
Abstract
Background and aims
Iron (Fe) plaque which normally coats rice roots has a strong affinity for phosphorus (P), with a debated effect on plant P uptake. Furthermore, plant responses to P availability shape the rhizospheric environment, possibly affecting the rates of Fe plaque formation and dissolution. The role of Fe plaque to serve as a sink or source of available P may depend on root traits, themselves influenced by P availability. However, the underlying mechanism regulating these interactions remains unclear. In this study, we investigated the effects of P availability on root traits, Fe plaque dynamics and their implications for P uptake and rice plant growth.
Methods
Plants were hydroponically grown for 60 days under P-sufficiency or P-deficiency, with or without Fe plaque. Root traits, rhizosphere acidification, and the rates of Fe plaque formation and dissolution were investigated and linked to differences in rice P content and growth.
Results
P-deficient conditions stimulated root development and promoted Fe plaque formation on the root surface compared to P-sufficient conditions. However, P limited plants exhibited a faster Fe plaque dissolution, along with increased net proton exudation. After 60 d, P-deficient plants showed higher P uptake in the presence of Fe plaque, whereas the opposite was observed in P-sufficient plants, where Fe plaque limited plant P uptake.
Conclusions
The role of Fe plaque in regulating P uptake highly depends on the dynamic nature of this Fe pool that is strictly linked to P availability and regulated by plant responses to P deficiency.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rice (Oryza sativa L.) is the major food crop for over half of the world’s population (Seck et al. 2012). It is commonly cultivated in flooded paddy fields, resulting in oxygen (O2) depleted soils. The decreased redox potential causes the reductive dissolution of iron (Fe) (hydr)oxides, and the release of Fe(II) in soil porewater (Ponnamperuma 1972; Marschner 2021). High concentrations of Fe(II) in porewater may be toxic to rice plants (Sahrawat 2005), which in turn have developed a series of adaptation strategies to cope with high Fe concentrations (Yoshida 1967). Indeed, rice plants respond to anoxic soil conditions by enhancing the longitudinal transport of O2 from shoots to root tips, inducing the formation of aerenchyma, a specialized tissue comprising enlarged gas spaces that can supply the necessary O2 for root respiration (Evans 2004). Some of the O2 conveyed to the roots diffuses into the soil immediately adjacent to the root through radial oxygen loss (ROL), generating redox gradients in the rhizosphere (Colmer 2003). Here, Fe(II) can be rapidly re-oxidized and precipitated as Fe(III) (oxyhydr)oxides on the root surface, resulting in the formation of Fe plaque (Armstrong 1967; Hansel et al. 2001). Iron plaque shows chemical and physical properties similar to those of pedogenic Fe oxides present in soil, and therefore has a high capacity to retain various anions in the soil solution, including phosphorus (P), which is an essential plant macronutrient (Zhang et al. 1999; Hansel et al. 2001; Seyfferth et al. 2010; Amaral et al. 2017).
The availability of P in paddy soils is strongly driven by redox conditions, although the higher soil solution P concentrations under reducing conditions do not necessarily result in a higher plant P uptake (Martinengo et al. 2023). Previously, low P uptake by rice has been attributed to high P sorption capacity of newly formed Fe minerals near the root surface (Zhang et al. 2003; Kirk 2004; Veelen et al. 2020). Despite these findings, the role of Fe plaque in controlling P plant uptake remains controversial (Khan et al. 2016), as previous studies have reported its role as both a barrier/sink (Liang et al. 2006) and a source of plant available P (Zhang et al. 1999). These contrasting observations probably arise from studies overlooking the varying responses of rice plants to different P availabilities, and/or neglecting the interaction between the temporal dynamics in Fe plaque formation/dissolution and plant P acquisition strategies.
The dynamics affecting P retention on Fe plaque minerals can be confounded by the ability of rice plants to alter the rhizosphere environment in response to nutrient deficiency (Zhang et al. 2004). Indeed, it is well acknowledged that many plant species can modify their root traits and architecture as a function of changes in P availability, with a higher partitioning of photosynthates towards root growth under P deficiency (Richardson et al. 2009; Rose et al. 2013). In rice plants, increased root development observed under low P availability is related to an increase in root aerenchyma formation to reduce the metabolic cost of this adaptation mechanism (Kirk and Van Du 1997; Lynch and Brown 2008). This leads to a consequent increase in O2 release into the rhizosphere under P-limiting conditions (Fu et al. 2014).
Notwithstanding a higher formation of Fe plaque was previously attributed to to a higher O2 release in the rhizosphere (Wu et al. 2012), the effects of root modifications in response to P availability on Fe plaque formation, and the implications for P acquisition in rice plants are however still largely debated. Indeed, some studies have observed an increase in Fe plaque formation under low P availability (Liang et al. 2006; Jiaofeng et al. 2022), but mechanisms underlying this phenomenon are not yet clear. Some authors argue that this increase may be due to enhanced ROL, as reported by Fu et al. (2014). Others suggest that it may be related to an increase in root volume, as Kirk and Van Du (1997) did not find a significant difference in O2 release per unit of root volume. Moreover, it remains unclear whether the high affinity of Fe plaque minerals for P combined with the higher amounts of root-associated Fe might emphasize their role as P sink under P limiting conditions, thereby hindering plant growth by impeding P acquisition.
Rice plants growing under P limited conditions are known to have the capability to activate strategies to access mineral-bound P forms through different rhizospheric mechanisms (Kirk and Van Du 1997; Hoffland et al. 2006; Rose et al. 2013). Specifically, the exudation of protons (H+) and low molecular weight organic acids (LMWOAs) from roots may promote the release of sparingly available soil P forms, such as P retained by metal oxides (Hinsinger 2001; Hoffland et al. 2006; Bhattacharyya et al. 2013; Santoro et al. 2022). Unlike other plant species that rely on organic acids to solubilize P by competing for adsorption sites (Hoffland et al. 2006; Bhattacharyya et al. 2013), proton exudation by rice plants is considered as the primary mechanism by which these lowland plants release P from Fe minerals in response to P deficiency, especially considering their relatively low rates of organic acid exudation (Mori et al. 2016). Given the similarity between Fe plaque and Fe oxides present in soil (Hansel et al. 2001; Seyfferth et al. 2010; Amaral et al. 2017), these strategies could be also effective in enhancing the uptake of P associated with Fe plaque, thereby converting this P pool from a P sink into a P source for the plant. The role of Fe plaque to serve as a sink or a source of plant available P and consequently regulate P uptake by rice plants, could therefore be viewed as a dynamic process influenced by the rates of both Fe plaque formation and dissolution as a function of P availability, although evidence remains lacking.
Based on these considerations, we hypothesised that the response of rice plants to P deficiency could (a) induce a greater formation of root-associated Fe plaque as a result of an enhanced root development and ROL, but at the same time (b) favour Fe plaque dissolution (and the release of plaque-associated P) through increased proton exudation, thereby promoting P uptake. As such, we therefore further hypothesise that (c) the capability of Fe plaque to serve as either a P sink or a source of plant available P will be strongly influenced by the dynamic nature of this Fe pool, which is dependent on P availability (i.e., P source under low P availability vs. P sink under high P availability). We tested these hypotheses by growing rice plants in a hydroponic system under P-sufficient or P-deficient conditions. Plants were exposed to multiple Fe-plaque induction periods and analysed for differences in root traits, changes in the rates of Fe plaque formation and contents, and net proton exudation, and differences in P uptake over a 62-d growth period.
Materials and methods
Plant cultivation and Fe plaque induction
To investigate the impact of P availability on the formation and evolution of Fe plaque in rice (O. sativa cv. Selenio) and, consequently, on P uptake by rice, a hydroponic experiment was performed using a factorial design in which plants were grown under either P-deficient (‒P) or P-sufficient (+ P) conditions for 62 days. During this period, half of the plants for each treatment were exposed to Fe plaque forming conditions (+ Fe) while the other half served as a control group without Fe plaque induction (‒Fe). In detail, rice seeds were surface sterilised with H2O2 for 10 min, rinsed thoroughly with distilled water and germinated for three days in the dark at 20 °C. Afterwards, a total of sixteen seedlings of uniform size per treatment were transferred to plastic pots containing half-strength modified Yoshida solution (Yoshida et al. 1972) for the first two weeks and then shifted to full strength nutrient solution (day 0), that was replaced every 4 days. The modified Yoshida solution had the following macronutrient composition (mM): NH4NO3, 1.43; K2SO4, 0.51; CaCl2, 1.0; MgSO4, 1.65; and micronutrient composition (µM): MnCl2, 9.1; (NH4)6Mo7O24·2H2O, 0.52; H3BO3, 18.5; ZnSO4·7H2O, 0.15; CuSO4·5H2O, 0.16 and FeCl3(EDTA), 35.8. The nutrient solution used for plants grown under P-deficient conditions contained 10 µM NaH2PO4, while that for plants grown under P-sufficient conditions contained 100 µM NaH2PO4. The pH of the solution was adjusted to 6.0 by using diluted HCl or NaOH.
Fe plaque induction was carried out twice during the growth period. On the 30th and 45th days after seeding (DAS), half of the plants for each P treatment were removed from the nutrient solution, and their roots were rinsed in deionised water and subsequently transferred to a 0.9 mM (as FeCl2·7H2O) solution buffered at pH 5.5 (15 mM 2-(N-morpholino)ethanesulfonic acid). Fe plaque induction was carried out in deionized water to avoid any possible formation of Fe precipitates with other elements present in the nutrient solution (Zhang et al. 1999; Liang et al. 2006). Fe plaque on the root surfaces was allowed to form for 48 h. The plants were then transferred back to their respective P-deficient or P-sufficient nutrient solutions, as described above (i.e., +P + Fe and ‒P + Fe treatments). Plants not exposed to Fe plaque induction (i.e. +P ‒Fe and ‒P ‒Fe treatments) were treated in the same way, by replacing the ferrous solution with deionized water. To limit the oxidation of Fe(II) by external sources of O2 other than ROL during Fe plaque induction, the Fe(II) solution was previously flushed with N2 and, after plant root insertion, the containers were wrapped in foil and their mouths sealed with parafilm around the shoots to limit O2 diffusion. Separate containers with the Fe(II) solution but without plants were also included to account for any possible oxidation of Fe(II) not induced by ROL. The Fe(II) concentration in all the treatment was constantly monitored during the 48 h induction, more detailed information are reported in "Rate of Fe plaque formation and net proton release" section. A graphical summary of the experimental set-up and of the measurements conducted is provided in supplementary material (Fig. S1).
Plant analyses
Plant samples were collected (four replicates for each treatment) at four different time points throughout the experiment. They were first sampled at 30 DAS in correspondence with the first Fe plaque induction period, then before (45 DAS) and after (48 DAS) the second Fe plaque induction period. The final plants were collected at 62 DAS. Plants that were sampled at 30 DAS were used to evaluate the impact of P availability on root development and morphology. In addition to determining root biomass, three plants from each treatment were imaged using a Nikon D3500 digital camera. Root traits including total root length (cm), surface area (cm2), total root volume (cm3), and number of root tips were quantified using saRIA software (Narisetti et al. 2019). The root density (mg cm−3) was estimated as the ratio between the root biomass and the root volume. The development of root aerenchyma was assessed qualitatively on three roots per plant after sectioning 3 cm from the root tip and examined under a light microscope (Leitz Dialux 20 EB) with a 24 MP digital camera (Nikon D3500). At 62 DAS, plants were collected to evaluate the final dry biomass (shoot + root) and P contents in all plant parts. The P concentration in root and shoot tissues was determined on dry plant material (50 mg) after sulfuric-perchloric digestion and colorimetric quantification of P in the extracts using the malachite green method (Ohno and Zibilske 1991). To evaluate the changes in Fe plaque contents during the growth period as a function of P availability, all root samples collected before and/or after Fe plaque induction were analysed for root-associated Fe. For each plant, the entire root system was first rinsed with deionized water and subsequently extracted with 0.1 M acidic ammonium oxalate (Loeppert and Inskeep 1996; Hossain et al. 2009). Fe concentration in the extracts was quantified by atomic absorption spectroscopy (AAS, PerkinElmer AAnalyst 400, Norwalk, CT, USA).
Rate of Fe plaque formation and net proton release
The rate of Fe(II) oxidation for the P-sufficient + Fe and P-deficient + Fe treatments during the second induction period (45 DAS) was determined on four independent replicates for each treatment. During the 48 h-induction period, small aliquots of the ferrous Fe solution were sampled every hour and the residual dissolved Fe(II) concentrations were quantified spectrophotometrically, immediately after sampling using the 1,10-phenanthroline method (Loeppert and Inskeep 1996). The decrease in the concentration of Fe(II) in the solution over time was related to the kinetics of Fe(II) oxidation that served as a proxy for the rate of Fe plaque formation by ROL, after accounting for the minimal Fe(II) oxidation in the absence of plants and assuming that plant uptake of dissolved Fe over this short period was negligible. Moreover, it was ensured that the precipitation of Fe minerals occurred only on the root surfaces as the solution remained clear and no precipitation of Fe (oxyhydr)oxides was observed on the container walls during the Fe plaque induction period. Under these experimental conditions, the rate of Fe(II) oxidation by O2 continuously released from the roots and at constant pH was described by a pseudo-first order rate expression with respect to Fe(II) according to the equation
where [Fe(II)]t and [Fe(II)]0 where the concentrations of Fe(II) at time t and at the beginning of the induction period, respectively, while k was the pseudo-first order rate constant (in h−1). Since we expected P availability to influence other root traits (including root surface area) in addition to ROL, the cumulative amount of Fe(III) precipitated on the root surfaces (in mmol Fe) over the first hours of Fe plaque induction, calculated as the difference between [Fe(II)]0 and [Fe(II)]t was normalised on the basis of root surface area (mmol Fe m−2) and plotted against time, as proposed by Kirk and Van Du (1997). Similarly, rate constants were also normalised for root surface area for comparison between treatments.
To quantify the capacity of rice roots to exude protons as a function of the different treatments, the pH of the growth solutions was monitored every 24 h in the 3 days preceding and succeeding each Fe induction event, by means of a pH-sensitive electrode (inoLab pH 7110, WTW GmbH, Weilheim, Germany). The net H+ release (mM) over time was calculated from the decrease in pH with respect to the initial pH of the growth solution.
Statistical analyses
The statistical analyses were performed using the R version 4.1.1. Normality and data homoscedasticity were checked with Shapiro-Wilk and Levene tests, respectively. When necessary, data were transformed according to the data distribution. All the variables were tested for the analysis of variance (two-way ANOVA), followed by pair-wise post hoc analyses (Student-Newman-Keuls test) to determine the difference among the mean value at p < 0.05. The ggplot2 package was used to plot all the figures.
Results
Plant growth, root traits and elemental composition
The root dry biomass measured at 30 DAS (Table 1) did not show significant differences between P-deficient and P-sufficient plants. However, the root traits were strongly influenced by P treatment (p < 0.001), with P-deficient plants showing a 2 to 3-fold higher root volume, surface area, total length, and number of root tips compared to P-sufficient plants (Table 1). The root density was lower in the P-deficient plants, which is consistent with the higher development of aerenchyma as evidenced through the microscopic observation of the root sections (Fig. 1). Similarly, at the end of the growing period (62 DAS), the differences observed in the total dry biomass, were mainly attributed to differences in shoot biomass, as the root biomass was unaffected by both P and Fe treatments (Table 2). The shoot biomass at 62 DAS was highly dependent on P treatment (p < 0.01), Fe treatment (p < 0.01) and their interaction (p < 0.05; Table 2). Although P-sufficient plants showed a higher shoot biomass than P-deficient plants in the absence of Fe plaque, shoot growth in the former was strongly hindered when Fe plaque (P-sufficient + Fe) was present, compared to plants grown without Fe plaque (P-sufficient–Fe). On the other hand, no significant differences were observed in P-depleted plants with or without Fe plaque (P-deficient + Fe vs. P-deficient –Fe; Table 2). These effects on plant growth resulted in significant differences on the root-to-shoot ratio (R/S), based on P treatment (p < 0.05), Fe treatment (p < 0.05) and their interaction (p < 0.05; Table 2). Particularly, P-sufficient plants showed a lower R/S ratio compared to P-deficient plants, and in the latter a decrease R/S in absence of Fe plaque was observed. Conversely, P-sufficient plants did not result in significant alteration of R/S when Fe plaque was induced, but the average R/S ratio was close to P-deficient plants with Fe plaque (Table 2).

Root sections of representative rice plants grown for 30 days in P-sufficient (a) and P-deficient (b) nutrient solutions. Scale bar is 0.1 mm
The root P concentration and total root P content were higher in P-sufficient compared to P-deficient plants (p < 0.05), whereas the presence or absence of Fe plaque did not cause any significant differences in root P within the P treatments (Fig. 2). Conversely, both shoot P concentration and content showed a significant dependence on the P treatment (p < 0.001 and p < 0.01, respectively), Fe treatment (p < 0.01 and p < 0.001, respectively), and their interaction (p < 0.05 and p < 0.001, respectively). In all cases, P-sufficient plants accumulated more P in the shoot than P-deficient plants (Fig. 2). However, whereas under P-sufficient conditions the presence of Fe plaque on the roots strongly limited P uptake in comparison to plants without Fe plaque, the opposite was evident for plants grown under P-deficiency (Fig. 2). Consistently with shoot P accumulation, the difference in total P uptake between plants grown under P-sufficient and P-deficient conditions was much more evident in the absence rather than in the presence of Fe plaque (Fig. 2; p < 0.001).

P concentrations (a) and contents (b) in the roots and shoots of rice plants grown under P-sufficient (+ P) and P-deficient (‒P) conditions, in the presence (+ Fe) or absence (–Fe) of Fe plaque at 62 DAS. Each value represents the mean of four replicates (± SE). Lowercase letters above bars indicate significant differences between Fe treatment within the same P treatment (p < 0.01), uppercase letters above bars indicate significant differences between P treatment within the same Fe treatment (p < 0.01). Uppercase letters with brackets represent significant differences between P treatments when no interaction with Fe treatment was observed (p < 0.05)
Fe plaque formation and dissolution
The rate of Fe(II) oxidation during Fe plaque induction followed pseudo-first order kinetics with respect to Fe(II). Notably, the rate of Fe(II) oxidation was considerably faster in the presence of plants roots grown under P-deficient conditions and the rate constant (k) was 8 times higher than that observed in the presence of P-sufficient plants (Fig. 3a; Table 3). Although P-deficient plants had a significantly larger total root surface area (Table 1), they still displayed a faster rate of Fe plaque formation when the cumulative amount of Fe(III) precipitated on the root surface was normalised to root surface area (Fig. 3b). Similarly, surface area-normalised rate constants were 2.5 times faster for P-deficient with respect to P-sufficient plants (Table 3). Although initially P-sufficient plants showed higher amounts of precipitated Fe per unit root surface area compared to P-deficient plants, the differences were merely transient and soon reached similar values within 5 h of Fe plaque induction (Fig. 3b).

Fe(II) oxidation kinetics during the 48 h Fe plaque induction period approximated by a pseudo-first order reaction for plants grown for 45 d under P-sufficient (+ P) and P-deficient (‒P) conditions, as well as for the control without plants (a), and the amount of precipitated Fe(III) with time over the first 10 h of Fe plaque induction normalized per unit root surface area (b). Values represent the mean of measurements on four independent replicates per treatment. For the sake of graph readability, SEs are not reported
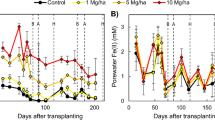
The amount of Fe plaque formed after the first Fe plaque induction period (on 30 DAS) was slightly higher for P-deficient than P-sufficient plants (Fig. 4). However, by 45 DAS most of the Fe plaque was redissolved for both treatments, even though P-deficient plants still showed significantly lower root-associated Fe concentrations than P-sufficient plants. After the second Fe plaque induction period (on 48 DAS), the quantity of root-associated Fe was significantly higher in P-deficient compared to P-sufficient plants (in line with the faster rate of Fe(II) oxidation). Over the following 15 d, the amount of Fe plaque on P-sufficient plants remained relatively unchanged, while in P-deficient plants, a substantial dissolution of Fe plaque occurred, resulting in no significant differences in Fe plaque contents between the two P treatments by 62 DAS. In all cases, no root-associated Fe plaque was detected in control roots of both P-deficient –Fe and P-sufficient –Fe plants.

Changes in the Fe plaque content over time in P-sufficient (+ P) and P-deficient (‒P) plants over the growth period (Root analysis carried out on 30, 45, 48 and 62 DAS). Yellow arrows in the top bar of the figure indicate the timing of Fe plaque induction. Oxalate-extractable Fe in –Fe control plants were under the detection limits and not included. Values represents the mean of four replicates (± SE). Different letters above bars indicate significant differences between treatment (p < 0.05)
Net proton release
The net proton release by rice plants over time differed as a function of P availability, presence of Fe plaque as well as the plant growth stage (Fig. 5). In the 72 h before the first Fe plaque induction period, the net proton release was only influenced by the P treatment (p < 0.001), as no Fe treatment was yet applied. In particular, P-deficient plants showed significantly higher rates of H+ exudation than P-sufficient plants throughout the monitoring period (Fig. 5a). After the first Fe plaque induction period, the kinetics of net H+ release was influenced by the combined effects of both P and Fe treatments (p < 0.001). In the absence of Fe plaque, P-deficient plants continued to exhibit higher values of H+ exudation than P-sufficient plants, with rates very similar to those observed earlier. On the other hand, net H+ release in solution in the presence of Fe plaque on the root surface, which was only observed after 48 h from induction, was not significantly different between P-sufficient and P-deficient plants. Moreover, the rates of H+ release were much lower compared to those observed in the absence of Fe plaque, regardless of P availability (Fig. 5b). During the 72 h before the second Fe plaque induction, H+ exudation kinetics in the absence of Fe plaque followed similar trends to those described previously, with significantly higher values observed for the P-deficient treatment in comparison to the P-sufficient treatment (Fig. 5c). However, in the presence of Fe plaque, the net release of H+ by the roots was significantly higher in the P-sufficient treatment. Similar results were observed immediately after the second Fe plaque induction period, with P-deficient plants showing the highest values of net H+ release in the absence of Fe plaque and the lowest values in the presence of Fe plaque (Fig. 5d). During this period, the kinetics of H+ release appeared to be faster compared to the 72 h succeeding the first Fe plaque induction (Fig. 5b).

Kinetics of net proton exudation over 72 h before (a) and after (b) the first Fe plaque induction (30 DAS), and before (c) and after (d) the second Fe plaque induction (45 DAS) in P-sufficient (+ P) and P-deficient (‒P) plants, with (+ Fe) or without Fe (–Fe) plaque induction. Values represent the mean of measurements on four independent replicates per treatment. For the sake of graph readability, SEs are not reported, while different letters indicate significant differences between treatments for each analysis time (p ≤ 0.001). Note the different range of values on the y-axis between the two induction periods
Discussion
Influence of P availability on rice root traits and Fe plaque dynamics
As expected, P-deficiency induced rice plants to modify their root morphology and traits. Specifically, P-limited plants significantly enhanced their root length, volume, surface area, and tips number compared to plants grown under P-sufficient conditions, without affecting root dry weight. This response was accompanied by a greater development of aerenchyma as previously reported by Fu et al. (2014). Aerenchyma develops in rice plants to facilitate the diffusion of O2 from the shoot to the submerged roots and, under P limitation, the development of these tissues is further promoted to sustain the higher O2 requirements associated with a greater soil exploration by the roots, without increasing the C cost of a higher root biomass (Lynch and Brown 2008). Increased O2 transport to the root is crucial to support energy-dependent processes related to plant growth and nutrition under P limitation and indicates a relationship between P availability and O2 dynamics in the root environment.
Due to enhanced ROL, in the presence of sufficient amounts of dissolved Fe(II) as those typically found in the porewaters of anaerobic rice paddy soils, P-deficient plants showed a faster precipitation of Fe(III) (oxyhydr)oxides on their root surfaces, resulting in significantly higher amounts of Fe plaque immediately following induction, compared to P-sufficient plants. This finding supports our initial hypothesis that changes in rice root traits in response to low P availability lead to an increase in Fe plaque formation on the root surface. Despite similar results being reported in other studies, the underlying factors that drive the increase in Fe plaque formation at low P supply remain controversial. In fact, previous research conducted by Kirk and Van Du (1997) investigated the changes in root architecture, porosity and O2 release in 30 d-old rice plants grown under P concentrations similar to those used in our experiment, and observed a higher apparent rate of O2 release in P-limited compared to P-sufficient plants. Such an effect was entirely attributed to an increased root volume rather than to higher rates of O2 release per unit of root volume. On the other hand, in line with our findings, Fu et al. (2014) observed an increase in oxygenation capacity of rice roots under P-deficient conditions, which was associated to enhanced root aerenchyma development. However, it is worth noting that these authors did not evaluate the consequences of this oxygenation on Fe plaque formation. Also, Wu et al. (2012) did not find a significant relationship between root volume and the degree of Fe plaque formation under similar growing conditions, but their study revealed a positive correlation between Fe plaque formation and the ROL when comparing different rice genotypes. Our results indicate that the higher rate of Fe(II) oxidation and Fe plaque formation under P-deficient conditions was not solely attributable to a larger total root surface area but, considering the values of root surface area-normalized rate constants, also to higher ROL. Although we did not measure the rates of root O2 release directly, our study strongly supports the influence of ROL on the observed phenomena.
Despite the significantly higher amount of Fe precipitated on the root surfaces of P-deficient plants following the Fe plaque induction periods, the subsequent decrease in root-associated Fe over the entire growth period clearly indicated the dynamic nature of this Fe pool, and presumably any associated P. We speculate that the amount of Fe plaque present on the roots is likely determined by the interplay between the rates of formation and dissolution, both of which are notably influenced by P availability. In support of this, albeit the higher rates of Fe mineral precipitation on the root surfaces, P-deficient plants are capable of up-regulating strategies for the dissolution of Fe plaque. This adaptive response enables them to release the required P associated with the Fe plaque to cope with the limited P availability. Indeed, it is well known that P-deficient plants can acidify the rhizosphere via proton exudation to promote the dissolution of sparingly available P forms, as well as the release of P adsorbed onto Fe oxides (Kirk and Van Du 1997; Hinsinger 2001; Santoro et al. 2022). Given that Fe plaque is composed of Fe (oxy)hydroxide phases similar to those found in soil (Hansel et al. 2001; Seyfferth et al. 2010; Amaral et al. 2017), we expect similar mechanisms to be more effective at the root surface. This observation would explain the faster decline of root-associated Fe over time in P-deficient compared to P-sufficient plants following Fe plaque induction, albeit differences in root elongation and architecture as a function of P availability could have also been partially responsible for the observed changes in the amount of root-associated Fe with time. Our second hypothesis is nevertheless supported by the higher net H+ release in P-deficient compared to P-sufficient plants in the absence of Fe plaque, and the significant consumption of these protons during Fe plaque dissolution in the former when Fe plaque was present on the root surfaces (Fig. 5d). In fact, the dissolution of Fe oxide phases always involves a stoichiometric consumption of protons that explains the lower net H+ exudation in P-deficient with respect to P-sufficient plants in the presence of Fe plaque (Schwertmann 1991). Therefore, assuming that root H+ exudation was only influenced by P availability but not by the presence or absence of Fe on the root surface, the lowest values of net H+ release in solution under P-deficient conditions suggest that most of the protons released were used for Fe plaque dissolution. In contrast, the lack of significant differences in net H+ release between + Fe and –Fe treatments under P-sufficient conditions suggest that Fe plaque dissolution mechanisms were not activated when P supply was sufficient. This finding is further confirmed by the smaller change in Fe plaque contents over time following each induction.
The role of Fe plaque in regulating P uptake by rice plants
The importance of Fe plaque dynamics over time has been recently related to their role in regulating nutrient and/or contaminants availability for rice (Li et al. 2015; Zhou et al. 2018; Limmer et al. 2022). Nonetheless the majority of the previous hydroponic studies generally focused on a very short time frame (Zhang et al. 1999; Liang et al. 2006; Liu et al. 2019) after a single Fe plaque induction period (Jiaofeng et al. 2022), thus failing to consider the effect of the temporal transformations in Fe plaque in relation to plant development. In this study we finally hypothesized that the different findings reported in literature regarding the role of Fe plaque in serving either as a sink or a source of P for rice acquisition, mainly arise from a lack of consideration of the interdependence and temporal variation of various rhizosphere processes that regulate P availability throughout the rice growth period. For example, the studies by Zhang et al. (1999), Liang et al. (2006), and Jiaofeng et al. (2022) related a higher Fe plaque formation to an inhibitory effect on plant P uptake, however the tested range of P concentrations (> 150 µM) were higher than the threshold (6 µM) above which P availability is considered to be sufficient for rice plants (Shahandeh et al. 1995; Frossard et al. 2000). With these experimental designs they were thus not able to appreciate the plant adaptation mechanisms (e.g. rhizosphere acidification) on Fe plaque dissolution and P uptake under low P availability.
The results of our study suggest that the equilibrium between Fe plaque formation and dissolution depends on the availability of P in relation to plant requirements. Whereas the presence of Fe plaque resulted in a limitation of plant P uptake under P-sufficient conditions over a growth period of 60 d, a higher P uptake was observed in the presence rather than in the absence of Fe plaque under P-deficient conditions. By combining these findings with the enhanced Fe plaque dissolution observed at low P availability, we provide support for our third hypothesis. Indeed, Fe (oxyhydr)oxides on the root surface can strongly bind P (Zhang et al. 1999; Liang et al. 2006; Khan et al. 2016), and their dissolution facilitated by rhizosphere acidification might have promoted the release and uptake of the bound P at rhizosphere level. Our results therefore confirm that Fe plaque can act as a barrier to plant P uptake under high P availability but serve as a P source under low P availability partially attenuating the nutrient deficiency.
Undoubtedly, the current study has only investigated plant responses on rhizospheric Fe and P interactions and did not evaluate the influence of microbially-driven processes that are known to play an important role in Fe redox cycling the rice rhizosphere. Furthermore, under field conditions it is likely during the oxidative precipitation of Fe minerals on the root surface, P retention does not only involve adsorption but also co-precipitation that, depending of the P:Fe ratio and P forms may involve adsorption, precipitation and physical entrapment within the newly formed particles (Voegelin et al. 2013; Santoro et al. 2019). These mechanisms together could contribute to changes in the mineralogy of Fe plaque that are known to occur during the rice growing period (Maisch et al. 2019; Limmer et al. 2022), possibly affecting the rhizospheric P cycling and its availability for plant uptake. However, the current observations extend our understanding of the role that root traits play in driving Fe plaque formation and dissolution as a function of nutrient availability, considering a range of P concentrations in the nutrient solution very close to those present in paddy soil solutions, thereby providing mechanistic explanations for the interactions between P availability and Fe plaque dynamics.
Conclusions
Due to the high affinity of Fe (oxyhydr)oxides for P, Fe plaque generally associated with rice roots under anoxic soil conditions, can influence P uptake for sustaining growth. In this study we show that both Fe plaque formation and dissolution during the early stages of plant growth, are influenced by root traits that are themselves regulated by P availability. Whereas P-deficiency leads to modifications in root morphology and traits that induce a faster rate and higher amount of Fe plaque formation, the dissolution of Fe plaque through mechanisms induced by P starvation (i.e. rhizosphere acidification) is also enhanced, such that plaque-associated P can potentially serve as a nutrient source facilitating P uptake under limiting conditions. On the other hand, under P-sufficient conditions Fe plaque may serve as a P sink limiting plant uptake and growth to some extent, as plaque formation is not accompanied by its dissolution. This study therefore contributes to further understanding the complex interactions between modifications in rice root traits and temporal Fe plaque dynamics on root surfaces as a function of P availability, which deeply shape nutrient cycling in the rhizosphere, albeit further investigations should also consider the active role of microbial communities associated with the roots in rhizospheric Fe cycling in paddy soils, which together with plant responses, may also play an important role in controlling P availability to rice plants.
Abbreviations
- +P + Fe:
-
Phosphorus sufficient plants with iron plaque
- +P –Fe:
-
Phosphorus sufficient plants without iron plaque
- –P + Fe:
-
Phosphorus limited plants with iron plaque
- –P –Fe:
-
phosphorus
limited plants without iron plaque
References
Amaral DC, Lopes G, Guilherme LRG, Seyfferth AL (2017) A new approach to sampling intact fe plaque reveals si-induced changes in fe mineral composition and shoot as in rice. Environ Sci Technol 51(1):38–45. https://doi.org/10.1021/acs.est.6b03558
Armstrong W (1967) The oxidising activity of roots in waterlogged soils. Physiol Plant 20(4):920–926. https://doi.org/10.1111/j.1399-3054.1967.tb08379.x
Bhattacharyya P, Das S, Adhya TK (2013) Root exudates of rice cultivars affect rhizospheric phosphorus dynamics in soils with different phosphorus statuses. Commun Soil Sci Plant Anal 44(10):1643–1658. https://doi.org/10.1080/00103624.2013.769562
Colmer TD (2003) Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots: gas transport in plants. Plant Cell Environ 26(1):17–36. https://doi.org/10.1046/j.1365-3040.2003.00846.x
Evans DE (2004) Aerenchyma formation. New Phytol 161(1):35–49. https://doi.org/10.1046/j.1469-8137.2003.00907.x
Frossard E, Condron LM, Oberson A, Sinaj S, Fardeau JC (2000) Processes governing phosphorus availability in temperate soils. J Environ Qual 29(1):15–23. https://doi.org/10.2134/jeq2000.00472425002900010003x
Fu YQ, Yang XJ, Shen H (2014) The physiological mechanism of enhanced oxidizing capacity of rice (Oryza sativa L.) roots induced by phosphorus deficiency. Acta Physiol Plant 36(1):179–190. https://doi.org/10.1007/s11738-013-1398-3
Hansel CM, Fendorf S, Sutton S, Newville M (2001) Characterization of Fe plaque and associated metals on the roots of mine-waste impacted aquatic plants. Environ Sci Technol 35(19):3863–3868. https://doi.org/10.1021/es0105459
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237(2):173–195. https://doi.org/10.1023/A:1013351617532
Hoffland E, Wei C, Wissuwa M (2006) Organic Anion Exudation by Lowland Rice (Oryza sativa L.) at Zinc and Phosphorus Deficiency. Plant Soil 283(1–2):155–162. https://doi.org/10.1007/s11104-005-3937-1
Hossain MB, Jahiruddin M, Loeppert RH, Panaullah GM, Islam MR, Duxbury JM (2009) The effects of iron plaque and phosphorus on yield and arsenic accumulation in rice. Plant Soil 317(1–2):167–176. https://doi.org/10.1007/s11104-008-9798-7
Jiaofeng G, Yang H, Peng Z, Bohan L, Hang Z (2022) Increasing phosphorus inhibits the retention and prevention of cadmium by iron plaque and promotes cadmium accumulation in rice plants. Chemosphere 307:135642. https://doi.org/10.1016/j.chemosphere.2022.135642
Khan N, Seshadri B, Bolan N, Saint CP (2016) Root iron plaque on wetland plants as a dynamic pool of nutrients and contaminants. Adv Agron 138:1–96. https://doi.org/10.1016/bs.agron.2016.04.002
Kirk G (2004) The biogeochemistry of submerged soils, 1st edn. Wiley
Kirk GJD, Van Du L (1997) Changes in rice root architecture, porosity, and oxygen and proton release under phosphorus deficiency. New Phytol 135(2):191–200. https://doi.org/10.1046/j.1469-8137.1997.00640.x
Li R, Zhou Z, Zhang Y, Xie X, Li Y, Shen X (2015) Uptake and accumulation characteristics of arsenic and iron plaque in rice at different growth stages. Commun Soil Sci Plant Anal 46(19):2509–2522. https://doi.org/10.1080/00103624.2015.1089259
Liang Y, Zhu Y-G, Xia Y, Li Z, Ma Y (2006) Iron plaque enhances phosphorus uptake by rice (Oryza sativa) growing under varying phosphorus and iron concentrations. Ann Appl Biol 149(3):305–312. https://doi.org/10.1111/j.1744-7348.2006.00095.x
Limmer MA, Thomas J, Seyfferth AL (2022) The effect of silicon on the kinetics of rice root iron plaque formation. Plant Soil 477(1–2):171–181. https://doi.org/10.1007/s11104-022-05414-4
Liu J, Sui P, Cade-Menun BJ, Hu Y, Yang J, Huang S, Ma Y (2019) Molecular-level understanding of phosphorus transformation with long-term phosphorus addition and depletion in an alkaline soil. Geoderma 353:116–124. https://doi.org/10.1016/j.geoderma.2019.06.024
Loeppert RH (1996) Iron. In: Bigham JM (ed) Part 3 methods of soil analysis. Chemical Methods. SSSA, Madison, pp 639–664
Lynch JP, Brown KM (2008) Root strategies for phosphorus acquisition. In: White PJ, Hammond JP (eds) The ecophysiology of plant-phosphorus interactions. Springer Netherlands, Dordrecht, pp 83–116
Maisch M, Lueder U, Kappler A, Schmidt C (2019) Iron lung: how rice roots induce iron redox changes in the rhizosphere and create niches for microaerophilic Fe(II)-oxidizing bacteria. Environ Sci Technol Lett 6:600–605. https://doi.org/10.1021/acs.estlett.9b00403
Marschner P (2021) Processes in submerged soils – linking redox potential, soil organic matter turnover and plants to nutrient cycling. Plant Soil 464(1–2):1–12. https://doi.org/10.1007/s11104-021-05040-6
Martinengo S, Schiavon M, Santoro V, Said-Pullicino D, Romani M, Miniotti EF, Celi L, Martin M (2023) Assessing phosphorus availability in paddy soils: the importance of integrating soil tests and plant responses. Biol Fertil Soils 59(4):391–405. https://doi.org/10.1007/s00374-023-01714-8
Mori A, Fukuda T, Vejchasarn P, Nestler J, Pariasca-Tanaka J, Wissuwa M (2016) The role of root size versus root efficiency in phosphorus acquisition in rice. EXBOTJ 67(4):1179–1189. https://doi.org/10.1093/jxb/erv557
Narisetti N, Henke M, Seiler C, Shi R, Junker A, Altmann T, Gladilin E (2019) Semi-automated root image analysis (saRIA). Sci Rep 9(1):19674. https://doi.org/10.1038/s41598-019-55876-3
Ohno T, Zibilske LM (1991) Determination of low concentrations of phosphorus in soil extracts using malachite green. Soil Sci Soc Am J 55(3):892–895. https://doi.org/10.2136/sssaj1991.03615995005500030046x
Ponnamperuma FN (1972) The chemistry of submerged soils. Adv Agron 24:29–96
Richardson AE, Hocking PJ, Simpson RJ, George TS (2009) Plant mechanisms to optimise access to soil phosphorus. Crop Pasture Sci 60(2):124. https://doi.org/10.1071/CP07125
Rose TJ, Impa SM, Rose MT, Pariasca-Tanaka J, Mori A, Heuer S, Johnson-Beebout SE, Wissuwa M (2013) Enhancing phosphorus and zinc acquisition efficiency in rice: a critical review of root traits and their potential utility in rice breeding. Ann Botany 112(2):331–345. https://doi.org/10.1093/aob/mcs217
Sahrawat KL (2005) Iron toxicity in wetland rice and the role of other nutrients. J Plant Nutr 27(8):1471–1504. https://doi.org/10.1081/PLN-200025869
Santoro V, Martin M, Persson P, Lerda C, Said-Pullicino D, Magnacca G, Celi L (2019) Inorganic and organic P retention by coprecipitation during ferrous iron oxidation. Geoderma 348:168–180. https://doi.org/10.1016/j.geoderma.2019.04.004
Santoro V, Schiavon M, Visentin I, Martin M, Said-Pullicino D, Cardinale F, Celi L (2022) Tomato plant responses induced by sparingly available inorganic and organic phosphorus forms are modulated by strigolactones. Plant Soil 474(1–2):355–372. https://doi.org/10.1007/s11104-022-05337-0
Schwertmann U (1991) Solubility and dissolution of iron oxides. Plant Soil 130(1–2):1–25. https://doi.org/10.1007/BF00011851
Seck PA, Diagne A, Mohanty S, Wopereis MCS (2012) Crops that feed the world 7: Rice. Food Sec 4(1):7–24. https://doi.org/10.1007/s12571-012-0168-1
Seyfferth AL, Webb SM, Andrews JC, Fendorf S (2010) Arsenic localization, speciation, and co-occurrence with iron on rice (Oryza sativa L.) roots having variable Fe coatings. Environ Sci Technol 44(21):8108–8113. https://doi.org/10.1021/es101139z
Shahandeh H, Hossner LR, Turner FT (1995) Evaluation of soil phosphorus tests for flooded rice soils under oxidized and reduced soil conditions. Commun Soil Sci Plant Anal 26(1–2):107–121. https://doi.org/10.1080/00103629509369284
Veelen A, Koebernick N, Scotson CS, McKay-Fletcher D, Huthwelker T, Borca CN, Mosselmans JFW, Roose T (2020) Root‐induced soil deformation influences Fe, S and P: rhizosphere chemistry investigated using synchrotron XRF and XANES. New Phytol 225(4):1476–1490. https://doi.org/10.1111/nph.16242
Voegelin A, Senn A-C, Kaegi R, Hug SJ, Mangold S (2013) Dynamic Fe-precipitate formation induced by Fe(II) oxidation in aerated phosphate-containing water. Geochim Cosmochim Acta 117:216–231. https://doi.org/10.1016/j.gca.2013.04.022
Wu C, Ye Z, Li H, Wu S, Deng D, Zhu Y, Wong M (2012) Do radial oxygen loss and external aeration affect iron plaque formation and arsenic accumulation and speciation in rice? J Exp Bot 63(8):2961–2970. https://doi.org/10.1093/jxb/ers017
Yoshida D (1967) Effects of nutrient deficiencies on tne biosynthesis of nicotine in tobacco plants. Soil Sci Plant Nutr 13(4):107–111. https://doi.org/10.1080/00380768.1967.10431983
Yoshida S, Forno DA, Cock JH, Gomez K (1972) Laboratory manual for physiological studies of rice, 2nd edn. The International Rice Research Institute, Los Baños, Philippines, pp 53–57
Zhang Y, Lin X, Werner W (2003) The effect of soil flooding on the transformation of Fe oxides and the adsorption/desorption behavior of phosphate. Z Pflanzenernähr Bodenk 166(1):68–75. https://doi.org/10.1002/jpln.200390014
Zhang Y, Lin X, Werner W (2004) Effects of aerobic conditions in the rhizosphere of rice on the dynamics and availability of phosphorus in a flooded soil – a model experiment. Z Pflanzenernähr Bodenk 167(1):66–71. https://doi.org/10.1002/jpln.200320349
Zhang X, Zhang F, Mao D (1999) Effect of iron plaque outside roots on nutrient uptake by rice (Oryza sativa L.): phosphorus uptake. Plant Soil 209(2):187–192. https://doi.org/10.1023/A:1004505431879
Zhou H, Zhu W, Yang W-T, Gu J-F, Gao Z-X, Chen L-W, Du W-Q, Zhang P, Peng P-Q, Liao B-H (2018) Cadmium uptake, accumulation, and remobilization in iron plaque and rice tissues at different growth stages. Ecotoxicol Environ Saf 152:91–97. https://doi.org/10.1016/j.ecoenv.2018.01.031
Acknowledgements
The authors would like to show their gratitude to Prof. L. Tavella and Dr. F. Tortorici for access and assistance with the use of laboratory equipment for the qualitative assessment of root aerenchyma.
Funding
Open access funding provided by Università degli Studi di Torino within the CRUI-CARE Agreement. This study was financially supported by the Italian Ministry of University and Research (MUR) through the PRIN project “Roots in armour: Formation and functions of iron plaques at the root-soil interface of wetland plants” (ROOTARMOUR; Project Nr. 2022T3LMSS). SM was supported by the Cassa di Risparmio di Torino through the research project “Enhanced biological N fixation through improved P acquisition by leguminous plants” (FOS4FIX).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Conceptualization: Sara Martinengo, Daniel Said-Pullicino. Methodology: Sara Martinengo, Daniel Said-Pullicino. Formal analysis and investigation: Sara Martinengo, Veronica Santoro. Writing—original draft preparation: Sara Martinengo, Veronica Santoro. Writing—review and editing: all authors. Funding acquisition: Daniel Said-Pullicino. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Additional information
Responsible Editor: Terry James Rose.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 660 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Martinengo, S., Santoro, V., Schiavon, M. et al. The influence of phosphorus availability on rice root traits driving iron plaque formation and dissolution, and implications for phosphorus uptake. Plant Soil 494, 603–616 (2024). https://doi.org/10.1007/s11104-023-06306-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-023-06306-x