
Abstract
Aims
The cycling of nutrients from plant litter has key implications for the functioning of terrestrial ecosystems by controlling nutrient availability and net primary production. Despite extensive research on the effects of global change on ecosystem functioning, the direct implications of global change on stoichiometry and nutrient dynamics during litter decomposition remain poorly understood. To address this gap, we conducted a meta-analysis.
Methods
We analysed 178 experiments that simulated (i) warming, (ii) drought, (iii) increased water availability, (iv) N enrichment, (v) P enrichment, and (vi) combined N and P (N + P) enrichment. We compared earlier (approximately six months) and later (approximately one year) stages of decomposition and analysed the specific effects taking into account climate and plant type.
Results
The C:N and C:P ratios decreased in most warming and nutrient enrichment scenarios, leading to losses of litter C content, while the N:P ratio remained more resilient and affected by water availability. Furthermore, the abundance of resources (water and N + P) fosters the decomposition of litter. The nutrient mobilisation increases for both P and N under non-limited nutrient enrichment and it is faster for N than for P when water increases its availability. Nutrient enrichment was relevant in later stages of decomposition.
Conclusions
Our study provides insights into the fate of litter decomposition and its stoichiometric dynamics in response to drivers of global change. Concerning scenarios of C release and N and P immobilisation were identified. However, further experimentation and analysis are necessary to consider all interacting drivers.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Nutrient stoichiometry is a crucial factor in controlling important ecological processes, such as the growth rate and life strategies of organisms, as well as ecosystem diversity, structure, and functionality (Sterner and Elser 2002; Sardans et al. 2012b; Peñuelas et al. 2019). In particular, litter decomposition is one of the key processes that determines ecosystem functioning (Zhang et al. 2018; Lanuza et al. 2019). Currently, the biosphere is facing various transformations, including climate-related changes like global warming and shifts in precipitation patterns (IPCC 2021), as well as changes in the availability of elements such as C and N due to anthropogenic activities (Gruber and Galloway 2008; Sundquist 1993). However, the availability of other essential nutrients like P and K are not increasing to the same extent (Sardans et al. 2012a, 2021; Peñuelas et al. 2020). These changes in nutrient stoichiometry and imbalances are altering the "elementome" (i.e., the elemental composition of ecosystems) of soils (Peñuelas et al. 2019; Fernández-Martínez 2022; Peñuelas and Sardans 2022). Understanding how these shifts in elementome, together with climate change, affect nutrient cycles in plant-soil systems is crucial for the development of global change models, conservation strategies, and food security policies.
Litter decomposition is an important process for understanding plant-soil nutrient cycles, as litter serves as a crucial source of mineralizable nutrients for microbial and plant metabolism in terrestrial ecosystems (Silveira et al. 2011), that controls the availability of nutrients in soils, cation-exchange capacity, soil pH, and the formation of humus (Berg and McClaugherty 2008). The release of nutrients during litter decomposition allows microbes and plants to take them up. Litter decomposition also plays a significant role at the biospheric level, affecting the exchange of greenhouse gases and the carbon stocks in soil (Walkiewicz et al. 2021), which can, in turn, affect climate change (Parker et al. 2018). Drivers of global change such as increasing temperatures and precipitation can accelerate the rate of litter decomposition (Zhang et al. 2008), while droughts can slow it down and reduce the C content of litter (Deng et al. 2021). Additionally, nutrient enrichment can have diverse effects on litter decomposition rates, increasing them in grasslands and wetlands but decreasing them in forests (Su et al. 2022).
Litter decomposition and nutrient mineralisation also depends on the species that compose the vegetation cover, or the environmental factors, like climate, that characterise the ecosystem; and will determine the shifting of elementomes in soils. Temperature and precipitation can affect litter decomposition directly by regulating the activity of decomposers, and indirectly through changes in plant species composition and abundance, as a result, in variations in litter quality and quantity (Petraglia et al. 2019). Colder and drier climates present higher stabilisation of organic material (Mueller et al. 2018), while warmer and wetter climates are more likely to promote faster decomposition by the soil food webs (García-Palacios et al. 2013). On the other hand, litter quality is considered a key regulator of the decomposition dynamics in litter (Zhang et al. 2008). Decomposing plant litter from broadleaved forest beds of grass species is less constrained than conifer tree litter, whose lignin content and allelopathic substances are generally higher (Rahman et al. 2013; Liu et al. 2019). The effect of global change drivers then differ depending on the litter quality. N additions had been reported accelerating decomposition in low-lignin litters, while this effect was inhibiting in high-ligning litters (Hobbie et al. 2012; Knorr et al. 2005). Another aspect to consider is the stage of the litter decomposition, with earlier stages characterised by the decomposition of non-lignified cellulose and hemicellulose; and the leaching of soluble compounds; while later stages encompasses the degradation of the lignified tissues, requiring a more complex metabolism (Djukic et al. 2018).
Litter decomposition has been a widely studied field, even in relation to global change analyses (Zhang et al. 2008; Deng et al. 2021; Su et al. 2022). However, there is still a lack of an all-encompassing understanding of how litter stoichiometry during the decomposition process varies in relation to climate change and nutrient enrichment. Most review studies focus on one aspect of litter dynamics, such as litter decomposition rate (Zhang et al. 2008; Su et al. 2022) or the effect on a particular elemental content (Deng et al. 2021), and others, are barely focused on the dynamics of litter decomposition (Sardans et al. 2012a). Our objective with this analysis is to examine how nutrient addition and climate change, two important indicators of global change, affect the stoichiometry of litter decomposition, and to detect how it differs depending on and climate and the plant group producing the litter through a global assessment. To achieve this goal, we gathered data from experiments measuring the stoichiometric relationships of at least two elements (C, N, P; K was also contempled) in decomposing litter and where treatments simulated one of four drivers of global change: (i) warming, (ii) drought, (iii) irrigation (an increase in water availability), or (iv) N and P enrichment (separately or combined). We evaluated nutrient dynamics in the earlier (approximately six months) and later (approximately one year) stages of decomposition and also considered factors such as climate and plant type. By focusing only on studies that examined the simultaneous response of at least two elements, we reduced the number of studies available for the meta-analysis, but we were able to investigate elemental ratios. Additionally, we examined the effects on the decomposition rate and individual contents of C, N, and P.
Global change affects litter decomposition dynamics: an increment in the decomposition rate results in a preferential mineralisation of C to N and P; and N to P (Sardans et al. 2012a). When comparing experimental and control plots, we expect to find the following: (i) Increases in temperature will reduce the C:N, C:P and N:P ratios as decomposer activity is enhanced; (ii) Water scarcity will elevate C:N, C:P and N:P ratios, as decomposition is hampered, but the opposite effect is expected if water availability increases; (iii) Nutrient enrichments would also increase decomposition activity, leading to reductions of C:N and C:P ratios. However, the N:P ratio would differ depending on the enrichment – increasing if the enrichment in N dominates and decreasing if it is P dominated. This effect would be expected to be (iv) greater in earlier decomposition stages than in later, as the preferential mineralization of nutrients by decomposers may disappear with time when the carbon-rich – easy-to-decompose – litter component disappears. Additionally, we expect to find different responses to different climates and plant types. As an example, (v) we expect to find that increases of temperature and changes in precipitation are more likely to affect colder and dry climates compared to warm and wetter ones; and finally. (vi) we expect conifer, high-ligning, litter being less affected by nutrient enrichments compared to grasses of broadleaved species.
Materials and methods
Data collection
To compile a dataset of the effects of global change drivers on litter stoichiometry, we conducted a bibliographic search on the 1st of February 2022 for peer-reviewed papers on Scopus, Web of Science, and Google Scholar. Three search strings were performed on title, keywords, and abstract: (i) “(litter AND decomposition) AND (warming OR (increase* AND temperature*)) AND experiment* AND ((N:P) OR (C:N) OR (C:P))” for global warming as an agent of change; (ii) “(litter AND decomposition) AND ((N OR P) AND (deposit* OR fertiliz* OR addition)) AND experiment* AND ((N:P) OR (C:N) OR (C:P))” for nutrient enrichment (e.g. fertilisation or enhanced deposition) as an agent of change, and (iii) “(litter AND decomposition) AND (precipitation OR rainfall OR drought) AND (reduc* OR addition OR increment* OR exclusion) AND experiment* AND ((N:P) OR (C:N) OR (C:P))” for changes in the availability of precipitation as an agent of global change. After screening the manuscripts, we selected studies that reported: (i) plots with experimental and control conditions, (ii) measurements of the variables in the experimental and control plots that were conducted under the same temporal and spatial scales, and where the magnitude of the treatment and experimental duration were clearly recorded and (iii) sufficientdata to calculate the C:N, C:P, and/or N:P ratios of the remaining litter. Our final database contained 43 studies (highlighted with an asterisk in the reference section; Table S1). All data for each study were obtainedfrom the associated published data sets provided by the author/s while extracting data from figures was done using WebPlotDigitizer (Rohatgi 2021). We considered different experiments when (i) the litter type differed, (ii) plots differed geographically within the same study, (iii) different treatments were applied independently, and (iv) plots varied in the intensity of treatment. A PRISMA formatted flow diagram (Moher et al. 2009) is provided in the supplementary material (Figure S1).
We recorded ancillary data for each study, including information for the study site such as geographic location of the sampling sites (latitude, longitude, and elevation), information for the litter species sampled, the litter sampling method (litterbag experiments vs litter collection), the methodological approach to measure each of the elements, and the initial litter biomass and elemental concentrations. The experimental conditions were characterised in three groups: (i) increase in temperature, the intensity of which was recorded in °C, (ii) enrichment of N, P, or both N and P, the intensity of which was recorded as g m−2 y−1, and (iii) changes in the amount of precipitation, which was recorded as the percent decrease or increase in water availability relative to the control plots in a year. While we recorded this ancillary data from the paper, we also recorded the climate zone of the location (polar, cold, temperate, arid, or tropical) using the recorded longitude and latitude, and the kgc package of R (Bryant et al. 2017). If the plant type (angiosperm tree, gymnosperm tree, shrubs, and grasses; all herbaceous species were considered grasses) was not specified in the text, we filled the gaps using the Global Biodiversity Information Facility (GBIF 2021). We ultimately analysed six data sets: i) warming treatments (47 experiments), ii) enrichment with N only (55 experiments), iii) enrichment with P only (13 experiments), iv) enrichment with both N and P (38 experiments), vi) decreases in the amount of precipitation (drought treatment, 13 experiments), vii) increases in the amount of precipitation (irrigation treatment, 12 experiments). We rejected experiments that entailed more than one of the selected treatments (warming, irrigation, drought, N enrichment, P enrichment, and N + P enrichment) at the same time. After applying these requirements our database contained 178 experiments. The starting date and associated duration (days) for each experiment were compiled for 123 of the experiments. We then compiled both the percentage of remaning control and experimental litter masses (only for the litterbag experiments), the elemental concentrations, elemental contents (percentages of the initial quantity of the element remaining after the experiment) C, N and P; and the C:N and C:P litter ratios. The standard deviation and number of replicates for all these variables for both the control and treatment plots were also recorded. If provided, we also collected the K concentration.
Calculation of variables
Depending on the study, the elemental data were the concentration of the element, the content that remained relative to the initial value, and/or the ratios of the elements. We calculated the concentrations of C, N, P, and K, the contents of C, N, and P remaining relative to the initial values, and the C:N, C:P, and N:P ratios (on a mass basis, for the mass remaining) for all experiments when possible. We initially recorded all sampling dates for each experiment. First, we subsetted our database considering sampling dates resembling experiments that last for approximately half a year (180 d). This subset contained the samples representing the "earlier" stage of decomposition with a median of 233 ± 26 days (see below). To test changes in later decomposition stages (prediction iv), we subsetted the database filtering sampling dates closer to three years (1080 d), but the median resulted similar to one year: 412 ± 32 day. Thus, this database contained experiments where litter was at a “later” stage of decomposition. A detailed information about the days of experiments foe each study can be found in Table S2. We thus constructed a database for all experiments at an “earlier” stage of decomposition, and others at a “later” stage. Additionally, we calculated the decomposition rate based on the classical definition (Olson 1963):
where \(k\) is the decomposition rate, \(t\) is the duration (in days) of the experiment, which was recorded for every experiment, and \({X}_{0}\) and \({X}_{t}\) are the initial litter mass and the litter mass remaining at the time of sampling, respectively (in grams). We also applied this formula to the initial and remaining litter elemental contents (for obtaining the rates of release of the elements), and to the initial and remaining litter elemental ratios (for obtaining the rates of change of the elemental ratios).
Before determining the stoichiometric effects of global change drivers, we assessed the effects of each treatment on the mass remaining at the end of the experiment and the decomposition rates of the litter biomasses. We determined the effects of the treatments on the litter concentrations of C, N, and P and calculated the elemental contents remaining (% from the initial) after the experiment (when the remaining contents were not provided). Due to the lower number of experiments providing initial litter masses compared to those providing elemental concentrations, the sample size used for the content analysis was smaller than that for the concentration analysis. Ratios were calculated as the quotient of the elemental contents, but we used the ratios provided when these data were not available. To calculate k (rate of change) associated with the shifting ratios, we needed the initial ratios at the start of the decomposition experiments.
The final data set
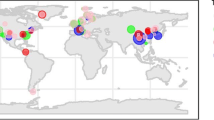
Our final data set contained 42 sampling locations, (the locations of the mesocosms were not considered) (Fig. 1); and data for litter from 71 species. From the 178 experiments selected, and based on the Köppen-Geiger classification (Beck et al. 2018), we found that 7 of the experiments were carried out in tropical climates, 48 in arid climates, 55 in temperate climates, 33 in cold climates, and 32 in polar climates. The starting months for the litterbag experiments differed across studies: 10 experiments started from January to March, 63 from April to June, 12 from July to September, and 21 from October to December. Plant species and climate were correlated variables.

Global map of the sampling locations (coloured dots) used in the meta-analysis
Statistical analysis
We used the metafor package (v2.4–0; Viechtbauer 2010) in R Studio (v4.1.2; R Core Team 2021) for the statistical analyses. We calculated the sizes of the effects of each treatment (warming, N enrichment, P enrichment, N + P enrichment, increase in water availability, and decrease in water availability) on the remaining litter biomass, the contents of C, N, P, and K, the release rate of these contents, the C:N, C:P, and N:P ratios, and the rates of change of these ratios. To calculate the effect size of a treatment, we calculated the standard mean difference (SMD; Hedges 1981) with the function escalc, where the numerator of SMD was set as the value of the variable under the treatment minus the value of the variable under the control (retaining negative signs). Thus, a positive treatment effect (SMD) indicates that the treatment group has, on average, a higher value than the control group. On the other hand, a negative treatment effect indicates that the treatment group has a lower value compared to the control group. We constructed meta-analytic models as random-effect models using the Restricted Maximum Likelihood method (REML; Cook 2014) with the rma.mv function. We rejected the meta-analytic models when the number of experiments was lower than 5, and those rejected models can be consulted in Table S3We calculated the general effects of the treatments over the experiments using the study as the random factor to account for the possible non-independence of experiments from the same study. For the treatments that significantly affected the variables, we calculated the sensitivity of the effect size to the intensity of the treatment by including the intensity as a moderator. We also explored the effects of the treatments over the experiments taking into account factors such as climate (arid, cold, temperate, polar, or tropical; according to Köppen-Geiger classification) and plant type (broadleaved tree, conifer tree, grass, or shrub). We added the factor (climate orplant type) as a moderator of the random model, with the study as the random factor. In the case of plant type, the random factor was plant nested within the study. For those analyses, we only used data from the earlier decomposition database and the effect of decomposition rate, C, N, P, and K contents and ratios.
We also conducted a publication bias analysis using Egger’s regression test (Egger et al 1997), adapted to the rma.mv function. We adjusted the metaregression models by including the standard error of the effect sizes as a moderator. If the intercept significantly differed from zero (P < 0.1, following Egger et al. 1997), we inferred that the relationship between precision and the size of the studies is asymmetrical, indicating bias.
Results
General effects
We explored the general effects of the treatments over the experiments. The detailed data of the significant effects found in the general effect analysis are presented in Table S4. We display the data in C, N, and P ratios and contents in the litter remaining after decomposition. More analysis on C, N, and P concentration are available on Supplementary Material (Figure S2 and Table S5).
We identified publication bias in 26.1% of the analyses, which warrants some caution on the inferences presented here.’. The analysis affected by the bias are depicted in Figs. 2, 3 and 4 (see figure captions) and in Table S4.

Effect sizes (± 95% CI) of the treatments on the amount of the initial litter mass remaining after approximately half a year (earlier stage, green bars, above) or approximately a year (later stage, pink bars, below) of decomposition (a) and the decomposition rates (b). Coloured bars indicate significant effect sizes. The boxes depict the number of experiments (similar to earlier and later stages).. Treatments are only represented in the plots if the number of experiments is ≥ 5. Publication bias is highlighted with warning symbols (triangle with exclamation mark inside) in the title of the treatment. Green warning symbols depict publication analysis for the earlier stage; while pink warning symbols are for the later stages

Effect sizes (± 95% CI) of the treatments on the content and k for the contents and release rates of C (a, b), N (c, d), and P (e, f) approximately half a year (earlier stage, green bars, above) or approximately a year (later stage, pink bars, below) of decomposition (a) and the decomposition rates (b). Coloured bars indicate significant effect sizes. The boxes depict the number of experiments (similar to earlier and later stages). Treatments are only represented in the plots if the number of experiments is ≥ 5. Publication bias is highlighted with warning symbols (triangle with exclamation mark inside) in the title of the treatment. Green warning symbols depict publication analysis for the earlier stage; while pink warning symbols are for the later stages

Effect sizes (± 95% CI) of the treatments on the stoichiometric ratios and associated change rates for C:N (a, b), C:P (c, d), and N:P (e, f) approximately half a year (earlier stage, green bars, above) or approximately a year (later stage, pink bars, below) of decomposition (a) and the decomposition rates (b). Coloured bars indicate significant effect sizes. The boxes depict the number of experiments (similar to earlier and later stages). Treatments are only represented in the plots if the number of experiments is ≥ 5. Publication bias is highlighted with warning symbols (triangle with exclamation mark inside) in the title of the treatment. Green warning symbols depict publication analysis for the earlier stage; while pink warning symbols are for the later stages
Litter decomposition
The amount of mass remaining was lower in N + P enriched plots than in controls in both the earlier (6 months) and later stages (12 months). However, an increasing effect in litter decomposition rates was only found for later stages, while earlier stages did not show a significant effect. Plots under the irrigation treatment presented significantly higher decomposition rates than control plots in the later stages of decomposition (Fig. 2). When analysing the sensitivity of the effects to the treatment intensity, we found that in the case of N + P enrichment experiments, the more N is added the higher the effect, but this was only found for later stages. Warming, drought, and N or P enrichment (alone) did not show any effects on litter mass decomposition.
Effects on the nutrient contents remaining
Compared to control plots, C content in litter decreased in the warmed treatments during earlier and later stages of decomposition. C content in litter also decreased in N + P enriched plots in the later stages of decomposition. N content in litter was incremented in later stages of decomposition under N + P enrichment conditions. No effect was found for N and P remaining content or release rate (Fig. 3). There were not enough (< 5) P enrichment experiments valid for assessing its effects on element contents remaining since the beggining of the experiments.
Effect on the nutrient ratios
Compared to control plots, litter C:N ratios decreased in P enriched plots in both earlier and later stages of decomposition, and with N enrichment in the earlier stage of decomposition. C:N change rate increased under N enrichment in later stages of decomposition and under N + P enrichment in both stages of decomposition (Fig. 5). The sensitivity analysis revealed that the more P was accounted for in the P enrichment experiments, the higher the effect is over the C:N ratio. The same happened with N enrichment intensity in the N enrichment experiments, the more N was accounted for, the higher the effect was over the C:N ratio. Litter C:P ratio decreased, compared to control plots, under warming treatment in both earlier and later stages of decomposition. Warming also increased the C:N ratio change rate in earlier and later stages. N + P enrichment treatment decreased C:P ratios in the later stage of decomposition while increasing C:P change rate in the later stage of decomposition (Fig. 5). The sensitivity analysis revealed an indirect relationship between the intensity of the warming and the decreasing effect on the C:P ratio. The litter N:P ratio decreased, compared to control plots, with the irrigation treatment for the earlier stage of decomposition, however no other effects were found for N:P ratios of N:P change rates (Fig. 4).

Graphical summary of the general effects of the meta-analysis on decomposition rate and C, N, and P contents and their ratios. General effects, while specific effects only from earlier stages of decomposition. Significant effects from the climate and plant type-specific analysis that were only found from one experiment are excluded from the figure and the Discussion section. Consult Table S4, Table 2, and Table 3 for p values and number of studies for each analysis
Effects considering the climate of the sampling area
The warming experiments from cold climates presented a decreasing effect on C contents. Regarding N content, irrigation treatment had a decreasing effect on cold climates while drought had an increasing effect on arid climate. Although general changes in P content were not significant, different climate conditions led to significant changes in the P content significantly changed. In this regard, under warming experiments, we found an increasing effect on P content in cold climates, and decreasing in polar climates. In drought experiments, we found an increasing effect on P content in arid climates. In irrigation experiments, we found an increasing effect on P content in temperate climates. Last, in N + P enrichment experiments, we found an increasing effect on P content in temperate climates and a decreasing effect on P content in arid climates.
C:N ratio was also affected by climate in warming and N + P enrichment experiments despite not having any general significant effect. We found a decreasing effect of warming experiments on C:N ratio in cold climates. Also, in N + P enrichment experiments, we found decreasing effect on C:N for polar and temperate climates. In N enrichment experiments, which had a C:N ratio general decreasing effect, Climate-related effects were found for N enrichment experiments:C:N ratio, which showed a general decrease effect, decreasedin cold and tropical climates; C:P ratio was affected by climate in N + P and N enrichment experiments although no general effects were found. We found, for N + P enrichment experiments, a C:P ratio decreasedt in cold and tropical climates. We also found a N enrichment decreasing effect on C:P ratios in cold climates. For warming experiments, which showed a general decreasing of C:P ratios, we found that cold climates were driving the effect. All this information is summarised in Table 1.
Effects considering the plant type
Although there was no general significant effect on litter decomposition rates under warming treatments, we found a increasing effect when looking at grass species Also, in N + P enrichment experiments under conifer trees, an increasing effect on decomposition rate.
We found a general decreasing effect of C contents in warming experiments (Table 1). Additionally, during warming experiments under grass species, a decreasing effect of the C content in the remaining litter was found. Although there was not a general effect for C content on drought experiments, we found a decreasing effect for C content when lookint at shrub species. Despite not finding any general effect for N content in irrigation experiments, we found that in grass species there is a decreasing effect on the N content. P content did not have any general effect for N + P enrichment experiments, but we found an increasing effect on P content driven by litter from broadleaved tree leaves. C:N ratio did not show any general effect for the warming, drought, and N + P enrichment treatments. However, C:N ratios decreased in grass species on warming experiments; in conifer species on N + P enrichment experiments and in grass species in drought experiments; while increased in shrub species in drought experiments. For warming experiments, which showed a general decreasing effect on C:P ratios, we found a decreasing effect for grass species and conifer trees. Irrigation experiments showed a decreasing effect of N:P ratio. This effect was also found for broadleaved trees. All this information is summarised in Table 2.
K concentration
With the recorded data of K concentration in litter (Table S5; Figure S2). We detected a general decreasing effect in K concentration for warmed plots in later stages of decomposition (Table S5). However, while there is not a general effect during early stages of decomposition, there are climate specific effects in the early stage, increasing its concentration in cold climates (p = 0.007, from 5 of the 6 experiments addressing K concentrations in warming experiments), whereas we found an opposite effect for temperate climates, where K concentrations increase (p = 0.044 from 1 of the 6 experiments addressing K concentrations in warming experiments). We also find K concentrations decreasing in irrigated plots from temperate climates (p < 0.001, from 1 of the 6 experiments addressing K concentrations in warming experiments).
Summary of the general effects
To facilitate the comprehension of the results of the meta-analysis, a summary figure was constructed with results groups by the distinct scenarios that the global change experiments portrayed (Fig. 5).
Discussion
Our meta-analysis provides an overview of the dynamics and stoichiometry of litter decomposition under drivers of global change such as warming, changes in water availability, and nutrient enrichment and limitation. The results portray a guide of the C:N:P stoichiometry under the fate of global change.
Effect of warming
Our initial predictions (i) posited that increases in temperature would lead to a decrease in C:N and C:P ratios. Examining the general effects, elevated temperatures resulted in C release and reduced C:P ratios, consistent with the expectations. However, in warming experiments there were no general effects on litter decomposition rate or C:N ratios. Water limitations may be hindering the effect or warming on litter decomposition. Similar results were obtained by Petraglia et al.(2019) in a national decomposition study reflecting a broad range of vegetation types and microclimatic conditions. The effects of temperature increase on litter decomposition were present in moist conditions. Alternatively, they pointed to vegetation type as a possible explanation since most of the moist soils were classified as grasslands. We found similar results: although a general effect of warming on decomposition rate was absent, there was an increasing decomposition rate effect in litter from grass species.
Litter dynamics in cold climates are likely to be disturbed with increments in temperature and precipitation, potentially releasing the old and stabilised pools of organic matter (García-Palacios et al. 2013). When accounting for climate, we detected that C release under warming treatment was notably prominent in cold climates. The warming effect of litter decompostion in cold climates is enhanced since decomposition in these environments is limited by low temperatures which are raised in the warming treatments. Additionally, litter decomposition is promoted since cold climates are charecterised by precipitation exceeding evaporation (Beck et al. 2018), being non water-limited by definition. Consequently, our results confirmprevious predictions (Sardans et al. 2012a) that warming effects on C:N and C:P ratios are only pronounced under water-abundant scenarios. In cold climates, P is immobilised in litter, explaining why the warming-induced decrease was only found for C:P.
Concerning plant type, we found that warming in grasses significantly increased decomposition rates and reduced C contents, C:N, and C:P ratios, aligning with previous studies (Song et al. 2014). In the experiments with grass litter, which is generally characterised by lower N concentration than tree litter (Yuan and Chen 2009; Vallicrosa et al. 2022), decomposition was hindered. This could elucidate why warming effects on decomposition rate and C:N were exclusive to grasses.Warming in cold grasslands and conifer forests enhances C release and P immobilisation in litter. Released C may contribute to soil and atmospheric carbon, exacerbating greenhouse effects and global warming (Cornelissen et al. 2007; Hobbie et al 2002). P-limitation arises from P immobilisation in litter, rendering it unavailable as a resource for plants. Consequently, C:N and C:P ratios are reduced in these scenarios, potentially impacting soil trophic webs. Another concern, that was nos addressed in the meta-analysis, is the community shift towards a more recalcitrant leaf litter, shrub dominance, in response to increasing temperatures in cold climates (Cornelissen et al. 2007; Jackson et al. 2002).
Lastly, K release from the litter increased during the warming treatments. The effect was relevant in cold climates, that is non-water limited. However, we detected an opposite (increasing litter K concentration) effect in a case in a Mediterranean shrubland (Saura-Mas et al. 2012). Since K is an important element for plant water use efficiency (Xu et al 2021), it might be immobilised in litter as a result of biological strategies to cope with water limitation.
Effect of water availability
Our predictions (ii) postulated that water scarcity would elevate C:N and C:P ratios, hindering decomposition, with the opposite expected under increased water availability. C:N, C:P, and the contents of C, N, or P did not exhibit consistent increases or decreases in both drought and irrigation scenarios. General effects were observed for irrigation, increasing litter decomposition rate and decreasing the N:P ratio. Irrigation-induced reduction in the N:P ratio may be attributed to the low solubility and limited soil mobility of P (Manzoni et al. 2010), favoured by rising soil water content.
We also predicted (v) that increases in temperature and changes in precipitation will have greater effects in cold and dry climates (cold, arid and polar) compared to warm and wetter ones (temperate and tropical). In the case of warming, addressed in the section above, we did not find a greater effect of decomposition in cold climates, but we did for C release, P immobilisation and C:P ratio decrease. While there were no other specific effects related to climate. Regarding changes in the precipitation patterns, we did not find any effect regarding decomposition rates, but N and P were immobilised during drought treatment; and N released in irrigated plots from cold climates. Temperate and tropical climates were not affected specifically by any climate change driver. In cold climates (non water-limited), the accelerated N release may be caused by a facilitated hydrolysis of litter compounds rich in C and N, such as cellulose, lignin, and macroproteins. This could result in a higher mineralization rate for C and N compared to P. Conversely, under drought conditions in arid climates, N and P immobilisation suggesting an increased nutrient use efficiency under water-limiting conditions (Sardans et al. 2012a).
Examining the specific effects of plant type, the general decreasing effect on the N:P ratio was primarily driven by experiments involving litter from broadleaved trees. Additionally, a significant effect on N release was noted in grasses, potentially explained by the lower N and higher P concentrations in the senescent leaves of grasses compared to broadleaved trees (Yuan and Chen 2009; Vallicrosa et al. 2022). Broadleaved tree litter, with higher N:P ratios, may be more sensitive to changes in water availability than grass litter..
Effect of nutrient enrichment
Our prediction (iii) suggested that nutrient enrichments would increase decomposition activity, leading to reductions in C:N and C:P ratios. However, the N:P ratio would differ depending on the enrichment, increasing in N-enriched treatments and decreasing in P-enriched treatments. For C:N ratios, the prediction was confirmed, generally decreasing under N, P, and N + P enrichment treatments. Under N + P enrichment, the decomposition rate increased, C:P ratio decreased (in contrast to N or P enrichment scenarios), and C was released while N was immobilised. A meta-analysis by Ferreira et al. (2015) reported increased litter decomposition only when both N and P were applied, suggesting a co-limitation of these elements for faster decomposition. Imbalanced nutrient enrichment, favouring N over P, could lead to N-limitation, explaining the lack of general effects of N enrichment on decomposition (Colin and Waring 2017). The observed release of C and immobilisation of N in N + P enrichment scenarios suggest a preferential metabolism of C-rich compounds by decomposing microorganisms (Manzoni et al. 2021; Hobbie et al. 2012)).
In climate-specific analysis, cold climates exhibited a prominent decrease in C:N and C:P ratios under N enrichment and C:P decrease under N + P enrichment. The combination of nutrient enrichment and water-abundant environments intensified the activity of fast-decomposing communities, leading to a preferential mobilisation of C. A 17 year old experiment on N addition in cold climates (Maaroufi et al. 2017) suggests that C mobilisation from litter in response to N enrichment was driven by changes in soil microbiota. C:P ratio also prominently decreased in tropical climates, where water is not scarce (Beck et al. 2018). Under N + P enrichment, P immobilisation occurred in temperate climates but was released in arid climates. Further analysis addressing the interaction of nutrient enrichment and water availability is necessary to fully understand this effect.
We predicted (vi) that conifer, high-lignin, litter would be less affected by nutrient enrichments compared to broadleaved trees or grass litter. However, we detected the opposite: conifer tree litter had a prominent decomposition rate increment and C:N decrease. Broadleaved tree litter exhibited a significant P immobilisation effect in N + P treatment. The differing N:P ratios in conifer and broadleaved tree litter, with lower N:P ratios and higher lignin content in conifer leaves (higher P contents, Rahman et al. 2013), may explain why conifer tree litter is less P-limited, fostering higher decomposition rates. The correlation between climate and plant type as moderators could contribute to these effects, especially in temperate climates where broadleaved tree experiments predominated. Opposite effects between grass and tree species were not found, as could be expected by previous meta-analysis comparing the effects of N addition in forest and grasslands (Su et al. 2022).
Earlier vs later stages
Contrary to our predictions (iv) expecting a decrease in C:N and C:P ratios only in earlier stages of decomposition, we did not find a consistent pattern.Instead, distinct effects between earlier and later stages were observed, particularly under N + P enrichment. In cases where there was a difference between stages, the later stage exhibited significant effects. Earlier stages of decomposition are less specific, as they consist on the mineralisation of labile C. However, later stages of decomposition require complex metabolites to degrade the recalcitrant tissues (Djukic et al. 2018). Impacts on the microbiota due to nutrient enrichments (Maaroufi and De Long 2020) may be more critical during a less generalistc later stage of decompositon. Moreover, the systemic impact of nutrient enrichment on litter decomposition stoichiometry appears relevant in the long term, supporting the notion that N + P enrichment effects are cumulative (Hanley et al. 2021; Seabloom et al. 2021). This aligns with explanations for N immobilisation under N + P enrichment, where N-limitation results from a prolonged metabolic process.
Limitations of the study
Our initial question was about how drivers of global change interacted with the elemental stoichiometry in leaf litter, so we only selected articles that sampled at least two elements simultaneously in litter samples for control and treated plots. The number of studies/experiments to test our hypothesis was thus reduced. We, therefore, used data from experiments of decomposition in litterbags but also from litter sampling without decomposition experiments, from which we could only extract data for the differences in elemental concentrations or ratios between control and treated plots but not for the rates of mineralisation or decomposition. Caution is needed when interpreting these results, some aspects of the meta-analysis are low in number of experiments. Regarding the sampled sites (Fig. 1), there is a clear example of the geographic bias, frequently found in meta-analyses, where most of the field studies are within the latitudinal ranges of China, Europe, and North America. Also, all arid places are found in Asia. More experiments in other areas are needed in order to infer more precise global patterns.
Another issue to consider is that the processes of decomposition vary across ecosystems, chemical composition of litter, and stages of decomposition. The days of experiments were not consistent across studies; and even if they were, the experiments from each study began in different seasons, which made the appropriate comparison of experiments difficult. Thus, our results can serve as general information for determining how nutrient dynamics may behave under current global change and may differ across ecosystems and decomposing time. The methods used for analysing elemental compositions were not homogeneous and could introduce bias to our results. We provide in Table S2 information about the measurement methodologies applied on each study, and the experiment days that were considered for the earlier and later stages. For further information about specific climate and litter species, we detail the characteristics of each study used in Table S1. is important to highlight the presence of publication bias in 26.1% of the analysis. However, the models took into account the possible non-independence of experiments from the same study variability between studies. The use of plant type and climate, which are correlated variables, should also be carefully interpreted for specific analysis independently, avoiding cross-referencing. The use of the rates of elemental release and change of elemental ratios was useful for identifying effects that were not apparent when focusing only on the elemental contents in the litter.
Another aspect that was not contemplated in this study is the soil biota, crucial for the litter decomposition process (Njoroge et al. 2021), which is affected by nutrient enrichment (Nessel et al 2021) and climate change (García-Palacios et al. 2013; Maaroufi and De Long 2020). Finally, we focused on experiments and observations of one driving factor, but all drivers of global change interact in nature. Therefore, multifactorial experiments and observations are strongly recommended.
Conclusions
Our meta-analysis explored the effects of warming, changes in water availability, and nutrient enrichment, which are known drivers of global change, on the processes of elemental mineralisation/immobilisation and on the stoichiometric relationships during the decomposition of litter. The effects can vary across scales, but they can help us determine the generalised effects of global change on the nutrient dynamics of terrestrial ecosystems.
As the main general effects, we found that the abundance of resources (water and N + P) fosters the decomposition of litter. Warming and nutrient enrichment led to losses in litter C content and reduction of the C:N and C:P ratios. C can move from the litter to the soil but also to the atmosphere as CO2, contributing to the greenhouse effect and generating a feedback loop, because more warming derived from the greenhouse effect may intensify the release of C from litter. Subsequently, the C: N and C: P ratios were reduced in these scenarios, with potential impacts on the soil trophic webs. This effect is prominent in grassland and coniferous forests in cold climates.
The N:P ratios were only affected by the increase in the availability of water in broadleaved tree leaf litter, which may be caused by the higher solubility of N with respect to P. This effect was only apparent at earlier stages of decomposition. Our analysis found that N:P ratios, in contrast with C:N and C:P ratios, were more resilient to changes in temperature, water scarcity, and nutrient enrichment.
We identified immobilisation of P in the litter, which may hinder P-limitation scenarios, in warming and cold climates; arid environments experiencing droughts; temperate environments with increasing water availability; and in temperate or on broadleaved tree leaf litters experiencing N and P enrichment. N immobilisation was detected under the combined enrichment of N and P at later stages of decomposition. Arid environments experiencing droughts are subject to both N and P immobilisation. This exploratory analysis is useful for distinguishing between various impacts of climate change and nutrient enrichment on ecosystem nutrient dynamics. We aim to inspire new and warranted research on the unknown mechanisms responsible for the results and interactions amongst multiple drivers. Future research with experimental designs is encouraged research to simulate scenarios of global change for exploring the stoichiometric implications of the simulated scenarios, with the maximum possible number of drivers interacting and in different stages of decomposition.
Data availability
Data will be made available upon request.
References
The asterisk (*) indicates the studies used in the meta-analysis
*Aerts R, Callaghan TV, Dorrepaal E et al (2012) Seasonal climate manipulations have only minor effects on litter decomposition rates and N dynamics but strong effects on litter P dynamics of sub-arctic bog species. Oecologia 170:809–819. https://doi.org/10.1007/s00442-012-2330-z
*Aerts R, Wallén B, Malmer N, De Caluwe H (2001) Nutritional constraints on Sphagnum-growth and potential decay in northern peatlands. J Ecol 89:292–299. https://doi.org/10.1046/j.1365-2745.2001.00539.x
*Allison SD, Lu Y, Weihe C et al (2013) Microbial abundance and composition influence litter decomposition response to environmental change. Ecology 94:714–725. https://doi.org/10.1890/12-1243.1
*Andresen LC, Michelsen A, Jonasson S et al (2010) Plant nutrient mobilization in temperate heathland responds to elevated CO2, temperature and drought. Plant Soil 328:381–396. https://doi.org/10.1007/s11104-009-0118-7
Beck H, Zimmermann N, McVicar T et al (2018) Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci Data 5:180214. https://doi.org/10.1038/sdata.2018.214
Berg B, McClaugherty C (2008) Plant litter: decomposition, humus formation, carbon sequestration (2nd ed.). Springer
*Blok D, Elberling B, Michelsen A (2016) Initial stages of tundra shrub litter decomposition may be accelerated by deeper winter snow but slowed down by spring warming. Ecosystems 19:155–169. https://doi.org/10.1007/s10021-015-9924-3
Bryant C, Wheeler N, Rubel F, French R (2017). kgc: Koeppen-Geiger Climatic Zones. R package version 1.0.0.2. https://CRAN.R-project.org/package=kgc
Cook NR (2014) Restricted Maximum Likelihood: Introduction. Wiley StatsRef Stat Ref Online 1–2. https://doi.org/10.1002/9781118445112.stat05953
Colin A, Waring B (2017) Nitrogen limitation of decomposition and decay: How can it occur? Glo Chang Biol 24:1414–1427. https://doi.org/10.1111/gcb.13980
*Cornejo FH, Varela A, Wright SJ, Tropical SJ (1994) Tropical Forest Litter Decomposition under Seasonal Drought : Nutrient Release , Fungi and Bacteria Author ( s ): Fernando H . Cornejo , Amanda Varela and S . Joseph Wright Published by : Wiley on behalf of Nordic Society Oikos Stable URL https://www.js. OIKOS 70:183–190
Cornelissen J, Bodegom P, Aerts R (2007) Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes. Ecol Lett 10:619–627. https://doi.org/10.1111/j.1461-0248.2007.01051.x
*Day TA, Ruhland CT, Xiong FS (2008) Warming increases aboveground plant biomass and C stocks in vascular-plant-dominated Antarctic tundra. Glob Chang Biol 14:1827–1843. https://doi.org/10.1111/j.1365-2486.2008.01623.x
Deng L, Peng C, Kim DG et al (2021) Drought effects on soil carbon and nitrogen dynamics in global natural ecosystems. Earth-Sci Rev 214:103501. https://doi.org/10.1016/j.earscirev.2020.103501
Djukic I, Kepfer-Rojas S, Schmidt I (2018) Early stage litter decomposition across biome. Sci Total Environ. https://doi.org/10.1016/j.scitotenv.2018.01.012
Egger M, Smith G, Schneider M, Minder C (1997) Bias in meta-analysis detected by a simple, graphical test. Br Med J 315:629–634. https://doi.org/10.1136/bmj.315.7109.629
*Fang XM, Zhang XL, Zong YY et al (2021) Responses of leaf litter decomposability to nitrogen and phosphorus additions are associated with cell wall carbohydrate composition in a subtropical plantation. Plant Soil 467:359–372. https://doi.org/10.1007/s11104-021-05099-1
Fernández-Martínez M (2022) From atoms to ecosystems: elementome diversity meets ecosystem functioning. New Phytol 234:35–42. https://doi.org/10.1111/nph.17864
Ferreira V, Graça M, Elosegui A (2015) A meta-analysis of drought effects on litter decomposition in streams. Hydrobiologia 850:1715–1735. https://doi.org/10.1007/s10750-023-05181-y
García-Palacios P, Maestre F, Kattge J, Wall H (2013) Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol Lett 16(8):1045–1053. https://doi.org/10.1111/ele.12137
GBIF: The Global Biodiversity Information Facility (2021) What is GBIF?. Available from https://www.gbif.org/what-is-gbif. Accessed 13 Feb 2021
*Gong J, Zhu C, Yang L et al (2020) Effects of nitrogen addition on above-and belowground litter decomposition and nutrient dynamics in the litter-soil continuum in the temperate steppe of Inner Mongolia, China. J Arid Environ 172:104036. https://doi.org/10.1016/j.jaridenv.2019.104036
*Gossiaux A, Rollin M, Guérold F et al (2020) Temperature and nutrient effects on the relative importance of brown and green pathways for stream ecosystem functioning: A mesocosm approach. Freshw Biol 65:1239–1255. https://doi.org/10.1111/fwb.13474
Gruber N, Galloway J (2008) An Earth-system perspective of the global nitrogen cycle. Nature 451:293–296. https://doi.org/10.1038/nature06592
Hanley T, Bowen J, Kearns P et al (2021) Short- and long-term effects of nutrient enrichment on salt marsh plant production and microbial community structure. J Ecol 109:3779–3793. https://doi.org/10.1111/1365-2745.13756
Hedges L (1981) Distribution theory for Glass’s estimator of effect size and related estimators. J Educ Stat 6:107–128. https://doi.org/10.3102/10769986006002107
*Hong J, Lu X, Ma X, Wang X (2021) Five-year study on the effects of warming and plant litter quality on litter decomposition rate in a Tibetan alpine grassland. Sci Total Environ 750:142306. https://doi.org/10.1016/j.scitotenv.2020.142306
Hobbie S, Schimel J, Trumbore S (2002) Controls over carbon storage and turnover in high-latitude soils. Glob Chang Biol 6:196–210. https://doi.org/10.1046/j.1365-2486.2000.06021.x
Hobbie S, Eddy W, Buyarski C et al (2012) Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment. Ecol Monog 82:389–405. https://doi.org/10.1890/11-1600.1
*Hou S, Yin J, Yang J et al (2017) Consistent responses of litter stoichiometry to N addition across different biological organization levels in a semi-arid grassland. Plant Soil 421:191-202. https://doi.org/10.1007/s11104-017-3446-z
*Hu X, Sun Z (2021) Effects of exogenous nitrogen import on variations of nutrient in decomposing litters of Suaeda salsa in coastal marsh of the Yellow River estuary, China. Environ Sci Pollut Res 28:33165-33180. https://doi.org/10.1007/s11356-021-12926-6
*Huang X, Chen J, Wang D et al (2021) Simulated atmospheric nitrogen deposition inhibited the leaf litter decomposition of Cinnamomum migao H. W. Li in Southwest China. Sci Rep 11:1-13. https://doi.org/10.1038/s41598-021-81458-3
IPCC (2021) Climate Change 2021: The physical science basis. Contribution of working group i to the sixth assessment report of the intergovernmental panel on climate change. Cambridge, United Kingdom and New York, NY, US In press. https://doi.org/10.1017/9781009157896
Jackson R, Banner J, Jobbágy E (2002) Ecosystem carbon loss with woody plant invasion of grasslands. Nature 418:623–626. https://doi.org/10.1038/nature00910
*Jiang L, Kou L, Li S (2018) Alterations of early-stage decomposition of leaves and absorptive roots by deposition of nitrogen and phosphorus have contrasting mechanisms. Soil Biol Biochem 127:213–222. https://doi.org/10.1016/j.soilbio.2018.09.037
*Jonasson S, Castro J, Michelsen A (2004) Litter, warming and plants affect respiration and allocation of soil microbial and plant C, N and P in arctic mesocosms. Soil Biol Biochem 36:1129–1139. https://doi.org/10.1016/j.soilbio.2004.02.023
Knorr M, Frey D, Curtis P (2005) Nitrogen Additions and litter decomposition: a meta-analysis. Ecology 86:3252–3257. https://doi.org/10.1890/05-0150
Lanuza O, Casanoves F, Delgado D, Van den Meersche K (2019) Leaf litter stoichiometry affects decomposition rates and nutrient dynamics in tropical forests under restoration in Costa Rica. Restor Ecol 27:549–558. https://doi.org/10.1111/rec.12893
*Li L, Liu B, Gao X et al (2019) Nitrogen and phosphorus addition differentially affect plant ecological stoichiometry in desert grassland. Sci Rep 9:1–8. https://doi.org/10.1038/s41598-019-55275-8
Liu Y, Shen X, Wang L et al (2019) Litter chemical quality strongly affects forest floor microbial groups and ecoenzymatic stoichiometry in the subalpine forest. Ann for Sci. https://doi.org/10.1007/s13595-019-0890-3
*Longhini VZ, Cardoso AS, Berça AS et al (2021) Nitrogen fertilizer increased litter deposition and litter N in warm-climate grasslands. Nutr Cycl Agroecosyst 119:247–258. https://doi.org/10.1007/s10705-021-10119-8
Maaroufi N, Nordin A, Palmqvist K et al (2017) Nitrogen enrichment impacts on boreal litter decomposition are driven by changes in soil microbiota rather than litter quality. Sci Rep. https://doi.org/10.1038/s41598-017-04523-w
Maaroufi N, De Long J (2020) Global change impacts on forest soils: linkage between soil biota and carbon-nitrogen-phosphorus stoichiometry. Front For Glob Change. https://doi.org/10.3389/ffgc.2020.00016
*Madeira M, Araújo MC, Pereira JS (1995) Effects of water and nutrient supply on amount and on nutrient concentration of litterfall and forest floor litter in Eucalyptus globulus plantations. Plant Soil 168–169:287–295. https://doi.org/10.1007/BF00029340
Manzoni S, Trofymow J, Jackson R et al (2010) Stoichiometric controls dynamics on carbon, nitrogen, and phosphorus in decomposing litter. Ecol Monogr 80:89–106. https://doi.org/10.1890/09-0179.1
Manzoni S, Chakrawal A, Spohn M et al (2021) Modeling Microbial Adaptations to Nutrient Limitation During Litter Decomposition. Front For Glob Change. https://doi.org/10.3389/ffgc.2021.686945
*Mei L, Zhang P, Cui G et al (2022) Arbuscular mycorrhizal fungi promote litter decomposition and alleviate nutrient limitations of soil microbes under warming and nitrogen application. Appl Soil Ecol 171:104318. https://doi.org/10.1016/j.apsoil.2021.104318
Moher D, Liberati A, Tetzlaff J, Altman DG, PRISMA Group (2009) Preferred reporting items for systematic reviews and meta-analyses:the PRISMA statement. Ann Intern Med 151:264–269. https://doi.org/10.1016/j.jclinepi.2009.06.005
Mueller P, Schile-Beers M, Mozdzer J et al (2018) Global-change effects on early-stage decomposition processes in tidal wetlands – implications from a global survey using standardized litter. Biogeosciences 15:3189–3202. https://doi.org/10.5194/bg-15-3189-2018
Nessel M, Konnovitch T, Romero G (2021) Nitrogen and phosphorus enrichment cause declines in invertebrate populations: a global meta-analysis. Biol Rev 96:2617–2637. https://doi.org/10.1111/brv.12771
*Niu S, Sherry RA, Zhou X et al (2010) Nitrogen regulation of the climate-carbon feedback: Evidence from a long-term global change experiment. Ecology 91:3261–3273. https://doi.org/10.1890/09-1634.1
Njoroge D, Chen S, Zuo J (2021) Soil fauna accelerate litter mixture decomposition globally, especially in dry environments. J Ecol 110:659–672. https://doi.org/10.1111/1365-2745.13829
Olson J (1963) Energy storage and the balance of producers and decomposers in ecological systems. Ecology 44:322–331. https://doi.org/10.2307/1932179
Parker TC, Sanderman J, Holden RD et al (2018) Exploring drivers of litter decomposition in a greening Arctic: results from a transplant experiment across a treeline. Ecology 99:2284–2294. https://doi.org/10.1002/ecy.2442
Petraglia A, Cacciatori C, Chelli S et al (2019) Litter decomposition: effects of temperature driven by soil moisture and vegetation type. Plant Soil 435:187–200. https://doi.org/10.1007/s11104-018-3889-x
Peñuelas J, Fernández-Martínez M, Ciais P et al (2019) The bioelements, the elementome, and the biogeochemical niche. Ecology 100:1–15. https://doi.org/10.1002/ecy.2652
Peñuelas J, Janssens IA, Ciais P et al (2020) Anthropogenic global shifts in biospheric N and P concentrations and ratios and their impacts on biodiversity, ecosystem productivity, food security, and human health. Glob Chang Biol 26:1962–1985. https://doi.org/10.1111/gcb.14981
Peñuelas J, Sardans J (2022) The global nitrogen-phosphorus imbalance. Science (80- ) 375:266–267. https://doi.org/10.1126/science.abl4827
R Core Team (2021) R: A language and environment for statistical computing
Rahman M, Tsukamoto J, Rahman M et al (2013) Lignin and its effects on litter decomposition in forest ecosystems. Chem Ecol. https://doi.org/10.1080/02757540.2013.790380
*Ribeiro C, Madeira M, Araújo MC (2002) Decomposition and nutrient release from leaf litter of Eucalyptus globulus grown under different water and nutrient regimes. For Ecol Manage 171:31–41. https://doi.org/10.1016/S0378-1127(02)00459-0
Rohatgi A (2021) WebPlotDigitizer. Version 4.5. https://automeris.io/WebPlotDigitizer
*Rustad LE, Fernandez IJ (1998) Soil warming: consequences for foliar litter decay in a spruce-fir forest in Maine, USA. Soil Sci Soc Am J 62:1072–1080. https://doi.org/10.2136/sssaj1998.03615995006200040031x
*Sanaullah M, Chabbi A, Girardin C et al (2014) Effects of drought and elevated temperature on biochemical composition of forage plants and their impact on carbon storage in grassland soil. Plant Soil 374:767–778. https://doi.org/10.1007/s11104-013-1890-y
Sardans J, Janssens IA, Ciais P et al (2021) Recent advances and future research in ecological stoichiometry. Perspect Plant Ecol Evol Syst 50:125611. https://doi.org/10.1016/j.ppees.2021.125611
Sardans J, Peñuelas J (2015) Potassium: A neglected nutrient in global change. Glob Ecol Biogeogr 24:261–275. https://doi.org/10.1111/geb.12259
Sardans J, Rivas-Ubach A, Peñuelas J (2012a) The C:N: P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect Plant Ecol Evol Syst 14:33–47. https://doi.org/10.1016/j.ppees.2011.08.002
Sardans J, Rivas-Ubach A, Peñuelas J (2012b) The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives. Biogeochemistry 111:1–39. https://doi.org/10.1007/s10533-011-9640-9
*Saura-Mas S, Estiarte M, Peñuelas J, Lloret F (2012) Effects of climate change on leaf litter decomposition across post-fire plant regenerative groups. Environ Exp Bot 77:274–282. https://doi.org/10.1016/j.envexpbot.2011.11.014
Seabloom E, Adler P, Alberti J et al (2021) Increasing effects of chronic nutrient enrichment on plant diversity loss and ecosystem productivity over time. Ecology. https://doi.org/10.1002/ecy.3218
*Shi J, Gong J, Baoyin T tao et al (2021) Short-term phosphorus addition increases soil respiration by promoting gross ecosystem production and litter decomposition in a typical temperate grassland in northern China. Catena 197:104952. https://doi.org/10.1016/j.catena.2020.104952
Silveira ML, Reddy KR, Comerford NB (2011) Litter decomposition and soluble carbon, nitrogen, and phosphorus release in a forest ecosystem. Open J Soil Sci 01:86–96. https://doi.org/10.4236/ojss.2011.13012
*Song P, Zhang K, Ma K, Guo J (2014) Impacts of global warming on litter decomposition. Acta Ecologica Sinica 34(6):1327–1339. https://doi.org/10.5846/stxb201210251479
Sterner RW, Elser JJ (2002) Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere
*Su Y, Le J, Ma X et al (2021a) Soil burial has a greater effect on litter decomposition rate than nitrogen enrichment in alpine grasslands. J Plant Ecol 14:1047–1059. https://doi.org/10.1093/jpe/rtab044
*Su Y, Ma X, Gong Y et al (2022) Global Patterns and Drivers of Litter Decomposition Under Nitrogen Enrichment: A Meta-Analysis. Front For Glob Chang 5:1–11. https://doi.org/10.3389/ffgc.2022.895774
Su Y, Ma X, Le J et al (2021b) Decoupling of nitrogen and phosphorus in dominant grass species in response to long-term nitrogen addition in an Alpine Grassland in Central Asia. Plant Ecol 222:261–274. https://doi.org/10.1007/s11258-020-01103-3
Sundquist E (1993) The Global Carbon Dioxide Budget. Science 259:934–941. https://doi.org/10.1126/science.259.5097.934
*Tie L, Fu R, Peñuelas J et al (2020) The additions of nitrogen and sulfur synergistically decrease the release of carbon and nitrogen from litter in a subtropical forest. Forests 11:1–16. https://doi.org/10.3390/f11121280
Vallicrosa H, Sardans J, Maspons J, Penuelas J (2022) Global distribution and drivers of forest biome foliar nitrogen to phosphorus ratio (N:P). Glob Ecol Biogeogr 31(10). https://doi.org/10.1111/geb.13457
*Van Meeteren MJM, Tietema A, Westerveld JW (2007) Regulation of microbial carbon, nitrogen, and phosphorus transformations by temperature and moisture during decomposition of Calluna vulgaris litter. Biol Fertil Soils 44:103–112. https://doi.org/10.1007/s00374-007-0184-z
Viechtbauer W (2010) Conducting meta-analyses in R with the metafor. J Stat Softw 36:1–48. https://doi.org/10.18637/jss.v036.i03
Walkiewicz A, Rafalska A, Bulak P et al (2021) How can litter modify the fluxes of CO2 and CH4 from forest soils? A mini-review. Forests 12(9). https://doi.org/10.3390/f12091276
*Wang Q, Kwak JH, Choi WJ, Chang SX (2018) Decomposition of trembling aspen leaf litter under long-term nitrogen and sulfur deposition: Effects of litter chemistry and forest floor microbial properties. For Ecol Manage 412:53–61. https://doi.org/10.1016/j.foreco.2018.01.042
*Wang X, Xu Z, Lü X et al (2017) Responses of litter decomposition and nutrient release rate to water and nitrogen addition differed among three plant species dominated in a semi-arid grassland. Plant Soil 418:241–253. https://doi.org/10.1007/s11104-017-3288-8
*Xie T, Shan L, Su P (2020) Drought conditions alter litter decomposition and nutrient release of litter types in an agroforestry system of China. Ecol Evol 10:8018–8029. https://doi.org/10.1002/ece3.6264
Xu Q, Fu H, Zhu B et al (2021) Potassium improves drought stress tolerance in plants by affecting root morphology, root exudates and microbial diversity. Metabolites 11:1–17. https://doi.org/10.3390/metabo11030131
*Xu ZF, Pu XZ, Yin HJ et al (2012) Warming effects on the early decomposition of three litter types, Eastern Tibetan Plateau, China. Eur J Soil Sci 63:360–367. https://doi.org/10.1111/j.1365-2389.2012.01449.x
*Yahdjian L, Sala OE, Austin AT (2006) Differential controls of water input on litter decomposition and nitrogen dynamics in the patagonian steppe. Ecosystems 9:128–141. https://doi.org/10.1007/s10021-004-0118-7
Yan W, Shangguan Z, Zhong Y (2021) Responses of mass loss and nutrient release in litter decomposition to ultraviolet radiation. J Soils Sediments 21:698–704. https://doi.org/10.1007/s11368-020-02810-0
*Yavitt JB, Wright SJ, Wieder RK (2004) Seasonal drought and dry-season irrigation influence leaf-litter nutrients and soil enzymes in a moist, lowland forest in Panama. Austral Ecol 29:177–188. https://doi.org/10.1111/j.1442-9993.2004.01334.x
Yuan ZY, Chen HYH (2009) Global trends in senesced-leaf nitrogen and phosphorus. Glob Ecol Biogeogr 18:532–542. https://doi.org/10.1111/j.1466-8238.2009.00474.x
Zhang D, Hui D, Luo Y, Zhou G (2008) Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. J Plant Ecol 1:85–93. https://doi.org/10.1093/jpe/rtn002
Zhang H, Goll DS, Manzoni S et al (2018) Modeling the effects of litter stoichiometry and soil mineral N availability on soil organic matter formation using CENTURY-CUE (v1.0). Geosci Model Dev 11:4779–4796. https://doi.org/10.5194/gmd-11-4779-2018
*Zheng Z, Mamuti M, Liu H et al (2017) Effects of nutrient additions on litter decomposition regulated by phosphorus-induced changes in litter chemistry in a subtropical forest, China. For Ecol Manage 400:123–128. https://doi.org/10.1016/j.foreco.2017.06.002
Zhou K, Lu X, Mori T et al (2018) Effects of long-term nitrogen deposition on phosphorus leaching dynamics in a mature tropical forest. Biogeochemistry 138:215–224. https://doi.org/10.1007/s10533-018-0442-1
Funding
Open Access Funding provided by Universitat Autonoma de Barcelona. This work was supported by the Spanish Government grants PID2020115770RB-I, PID2022-140808NB-I00, and TED2021-132627 B–I00 funded by MCIN, AEI/10.13039/ 501100011033 European Union Next Generation EU/PRTR, the Fundación Ramón Areces grant CIVP20A6621, and the Catalan Government grants SGR 2021–1333 and AGAUR2023 CLIMA 00118.
Author information
Authors and Affiliations
Contributions
Javier de la Casa: Conceptualization, Data extraction, Statistical analysis, Writing. Jordi Sardans.: Conceptualization, Supervision, Reviewing and Editing. Marta Galindo-Díaz: Data Extraction, Reviewing and Editing. Josep Peñuelas: Conceptualization, Supervision, Reviewing and Editing.
Corresponding author
Ethics declarations
Competing interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Ivika Ostonen.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
de la Casa, J., Sardans, J., Galindo, M. et al. Stoichiometry of litter decomposition under the effects of climate change and nutrient enrichment: A meta-analysis. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06718-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06718-3