. Thus, factors that influence and regulate SOC content must be clarified when addressing global climate change.
Methods
We investigated SOC in the top 10 cm of the soil layer from 109 plots of the Mongolian Plateau, including both desert-steppe and steppe landscapes. We then examined the association between SOC with grazing intensity, climatic factors, soil properties, and vegetation diversity indices.
Results
Desert-steppe samples had lower mean SOC than steppe samples (0.3% versus 1.5%). Desert-steppe SOC did not vary with grazing intensity, but steppe SOC did. Instead, desert-steppe SOC decreased and increased, respectively, with higher growing season temperature and soil electrical conductivity; these two variables were the major factors influencing SOC in this grassland type. Besides grazing, outside growing season precipitation and soil pH were the major environmental factors that correlated positively with steppe SOC.
Conclusion
The SOC of steppe grasslands was more sensitive to grazing than SOC of desert-steppe grasslands. Additionally, while climate, grazing, soil, and vegetation all regulated SOC, the most influential variables differed between the two grassland types. These findings enhance our understanding of SOC regulatory mechanisms in different grasslands on the Mongolian Plateau. Such knowledge is crucial for predicting the consequences of environmental change on carbon sequestration.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The soil stores three times the carbon of terrestrial vegetation and twice the amount of atmospheric carbon (Smith 2012). Thus, soil is a crucial part of organic C stock in terrestrial ecosystems (Chu et al. 2018; Wang et al. 2016) and determines regional C storage capacity. Small fluctuations in soil organic carbon (SOC) pools can in fact disturb atmospheric carbon concentrations (Zhou et al. 2019) and influence global warming (Harrison et al. 2021).
Grasslands are the largest type of terrestrial ecosystem on Earth, covering about 40% of total land area (Sun et al. 2022). Plant-soil interactions cause tremendous carbon sequestration in grasslands, on the level of 525–634 Pg C (Yang et al. 2019c). Hence, research on global terrestrial carbon cycle and carbon budget management necessitates investigating the factors that influence SOC content in grasslands (Dong et al. 2022; Wu et al. 2022; Yu et al. 2023).
Plant community composition, and their variation across grassland types, exerts an effect on SOC that is critical for ecosystem function and soil biogeochemical cycling. Concentrations and storage of SOC and soil total nitrogen decrease along a decreasing elevation gradient of alpine meadows > mountain meadows > temperate typical steppes > temperate steppe deserts > temperate desert steppes (Bi et al. 2018). Additionally, vegetation composition is a determinant of grazing effects on soil C sequestration and thus SOC content in rangelands (Bagchi and Ritchie 2010; Reeder et al. 2004).
Livestock grazing is a common strategy to manage grasslands and exerts notable effects on SOC (Abdalla et al. 2018; Bork et al. 2023; Dlamini et al. 2016). Grazing can stimulate shallow and fine roots in C4 species, leading to an SOC increase (Derner et al. 2006). However, inappropriate grazing of grassland ecosystems can result in SOC depletion (Wang et al. 2011), whereas grazing exclusion measurements can enhance SOC sequestration (Sun et al. 2022). Globally, livestock grazing establishments have decreased SOC stocks by an average of 10.3% (Zhou et al. 2017). Besides, adverse effects of grazing on SOC vary considerably across climatic regions (arid, semi-arid, and humid), in response to precipitation (Phukubye et al. 2022). Additionally, grassland type affects SOC sensitivity to grazing. In desert steppes, grazing disturbance did not appear to influence SOC (Liu et al. 2012; Zhang et al. 2018). In steppes, the influence of livestock grazing is more variable. Overall, free grazing depletes SOC content the most (Wang et al. 2017), and as grazing intensity increases, SOC appears to increases before declining (Baohong et al. 2018; Han et al. 2008). When comparing steppes and other grasslands, grazing significantly reduced SOC content in the Stipa steppes and scattered tree grassland, but had no significant effect on meadow or sandy grasslands (Zhang et al. 2017a). Alpine meadows were least sensitive to changes in grazing intensity, with SOC content largely remaining unaltered. Lowland meadows and temperate desert grasslands may be the most susceptible to grazing intensity because their primary production of fixed carbon is plant litter. Livestock grazing considerly decreases the amount of plant litter and thus lowers carbon input to the soil from that source (Derner et al. 2006; Wang et al. 2022a).
Beyond grassland management practices, environmental factors can substantially influence carbon balance in grasslands (Wang et al. 2021). In particular, temperature exerts a substantial influence on the physiological, chemical, and biochemical processes that affect SOC input/output. Notably, litter disintegration slows under low temperatures (Zhang et al. 2017b). Moreover, a temperature increase stimulates plant productivity in grasslands through enhancing photosynthesis and extending the growing season (Piao et al. 2020). Increased temperatures can also bolster microbial activity, speeding up decomposition of soil organic matter and carbon loss (Dutta and Dutta 2016). Hence, in both desert steppes and steppes, SOC is negatively correlated with temperature (Bi et al. 2018; Ding et al. 2022; Xu et al. 2018).
Aridity is another environmental factor that alters SOC, with the amount of precipitation mediating both above- and below-ground C inputs through influencing soil water availability, vegetation biomass, and biological weathering (Wang et al. 2010; Nielsen and Ball 2015). In desert ecosystems, elevated aridity decreases SOC content (Zhang et al. 2019a). Similarly, in some steppe grasslands, SOC stocks and their light fractions are negatively correlated with aridity (Hu et al. 2022); that is, positively correlated with rainfall (Bi et al. 2018). In desert steppes, SOC concentration and stock are also positively correlated with mean annual rainfall (Yang et al. 2023). However, the relationship is not always straightforward, as SOC density and mean annual precipitation are negatively correlated in the eastern transect region of Eurasian steppes (Zhao et al. 2017).
This variation in effects from aridity and precipitation may be related to environmental conditions outside of the growing season; such forms of precipitation tend to be snowfall (Yang et al. 2019a), a major contributor to moisture levels and nutrient cycling in desert regions particularly (Ball et al. 2011; Hu et al. 2019; Zhao et al. 2016). Snow depth is positively correlated with net ecosystem exchange (NEE) of CO2, thus lowering its intra- and inter-annual variation. Deep snow significantly elevates root biomass in temperate grasslands (Li et al. 2020), while also increasing the capacity of subsoil to incorporate C leached from topsoil in both desert steppe and steppe grasslands. Indeed, research on the effects of snowfall suggest that vertical C transport plays a major role in turning the subsoil into a C sink, overturning prior ideas that this soil layer is climate-insensitive (Deng et al. 2023). Considering such effects, more consideration should be given to climatic conditions outside the growing season when researching SOC. Currently, most studies have investigated environmental conditions during the growing season only.
Soil physiochemical properties are also another crucial factor that modulates the decomposition rate of organic matter via influencing soil microbial abundance (Kögel-Knabner et al. 2008). Soil pH and electrical conductivity (EC), for example, significantly influence SOC content through their effects on plant growth and soil microbial activity (Xu et al. 2021; Zhu and Shao 2018).
Any analysis of SOC flux and accumulation must account for interactions between the many complex factors involved, including grazing intensity, environmental conditions, plant communities, and soil characteristics (Cao and Woodward 1998; Jobbágy and Jackson 2000; Piao et al. 2005; Wiesmeier et al. 2012). Structural equation modeling (SEM) is thus an excellent method for investigating SOC dynamics. This methodology is used to investigate causal network relationships among variables within a system, offering a systematic framework for comprehensively exploring changes in soil C and uncovering underlying mechanisms (Luo et al. 2017). Thus far, research examining SOC dynamics have largely focused on a single vegetation type or solely on climatic factors, with limited attention on combining effects from in-season/out-of-season conditions, grazing, and their interactions. Furthermore, grassland types should be accounted for in any explanatory model, given that dominant SOC-influencing factors differ between them (Liu et al. 2023; Lu et al. 2023; Yu et al. 2023).
The Mongolian Plateau of Asia, comprising mainly of desert steppe and steppe grasslands, is an integral component of worldwide rangeland, providing major benefits to global livestock development and ecosystem services (Wu et al. 2015). Understanding how environmental variables and grazing intensity affect SOC on a regional level such as the Plateau will be useful for predicting global SOC dynamics. In this study, we used data from 109 field plots of the Mongolian Plateau to test two hypotheses. First, SOC of steppe grasslands is more sensitive to grazing than SOC of desert-steppe grasslands. Second, climate, grazing, soil properties, and vegetation community all combine to regulate SOC, but their relative contributions differ between desert-steppes and steppes. To test our two hypotheses, we specifically aimed to answer three questions: (1) Which are the main factors (grazing, climate, plant composition) controlling SOC between steppes and desert-steppes? (2) How does grazing intensity interact with climate and other environmental variables to influence SOC concentrations in the Plateau’s grassland ecosystems? (3) What is the mechanism driving SOC dynamics across the two grassland types?
Materials and methods
Study sites
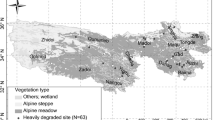
Survey data were collected from seven sites distributed across the central and southern regions of the Mongolian Plateau, ranging from 43.80–47.04°N to 100.29 –109.94°E (Fig. 1; Table 1). Study-site elevation spans 935–2127 m. The study area is characterized by a seasonal, dry, continental climate with a mean annual temperature of -2.1–5.2℃ and mean annual precipitation of 190–487 mm. The growing season is May through September and the out-of-growing season is October through April (Li et al. 2022; Wang et al. 2014). Dominant natural vegetation types are Stipa gobica, Stipa glareosa, and Allium polyrrhizum in desert-steppes, as well as Stipa krylovii and Cleistogenes soongorica in steppes.

Locations of the study sample sites on the Mongolian Plateau
Data collection
Grazing intensity treatment
Grazing is the primary land use in the Mongolian Plateau, a region with a nomadic history spanning over 3,000 years (Zhang et al. 2019a, b). Settlements and water sources usually considered the areas of highest grazing intensity. Thus, for this study, plots for measuring grazing intensity were selected to decrease with increasing distance from settlements. Transect sampling along the grazing gradient was chosen with four or five replicates per site. The farthest plots were positioned equidistant from water sources or settlements. Regions surrounding each settlement was stratified into five concentric levels spaced at 5 km intervals. These levels were denoted as follows: overgrazing, 0 km away from a settlement; high grazing, 5 km away; medium grazing, 10 km away; and light grazing, 15 km away. The main study area, located 20 km from all settlements, is a no-grazing site. Sheep density data for 2007 were extracted for each site (Table S1) from the National Statistics Office of Mongolia (https://www.1212.mn).
Field sampling design
Field sampling was conducted from late July to mid-August 2008, when grassland biomass peaks. For sampling, 109 field plots were established on topographically level areas (Fig. 1), comprising 48 desert-steppe plots and 61 steppe plots. Sampling site location was determined using GPS. In each plot, 10 m × 10 m subsections were selected under different grazing gradients to survey vegetation and soil.
Vegetation surveys
Vegetation was surveyed at seven sites using published phytosociological methods (Braun-Blanquet 1964). Species composition per plot was documented using Braun-Blanquet’s cover-abundance scale (Braun-Blanquet 1964; Ellenberg and Mueller-Dombois 1974). Plant height and coverage per plot were also measured.
Three indices were used for community biodiversity: species richness (SR) (Brown et al. 2007), the Shannon–Wiener index (H) (Shannon and Weaver 1949), and the Pielou evenness index (E) (Pielou 1966). Species richness of grasses, forbs, and shrubs (SRgrass, SRforb, and SRshrub) was calculated as the average number of species per plot (number of species/16 mp2). The S/G ratio refers to species richness of shrubs divided by species richness of grasses. The following equations were used to calculate H (Eq. 2) and E (Eq. 3):
where N is number of individuals in the community, Ni is number of individuals in species i, S is number of species in the community, and Pi is proportion of individual members in species i to total number of individuals in the community.
Soil sampling and measurements
Samples per plot were collected from 0–10 cm soil depths. We collected three soil samples from each grassland plot using excavation and thoroughly mixed them to ensure uniformity. These soil samples were air-dried and subsequently sieved through a 2 mm sieve to remove visible plant roots and rock fragments. Soil EC was measured in a 1:5 (soil: water) suspension using an EC meter. Soil pH was measured with a pH meter in a 1:2.5 (soil:water) suspension (Mettler Toledo, Shanghai, China). Soil hardness was measured by SOIL HARDNESS TESTER (YAMANAKA, STANDARD TYPE). Soil moisture was measured by TDR Soil Moisture Detector. Soil SOC was quantified using the potassium dichromate oxidation titration method (Walkley and Black 1934). Total nitrogen (TN) was determined using Kjeldahl analysis (Nelson and Sommers 1975). To measure total phosphorus (TP), soil samples were extracted with HClO4-H2SO4 and analyzed using Mo-Sb colorimetry (Olsen et al. 1983).
Climatic data
Five years (2008–2012) of meteorological data in Mongolia were all obtained from the European Center for Medium-Range Weather Forecasts Reanalysis v5 (https://cds.climate.copernicus.eu) (Hersbach et al. 2020). ERA5 variables are provided on a fine-resolution grid, that is, 0.25° × 0. 25°.The data included 2 m temperature (MAT, °C), total precipitation (MAP, mm) and evaporation (E0, mm). Growing season temperature (GST) and growing season precipitation (GSP) were calculated using data from May to September. Outside growing season temperature (OGST) and outside growing season precipitation (OGSP) were calculated using data from October to April. The aridity index (R)—both total and within the growing season (GSR)—was calculated using Eq. 4:
where E0 is annual evaporation (mm) and P is annual precipitation (mm). Higher R indicates that a more severe drought (evaporative capacity exceeds precipitation).
Data analysis
Random forest modeling with 500 trees was performed with the “randomForest” and “rfPermute” packages of R. This model can identify the relative contributions of climate variables (MAT, GST, OGST, MAP, GSP, OGSP, R, and GSR; see Sect. 2.2.5) to SOC. A two-way ANOVA was used to examine whether SOC differs significantly across levels of grazing intensity, GST (in desert-steppes), OGSP (in steppes), and their interactions. Relationships of SOC with climate, soil properties, and plant species composition were tested using Pearson’s correlations (Wang et al. 2022b).
A stepwise regression was used to verify the main drivers of SOC variation (Huang et al. 2022). Tested variables include GST, OGSP, soil EC, soil PH, soil moisture, vegetation index, and grazing intensity. In this step, one indicator was selected for each climate, soil property, and vegetation type to explain changes in SOC.
Based on current literature (Fig. S1), SEM was conducted using the “lavaan” package in R (Rosseel 2012) to examine direct and indirect effects of environment (climate and soil) and vegetation on SOC. This model included SOC as the dependent variable, while endogenous/independent variables were soil physical properties (pH, EC, moisture, hardness), climate (temperature, precipitation, aridity), plant composition (diversity, species richness, evenness), and grazing intensity. The results of SEM are interpreted using the following outputs. Standardized effect estimates represent the effect size of the relationship between two variables; a higher absolute value indicates a stronger relationship. R2 reflects proportion of variance in the dependent variable that each endogenous/independent variable explains (Grace 2006). Overall model interpretation typically involves three fit indices: ratio of maximum likelihood χ2 values to degrees of freedom (df), comparative fit index (CFI), and root mean square error of approximation (RMSEA). Model fit is considered acceptable when χ2/df < 3, RMSEA < 0.10 and CFI ≥ 0.90 (W. Zhao et al. 2019). Finally, path coefficients of endogenous variables indicate the strength and direction of their relationships with other variables (Kline 2011).
All statistical analyses were conducted in SPSS (IBM Corp 2019) and R-4.3.3 (R Core Team 2024).
Results
Grazing effect on the SOC changes of desert-steppe and steppe grasslands
The two grassland types varied in their SOC response to grazing intensity. Specifically, desert-steppe SOC (~ 0.5%) did not vary with grazing intensity (Fig. 2a, Table 2). However, steppe SOC altered significantly (Fig. 2b, Table 2), being highest (1.48%) under high grazing intensity and lowest (0.97%) under middle grazing intensity.

The effect of grazing intensity on SOC in the desert-steppe grassland (DS) (a) and the steppe grassland (S) (b) under different climate conditions. The value is mean ± SD. Different capital letters indicate significant differences between the grazing intensity of different grasslands at the same climate condition at the 0.05 level, and different lowercase letters indicate significant differences between the climate conditions at the same grazing intensity at the 0.05 level. NG, no grazing; LG, light grazing intensity; MG, middle grazing intensity; HG, high grazing intensity; OG, overgrazing intensity. GST, growing season temperature; OGSP: outside growing season precipitation
Random forest modeling revealed that GST in desert-steppes and OGSP in steppes were the main climatic factors affecting SOC (Fig. 3). Desert-steppes and steppes also differed when other variables were considered in conjunction with grazing intensity. Steppe SOC responded to the interaction between grazing intensity and OGSP, whereas desert-steppe SOC did not respond to the interactions of any variables.

Climate Variable importance derived from the random forest model for soil organic carbon. GST: growing season temperature; GSR: growing season aridity index; MAT: mean annual temperature; MAP: mean annual precipitation; OGSP: outside-growing season precipitation; OGST: outside-growing season temperature; R: the aridity index; GSP: growing season precipitation. *, **,*** represent significance at the P < 0.05, P < 0.01, and P < 0.001 levels, respectively
Factors influencing desert-steppe and steppe SOC variation
Desert-steppe SOC was negatively correlated with GST, MAP, GSR and TP, but positively correlated with EC, TN, and moisture (Fig. S2a). The results demonstrated that there was no significant correlation between vegetation factors and grazing intensity with respect to SOC (Fig. S2a). In the steppe grassland, precipitation (MAP, GSP, and OGSP) was positively correlated with steppe SOC, while temperature factors (MAT, GST, and OGST) and the drought index (GSR) were negatively correlated with SOC (Fig. S2b). Soil factors exhibited a positive correlation with SOC. In terms of vegetation, SOC was negatively correlated with species richness of shrubs (SRshrub) and the species richness ratio of shrubs to grasses (S/G) (Fig. S2b) (Li et al. 2021b).
Additionally, stepwise regression explained 32.4% and 83.8% of SOC variation in desert-steppe and steppe grasslands, respectively (Table 3). GI had a critical influence on SOC for the steppe grassland. The two grassland types were regulated by different climatic factors, among which the desert-steppe was affected by GST, and the steppe was influenced by OGSP. In terms of soil variables, stepwise regression (Table 3) found that EC played an important role in desert-steppe SOC, along with moisture and SRf. Steppe SOC was affected by pH in addition to moisture and SRf.
Mechanisms driving variation in desert-steppe and steppe SOC
Goodness-of-fit metrics for our SEM models were satisfactory (desert-steppe SOC: χ2/df = 0.698, CFI > 0.99, and RMSEA < 0.05; steppe SOC: χ2/df = 0.083, CFI > 0.99, and RMSEA < 0.05) (Fig. 4a and b). The SEM model for desert-steppe SOC explained 42% (R2 = 0.42) of the variation. Of that 42%, EC, moisture, and SRf contributed 25%, 43%, and 14%, respectively (Fig. 4a). Climate variable GST had a direct negative impact on SOC (-0.33), while also mediating moisture variables and SRf to exert indirect effects. Both EC and SRf had direct positive effects on SOC (0.48, 0.27), whereas moisture played an indirect role (Fig. 4a). Overall, EC had the greatest positive effect on desert-steppe SOC, with a total effect of 0.483 (Fig. 4c, Table S2).

Structural equation model results for environmental, soil, and vegetation factors influencing soil organic carbon (SOC) in the desert-steppe grassland (a) and steppe grassland (b). And total, direct, and indirect effects of influencing factors on soil carbon were analyzed by structural equation modeling in the desert-steppe grassland (c) and steppe grassland (d). The blue solid arrows and red solid arrows indicate significant positive and negative correlations, respectively, while grey dotted arrows indicate non-significant relationships. Path arrow thickness reflects the strength of the relationship (thicker = stronger relationship). The numbers next to the path arrows indicate the standardized effect estimates and the direction of their relationship (+ or −). “*”, “**”, “***” indicate significant differences at P < 0.05, P < 0.01, and P < 0.001.The proportion of variance explained (R2) appears alongside each response variable in the model. Goodness-of-ft statistics for the model are shown below the SEM model. GI denotes grazing intensity; GST denotes growing season temperature; OGSP denotes outside growing season precipitation; EC denotes electric conductivity; PH denotes potential of hydrogen; Moisture denotes soil moisture; SRf denotes species richness of forb
The SEM model for steppe SOC explained 85% (R2 = 0.85) of the variation, with pH (18%), moisture (86%), and SRf (31%) being the greatest contributors (Fig. 4b). Grazing intensity and OGSP regulated moisture and forb species richness, thereby indirectly affecting SOC content (Fig. 4b). Soil pH was another indirect regulator of SOC through its influence on plants, while itself being regulated by moisture (Fig. 4b). Overall, the most important variable affecting steppe SOC was OGSP, with a total effect of 0.899 (Fig. 4d, Table S2).
Discussion
Sensitivity of soil organic carbon content to grazing in steppe and desert—steppe grasslands
The results of our study demonstrate that grazing differentially affected SOC depending on the grassland type. Overall, steppe SOC was more sensitive to grazing than desert-steppe SOC (Fig. S3), validating our first hypothesis. Steppe plots with low grazing exhibit higher SOC levels than other grazing conditions (Table 2, Fig. 2b), aligning with previous findings (Liu et al. 2012; Schuman et al. 1999). Instead of grazing, desert-steppe SOC was primarily modulated by climate, soil, and vegetation communities (Fig. 4a). The difference between these two grasslands is likely related to aridity. Drier environments (i.e., desert-steppes) are more affected than wetter environments by unpredictable, abiotic factorsn (Wiens 1984). As such, desert-steppe grasslands behave like nonequilibrium ecosystem models, where vegetation dynamics are largely driven by external drivers such as climatic variability (Derry and Boone 2010; Sullivan and Rohde 2002). In contrast, steppe grasslands follow equilibrium models, where ecosystems are mostly regulated by internal biotic factors (e.g., interspecific interactions such as herbivory) (Briske 2016; Wu and Loucks 1995).
Interestingly, a complete absence of grazing did not increase SOC over low grazing in steppe grasslands. This outcome may be because a small amount of grazing causes more litter accumulation, increasing carbon input from both aboveground and belowground sources. Animal traffic also aids in litter breakdown, enhancing decomposition and carbon transfer to the soil.
Another factor that likely played a role in the effect of grazing on steppe grasslands is the mix of C3 versus C4 plants. Previous studies have observed that in C4-dominant or mixed C3/C4 grasslands, SOC increases as grazing intensity increases; however, the correlation becomes negative in C3-dominant grasslands (McSherry and Ritchie 2013; Schönbach et al. 2011). We observed a similar pattern in our study. High grazing of steppe grasslands also increased SOC (Fig. 2b), apparently through stimulating rapid turnover in plant species composition that favored more C4 and mixed C3-C4 plants (Table S3). Previous research has uncovered the same phenomenon under heavy grazing (Reeder and Schuman 2002). Through its effects on plant community turnover, high grazing intensity may also alter the soil microbiome and influence SOC in that direction (Eldridge et al. 2017).
Nevertheless, grazing intensity overall had a weak effect on grasslands in our study (Fig. 4). In desert-steppe grasslands, the effect of environmental factors, particularly climate, appears to have overwritten the influence of livestock grazing entirely (Tables 2 and 3). Our findings thus support the need to holistically investigate interactions between grazing, climate, soil, and plant/microbial biodiversity when evaluating ecosystem services such as SOC storage (Maestre et al. 2022). The interplay between grazing intensity and environmental factors, plus the relative sensitivity of ecosystems to these influences, accounts for global variation in soil carbon dynamics (Ren et al. 2024).
Regulatory mechanisms of SOC content in steppe and desert-steppe grasslands
Our data supported our second hypothesis, namely that SOC is regulated by a combination of climate, grazing, soil properties, and vegetation. In desert-steppe grasslands, SOC was influenced by GST, EC, and SRf (Fig. 4a), while in steppe grasslands, SOC was influenced by grazing, OGSP, soil pH, and SRf (Fig. 4b). The relatively low R2 for SOC in desert-steppes (Table 3) suggests that our analysis did not account for critical variables, such as aboveground biomass (Wang et al. 2023) and soil microbial communities (Fan et al. 2021).
Higher temperatures and increased aridity caused SOC to decline, particularly during the growing season (Fig. S2). Increasing temperatures during the growing season increases evapotranspiration and can eventually alter plant type through negative effects on photosynthetic rate and productivity (Fig. S2) (Yang et al. 2019a). Drought also tends to lower plant diversity and biomass, causing corresponding decreases in SOC stock (Yang et al. 2019a). Furthermore, the influence of high temperatures and drought on soil microbial communities will alter enzyme decomposition kinetics (Conant et al. 2011). The resultant changes to above- and below-ground community composition (Fig. S2) (Berdugo et al. 2020) can lead to loss of C stock (Zhang et al. 2015).
Steppe and desert-steppe SOC differed completely in response to precipitation (Fig. S2). Precipitation negatively affected SOC in desert-steppes but positively affected SOC in steppes. This discrepancy is related to the complex interaction of regional humidity, plant communities, and soil microbiota. Elevated precipitation can promote plant growth and increase carbon input into the soil (Qin et al. 2023), but can also enhance soil microbial activity, accelerating decomposition and SOC loss (Hu et al. 2019; Liu et al. 2009). Responses to precipitation is associated with variation in vegetation characteristics and plant biomass allocation across regions (Li et al. 2021a; Ma et al. 2010; Meng et al. 2017). For example, plant species in relatively arid regions may have high drought tolerance. As a result, lower precipitation largely impedes microbial decomposition of SOC rather than hampering plant productivity and decreasing carbon inputs to the soil (Meng et al. 2017). In contrast, areas with normally high precipitation are unlikely to have drought-tolerant plants. Therefore, a drop in precipitation will impede plant growth and cause a decline in soil organic matter (Li et al. 2021a, 2021b; Stocker et al. 2019).
We found that OGSP (snowfall) was positively correlated with SOC (Fig. S2) (Hui et al. 2022; Li et al. 2020) and propose three mechanisms underlying this link. First, snow alters soil moisture and temperature (Fig. S2), thus influencing soil microbial activity and nutrient cycling dynamics (Yang et al. 2010). These changes then affect vegetation growth conditions and community composition (Fig. S2) (Wipf and Rixen 2010). Second, snowfall itself is a temporary reservoir for water and nutrients that can then be exploited by plants at the beginning of the growing season (Clark et al. 2011; Wipf and Rixen 2010). Third, snow slows surface soil freezing and has a protective effect on plant roots (Tierney et al. 2001), soil structure (Kværnø and Øygarden 2006), and microbial communities (Grogan et al. 2004).
Soil EC was the biggest factor influencing desert-steppe SOC (Fig. 4 and Table S2), consistent with prior results (Dong et al. 2022). Two factors could potentially explain a positive correlation between EC and SOC. First, an increase in EC elevates the osmotic pressure of the soil solution, disrupting microbial cells and inhibiting their metabolic processes, including SOC decomposition rates (Yang et al. 2019b). Second, arid ecosystems characterized by high EC often exhibit substantial reserves of shallow groundwater (Fan et al. 2012). These reserves facilitate the growth of salt-tolerant plant species and the decomposition of aboveground plant litter, promoting SOC accumulation (Volik et al. 2021).
In steppe grasslands, although soil EC significantly affected SOC, the influence of soil pH was considerably greater (Table 3; Fig. 4b). Soil pH directly contributes to the decomposition of soil organic matter (Dlamini et al. 2016). Furthermore, species richness of vascular plant communities often exhibits a positive correlation with soil pH (Pärtel 2002). Here, we observed that species richness decreased on both ends of the pH spectrum. This relationship may be because many plants are not well-adapted to extremely acidic or alkaline soils, resulting in lower species richness (Currie et al. 2004; Palpurina et al. 2017).
Forb species richness (SRf) positively influenced SOC in both grasslands, consistent with the plant diversity–soil C storage hypothesis. This hypothesis states that high plant diversity generally leads to increased net primary productivity and, consequently, higher SOC storage (Chen et al. 2018). In line with ours, previous studies have also found that high plant diversity seems to increase carbon output and enhance SOC accumulation (Liu et al. 2018; Shi et al. 2016).
The implications and limitations of the study
This study contributes to the understanding of SOC dynamics in arid and semi-arid grassland ecosystems. We advocate differentiated management strategies tailored to enhance carbon sequestration in specific grassland types. For steppe grasslands, attention should be paid to the simultaneous impact of climate change and grazing. In contrast, for desert-steppe grasslands, more emphasis should be placed on addressing the impact of climate change. Management efforts should also account for precipitation seasonality when considering fluctuations in SOC content.
However, this study has several limitations. First, the geographical scope and one-year duration limits our ability to capture the full spatiotemporal variability of SOC processes. Second, we collected no data on soil microbial activity. Considering the importance of microbial communities in SOC decomposition and sequestration, we are likely missing several key elements to the underlying ecological mechanisms. Third, the absence of soil bulk density measurements impeded the calculation of SOC stocks. Consequently, we could only provide insights into SOC concentration, not actual stock levels. Obtaining accurate measures of SOC stock are essential for understanding the implications of environmental change for the carbon cycle. Future research should aim to address these limitations, thereby providing a more robust and comprehensive view of grassland SOC dynamics.
Conclusion
In this study, we examined the effects of grazing, climate, soil properties, and vegetation communities on desert-steppe and steppe SOC in the Mongolian Plateau. We observed that grazing generally had a weak effect on the tested grasslands, particularly in desert-steppes. This outcome was likely due to the stronger influence of other environmental variables, particularly climate (growing-season temperature for desert-steppe SOC and out-of-growing-season precipitation for steppe SOC). Soil variables such as EC and pH were also crucial through direct and indirect contributions to SOC. Overall, our study offers a new perspective for understanding how grassland type mediates the response of SOC to grazing and climate. The findings will benefit pasture management in arid and semi-arid regions under climate change.
Data availability
The datasets are available from the corresponding author upon reasonable request.
References
Abdalla M, Hastings A, Chadwick DR, Jones DL, Evans CD, Jones MB, Rees RM, Smith P (2018) Critical review of the impacts of grazing intensity on soil organic carbon storage and other soil quality indicators in extensively managed grasslands. Agric Ecosyst Environ 253:62–81. https://doi.org/10.1016/j.agee.2017.10.023
Bagchi S, Ritchie ME (2010) Introduced grazers can restrict potential soil carbon sequestration through impacts on plant community composition. Ecol Lett 13(8):959–968. https://doi.org/10.1111/j.1461-0248.2010.01486.x
Ball BA, Barrett JE, Gooseff MN, Virginia RA, Wall DH (2011) Implications of meltwater pulse events for soil biology and biogeochemical cycling in a polar desert. Polar Res 30:14555. https://doi.org/10.3402/polar.v30i0.14555
Baohong D, Cuiping G, Hadachaolu (2018) Effect of grazing on plant biomass and carbon storage of typical grassland in Xilinguole. Research of soil and water conservation. Res Soil Water Conserv 25(1):139–146, 152. https://doi.org/10.13869/j.cnki.rswc.2018.01.023
Berdugo M, Delgado-Baquerizo M, Soliveres S, Hernández-Clemente R, Zhao Y, Gaitán JJ, Gross N, Saiz H, Maire V, Lehmann A, Rillig MC, Solé RV, Maestre FT (2020) Global ecosystem thresholds driven by aridity. Science 367(6479):787–790. https://doi.org/10.1126/science.aay5958
Bi X, Li B, Nan B, Fan Y, Fu Q, Zhang X (2018) Characteristics of soil organic carbon and total nitrogen under various grassland types along a transect in a mountain-basin system in Xinjiang, China. J Arid Land 10(4):612–627. https://doi.org/10.1007/s40333-018-0006-1
Bork EW, Hewins DB, Lamb EG, Carlyle CN, Lyseng MP, Chang SX, Alexander MJ, Willms WD, Iravani M (2023) Light to moderate long-term grazing enhances ecosystem carbon across a broad climatic gradient in northern temperate grasslands. Sci Total Environ 894:164978. https://doi.org/10.1016/j.scitotenv.2023.164978
Braun-Blanquet J (1964) Pflanzensoziologie: Grundzüge der vegetationskunde. Springer-Verlag, Berlin
Briske (2016) Rangeland systems: processes, management and challenges. Springer, Cham
Brown RL, Reilly LAJ, Peet RK (2007) Species richness: small scale. Encyclopedia of life sciences, 1st ed. Wiley. https://doi.org/10.1002/9780470015902.a0020488
Cao M, Woodward FI (1998) Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their responses to climate change. Glob Change Biol 4(2):185–198. https://doi.org/10.1046/j.1365-2486.1998.00125.x
Chen S, Wang W, Xu W, Wang Y, Wan H, Chen D, Tang Z, Tang X, Zhou G, Xie Z, Zhou D, Shangguan Z, Huang J, He J-S, Wang Y, Sheng J, Tang L, Li X, Dong M, … Bai Y (2018) Plant diversity enhances productivity and soil carbon storage. Proc Natl Acad Sci 115(16):4027–4032. https://doi.org/10.1073/pnas.1700298114
Chu S, Jacobs DF, Sloan JL, Xue L, Wu D, Zeng S (2018) Changes in soil properties under eucalyptus relative to Pinus massoniana and natural broadleaved forests in South China. J For Res 29(5):1299–1306. https://doi.org/10.1007/s11676-017-0546-9
Clark MP, Hendrikx J, Slater AG, Kavetski D, Anderson B, Cullen NJ, Kerr T, Örn Hreinsson E, Woods RA (2011). Representing spatial variability of snow water equivalent in hydrologic and land-surface models: a review. Water Resour Res 47(7). https://doi.org/10.1029/2011WR010745
Conant RT, Ryan MG, Ågren GI, Birge HE, Davidson EA, Eliasson PE, Evans SE, Frey SD, Giardina CP, Hopkins FM, Hyvönen R, Kirschbaum MUF, Lavallee JM, Leifeld J, Parton WJ, Megan Steinweg J, Wallenstein MD, Martin Wetterstedt JÅ, Bradford MA (2011) Temperature and soil organic matter decomposition rates – synthesis of current knowledge and a way forward. Glob Change Biol 17(11):3392–3404. https://doi.org/10.1111/j.1365-2486.2011.02496.x
Currie DJ, Mittelbach GG, Cornell HV, Field R, Guégan J-F, Hawkins BA, Kaufman DM, Kerr JT, Oberdorff T, O’Brien E, Turner JRG (2004) Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol Lett 7(12):1121–1134. https://doi.org/10.1111/j.1461-0248.2004.00671.x
Deng M, Li P, Liu W, Chang P, Yang L, Wang Z, Wang J, Liu L (2023) Deepened snow cover increases grassland soil carbon stocks by incorporating carbon inputs into deep soil layers. Glob Change Biol 29(16):4686–4696. https://doi.org/10.1111/gcb.16798
Derner JD, Boutton TW, Briske DD (2006) Grazing and ecosystem carbon storage in the North American Great Plains. Plant and Soil 280(1–2):77–90. https://doi.org/10.1007/s11104-005-2554-3
Derry JF, Boone RB (2010) Grazing systems are a result of equilibrium and non-equilibrium dynamics. J Arid Environ 74(2):307–309. https://doi.org/10.1016/j.jaridenv.2009.07.010
Ding J, Wang W, Mi W, Hou K, Zhang X, Zhao Y, Wen Q (2022) Spatial characteristics of soil organic carbon in grassland of Ningxia and its influencing factors. Acta Ecol Sin 43(5):1913–1922. https://doi.org/10.5846/stxb202201040022
Dlamini P, Chivenge P, Chaplot V (2016) Overgrazing decreases soil organic carbon stocks the most under dry climates and low soil pH: a meta-analysis shows. Agric Ecosyst Environ 221:258–269. https://doi.org/10.1016/j.agee.2016.01.026
Dong L, Sun Y, Ran J, Hu W, Ji M, Du Q, Xiong J, Gong H, Yao S, Adnan Akram M, Zhang Y, Hou Q, Li H, Sun Y, Lu J, Wang X, Aqeel M, Zhu J, Schmidt MWI, … Deng J (2022) Ecosystem organic carbon storage and their drivers across the drylands of China. Catena 214:106280. https://doi.org/10.1016/j.catena.2022.106280
Dutta H, Dutta A (2016) The microbial aspect of climate change. Energy Ecol Environ 1(4):209–232. https://doi.org/10.1007/s40974-016-0034-7
Eldridge DJ, Delgado-Baquerizo M, Travers SK, Val J, Oliver I (2017) Do grazing intensity and herbivore type affect soil health? Insights from a semi-arid productivity gradient. J Appl Ecol 54(3):976–985. https://doi.org/10.1111/1365-2664.12834
Ellenberg D, Mueller-Dombois D (1974) Aims and methods of vegetation ecology. Wiley
Fan J, Jin H, Zhang C, Zheng J, Zhang J, Han G (2021) Grazing intensity induced alternations of soil microbial community composition in aggregates drive soil organic carbon turnover in a desert steppe. Agric Ecosyst Environ 313:107387. https://doi.org/10.1016/j.agee.2021.107387
Fan X, Pedroli B, Liu G, Liu Q, Liu H, Shu L (2012) Soil salinity development in the yellow river delta in relation to groundwater dynamics. Land Degrad Dev 23(2):175–189. https://doi.org/10.1002/ldr.1071
Grace JB (2006) Structural equation modeling and natural systems. Cambridge University Press, Cambridge
Grogan P, Michelsen A, Ambus P, Jonasson S (2004) Freeze–thaw regime effects on carbon and nitrogen dynamics in sub-arctic heath tundra mesocosms. Soil Biol Biochem 36(4):641–654. https://doi.org/10.1016/j.soilbio.2003.12.007
Han G, Hao X, Zhao M, Wang M, Ellert BH, Willms W, Wang M (2008) Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agric Ecosyst Environ 125(1):21–32. https://doi.org/10.1016/j.agee.2007.11.009
Harrison MT, Cullen BR, Mayberry DE, Cowie AL, Bilotto F, Badgery WB, Liu K, Davison T, Christie KM, Muleke A, Eckard RJ (2021) Carbon myopia: the urgent need for integrated social, economic and environmental action in the livestock sector. Glob Change Biol 27(22):5726–5761. https://doi.org/10.1111/gcb.15816
Hersbach H, Bell B, Berrisford P, Hirahara S, Horányi A, Muñoz-Sabater J, Nicolas J, Peubey C, Radu R, Schepers D, Simmons A, Soci C, Abdalla S, Abellan X, Balsamo G, Bechtold P, Biavati G, Bidlot J, Bonavita M, … Thépaut J-N (2020) The ERA5 global reanalysis. Q J R Meteorol Soc 146(730):1999–2049. https://doi.org/10.1002/qj.3803
Hu Z, Song X, Wang M, Ma J, Zhang Y, Xu H-J, Hu Z, Zhu X, Liu H, Ma J, Yu Q, Ostle NJ, Li Y, Yue C (2022) Aridity influences root versus shoot contributions to steppe grassland soil carbon stock and its stability. Geoderma 413:115744. https://doi.org/10.1016/j.geoderma.2022.115744
Hu Y, Li J-T, Zhao S, Zeng D-H (2019) Soil respiration response to precipitation reduction in a grassland and a mongolian pine plantation in semi-arid northeast china[J]. J For Res 30(5):10
Huang Y, Xin Z, Liu J, Liu Q (2022) Divergences of soil carbon turnover and regulation in alpine steppes and meadows on the Tibetan Plateau. Sci Total Environ 814:152687. https://doi.org/10.1016/j.scitotenv.2021.152687
Hui R, Zhao R, Liu L, Li X (2022) Effect of snow cover on water content, carbon and nutrient availability, and microbial biomass in complexes of biological soil crusts and subcrust soil in the desert. Geoderma 406:115505. https://doi.org/10.1016/j.geoderma.2021.115505
IBM Corp. (2019) IBM SPSS Statistics for Windows (Version 26.0) [Computer software]. IBM Corp., Armonk, NY
Jobbágy EG, Jackson RB (2000) The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol Appl 10(2):423–436. https://doi.org/10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2
Kline R (2011) Principles and practice of structural equation modeling, 3rd ed. Guilford Publications
Kögel-Knabner I, Ekschmitt K, Flessa H, Guggenberger G, Matzner E, Marschner B, von Lützow M (2008) An integrative approach of organic matter stabilization in temperate soils: linking chemistry, physics, and biology. J Plant Nutr Soil Sci 171(1):5–13. https://doi.org/10.1002/jpln.200700215
Kværnø SH, Øygarden L (2006) The influence of freeze–thaw cycles and soil moisture on aggregate stability of three soils in Norway. Catena 67(3):175–182. https://doi.org/10.1016/j.catena.2006.03.011
Li H, Wu Y, Chen J, Zhao F, Wang F, Sun Y, Zhang G, Qiu L (2021a) Responses of soil organic carbon to climate change in the Qilian Mountains and its future projection. J Hydrol 596:126110. https://doi.org/10.1016/j.jhydrol.2021.126110
Li H, Wu Y, Liu S, Xiao J (2021b) Regional contributions to interannual variability of net primary production and climatic attributions. Agric Meteorol 303:108384. https://doi.org/10.1016/j.agrformet.2021.108384
Li P, Sayer EJ, Jia Z, Liu W, Wu Y, Yang S, Wang C, Yang L, Chen D, Bai Y, Liu L (2020) Deepened winter snow cover enhances net ecosystem exchange and stabilizes plant community composition and productivity in a temperate grassland. Glob Change Biol 26(5):3015–3027. https://doi.org/10.1111/gcb.15051
Li Q, Liu Y, Kou D, Peng Y, Yang Y (2022) Substantial non-growing season carbon dioxide loss across Tibetan alpine permafrost region. Glob Change Biol 28(17):5200–5210. https://doi.org/10.1111/gcb.16315
Liu W, Zhang Z, Wan S (2009) Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob Change Biol 15:184–195. https://doi.org/10.1111/j.1365-2486.2008.01728.x
Liu H, Mi Z, Lin L, Wang Y, Zhang Z, Zhang F, Wang H, Liu L, Zhu B, Cao G, Zhao X, Sanders NJ, Classen AT, Reich PB, He J-S (2018) Shifting plant species composition in response to climate change stabilizes grassland primary production. Proc Natl Acad Sci 115(16):4051–4056. https://doi.org/10.1073/pnas.1700299114
Liu N, Zhang Y, Chang S, Kan H, Lin L (2012) Impact of grazing on soil carbon and microbial biomass in typical steppe and desert steppe of inner Mongolia. PLoS One 7(5):e36434. https://doi.org/10.1371/journal.pone.0036434
Liu Z, Zhou Q, Ma Q, Kuang W, Daryanto S, Wang L, Wu J, Liu B, Zhu J, Cao C, Li X, Kou Z, Shou W, Qian J, Liu M, Xin Z, Cui X, Liang W (2023) Scale effect of climate factors on soil organic carbon stock in natural grasslands of northern China. Ecol Ind 146:109757. https://doi.org/10.1016/j.ecolind.2022.109757
Lu J, Feng S, Wang S, Zhang B, Ning Z, Wang R, Chen X, Yu L, Zhao H, Lan D, Zhao X (2023) Patterns and driving mechanism of soil organic carbon, nitrogen, and phosphorus stoichiometry across northern China’s desert-grassland transition zone. Catena 220:106695. https://doi.org/10.1016/j.catena.2022.106695
Luo Z, Feng W, Luo Y, Baldock J, Wang E (2017) Soil organic carbon dynamics jointly controlled by climate, carbon inputs, soil properties and soil carbon fractions. Glob Change Biol 23(10):4430–4439. https://doi.org/10.1111/gcb.13767
Ma F, Zhao C, Milne R, Ji M, Chen L, Liu J (2010) Enhanced drought-tolerance in the homoploid hybrid species Pinus densata: implication for its habitat divergence from two progenitors. New Phytol 185(1):204–216. https://doi.org/10.1111/j.1469-8137.2009.03037.x
Maestre FT, Le Bagousse-Pinguet Y, Delgado-Baquerizo M, Eldridge DJ, Saiz H, Berdugo M, Gozalo B, Ochoa V, Guirado E, García-Gómez M, Valencia E, Gaitán JJ, Asensio S, Mendoza BJ, Plaza C, Díaz-Martínez P, Rey A, Hu H-W, He J-Z, … Gross N (2022) Grazing and ecosystem service delivery in global drylands. Science 378(6622):915–920. https://doi.org/10.1126/science.abq4062
McSherry ME, Ritchie ME (2013) Effects of grazing on grassland soil carbon: a global review. Glob Change Biol 19(5):1347–1357. https://doi.org/10.1111/gcb.12144
Meng L-H, Yang J, Guo W, Tian B, Chen G-J, Yang Y-P, Duan Y-W (2017) Differentiation in drought tolerance mirrors the geographic distributions of alpine plants on the Qinghai-Tibet Plateau and adjacent highlands. Sci Rep 7(1):42466. https://doi.org/10.1038/srep42466
Nelson DW, Sommers LE (1975) Determination of total nitrogen in natural waters. J Environ Qual 4(4):465–468. https://doi.org/10.2134/jeq1975.00472425000400040009x
Nielsen UN, Ball BA (2015) Impacts of altered precipitation regimes on soil communities and biogeochemistry in arid and semi-arid ecosystems. Glob Change Biol 21(4):1407–1421. https://doi.org/10.1111/gcb.12789
Olsen SR, Watanabe FS, Bowman RA (1983) Evaluation of fertilizer phosphate residues by plant uptake and extractable phosphorus. Soil Sci Soc Am J 47(5):952–958. https://doi.org/10.2136/sssaj1983.03615995004700050022x
Palpurina S, Wagner V, von Wehrden H, Hájek M, Horsák M, Brinkert A, Hölzel N, Wesche K, Kamp J, Hájková P, Danihelka J, Lustyk P, Merunková K, Preislerová Z, Kočí M, Kubešová S, Cherosov M, Ermakov N, German D, … Chytrý M (2017). The relationship between plant species richness and soil pH vanishes with increasing aridity across Eurasian dry grasslands. Glob Ecol Biogeogr 26(4):425–434. https://doi.org/10.1111/geb.12549
Pärtel M (2002) Local plant diversity patterns and evolutionary history at the regional scale. Ecology 83(9):2361–2366. https://doi.org/10.2307/3071796
Phukubye K, Mutema M, Buthelezi N, Muchaonyerwa P, Cerri C, Chaplot V (2022) On the impact of grassland management on soil carbon stocks: a worldwide meta-analysis. Geoderma Reg 28:e00479. https://doi.org/10.1016/j.geodrs.2021.e00479
Piao S, Fang J, Zhou L, Zhu B, Tan K, Tao S (2005) Changes in vegetation net primary productivity from 1982 to 1999 in China. Glob Biogeochem Cycles 19(2). https://doi.org/10.1029/2004GB002274
Piao S, Wang X, Park T, Chen C, Lian X, He Y, Bjerke JW, Chen A, Ciais P, Tømmervik H, Nemani RR, Myneni RB (2020) Characteristics, drivers and feedbacks of global greening. Nat Rev Earth Environ 1(1):14–27. https://doi.org/10.1038/s43017-019-0001-x
Pielou EC (1966) Shannon’s formula as a measure of specific diversity: its use and misuse. Am Nat 100(914):463–465. https://doi.org/10.1086/282439
Qin W, Wang Y, Yuan X, Zhang Q, Wang X, Zhao H, Zhu B (2023) Responses of soil carbon dynamics to precipitation and land use in an Inner Mongolian grassland. Plant Soil 491(1):85–100. https://doi.org/10.1007/s11104-022-05858-8
R Core Team (2024) R: a language and environment for statistical computing [Computer software]. R Foundation for Statistical Computing, Vienna
Reeder JD, Schuman GE (2002) Influence of livestock grazing on C sequestration in semi-arid mixed-grass and short-grass rangelands. Environ Pollut 116(3):457–463. https://doi.org/10.1016/S0269-7491(01)00223-8
Reeder JD, Schuman GE, Morgan JA, LeCain DR (2004) Response of organic and inorganic carbon and nitrogen to long-term grazing of the shortgrass steppe. Environ Manage 33(4):485–495. https://doi.org/10.1007/s00267-003-9106-5
Ren S, Terrer C, Li J, Cao Y, Yang S, Liu D (2024) Historical impacts of grazing on carbon stocks and climate mitigation opportunities. Nat Clim Chang 14(4):380–386. https://doi.org/10.1038/s41558-024-01957-9
Rosseel Y (2012) lavaan: an R package for structural equation modeling. J Stat Softw 48:1–36. https://doi.org/10.18637/jss.v048.i02
Schönbach P, Wan H, Gierus M, Bai Y, Müller K, Lin L, Susenbeth A, Taube F (2011) Grassland responses to grazing: effects of grazing intensity and management system in an Inner Mongolian steppe ecosystem. Plant Soil 340(1):103–115. https://doi.org/10.1007/s11104-010-0366-6
Schuman GE, Reeder JD, Manley JT, Hart RH, Manley WA (1999) Impact of grazing management on the carbon and nitrogen balance of a mixed-grass rangeland. Ecol Appl 9(1):65–71. https://doi.org/10.1890/1051-0761(1999)009[0065:IOGMOT]2.0.CO;2
Shannon C, Weaver W (1949) The mathematical theory of communication. University of Illinois 34(310):312–313. https://doi.org/10.2307/3611062
Shi Z, Xu X, Souza L, Wilcox K, Jiang L, Liang J, Xia J, García-Palacios P, Luo Y (2016) Dual mechanisms regulate ecosystem stability under decade-long warming and hay harvest. Nat Commun 7(1):11973. https://doi.org/10.1038/ncomms11973
Smith P (2012) Soils and climate change. Curr Opin Environ Sustain 4(5):539–544. https://doi.org/10.1016/j.cosust.2012.06.005
Stocker BD, Zscheischler J, Keenan TF, Prentice IC, Seneviratne SI, Peñuelas J (2019) Drought impacts on terrestrial primary production underestimated by satellite monitoring. Nat Geosci 12(4):264–270. https://doi.org/10.1038/s41561-019-0318-6
Sullivan S, Rohde R (2002) On non-equilibrium in arid and semi-arid grazing systems. J Biogeogr 29(12):1595–1618. https://doi.org/10.1046/j.1365-2699.2002.00799.x
Sun J, Wang Y, Piao S, Liu M, Han G, Li J, Liang E, Lee TM, Liu G, Wilkes A, Liu S, Zhao W, Zhou H, Yibeltal M, Berihun ML, Browning D, Fenta AA, Tsunekawa A, Brown J, … Tsubo M (2022) Toward a sustainable grassland ecosystem worldwide. Innovation 3(4). https://doi.org/10.1016/j.xinn.2022.100265
Tierney GL, Fahey TJ, Groffman PM, Hardy JP, Fitzhugh RD, Driscoll CT (2001) Soil freezing alters fine root dynamics in a northern hardwood forest. Biogeochemistry 56(2):175–190. https://doi.org/10.1023/A:1013072519889
Volik O, Petrone RM, Price JS (2021) Soil respiration and litter decomposition along a salinity gradient in a saline boreal fen in the Athabasca Oil Sands Region. Geoderma 395:115070. https://doi.org/10.1016/j.geoderma.2021.115070
Walkley A, Black IA (1934) An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37(1):29
Wang G, Mao J, Fan L, Ma X, Li Y (2022a) Effects of climate and grazing on the soil organic carbon dynamics of the grasslands in Northern Xinjiang during the past twenty years. Glob Ecol Conserv 34:e02039. https://doi.org/10.1016/j.gecco.2022.e02039
Wang N, Xia L, Goodale CL, Butterbach-Bahl K, Kiese R (2021) Climate change can accelerate depletion of montane grassland C stocks. Glob Biogeochem Cycles 35(10):e2020GB006792. https://doi.org/10.1029/2020GB006792
Wang S, Wang Q, Adhikari K, Jia S, Jin X, Liu H (2016) Spatial-temporal changes of soil organic carbon content in Wafangdian, China. Sustainability 8(11):11. https://doi.org/10.3390/su8111154
Wang S, Wilkes A, Zhang Z, Chang X, Lang R, Wang Y, Niu H (2011) Management and land use change effects on soil carbon in northern China’s grasslands: a synthesis. Agric Ecosyst Environ 142(3):329–340. https://doi.org/10.1016/j.agee.2011.06.002
Wang S, Zhang S, Lin X, Li X, Li R, Zhao X, Liu M (2022b) Response of soil water and carbon storage to short-term grazing prohibition in arid and semi-arid grasslands of China. J Arid Environ 202:104754. https://doi.org/10.1016/j.jaridenv.2022.104754
Wang T, Zhang Z, Li Z, Li P (2017) Grazing management affects plant diversity and soil properties in a temperate steppe in northern China. Catena 158:141–147. https://doi.org/10.1016/j.catena.2017.06.020
Wang Y, Li Y, Ye X, Chu Y, Wang X (2010) Profile storage of organic/inorganic carbon in soil: from forest to desert. Sci Total Environ 408(8):1925–1931. https://doi.org/10.1016/j.scitotenv.2010.01.015
Wang Y, Liu H, Chung H, Yu L, Mi Z, Geng Y, Jing X, Wang S, Zeng H, Cao G, Zhao X, He J-S (2014) Non-growing-season soil respiration is controlled by freezing and thawing processes in the summer monsoon-dominated Tibetan alpine grassland. Glob Biogeochem Cycles 28(10):1081–1095. https://doi.org/10.1002/2013GB004760
Wang Y, Wang Z, Li H, Shen T, Zhang X, Li J, Han G (2023) Grazing decreased soil organic carbon by decreasing aboveground biomass in a desert steppe in Inner Mongolia. J Environ Manage 347:119112. https://doi.org/10.1016/j.jenvman.2023.119112
Wiens JA (1984) Resource systems, populations, and communities. A new ecology novel approaches to interactive systems
Wiesmeier M, Spörlein P, Geuss U, Hangen E, Haug S, Reischl A, Schilling B, von Lützow M, Kögel-Knabner I (2012) Soil organic carbon stocks in southeast Germany (Bavaria) as affected by land use, soil type and sampling depth. Glob Change Biol 18(7):2233–2245. https://doi.org/10.1111/j.1365-2486.2012.02699.x
Wipf S, Rixen C (2010) A review of snow manipulation experiments in Arctic and alpine tundra ecosystems. Polar Res 29(1):1. https://doi.org/10.3402/polar.v29i1.6054
Wu J, Loucks OL (1995) From balance of nature to hierarchical patch dynamics: a paradigm shift in ecology. Q Rev Biol 70(4):439–466
Wu J, Zhang Q, Li A, Liang C (2015) Historical landscape dynamics of Inner Mongolia: patterns, drivers, and impacts. Landsc Ecol 30(9):1579–1598. https://doi.org/10.1007/s10980-015-0209-1
Wu Y, Guo Z, Li Z, Liang M, Tang Y, Zhang J, Miao B, Wang L, Liang C (2022) The main driver of soil organic carbon differs greatly between topsoil and subsoil in a grazing steppe. Ecol Evol 12(8):e9182. https://doi.org/10.1002/ece3.9182
Xu T, Zhang M, Ding S, Liu B, Chang Q, Zhao X, Wang Y, Wang J, Wang L (2021) Grassland degradation with saline-alkaline reduces more soil inorganic carbon than soil organic carbon storage. Ecol Ind 131:108194. https://doi.org/10.1016/j.ecolind.2021.108194
Xu Z, Li Z, Liu H, Zhang X, Hao Q, Cui Y, Yang S, Liu M, Wang H, Gielen G, Song Z (2018) Soil organic carbon in particle-size fractions under three grassland types in Inner Mongolia, China. J Soils Sediments 18(5):1896–1905. https://doi.org/10.1007/s11368-018-1951-1
Yang B, Gong J, Li X, Zhang Z, Shi J, Wang B, Zhang W, Liu Y (2023) Integrated response of carbon sequestration along co-varying gradients of climate and grazing intensity in a desert steppe ecosystem. Appl Soil Ecol 186:104845. https://doi.org/10.1016/j.apsoil.2023.104845
Yang B, Gong J, Zhang Z, Wang B, Zhu C, Shi J, Liu M, Liu Y, Li X (2019a) Stabilization of carbon sequestration in a Chinese desert steppe benefits from increased temperatures and from precipitation outside the growing season. Sci Total Environ 691:263–277. https://doi.org/10.1016/j.scitotenv.2019.06.481
Yang K, Zhu J, Zhang M, Yan Q, Sun OJ (2010) Soil microbial biomass carbon and nitrogen in forest ecosystems of Northeast China: a comparison between natural secondary forest and larch plantation. J Plant Ecol 3(3):175–182. https://doi.org/10.1093/jpe/rtq022
Yang X-D, Ali A, Xu Y-L, Jiang L-M, Lv G-H (2019b) Soil moisture and salinity as main drivers of soil respiration across natural xeromorphic vegetation and agricultural lands in an arid desert region. Catena 177:126–133. https://doi.org/10.1016/j.catena.2019.02.015
Yang Y, Tilman D, Furey G, Lehman C (2019c) Soil carbon sequestration accelerated by restoration of grassland biodiversity. Nat Commun 10(1):718. https://doi.org/10.1038/s41467-019-08636-w
Yu Z, Zhang W, Liu Y, Cao L, Bai Y, Xin X, Zhou Y, Xie Z, Hu T, Yang P (2023) Dynamics of SOC density and driving factors during the restoration of artificial grassland and abandoned farmland in Mu Us Desert, China. Catena 224:106991. https://doi.org/10.1016/j.catena.2023.106991
Zhang J, Zuo X, Zhou X, Lv P, Lian J, Yue X (2017a) Long-term grazing effects on vegetation characteristics and soil properties in a semiarid grassland, northern China. Environ Monit Assess 189(5):216. https://doi.org/10.1007/s10661-017-5947-x
Zhang K, Su Y, Yang R (2019a) Variation of soil organic carbon, nitrogen, and phosphorus stoichiometry and biogeographic factors across the desert ecosystem of Hexi Corridor, northwestern China. J Soils Sediments 19(1):49–57. https://doi.org/10.1007/s11368-018-2007-2
Zhang N, Wan S, Guo J, Han G, Gutknecht J, Schmid B, Yu L, Liu W, Bi J, Wang Z, Ma K (2015) Precipitation modifies the effects of warming and nitrogen addition on soil microbial communities in northern Chinese grasslands. Soil Biol Biochem 89:12–23. https://doi.org/10.1016/j.soilbio.2015.06.022
Zhang R, Wang Z, Han G, Schellenberg MP, Wu Q, Gu C (2018) Grazing induced changes in plant diversity is a critical factor controlling grassland productivity in the Desert Steppe, Northern China. Agric Ecosyst Environ 265:73–83. https://doi.org/10.1016/j.agee.2018.05.014
Zhang W, Zhang F, Qi J, Hou F (2017b) Modeling impacts of climate change and grazing effects on plant biomass and soil organic carbon in the Qinghai-Tibetan grasslands. Biogeosciences 14(23):5455–5470. https://doi.org/10.5194/bg-14-5455-2017
Zhang Y, Shao C, Chen J, Lu G (2019) Understanding 3000 years of nomadic history of Mongolian Grassland: Grassland, livestock and population. J Arid Land Resour Environ 33(6):40–45. https://doi.org/10.13448/j.cnki.jalre.2019.166
Zhao R, Hui R, Wang Z, Liu L, Xie M, An L (2016) Winter snowfall can have a positive effect on photosynthetic carbon fixation and biomass accumulation of biological soil crusts from the Gurbantunggut Desert, China. Ecol Res 31(2):251–262. https://doi.org/10.1007/s11284-016-1335-1
Zhao W, Zhang R, Cao H, Tan W (2019) Factor contribution to soil organic and inorganic carbon accumulation in the Loess Plateau: Structural equation modeling. Geoderma 352:116–125. https://doi.org/10.1016/j.geoderma.2019.06.005
Zhao Y, Ding Y, Hou X, Li FY, Han W, Yun X (2017) Effects of temperature and grazing on soil organic carbon storage in grasslands along the Eurasian steppe eastern transect. PLoS One 12(10):e0186980. https://doi.org/10.1371/journal.pone.0186980
Zhou G, Zhou X, He Y, Shao J, Hu Z, Liu R, Zhou H, Hosseinibai S (2017) Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: a meta-analysis. Glob Change Biol 23(3):1167–1179. https://doi.org/10.1111/gcb.13431
Zhou Y, Hartemink AE, Shi Z, Liang Z, Lu Y (2019) Land use and climate change effects on soil organic carbon in North and Northeast China. Sci Total Environ 647:1230–1238. https://doi.org/10.1016/j.scitotenv.2018.08.016
Zhu X, Shao M (2018) Distribution, stock, and influencing factors of soil organic carbon in an alpine meadow in the hinterland of the Qinghai-Tibetan Plateau. J Earth Syst Sci 127(5):71. https://doi.org/10.1007/s12040-018-0974-8
Acknowledgements
This study was funded by the Open Project Program of Ministry of Education Key Laboratory of Ecology and Resources Use of the Mongolian Plateau (KF2023004), the National Natural Science Foundation of Inner Mongolia (2021MS03032), the Major Science and Technology Projects of Inner Mongolia (2021ZD0044). The authors would also thank Yi Hu, Ziwen Wei, Haozhe Feng, and Zhenhao Zhang for their assistance and the anonymous reviewers for their insightful comments.
Funding
This study was funded by the Open Project Program of Ministry of Education Key Laboratory of Ecology and Resources Use of the Mongolian Plateau (KF2023004), the National Natural Science Foundation of Inner Mongolia (2021MS03032), the Major Science and Technology Projects of Inner Mongolia (2021ZD0044).
Author information
Authors and Affiliations
Contributions
MT and MS planned and designed the research. YC, MT, and MS performed the research. ZQ analyzed the data and wrote the manuscript. ZQ, YC, MT, and MS revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Responsible Editor: Ruzhen Wang.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Quan, Z., Cheng, Y., Tsubo, M. et al. Sensitivity and regulation factors of soil organic carbon content in steppe and desert—steppe grasslands of the Mongolian Plateau. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06867-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06867-5