
Abstract
The split-root system technique has been used to analyse plant biology for several decades, but woody plants have not received enough attention in this experimental approach. Historically, several methods have been developed, ranging from the simplest method of dividing a root into two parts and placing them in separate containers to more complex methods, such as grafting a second root from another plant. Each method has advantages and disadvantages that determine the goals of the experiment. Thus far, research using the split-root system has covered only 62 species of woody plants, mainly to investigate the water shortage effect on water acquisition. Many studies have also considered the significance of functional root-system diversity for plant fertilisation, which allows a better understanding of ion transport regulation mechanisms and some anatomical and functional features of woody plants. Ion uptake and transport have been studied frequently using isotope labelling. The split-root system method also offers interesting possibilities for studying the interactions of plants with other organisms. For example, this method was used to study root colonization strategies by mycorrhizal fungi. The comprehensive analysis of the split-root system technique in this review provides fine-scale information on the future concepts needed to study root-system biology, as the ability of roots to play a range of functions in the plant remains largely untested.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Plants, especially woody ones, are constantly exposed to changes in environmental conditions. The changing temperature, insolation, humidity, herbivore pressure, and competition within and between species affect the above-ground parts and the interrelationships at the root-soil contact. Thus, the influence of various biotic and abiotic factors on the root structure and metabolism has been intensively studied, and knowledge in this regard has increased. However, the root systems of trees and how they function remain a scientific mystery in many aspects.
The soil environment, in compere above ground, is characterised by a smaller amplitude of changes, especially physical factors, but is more heterogeneous, especially during the life of one tree plant (Hodge 2006). A technique enabling a detailed mechanistic and functional root-system survey (i.e. the split-root system or SRS) is employed to analyse the effect of a heterogeneous soil environment on plant growth under controlled conditions. This technique has been known for a long time; the first general studies were published in the 1940s (Long 1943) and, in the case of woody plants, in the 1960s (Fucik and Titus 1965). Whether dealing with herbaceous or woody plants, the SRS technique splits the root into two (or more) nontouching parts planted in separate containers. There are several ways to establish the SRS, which for herbaceous plants on the example of Arabidopsis thaliana (Brassicaceae) have been discussed in more detail in the review Saiz-Fernández et al. (2021). The roots of woody plants live longer and are larger than herbaceous plants, so they benefit from a more heterogeneous soil environment and they have to be more plastic. The vascular cambium originates in two different tissues (pericycle and procambium), and not only in the pericycle as in herbaceous plants. The roots of woody plants have a rich set of mycorrhizal fungi, which changes with the age of the tree (Dighton and Mason 1985). Therefore, even though the methods described for woody plants are similar to those for herbaceous plants, due to the different structure of the root, they have their own specificity.
The application of SRS technique is used in fundamental studies of plant morphology and physiology: plant signalling (Torres et al. 2021), sectoriality defined as the restriction of resource and hormone movement to limited regions of the plant, each of which comprises a quasi-autonomous “independent physiological unit” (Marquis 1996; Orians et al. 2004), water uptake and transport (Ameglio et al. 1999), ion transport (Gloser et al. 2009b) or ectomycorrhizal symbioses (Rose et al. 2023). The SRS in woody plants was most often used to study the effects of drought. Some studies aimed at optimizing water consumption were done on economically important species: vine grapes (Antolin et al. 2006, 2008; Dry and Loveys 1999; Dry et al. 2000; Poni et al. 2007, 2009), apples (Einhorn et al. 2009, 2012; Gowing et al. 1990; Zhao et al. 2008) or citrus (Allen et al. 2000; Contreras-Barragan et al. 2014; Miranda et al. 2018; Romero-Conde et al. 2014). The SRS method is also sometimes used in ecological research (e.g. plant interspecies competition (Marler 2013) and plant interaction with pathogens or mycorrhizal fungi (Kennedy et al. 2009)). However, these studies are still incidental, and their more common applications may introduce new research directions. This review aims to discuss the previous research results and mark research areas in which this technique may enhance the knowledge of woody plant anatomy, physiology and ecology. It presents various techniques for achieving SRS, giving the advantages and disadvantages of each method, and gives examples of SRS applications in the study of the root system and above-ground parts of woody plants.
Data collection methods
The use of the SRS method for studying woody plants is rare. In the Web of Science database (as of 26 January 2022), out of 902 publications where the split-root keyword occurs, less than 100 papers describe research on woody plants. In total, 25 botanical families were represented in these studies, with the most numerous species from Pinaceae (24) and Rosaceae (16) and 11 each from the Vitaceae and Fabaceae families. Of the 57 species, the most numerous were Vitis vinifera (Vitaceae) and Malus domestica (Rosaceae), both of great economic importance. During the review preparation, the experiments were divided according to the method of root-system separation, a factor that differentiates a) the substrate in which the split roots are placed, b) areas of application of the SRS and c) the use of the isotope technique.
Root-system separation methods
Saiz-Fernández et al. (2021) proposed five basic methods for generating an SRS in their A. thaliana review. For woody plants, several additional methods for obtaining an SRS are known (Table 1). Each of these approaches has advantages and disadvantages, which define the research applications (Table 2).
Split-root system
The most commonly used method in woody plants for experimenting with an SRS is dividing the developed root into two parts of comparable size (i.e. split-developed root or SDR), and more than half of all studies in this review used this method. The SDR was applied in all areas identified in this study (Table 1). The advantage of this method is that it is easy. It is sufficient to roughly clean soil from the roots, split the root into two similar parts and place each set in a different container or on both sides of a partition. Another advantage is that the gradient of heterogeneous soil can be tested (Allen et al. 2000). The disadvantage of the SDR method is its limited applicability for plants characterised by a taproot presence, as it is impossible to separate one axial root into two parts. However, modifying the unequal SDR (uSDR), as in the case of Juglans regia (Juglandaceae) research, can be performed (Ameglio and Archer 1996; Ameglio et al. 1999). Another modification of this method is the division of the root into more parts, called multiple SDR (mSDR), which is not a commonly used method because it requires the formation of several similarly sized side roots.
The second most frequently used method is the separation of newly formed lateral roots (i.e. split newly forming roots or SNR). Separation of the root system can be achieved in this case in two ways: pruning the taproot, which induces the formation of lateral roots, or rooting shoots. The advantages of this method include the possibility of including species producing the taproot in the research and, in its second variant, the possibility of researching genetically identical plants, vegetatively propagated from a single individual. Both of these experimental approaches result in the formation of lateral roots. However, they are associated with physical damage to the plant, which may make plants more susceptible to pathogen infection (Gordon et al. 2015; Lakomy et al. 2019), inducing plant defence responses (Feng et al. 2022) and an imbalance in the proportion of the above- to below-ground parts (Zadworny et al. 2021). All of these can be considered disadvantages. With the SNR method, the water balance (Beis and Patakas 2015; Marino et al. 2017), ion transport (Boukcim et al. 2006; Cubera et al. 2009), signal transmissions (Gowing et al. 1990; Portz et al. 2011) and interaction with microorganisms (Bogar et al. 2019; Hao et al. 2012) in the plant were investigated.
Another two methods of preparing the SRS are cutting the roots longitudinally (CLR) or cutting longitudinal cuttings (CLC). These methods are rarely used; only 11 publications on their use were found. The advantages and disadvantages of these methods are similar to those of SNR. The advantage of these methods, especially in the variant where the cuttings are split, is that there is no need to replant the plant, and testing the influence of the substrate on the rooting of cuttings is possible (Palm et al. 2021). The disadvantage is the higher exposure to infection by fungal pathogens because the wound area of the plant tissue is significantly larger. Another disadvantage is that, until the experiment is complete, we do not know whether the newly formed roots are of comparable size in both variants. These methods can only be used on plants that tolerate root pruning very well or root easily, for example to study the transport of water and ions in the plant for CLC (Dry and Loveys 1999; Dry et al. 2000; Palm et al. 2021) and CLR (Catchpoole and Blair 1990; Romero-Conde et al. 2014; Shani et al. 1993; Tan et al. 2021; Wang et al. 2016a)). Marler (2013) used this method to describe the recognition response of Cycas edentata (Cycadaceae) to related plants among competitors for soil resources.
The least common method of creating an SRS is horticultural techniques to graft two seedlings, which can be performed using one of two methods: attaching a second root (i.e. inverted grafting) or approach grafting (a longitudinal incision of a shoot of two plants at the same height, binding them together and after some time, when two plants grow together removing one of the tops). Grafting has some significant advantages. The first is the possibility of experimenting with plants forming a taproot. The roots of the two grafted plants are not damaged because the plant grafting is within the shoot. The second advantage is that this method reduces or avoids stress during root-system transplanting. The greatest advantage of this method is combining plants from different origins, such as varieties or populations of the same species. The SRS achieved by grafting help to identify whether physiological processes (e.g. the end of the shoot growth and the establishment of the apical bud or the end of root length growth), are under plant internal (genetics) or external (environment) control. Thus, such a combination of genetically diverse plants can help to study signal transmission in plants or growth and development regulation mechanisms.
However, grafting also has many disadvantages. First, this method can be used only in species for which the grafting efficiency is high. Second, the grafting procedure is time-consuming and can be performed only at a specific time of the year. In addition, grafting is associated with tissue damage, which may be associated with pathogen infections. Grafting as a method of obtaining SRS will have a very limited application in plants characterized by a high degree of sectoriality. Another disadvantage of grafting methods is the need to wait longer for the combined plants to fuse, which often requires waiting through the growing season. These methods were primarily applied in creating an SRS in fruit trees, such as apple and citrus trees. The influence of irrigation and water transport was investigated regarding their application (Einhorn et al. 2009, 2012; Miranda et al. 2018), ion transport (Wang et al. 2016b), or signal transmission regulation in the plant (Zhao et al. 2008).
The method of natural separation of the growing plant’s root system (i.e. natural split or NS) has sometimes been used, especially in rhizotron research, which is particularly important in root development analysis in a heterogeneous substrate. The advantage of the NS method is that the damage caused by artificial root separation can be reduced. The disadvantages of the NS method include hardware limitations, as this method is used in rhizotrons, allowing the observation of root growth. If it were not possible to observe roots through the visors, the existence of a root separation could only be verified after the end of the experiment. Glenn and Welker (1991); Glenn and Welker (1993) used this method in studies of the effect of turf created by grasses on the water and nitrogen uptake by peach seedlings (Prunus persica; Rosaceae). Lilleskov and Bruns (2003) also employed this method for research on root colonisation by ectomycorrhizal fungi.
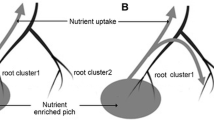
Some studies have presented more complex experimental setups using the SRS. Such an arrangement is the double split-root rhizotron, which separates the root systems of two tree seedlings into compartments with root strands of one seedling at each side and a shared root compartment in the centre where root strands of both tree seedlings can interact (Cesarz et al. 2013). In these studies, the effect of deciduous tree species on soil biota and carbon dynamics was analysed using isotope labelling.
Research using the split-root system
Effect of drought and water transport on plants
More than half of the publications using the SRS within trees concern the issues of water acquisition or its transport within plants. In this research, partial root drying (PRD) was the most common factor differentiating the substrate where the split roots were placed. Some researchers indicated that water transport from well-watered roots could compensate for the deficiencies in drained roots (Allen et al. 2000; Glenn and Welker 1993; Simonneau and Habib 1994). The results are sometimes not obvious because no clear compensation was seen, which may result from different experimental setups and a large variety of species. Nevertheless, we presented some of the most interesting ideas and examples of few studies and effort to make some generalizations. Experiments with the PRD have shown that partial restriction of water availability can increase the water use efficiency in Citrus paradis (Rutaceae) (Contreras-Barragan et al. 2014), Ginkgo biloba (Ginkgoaceae) (Wang et al. 2016a) and Persea americana (Lauraceae) (Neuhaus et al. 2007). This phenomenon may partially explain the presence of a hydraulic redistribution, transporting water from the well-watered part to the unwatered part, which occurs between neighbouring plants (Hafner et al. 2021, 2017; Hirota et al. 2004; Sakuratani et al. 1999; Töchterle et al. 2020). Without the use of SRS and a heterogeneous substrate, the discovery and description of a hydraulic redistribution would be impossible. Sufficient watering of only one part of the root while the other part was dried often has a negative effect on growth but increases the water use efficiency and does not reduce yield (Einhorn et al. 2009; Loveys et al. 1999; Poni et al. 2007).
The physiological explanation of these processes has often been based on the regulatory role of plant hormones. For example, drying out half of the root does not increase the abscisic acid (ABA) content or do not reduce the amount of water in the shoot (Fort et al. 1998). Moreover, gas exchange and ABA content in the leaves are related to the overall water consumption, and alternating (i.e. PRD) does not reduce the value of these traits (Einhorn et al. 2012). Experiments using PRD in the SRS of grapevines exhibited the regulatory effect of ABA and cytokinins on the reactions of the stomata (Beis and Patakas 2015). Some compensating mechanisms may be related to the intensity of the gas exchange, which is not always limited under mild stress conditions (Aganchich et al. 2009) because the sum of the diffusional resistances (i.e., stomatal and mesophyll resistances) sets the limit for photosynthetic rates. The results of many pot experiments with V. vinifera (Vitaceae) with an SRS (Antolin et al. 2006, 2008; Dry et al. 2000; Poni et al. 2007, 2009) resulted in interesting field experiments. Knowledge on the use of PRD in field conditions has been collected and presented in a recently published review (Slamini et al. 2022) and will not be discussed further in this paper.
A plant’s root system is characterised by high heterogeneity between and sometimes within species resulting from their life history, growth pattern, and other factors, which means that some root parts have a specific function in some species but not others. The SRS would considerably facilitate determining water uptake capacity in root systems that exhibit structural diversification. Research on the plant water supply using the SRS was conducted to analyse the sectorial character of plants (Dry and Loveys 1999; Shani et al. 1993). In some tree species (e.g. Acer rubrum (Sapindaceae) or Thuja occidentalis (Cupressaceae)), sectoriality is significant, which means that water transport from a specific root always takes place to a specific shoot (Gloser et al. 2008, 2009b). Ameglio et al. (1999) showed with the help of SRS experiments that the use of the predawn leaf water potential indicator is problematic because proper water supply of the root fragment (20%) is able to maintain the value of this indicator in walnut (Juglans regia; Juglandaceae) leaves at a level close to the control, while others were water shortage as indicated by sap flow measurements. Thus, SRS has so far been a very useful tool for studying many aspects of water uptake, transport and use by woody plants.
Nutrient uptake and transport
The experiments with the SRS also significantly contributed to the study of ion transport in plants. Usually, the experiments were based on the different fertilisation of individual parts of the roots. For example, the availability of nitrogen in the soil enhanced the proliferation of fine roots (e.g. A. rubrum (Sapindaceae) and Betula papyrifera (Betulaceae)) in the fertilised pot (Gloser et al. 2008). Moreover, heterogeneity was found in the maple in the above-ground part, with the leaves associated with the nitrogen-fertilised roots being larger and having a higher relative chlorophyll concentration. This finding may indicate that maple is more sectorial than birch. Tan et al. (2021) demonstrated a different reaction to the uptake of water and nutrients in a heterogeneous environment by birch (Betula platyphylla; Betulaceae) and larch (Larix olgensis; Pinaceae). Birch showed a higher foraging precision assessed by biomass and greater foraging plasticity assessed in morphology and physiology. In contrast, larch seedlings had higher root N concentration in the well-watered condition. Authors conclude that birch is better acclimatized to heterogeneous soil conditions and larch has a greater capacity to N uptake when water supply improves.
Interspecies differences are visible in many other studies, where the results sometimes indicate differences generated by uneven fertilisation (Boukcim et al. 2006; Cubera et al. 2009; Gentili 2006; Palm et al. 2021). However, this differentiation is not always revealed (George et al. 1997; McDonald et al. 1991). The lack of these differences may result from mechanisms to compensate for the uneven supply of nutrients (Gloser et al. 2008; Palm et al. 2021). The description of these mechanisms was possible thanks to the use of the SRS technique differentiating the availability of water and nutrients. The diversified nutrient supply in the soil increased the root proliferation in the fertilised soil of spruce (Picea abies; Pinaceae) and pine (Pinus sylvestris; Pinaceae), which was not found in Douglas fir (Pseudotsuga menziesii; Pinaceae) (George et al. 1997). The form of nitrogen fertilisation influenced the architecture of Cedrus atlantica (Pinaceae) roots, and an increased supply of ammonium (NH4) resulted in longer lateral roots than in the nitrate (NO3) fertilised variant (Boukcim et al. 2006). Different results were obtained for the oak (Quercus ilex; Fagaceae), where the increased availability of ammonium (NH4) limited root growth, and the SRS showed that unfavourable conditions of NH4 content have local consequences on the rooting depth and systemic consequences on the fine root density (Cubera et al. 2009). The most suitable NH4:NO3 ratio for a short-rotation poplar (Populus deltoides; Salicaceae) plantation was tested using the SRS in which 1–3% of the total root system was supplied with 2mMN at NH4:NO3 ratios of 0:100, 20:80, 40:60, 60:40, 80:20 and 100:0 (molar basis), with the rest of the plant supplied with 0 mM N, resulting in the whole plant becoming N deficient. The most developed root system was at a 20:80 ratio, therefore, the authors conclude that N form has a profound effect on root development and morphology in enriched patches (Woolfolk and Friend 2003). Tan et al. (2021) found that the nitrogen uptake in a heterogeneous environment is higher in birch (B. platyphylla; Betulaceae) than in larch (L. olgensis; Pinaceae) which was confirmed by greater biomass and greater morphological and physiological plasticity of the birch roots. On the other hand, larch responded better to the improvement of water conditions by increasing nitrogen uptake. Bogar et al. (2019) proved that in the SRS experiment (differentiated nitrogen availability and isolates of the mycorrhizal symbiont with different uptake efficiency) the allocation of carbon to the Pinus muricata (Pinaceae) roots and the associated ectomycorrhizal fungi is greater when they provided the plant with more nitrogen. Authors suggest that plants may be able to regulate this ectomycorrhizal symbiosis at a relatively fine scale, and that this regulation can be integrated across spatially separated portions of a root system.
In split-root experiments, soil nitrogen uptake by Eucalyptus grandis (Myrtaceae) roots was reduced with enhanced phosphorus concentrations, but if the enhance concerned only half of the root system, nitrogen uptake was similar to that of the control (Graciano et al. 2009). When an increased phosphorus concentration was used in only part of the root system, the nitrogen uptake did not differ from the control group because the decrease in nitrogen assimilation was counteracted by an increase in root hydraulic conductivity (Costa et al. 2017). In contrast, the deficiency of phosphorus in one part of the root system of an apple tree (Malus domestica; Rosaceae) may be supplemented by increased uptake in a part of the system better supplied with this element (Taylor and Goubran 1976).
Plants with the SRS are convenient models for studying the uptake and transport of trace and toxic elements. Wang et al. (2016b) showed using inverted grafting and labelledH310BO3 (B) that the inarched B-efficient rootstock genotype increases the boron supply of the plant, but decreases its content in the original Citrus sinensis (Rutaceae) root. Kuhn et al. (1995) using the SRS confirmed that the uptake of the basic elements magnesium (Mg), calcium (Ca), and potassium (K) by spruce roots (Picea abies; Pinaceae) is modified by the presence of aluminium (Al3+) ions and the pH of the substrate. The amount of elements (Ca, Mg and K) taken up by the root fragment placed in the Al substrate was higher, which was found thanks to the compounds labelled stable isotopes. The toxicity of high doses of Al3+ which was treated in one part of the eucalyptus roots is neutralised by adding phosphorus ions in the other part (Kang et al. 2020). The SRS method revealed that the toxic concentrations of zinc (Zn) for part of the willow root (Salix matsudana; Salicaceae) result in a greater allocation of carbon to the zone free of elevated concentrations of this element (Palm et al. 2021). This aspect may be useful in phytoremediation studies.
The use of the SRS of spruce (Picea abies; Pinaceae) and Douglas fir (Pseudotsuga menziesii; Pinaceae) in rhizotrons with a transparent front plate where microsuction cups were installed in the rhizosphere of the root made it possible to assess ion concentrations in individual root zones (Zhang and George 2009). The soil solution chemistry differs in the rhizosphere of various root types and different segments of single roots, and the high water consumption of the above-ground part causes the roots to take up more water than nutrients, leading to the accumulation of nutrients in the rhizosphere. This phenomenon was intensified after fertilisation, especially near the root tips. By placing micro-suction cups in different parts of the SRS, it was detected that old brown and suberized roots may also contribute to plant water and nutrients uptake. The SRS is also an excellent tool to study the effects of fertilisation on atmospheric nitrogen fixation by bacteria of the genus Frankia in sea buckthorn (Hippophaë rhamnoides; Elaeagnaceae) and results show that phosphorus stimulates a nitrogen (N2) fixation by direct effects on nodule (Gentili 2006). Through this experiment and the use of a heterogeneous substrate, it was also detected that phosphorus also stimulated N uptake from solution and influenced N isotope fractionation during N uptake.
Signal transmission
The signals derived from the root play critical roles in coordinating the shoot response to underground conditions, but recognition of these signals, especially over long distances, is scarce (Deng et al. 2021). The SRS appears to be a valuable tool for studying these processes. Lovisolo et al. (2002) used the SRS to study the reduction of plant hydraulic conductance during drought stress. Drying of only a part of the root resulted in an increase ABA in the whole plant, to the level observed where the whole root was subjected to drought stress. However, no reduction of whole-plant hydraulic conductance was observed. This suggests that the ABA drought signal does not affect the development and functionality of tissues transporting water at the level of the whole plant, determining its hydraulic conductance. Indeed, plants growing under harsh conditions may greatly benefit from an expanded root system that would communicate unfavourable conditions, increasing the ability of the entire plant to survive. Gowing et al. (1990) conducted an SRS experiment in which the dry part of the root system sends a signal that limits the growth of shoots and leaves of apple trees (Malus domestica; Rosaceae). These studies have proven that this signal is non-hydraulic because recovery to normal growth was achieved by re-irrigating or cutting off the desiccated root. Such conclusions would not be possible without the use of the SRS method.
Iron (Fe), as an essential element for the proper development of plants, plays a critical role in recognising signal transmission in the plant (Gayomba et al. 2015). Its availability depends on the soil pH. In alkaline soils, it is deficient, and in acidic soils, the concentrations may be toxic; therefore, maintaining its homeostasis requires a very efficient signal transmission system in the plant. Wu et al. (2012) demonstrated that depriving some of the roots of apple trees (Malus xiaojinensis; Rosaceae) of Fe ions induced a radical increase in Fe (III) reductase activity and extrusion of protons in the root part supplied with Fe. This outcome suggests that responses to iron deficiency are mediated by systemic signalling. Hu et al. (2014) presented the regulation at the whole-plant level and the local level of root growth towards areas richer in nitrogen. The SRS was used to gain information about the local and systemic consumption of soluble sugars by the pathogenic fungus of the genus Phytophthora (Peronosporaceae). Studies of Portz et al. (2011) showed that the response to the infection, expressed by decrease in the level of sucrose and increase in its degradation products (glucose and fructose), was very similar both in the infected and control root fragments. This proves the systemic response of beech (Fagus sylvatica; Fagaceae) roots infected with the pathogen.
Communication between individual organs, tissues or cells in plants, especially woody plants, is still poorly understood. Therefore we suppose that the use of SRS in the study of signal transmission in the plant may be particularly useful in distinguishing local or systemic responses to stress factors.
Interactions with microorganisms
The study of plant–pathogen/herbivore interaction or defence responses against pathogens requires a holistic approach, recording the responses in the affected and other parts of the plant. The SRS technique makes it possible to observe the defence reaction in, for example, the uninfected part of the root. Numerous publications have presented the results of experiments with SRS studies on the interaction of trees with soil microorganisms. For example, using the SRS technique with heterogeneous nutrient source by Lilleskov and Bruns (2003) showed that the colonisation of pine (Pinus muricata; Pinaceae) roots by late succession fungi (Tomentella sp.; Thelephoraceae) depends on the nutrient supply, which has not been found for early succession fungi (Rhizopogon sp.; Rhizopogonaceae). The authors stated that the strategy of Rhizpogon may be to colonize roots early in high resource soils; whereas that of Tomentella may be based upon slower colonization rates and greater competitive ability, so colonizes the roots also in the low nutrient soil. Vohnik et al. (2005) conducted an experiment using SRS to ensure spatial isolation of two fungi, inoculated into an individual root, and to screen the effects of inoculated fungi on their host plants. The root colonization of Rhododendron cv. Azurro (Ericaceae) by an ericoid mycorrhizal fungus improves the supply of phosphorus and nitrogen for leaves of whole plant, and that the supply is dependent on the species and strain of the fungus. Kennedy et al. (2009) confirmed that priority effects play a significant role in the dynamics of ectomycorrhizal fungi root tip colonisation. However, a study of the split-root experiment did show that the quantity of colonization on the side to which spores were added was not affected by the identity of the species present on the other part of the seedling. So establishing a partner reaction with the ectomycorrhizal fungi is very local. In contrast, Hao et al. (2012) documented a systemic response limiting the development of the parasitic nematode on both the roots of grapevine inoculated with arbuscular fungi and those not inoculated. A nonsystemic but local reaction was demonstrated for apple roots exposed to apple replant disease (Lucas et al. 2018). Bogar et al. (2019) found that plants can distinguish and select a mycorrhizal partner. The SRS experiments with Pinus muricata seedlings have shown that the root fragment associated with the ectomycorrhizal fungus, which provides the plant with more nitrogen, receives more carbon compounds in return. Moreover, carbon and nitrogen transfer depends on competition and soil resources (Bogar et al. 2022). These SRS experiments were also used to study the causes of apple replant disease. The disease is closely related to soil microorganisms, and the defence reactions of the roots at the biochemical level are local, as they were observed only in the part of the root exposed to apple replant disease (Balbin-Suarez et al. 2021, 2020; Rohr et al. 2021). The use of SRS in studies of arbuscular mycorrhizal and ectomycorrhizal symbioses, including several studies on woody plants, was presented by Kafle et al. (2022), so we will not discuss this issue in detail in this paper.
Studies of interactions of plants with other organisms using SRS can also help to determine the nature (systemic or local) of defensive reactions against pathogens, herbivores or parasitic plants. And one of the most interesting research areas offered by SRS is the study of the mechanisms of mycorrhizal establishment.
The remaining research areas in which the SRS was used are represented in the literature by single publications. Gorka et al. (2019) demonstrated that carbon obtained in the photosynthesis process by beech (Fagus sylvatica; Fagaceae) seedlings is transferred to the soil environment via the ectomycorrhizal hyphal sheath, and this process is selective and depends on the nitrogen resources in the soil. Cesarz et al. (2013) using a more complicated experimental setup (double SRS) hypothesized that modifications of the soil microorganism community and soil processes to be most pronounced in the mixed treatment with both tree species present due to complementary effects of the two tree species. They have found differences between beech (Fagus sylvatica; Fagaceae) and ash (Fraxinus excelsior; Oleaceae) in the influence on the soil environment. Beech more strongly affects the below-ground environment via exudates and the associated changes in rhizosphere microorganisms and carbon dynamics than ash. It has been shown that mixing of tree species increased plant biomass and mycorrhizal colonization but did not affect soil chemistry and microbial biomass. Marler (2013) studied the competition of cycad (Cycas edentate; Cycadaceae) roots and found that roots can recognise whether a neighbour is a related plant or from a remote population or another species. The related neighbourhood reduced the growth of the roots and above-ground parts, what has not been observed in unrelated neighbours. Bertoli et al. (2020) demonstrated that the ability to recognise related plants exists when analysing eucalyptus (Eucalyptus urophylla; Myrtaceae) seedlings. They found an inverse relationship: competition with individuals from other populations or half-siblings limited growth but not the competition with clonal specimens.
Use of isotope labelling
Differentiating the substrate and tracing the transport path of components in the roots employs a combination of the SRS technique and isotopically labelled compounds. The subject of these studies has primarily been various aspects of water and element uptake and their transport in the plant. In experiments using the SRS technique, the compounds labelled with stable isotopes of the elements were frequently used (about one-fourth of the studies). Nitrogen (15N) was used to study the following:
-
the transport of nitrogen between different plant species (Catchpoole and Blair 1990). They found nitrogen transfer from labelled half of the box to the roots of Leucaena leucocephala (Fabaceae) from unlabelled half one and then to other plants (e.g. grass)
-
the influence of different level of phosphorus on the uptake of nitrogen and N2 fixation in Hippophae rhamnoides (Elaeagnaceae) (Gentili 2006), where the stimulating effect of phosphorus was demonstrated,
-
the uptake, transport and use of nitrogen in photosynthesis in Picea abies (Pinaceae) and Thuja occidentalis (Cupressaceae) (Gloser et al. 2009b), it was found that, patchy N supply caused heterogeneous N distribution of newly acquired N within crown but both species have mechanisms that reduce adverse effects of patchy N supply,
-
the influence of two species, Fagus sylvatica (Fagaceae) and Fraxinus excelsior (Oleaceae), on soil microorganisms and carbon and nitrogen dynamics (Cesarz et al. 2013), where using a double split-root rhizotrons a significant impact of root exudate on the microbiome of the rhizosphere was demonstrated, hence the conclusion that the effects of living roots have to be included into studies on soil C dynamics to understand carbon and nutrient cycling,
-
nitrogen transfer in Pinus sylvestris (Pinaceae) from one part of the root system to another (Veerman et al. 2018), where the split-root method proved to be suitable to measure N rhizodeposition.
The methods using isotope labelling also tested the effect of different soil (infected with apple replant disease and control) on nitrate uptake efficiency, plants show no systemic response to apple replant disease although exchange between the two root parts occurred as indicated by 15N (Lucas et al. 2018). The influence of nitrogen availability on the relationship between Fagus sylvatica ectomycorrhiza hyphae and soil bacteria, when nitrogen was added to the litter compartments, bacterial biomass, and the amount of incorporated carbon strongly declined (Gorka et al. 2019). Whereas Bogar et al. (2022) show than Pinus muricata (Pinaceae) seedlings can exchange carbon for fungal nitrogen, but coupling of these resources can depend on the fungal species and soil nitrogen. Carbon (14C) was used to monitor tracers of distribution between the fruit and roots of Actinidia argute (Actinidiaceae). Cut off part of the root caused that more carbon went to the fruit, so in these plants, competitive strength for carbohydrate supply was higher in fruit compared with the remaining roots. This suggesting that the intact roots did not increase in function to compensate for the loss of root area (Black et al. 2012). Carbon (13C) was used to track the host P. muricata (Pinaceae) investment in fungi (Bogar et al. 2019). Where the results show that ectomycorrhizal plants can discriminate among potential fungal partners and that roots and fungi with greater nitrogen supplies received more plant carbon. This relationship is not universal because these results were not confirmed by studies on F. sylvatica, where, also with the use of SRS and labelled compounds (13C and 15N), a greater flow of assimilates to the roots providing more nitrogen was not found (Mayerhofer et al. 2021).
Calcium (42Ca and 44Ca), magnesium (25 Mg and 26 Mg) and potassium (41 K) were used to follow the path during uptake in the root and during long-range transport in the shoot (Kuhn et al. 1995). Phosphorus (32P) was used to examine within-plant phosphorus variability in Picea sitchensis (Pinaceae) and Pinus contorta (Pinaceae) (McDonald et al. 1991), it was shown that the varied content of P in the soil did not result in differentiation in the root tissues. Boron (10B) was used for studies of increasing boron uptake by inarching grafting of orange seedlings with a cultivar with higher uptake efficiency (Wang et al. 2016b). These results suggest that, under low B conditions, inarching B-efficient orange cultivar onto B-deficient one improves the plant’s B uptake. Oxygen (18O) was used to study water uptake in the halophytic mangrove Acicennia marina (Acanthaceae) (Reef et al. 2015). It is presented that Acicennia responds to transient freshwater patches by increasing water uptake from areas of the root zone where the saline enviroment is better for growth. Deuterium labelling (2H) was used to quantify the hydraulic redistribution in Picea abies (Pinaceae), Pseudotsuga menziesii (Pinaceae), Acer pseudoplatanus (Sapindaceae), Castanea sativa (Fagaceae), Fagus sylvatica (Fagaceae), Quercus robur (Fagaceae) and Abies alba (Pinaceae) (Hafner et al. 2020, 2021, 2017; Töchterle et al. 2020). For all these species, it has been proven that there is a hydraulic redistribution of water to less hydrated roots.
Perspectives
The presented research results using the SRS do not exhaust all possibilities offered by this method. The first issue that could be of assistance is the sectoriality/integrity of the vascular system of woody plants. Root functioning depends on the above-ground part and vice versa. Some researchers have assumed that this dependence is so strict that a specific leaf corresponds to a specific root (Shinozaki et al. 1964a, b). However, the degree of integration of the vascular system is species-specific, and there are systems ranging from full integration to full sectorial character (Lehnebach et al. 2018). Most research on tree sectoriality concerns deciduous species. Even among conifers that lack vessels, variation in sectoriality occurs (Augusto et al. 2011; Gloser et al. 2009a; Zanne et al. 2006). Experimental differentiation of soil conditions for fragments of the root system of one plant, especially with the use of stable isotope-labelled compounds, should help to identify the degree of the sectorial character of individual tree species. Understanding the sectoriality of woody plants can be very important in some cases, Marquis (1996) emphasizes that the sectoriality may influence a plants ability to tolerate tissue loss, Vuorisalo and Hutchings (1996) suggest that sectoriality affect competitive interactions between individual plants and Zanne et al. (2006) states that sectoriality may reduce embolism spread, which suggests that sectored species should be especially prominent in xeric environments.
Research using the SRS could facilitate determining the factors influencing the anatomical structure and functionality of roots, the moment of the end of the shoot in a given growing season, and other factors. Knowing which features of plants (trees) are genetically determined and which are environmentally related can help better predict the directions of environmental changes in the context of climate change. For example, biomass allocation to fine roots (Zadworny et al. 2016) or functional features related to photosynthesis and water use (de Miguel et al. 2014) are largely genetically determined. The discussion on the factors determining the moment of the end of shoot growth and the establishment of the apical bud in trees is still open. Some research has indicated photoperiodic and thermal regulation (Kvaalen and Johnsen 2008; Oleksyn et al. 2001), and others have proposed genetic determinants (Aldrete et al. 2008; Hodge et al. 2012; Sogaard et al. 2008). The moment of completion of the growth in height and the formation of the apical bud significantly influences root functioning because, from this point on, the assimilated carbon can be used to a greater extent for root growth. Experiments combining roots by inverted grafting from two individuals of differing origins (e.g. north population and south population) and cultivating them in the SRS could be used to determine the extent to which the environmental factors modify the genetic conditions of the root structure. Thanks to this, it could be determined whether the greater proportion of acquisitive roots in northern populations described by (Zadworny et al. 2016) is conditioned by the origin of the above-ground part of the tree and whether a change in environmental conditions, e.g. global warming, will change it. In natural conditions, the roots of even one tree very often live in different thermal conditions, which can be caused, for example, rooting depth, by the slope, the type of undergrowth plants, or artificial factors covering the soil. It is known that temperature changes can affect the desynchronization of the growth processes of the aboveground and belowground parts of plants (Abramoff and Finzi 2015). Root phenology is a poorly understood phenomenon, but it is known that it is dependent on soil temperature and in the context of climate change, recognition of these relationships seems very important (Radville et al. 2016). Experiments used SRS could simulate various temperatures in different parts of the roots, which would be a completely new experimental approach. They would allow to determine the influence of temperature as a regulator of various growth processes e.g. root respiration or inductor of signals sent by the roots.
Another area in which the SRS application is possible is the study of the holistic nature of defence responses, according to Mundim et al. (2017). Plants infected by pathogens or attacked by herbivores often react with various types of defence reactions (Karban and Baldwin 1997). This induced defence is often immediate chemical reactions to reduce infection, deteriorate food quality, or produce deterrents (Agrawal and Karban 1999). The answer to the following questions can be obtained through SRS experiments: Will the reaction to insect damage to one part of the root also occur in the other? Will the direction of carbon allocation be determined by the damage? Do trees, through the roots or mycorrhiza network, transmit information about damage to their neighbours, evoking similar defensive reactions? Certainly, there are many more questions that can be answered with the help of SRS.
The limitations cannot be ignored in presenting the areas in which the SRS method has been and can be used in tree research. In the case of trees, we can only test small individuals (i.e. young trees whose roots fit in pots or small containers). Another limitation is the time of conducting experiments because tree roots quickly fill the space available, which may inhibit their growth. Some negative factors include the potential damage and infection when preparing the seedlings. Although studies with SRS on mature trees are also conceivable, preparing such an experiment would require many years of waiting.
Conclusions
The review of the literature showed ten different ways of obtaining SRS. The selection of the appropriate method is conditioned by the goals set in the research as well as the limitations of each method. Research using SRS contributed most to the recognition of water transport in the plant, especially in partial root drying conditions, and to understanding the transport of key elements in heterogeneous soil conditions. Several interesting discoveries have been made through the use of SRS. For example, Sesbania rostrata (Fabaceae) has been shown to reverse sap flow to desiccated roots (Sakuratani et al. 1999). And the amount of water delivered by a hydraulic lift to the dry roots of Markhamia lutea (Bignoniaceae) and Quercus robur (Fagaceae) was sufficient for even neighbouring roots of other plants to survive (Hafner et al. 2021; Hirota et al. 2004). The ability of Cycas revoluta (Cycadaceae) seedlings to recognize neighbours was confirmed, and in the case of related plants, growth restriction was demonstrated (Marler 2013). Using SRS, it was shown that Picea abies (Pinaceae) and Thuja occidentalis (Cupressaceae) possess mechanisms that reduce adverse effects of patchy N supply and restricted nitrogen transport in xylem (Gloser et al. 2009b). It seems that the most interesting area where the use of the SRS technique can bring new results are the interactions of woody plants with organisms living in the soil. We mean both mycorrhizal fungi and many representatives of soil fauna. So far, there is little work in this field and it is difficult to make generalizations regarding, for example, the nature of the response (local or systemic) to pathogen infection or damage caused by herbivores.
Data availability
This literature review did not generate any new data that would not be presented in the paper.
References
Abramoff RZ, Finzi AC (2015) Are above- and below-ground phenology in sync? New Phytol 205:1054–1061. https://doi.org/10.1111/nph.13111
Aganchich B, Tahi H, Wahbi S, Elmodaffar C, Serraj R (2007) Growth, water relations and antioxidant defence mechanisms of olive (Olea europaea L.) subjected to Partial Root Drying (PRD) and Regulated Deficit Irrigation (RDI). Plant Biosystems 141:252–264. https://doi.org/10.1080/11263500701401893
Aganchich B, Wahbi S, Loreto F, Centritto M (2009) Partial root zone drying: regulation of photosynthetic limitations and antioxidant enzymatic activities in young olive (Olea europaea) saplings. Tree Physiol 29:685–696. https://doi.org/10.1093/treephys/tpp012
Agrawal AA, Karban R (1999) Why induced defenses may be favored over constitutive strategies in plants. In: RTaCD Harvell (ed) The ecology and evolution ofinducible defenses Princeton University PresSy, Princeton
Aldrete A, Mexal JG, Burr KE (2008) Seedling cold hardiness, bud set, and bud break in nine provenances of Pinus greggii Engelm. For Ecol Manage 255:3672–3676. https://doi.org/10.1016/j.foreco.2008.02.054
Allen LH, Brakke MP, Baker JT, Jones JW (2000) Gas exchange and biomass responses of young citrus trees to partial rooting-volume irrigation. Soil Crop Sci Soc Fla Proc 59:37–45
Ameglio T, Archer P (1996) Significance of predawn water potential in heterogeneous soil water content distribution. Agronomie 16:493–503. https://doi.org/10.1051/agro:19960803
Ameglio T, Archer P, Cruiziat P, Daudet FA, Valancogne C, Dayau S, Cohen M (1997) A limit in the use of predawn leaf water potential for tree irrigation. Acta Horticulturae (449):431–438. https://doi.org/10.17660/ActaHortic.1997.449.60
Ameglio T, Archer P, Cohen M, Valancogne C, Daudet FA, Dayau S, Cruiziat P (1999) Significance and limits in the use of predawn leaf water potential for tree irrigation. Plant Soil 207:155–167
Antolin MC, Ayari M, Sanchez-Diaz M (2006) Effects of partial rootzone drying on yield, ripening and berry ABA in potted Tempranillo grapevines with split roots. Aust J Grape Wine Res 12:13–20. https://doi.org/10.1111/j.1755-0238.2006.tb00039.x
Antolin MC, Santesteban H, Maria ES, Aguirreolea J, Sanchez-Diaz M (2008) Involvement of abscisic acid and polyamines in berry ripening of Vitis vinifera (L.) subjected to water deficit irrigation. Aust J Grape Wine Res 14:123–133. https://doi.org/10.1111/j.1755-0238.2008.00014.x
Augusto L, Zeller B, Midwood AJ, Swanston C, Dambrine E, Schneider A, Bosc A (2011) Two-year dynamics of foliage labelling in 8-year-old Pinus pinaster trees with 15N, 26Mg and 42Ca-simulation of Ca transport in xylem using an upscaling approach. Ann for Sci 68:169–178. https://doi.org/10.1007/s13595-011-0018-x
Balbin-Suarez A, Jacquiod S, Rohr AD, Liu BY, Flachowsky H, Winkelmann T, Beerhues L, Nesme J, Sorensen SJ, Vetterlein D, Smalla K (2021) Root exposure to apple replant disease soil triggers local defense response and rhizoplane microbiome dysbiosis. Fems Microbiol Ecol 97. https://doi.org/10.1093/femsec/fiab031
Balbin-Suarez A, Lucas M, Vetterlein D, Sorensen SJ, Winkelmann T, Smalla K, Jacquiod S (2020) Exploring microbial determinants of apple replant disease (ARD): a microhabitat approach under split-root design. Fems Microbiol Ecol 96. https://doi.org/10.1093/femsec/fiaa211
Beis A, Patakas A (2015) Differential physiological and biochemical responses to drought in grapevines subjected to partial root drying and deficit irrigation. Eur J Agron 62:90–97. https://doi.org/10.1016/j.eja.2014.10.001
Bertoli SC, Neris DM, Sala HR, Vieira WD, Souza GM (2020) The level of relatedness affects self/nonself discrimination in Eucalyptus urophylla seedlings. Can J Res 50:500–509. https://doi.org/10.1139/cjfr-2019-0376
Black MZ, Minchin PEH, Gould N, Patterson KJ, Clearwater MJ (2012) Measurement of Bremsstrahlung radiation for in vivo monitoring of 14C tracer distribution between fruit and roots of kiwifruit (Actinidia arguta) cuttings. Planta 236:1327–1337. https://doi.org/10.1007/s00425-012-1685-z
Bogar L, Peay K, Kornfeld A, Huggins J, Hortal S, Anderson I, Kennedy P (2019) Plant-mediated partner discrimination in ectomycorrhizal mutualisms. Mycorrhiza 29:97–111. https://doi.org/10.1007/s00572-018-00879-7
Bogar LM, Tavasieff OS, Raab TK, Peay KG (2022) Does resource exchange in ectomycorrhizal symbiosis vary with competitive context and nitrogen addition? New Phytol 233:1331–1344. https://doi.org/10.1111/nph.17871
Boukcim H, Pages L, Mousain D (2006) Local NO3- or NH4+ supply modifies the root system architecture of Cedrus atlantica seedlings grown in a split-root device. J Plant Physiol 163:1293–1304. https://doi.org/10.1016/j.jplph.2005.08.011
Catchpoole DW, Blair GJ (1990) Forage tree legumes. II. Investigation of nitrogen transfer to an associated grass using a split-root technique. Aust J Agric Res 41:531–537. https://doi.org/10.1071/ar9900531
Cesarz S, Fender AC, Beyer F, Valtanen K, Pfeiffer B, Gansert D, Hertel D, Polle A, Daniel R, Leuschner C, Scheu S (2013) Roots from beech (Fagus sylvatica L.) and ash (Fraxinus excelsior L.) differentially affect soil microorganisms and carbon dynamics. Soil Biol Biochem 61:23–32. https://doi.org/10.1016/j.soilbio.2013.02.003
Contreras-Barragan BA, Kusakabe A, Melgar JC, Nelson SD (2014) Understanding partial rootzone drying in citrus. 3rd International Symposium on Citrus Biotechnology. Int Soc Horticultural Science, Shizuoka, Japan
Costa L, Faustino LI, Graciano C (2017) The spatial distribution of phosphate in the root system modulates N metabolism and growth in Eucalyptus grandis young plants. Trees-Struct Funct 31:247–257. https://doi.org/10.1007/s00468-016-1480-8
Cubera E, Moreno G, Solla A (2009) Quercus ilex root growth in response to heterogeneous conditions of soil bulk density and soil NH4-N content. Soil Tillage Res 103:16–22. https://doi.org/10.1016/j.still.2008.09.002
de Miguel M, Cabezas JA, de Maria N, Sanchez-Gomez D, Guevara MA, Velez MD, Saez-Laguna E, Diaz LM, Mancha JA, Barbero MC, Collada C, Diaz-Sala C, Aranda I, Cervera MT (2014) Genetic control of functional traits related to photosynthesis and water use efficiency in Pinus pinaster Ait. drought response: integration of genome annotation, allele association and QTL detection for candidate gene identification. BMC Genomics 15:19. https://doi.org/10.1186/1471-2164-15-464
Deng ZY, Wu HY, Li DY, Li LP, Wang ZP, Yuan WY, Xing YZ, Li CD, Liang DC (2021) Root-to-Shoot Long-Distance Mobile miRNAs Identified from Nicotiana Rootstocks. Int J Mol Sci 22. https://doi.org/10.3390/ijms222312821
Dighton J, Mason P (1985) Mycorrhizal dynamics during forest tree development. In Development Biology of Higher Fungi. D., Moore, L.A., Castellano, D., Wood, J.C., Frankland (eds.). University Press Cambridge, Cambridge, pp. 117–139.
Dry PR, Loveys BR (1999) Grapevine shoot growth and stomatal conductance are reduced when part of the root system is dried. Vitis 38:151–156
Dry PR, Loveys BR, During H (2000) Partial drying of the rootzone of grape. I. Transient changes in shoot growth and gas exchange. Vitis 39:3–7
Einhorn T, Caspari HW, Green S (2009) Estimation of containerized single-stem and split-rooted, non-fruiting apple tree water use using miniaturized heat pulse probes. 7th International Workshop on Sap Flow, Sevilla, Spain
Einhorn TC, Caspari HW, Green S (2012) Total soil water content accounts for augmented ABA leaf concentration and stomatal regulation of split-rooted apple trees during heterogeneous soildrying. J Exp Bot 63:5365–5376. https://doi.org/10.1093/jxb/ers195
Feng XH, An P, Guo K, Li XG, Liu XJ, Zhang XM (2017) Growth root compensation and ion distribution in Lycium chinense under heterogeneous salinity stress Scientia Horticulturae 226:24–32. https://doi.org/10.1016/j.scienta.2017.08.011
Feng ZP, Kong DL, Kong YH, Zhang BH, Yang XT (2022) Coordination of root growth with root morphology, physiology and defense functions in response to root pruning in Platycladus orientalis. J Adv Res 36:187–199. https://doi.org/10.1016/j.jare.2021.07.005
Fort C, Muller F, Label P, Granier A, Dreyer E (1998) Stomatal conductance, growth and root signaling in Betula pendula seedlings subjected to partial soil drying. Tree Physiol 18:769–776. https://doi.org/10.1093/treephys/18.11.769
Fucik JE, Titus JS (1965) Split-root studies on calcium and manganese absorption and translocation in seedling apple trees. Proc Am Soc Hortic Sci 86:12–000
Gayomba SR, Zhai ZY, Jung HI, Vatamaniuk OK (2015) Local and systemic signaling of iron status and its interactions with homeostasis of other essential elements. Front Plant Sci 6. https://doi.org/10.3389/fpls.2015.00716
Gentili F (2006) Phosphorus, nitrogen and their interactions affect N2 fixation, N isotope fractionation and N partitioning in Hippophae rhamnoides. Symbiosis 41:39–45
George E, Seith B, Schaeffer C, Marschner H (1997) Responses of Picea, Pinus and Pseudotsuga roots to heterogeneous nutrient distribution in soil. Tree Physiol 17:39–45
Glenn DM, Welker WV (1991) Soil management affects shoot and root growth, nutrient availability, and water uptake of young peach trees. J Am Soc Hortic Sci Jashs 116:238–241. https://doi.org/10.21273/jashs.116.2.238
Glenn DM, Welker WV (1993) Water transfer diminishes root competition between peach and tall fescue. J Am Soc Hortic Sci 118:570–574. https://doi.org/10.21273/jashs.118.5.570
Gloser V, Ek P, Orians C (2009a) Shoot heterogeneity in trees: Consequences of patchy N availability and vascular transport. Nova Acta Leopoldina Neue Folge 96:135–141
Gloser V, Libera K, Orians CM (2008) Contrasting below- and aboveground responses of two deciduous trees to patchy nitrate availability. Tree Physiol 28:37–44. https://doi.org/10.1093/treephys/28.1.37
Gloser V, Sedlacek P, Gloser J (2009b) Consequences of nitrogen deficiency induced by low external N concentration and by patchy N supply in Picea abies and Thuja occidentalis. Trees-Struct Funct 23:1–9. https://doi.org/10.1007/s00468-008-0248-1
Gordon TR, Swett CL, Wingfield MJ (2015) Management of Fusarium diseases affecting conifers. Crop Prot 73:28–39. https://doi.org/10.1016/j.cropro.2015.02.018
Gorka S, Dietrich M, Mayerhofer W, Gabriel R, Wiesenbauer J, Martin V, Zheng Q, Imai B, Prommer J, Weidinger M, Schweiger P, Eichorst SA, Wagner M, Richter A, Schintlmeister A, Woebken D, Kaiser C (2019) Rapid transfer of plant photosynthates to soil bacteria via ectomycorrhizal hyphae and its interaction with nitrogen availability. Front Microbiol 10. https://doi.org/10.3389/fmicb.2019.00168
Gowing DJG, Davies WJ, Jones HG (1990) A positive root-sourced signal as an indicator of soil drying in apple, Malus x domestica Borkh. J Exp Bot 41:1535–1540. https://doi.org/10.1093/jxb/41.12.1535
Graciano C, Tambussi EA, Castan E, Guiamet JJ (2009) Dry mass partitioning and nitrogen uptake by Eucalyptus grandis plants in response to localized or mixed application of phosphorus. Plant Soil 319:175–184. https://doi.org/10.1007/s11104-008-9859-y
Hafner BD, Hesse BD, Bauerle TL, Grams TEE (2020) Water potential gradient, root conduit size and root xylem hydraulic conductivity determine the extent of hydraulic redistribution in temperate trees. Funct Ecol 34:561–574. https://doi.org/10.1111/1365-2435.13508
Hafner BD, Hesse BD, Grams TEE (2021) Friendly neighbours: Hydraulic redistribution accounts for one quarter of water used by neighbouring drought stressed tree saplings. Plant Cell Environ 44:1243–1256. https://doi.org/10.1111/pce.13852
Hafner BD, Tomasella M, Haberle KH, Goebel M, Matyssek R, Grams TEE (2017) Hydraulic redistribution under moderate drought among English oak, European beech and Norway spruce determined by deuterium isotope labeling in a split-root experiment. Tree Physiol 37:950–960. https://doi.org/10.1093/treephys/tpx050
Hao ZP, Fayolle L, van Tuinen D, Chatagnier O, Li XL, Gianinazzi S, Gianinazzi-Pearson V (2012) Local and systemic mycorrhiza-induced protection against the ectoparasitic nematode Xiphinema index involves priming of defence gene responses in grapevine. J Exp Bot 63:3657–3672. https://doi.org/10.1093/jxb/ers046
Hirota I, Sakuratani T, Sato T, Higuchi H, Nawata E (2004) A split-root apparatus for examining the effects of hydraulic lift by trees on the water status of neighbouring crops. Agrofor Syst 60:181–187. https://doi.org/10.1023/b:Agfo.0000013293.77907.64
Hodge A (2006) Plastic plants and patchy soils. J Exp Bot 57:401–411. https://doi.org/10.1093/jxb/eri280
Hodge GR, Dvorak WS, Tighe ME (2012) Comparisons between laboratory and field results of frost tolerance of pines from the southern USA and Mesoamerica planted as exotics. S For 74:7–17. https://doi.org/10.2989/20702620.2012.683637
Hu FQ, Mou PP, Weiner J, Li S (2014) Contrasts between whole-plant and local nutrient levels determine root growth and death in Ailanthus altissima (Simaroubaceae). Am J Bot 101:812–819. https://doi.org/10.3732/ajb.1400129
Kafle A, Frank HER, Rose BD, Garcia K (2022) Split down the middle: studying arbuscular mycorrhizal and ectomycorrhizal symbioses using split-root assays. J Exp Bot 73:1288–1300. https://doi.org/10.1093/jxb/erab489
Kang YC, Zhu SD, Li GY, Jiang YB, Teng WC, Luo M, Wei J, Cao F, Wang Z, Huang J (2020) Phosphorus applied to the root half without Al3+ exposure can alleviate Al toxicity on the other root half of the same eucalyptus seedling. J Plant Nutr 44:829–844. https://doi.org/10.1080/01904167.2020.1867579
Karban R, Baldwin IT (1997) Induced responses to herbivory. University of Chicago Press Chicago Illinois USA
Kennedy PG, Peay KG, Bruns TD (2009) Root tip competition among ectomycorrhizal fungi: Are priority effects a rule or an exception? Ecology 90:2098–2107. https://doi.org/10.1890/08-1291.1
Kuhn AJ, Bauch J, Schroder WH (1995) Monitoring uptake and contents of Mg, Ca and K in Norway spruce as influenced by pH and Al, using microprobe analysis and stable-isotope labeling. Plant Soil 168:135–150. https://doi.org/10.1007/bf00029322
Kvaalen H, Johnsen O (2008) Timing of bud set in Picea abies is regulated by a memory of temperature during zygotic and somatic embryogenesis. New Phytol 177:49–59. https://doi.org/10.1111/j.1469-8137.2007.02222.x
Lakomy P, Kuzminski R, Mucha J, Zadworny M (2019) Effects of oak root pruning in forest nurseries on potential pathogen infections. For Pathol 49. https://doi.org/10.1111/efp.12513
Lehnebach R, Beyer R, Letort V, Heuret P (2018) The pipe model theory half a century on: a review. Ann Bot 121:773–795. https://doi.org/10.1093/aob/mcx194
Lilleskov EA, Bruns TD (2003) Root colonization dynamics of two ectomycorrhizal fungi of contrasting life history strategies are mediated by addition of organic nutrient patches. New Phytol 159:141–151. https://doi.org/10.1046/j.1469-8137.2003.00794.x
Li X, Gao Y, Jian Q, Zhang Z, Liu S, Chen F, Zhang J (2017) Ferric reduction oxidase 2 gene from Pyrus betulifolia Bunge is regulated by iron deficiency and auxin. Can J Plant Sci 97:569–579. https://doi.org/10.1139/CJPS-2016-0213
Long EM (1943) The effect of salt additions to the substrate on intake of water and nutrients by roots of approach-grafted tomato plants. Am J Bot 30:594–601
Loveys BR, Dry PR, Stoll M, McCarthy MG (1999) Using plant physiology to improve the water use efficiency of horticultural crops. 3rd International Symposium on Irrigation of Horticultural Crops. International Society Horticultural Science, Estoril, Portugal
Lovisolo C, Hartung W, Schubert A (2002) Whole-plant hydraulic conductance and root-to-shoot flow of abscisic acid are independently affected by water stress in grapevines. Funct Plant Biol 29:1349–1356. https://doi.org/10.1071/fp02079
Lucas M, Balbin-Suarez A, Smalla K, Vetterlein D (2018) Root growth, function and rhizosphere microbiome analyses show local rather than systemic effects in apple plant response to replant disease soil. Plos One 13. https://doi.org/10.1371/journal.pone.0204922
Marino G, Brunetti C, Tattini M, Romano A, Biasioli F, Tognetti R, Loreto F, Ferrini F, Centritto M (2017) Dissecting the role of isoprene and stress-related hormones (ABA and ethylene) in Populus nigra exposed to unequal root zone water stress. Tree Physiol 37:1637–1647. https://doi.org/10.1093/treephys/tpx083
Marler TE (2013) Kin recognition alters root and whole plant growth of split-root Cycas edentata seedlings. HortScience 48:1266–1269. https://doi.org/10.21273/hortsci.48.10.1266
Marquis RJ (1996) Plant architecture, sectoriality and plant tolerance to herbivores. Vegetatio 127:85–97. https://doi.org/10.1007/BF00054850
Mayerhofer W, Schintlmeister A, Dietrich M, Gorka S, Wiesenbauer J, Martin V, Gabriel R, Reipert S, Weidinger M, Clode P, Wagner M, Woebken D, Richter A, Kaiser C (2021) Recently photoassimilated carbon and fungus-delivered nitrogen are spatially correlated in the ectomycorrhizal tissue of Fagus sylvatica. New Phytol 232:2457–2474. https://doi.org/10.1111/nph.17591
Mazzola M, Zhao X, Cohen MF, Raaijmakers JM (2007) Cyclic lipopeptide surfactant production by Pseudomonas fluorescens SS101 is not required for suppression of complex Pythium spp. populations. Phytopathology 97(10):1348–1355. https://doi.org/10.1094/PHYTO-97-10-1348
Meng YL, Xu XF, Khanizadeh S, Zhang MJ, Wang Q, Han ZH (2008) Contribution of abscisic acid to sorbitol accumulation in drought-stressed Malus hupehensis. J Food Agric Environ 6:319–326
McDonald MA, Malcolm DC, Harrison AF (1991) The use of a 32P root bioassay to indicate the phosphorus status of forest trees. 2. Spatial variation. Can J Res 21:1185–1193. https://doi.org/10.1139/x91-166
Miranda MT, da Silva SF, Moura BB, Hayashi AH, Machado EC, Ribeiro RV (2018) Hydraulic redistribution in Citrus rootstocks under drought. Theor Exp Plant Physiol 30:165–172. https://doi.org/10.1007/s40626-018-0111-8
Mundim FM, Alborn HT, Vieira-Neto EHM, Bruna EM (2017) A whole-plant perspective reveals unexpected impacts of above- and belowground herbivores on plant growth and defense. Ecology 98:70–78. https://doi.org/10.1002/ecy.1619
Neuhaus A, Turner DW, Colmer TD, Kuo J, Eastham J (2007) Drying half the root-zone of potted avocado (Persea americana Mill., cv. Hass) trees avoids the symptoms of water deficit that occur under complete root-zone drying. J Horticult Sci Biotechnol 82:679–689. https://doi.org/10.1080/14620316.2007.11512291
Oleksyn J, Reich PB, Tjoelker MG, Chalupka W (2001) Biogeographic differences in shoot elongation pattern among European Scots pine populations. For Ecol Manage 148:207–220
Orians CM, van Vuuren MMI, Harris NL, Babst BA, Ellmore GS (2004) Differential sectoriality in long-distance transport in temperate tree species: evidence from dye flow, 15N transport, and vessel element pitting. Trees-Struct Funct 18:501–509. https://doi.org/10.1007/s00468-004-0326-y
Palm E, Nissim WG, Mancuso S, Azzarello E (2021) Split-root investigation of the physiological response to heterogeneous elevated Zn exposure in poplar and willow. Environ Exp Bot 183. https://doi.org/10.1016/j.envexpbot.2020.104347
Poni S, Bernizzoni F, Civardi S (2007) Response of “Sangiovese”’ grapevines to partial root-zone drying: Gas-exchange, growth and grape composition. Sci Hortic 114:96–103. https://doi.org/10.1016/j.scienta.2007.06.003
Poni S, Bernizzoni F, Civardi S, Gatti M, Porro D, Camin F (2009) Performance and water-use efficiency (single-leaf vs. whole-canopy) of well-watered and half-stressed split-root Lambrusco grapevines grown in Po Valley (Italy). Agric Ecosyst Environ 129:97–106. https://doi.org/10.1016/j.agee.2008.07.009
Portz RL, Fleischmann F, Koehl J, Fromm J, Ernst D, Pascholati SF, Osswald WF (2011) Histological, physiological and molecular investigations of Fagus sylvatica seedlings infected with Phytophthora citricola. For Pathol 41:202–211. https://doi.org/10.1111/j.1439-0329.2010.00667.x
Radville L, McCormack ML, Post E, Eissenstat DM (2016) Root phenology in a changing climate. J Exp Bot 67:3617–3628. https://doi.org/10.1093/jxb/erw062
Reef R, Markham HL, Santini NS, Lovelock CE (2015) The response of the mangrove Avicennia marina to heterogeneous salinity measured using a split-root approach. Plant Soil 393:297–305. https://doi.org/10.1007/s11104-015-2489-2
Rohr AD, Staudt J, Cziborra K, Fritz A, Schmitz M, Winkelmann T (2021) Split-root approach reveals localized root responses towards apple replant disease (ARD) in terms of ARD biomarker gene expression and content of phenolic compounds. Sci Hortic 286. https://doi.org/10.1016/j.scienta.2021.110117
Romero-Conde A, Kusakabe A, Melgar JC (2014) Physiological responses of citrus to partial rootzone drying irrigation. Sci Hortic 169:234–238. https://doi.org/10.1016/j.scienta.2014.02.022
Rose BD, Frank HER, Garcia K (2023) Development of split-root assays for loblolly pine (Pinus taeda L.) seedlings to study ectomycorrhizal symbioses. MethodsX 10:102046. https://doi.org/10.1016/j.mex.2023.102046
Saiz-Fernández I, Černý M, Skalák J, Brzobohatý B (2021) Split-root systems: detailed methodology, alternative applications, and implications at leaf proteome level. Plant Methods 17:7. https://doi.org/10.1186/s13007-020-00706-1
Sakuratani T, Aoe T, Higuchi H (1999) Reverse flow in roots of Sesbania rostrata measured using the constant power heat balance method. Plant Cell Environ 22:1153–1160. https://doi.org/10.1046/j.1365-3040.1999.00464.x
Shane MW, de Vos M, de Roock S, Cawthray GR, Lambers H (2003) Effects of external phosphorus supply on internal phosphorus concentration and the initiation growth and exudation of cluster roots in Hakea prostrata R. Br. Plant Soil 248(1/2):209–219. https://doi.org/10.1023/A:1022320416038
Shane MW, Lambers H (2006) Systemic suppression of cluster-root formation and net P-uptake rates in Grevillea crithmifolia at elevated P supply: a proteacean with resistance for developing symptoms of ‘P toxicity.’ J Exp Bot 57(2):413–423. https://doi.org/10.1093/jxb/erj004
Shani U, Waisel Y, Eshel A, Xue S, Ziv G (1993) Responses to salinity of grapevine plants with split root systems. New Phytol 124:695–701. https://doi.org/10.1111/j.1469-8137.1993.tb03860.x
Shinozaki K, Toda K, Hozumi K, Kira T (1964a) A quantitative analysis of plant form − the pipe model theory. 1. Basic analyses. Jpn J Ecol 14:97–105
Shinozaki K, Toda K, Hozumi K, Kira T (1964b) A quantitative analysis of plant form − the pipe model theory. 2. Further evidence ot the theory and its application in forest ecology. Jpn J Ecol 14:133–139
Simonneau T, Habib R (1994) Water-uptake regulation in peach-trees with split-root systems. Plant Cell Environ 17:379–388. https://doi.org/10.1111/j.1365-3040.1994.tb00306.x
Slamini M, Sbaa M, Arabi M, Darmous A (2022) Review on Partial Root-zone Drying irrigation: Impact on crop yield, soil and water pollution. Agric Water Manag 271:107807. https://doi.org/10.1016/j.agwat.2022.107807
Sogaard G, Johnsen O, Nilsen J, Junttila O (2008) Climatic control of bud burst in young seedlings of nine provenances of Norway spruce. Tree Physiol 28:311–320. https://doi.org/10.1093/treephys/28.2.311
Srikasetsarakul U, Sringarm K, Sruamsiri P, Chiang M, Thailand S, Ongprasert W, Spreer J, Müller J, Thailand W, Wiriya A, Thailand (2011). Biomass formation and nutrient partitioning in potted longan trees under partial rootzone drying. Acta Horticulturae 889:587–592
Tagliavini M, Marangoni B (1994) Effects of peach root residues in soil on growth, water relations and mineral status of peach trees with split root system. Acta Horticulturae 363:93–100. https://doi.org/10.17660/ActaHortic.1994.363.12
Tan L, Fan RF, Sun HF, Guo SL (2021) Root foraging of birch and larch in heterogeneous soil nutrient patches under water deficit. Plos One 16. https://doi.org/10.1371/journal.pone.0255848
Taylor BK, Goubran FH (1976) Phosphorus nutrition of apple tree. 2. Effects of localized phosphate placement on growth and phosphorus content of split-root trees. Aust J Agric Res 27:533–539. https://doi.org/10.1071/ar9760533
Thomas DS, Eamus D, Shanahan S (2000) Influence of season drought and xylem ABA on stomatal responses to leaf-to-air vapour pressure difference of trees of the Australian wet-dry tropics. Aust J Bot 48(2):143–151. https://doi.org/10.1071/BT98090
Töchterle P, Yang FL, Rehschuh S, Rehschuh R, Ruehr NK, Rennenberg H, Dannenmann M (2020) Hydraulic water redistribution by silver fir (Abies alba Mill.) occurring under severe soil drought. Forests 11:12 https://doi.org/10.3390/f11020162
Torres LF, de Andrade SAL, Mazzafera P (2021) Split-root, grafting and girdling as experimental tools to study root-to shoot-to root signaling. Environ Exp Bot 191:13. https://doi.org/10.1016/j.envexpbot.2021.104631
Turner DW, Menzel CM, Simpson DR (1996) Short term drying of half the root system reduces growth but not water status or photosynthesis in leaves of passionfruit (Passiflora sp.). Scientia Horticulturae 65(1):25–36. https://doi.org/10.1016/0304-4238(95)00849-7
Veerman L, Kalbitz K, Schoorl JC, Tietema A (2018) Split-root labelling to investigate 15N rhizodeposition by Pinus sylvestris and Picea abies. Isot Environ Health Stud 54:16–27. https://doi.org/10.1080/10256016.2017.1352586
Vohnik M, Albrechtova J, Vosatka M (2005) The inoculation with Oidiodendron maius and Phialocephala fortinii alters phosphorus and nitrogen uptake, foliar C: N ratio and root biomass distribution in Rhododendron cv. Azurro. Symbiosis 40:87–96
Vuorisalo T, Hutchings MJ (1996) On plant sectoriality, or how to combine the benefits of autonomy and integration. Vegetatio 127:3–8. https://doi.org/10.1007/BF00054841
Wang L, Shi H, Wu JS, Cao FL (2016a) Alternative partial root-zone irrigation enhances leaf flavonoid accumulation and water use efficiency of Ginkgo biloba. New For 47:377–391. https://doi.org/10.1007/s11056-015-9521-5
Wang NN, Wei QJ, Yan TS, Pan ZY, Liu YZ, Peng S (2016b) Improving the boron uptake of boron-deficient navel orange plants under low boron conditions by inarching boron-efficient rootstock. Sci Hortic 199:49–55. https://doi.org/10.1016/j.scienta.2015.12.014
Wiriya-Alongkorn W, Spreer W, Ongprasert S, Spohrer K, Muller J (2016) Influence of water supply on CO2 concentration in the rootzone of split-root potted longan trees. 9th International Conference on Intelligent Robotics and Applications (ICIRA). Springer Int Publishing Ag, Hachioji, JAPAN
Woolfolk WTM, Friend AL (2003) Growth response of cottonwood roots to varied NH4:NO3 ratios in enriched patches. Tree Physiol 23:427–432. https://doi.org/10.1093/treephys/23.6.427
Wu T, Zhang HT, Wang Y, Jia WS, Xu XF, Zhang XZ, Han ZH (2012) Induction of root Fe(lll) reductase activity and proton extrusion by iron deficiency is mediated by auxin-based systemic signalling in Malus xiaojinensis. J Exp Bot 63:859–870. https://doi.org/10.1093/jxb/err314
Zadworny M, McCormack ML, Mucha J, Reich PB, Oleksyn J (2016) Scots pine fine roots adjust along a 2000-km latitudinal climatic gradient. New Phytol 212:389–399. https://doi.org/10.1111/nph.14048
Zadworny M, Mucha J, Jagodzinski AM, Koscielniak P, Lakomy P, Modrzejewski M, Ufnalski K, Zytkowiak R, Comas LH, Rodriguez-Calcerrada J (2021) Seedling regeneration techniques affect root systems and the response of Quercus robur seedlings to water shortages. For Ecol Manage 479:11. https://doi.org/10.1016/j.foreco.2020.118552
Zanne AE, Sweeney K, Sharma M, Orians CM (2006) Patterns and consequences of differential vascular sectoriality in 18 temperate tree and shrub species. Funct Ecol 20:200–206. https://doi.org/10.1111/j.1365-2435.2006.01101.x
Zhang JL, George E (2009) Rhizosphere effects on ion concentrations near different root zones of Norway spruce (Picea abies (L.) Karst.) and root types of Douglas-fir (Pseudotsuga menziesii L.) seedlings. Plant Soil 322:209–218. https://doi.org/10.1007/s11104-009-9909-0
Zhao JY, Dai ZW, Li SH, Kong Y (2008) Artificially-induced leaf nitrate accumulation affects photosynthesis in micropropagated apple plants with different water supply. J Horticult Sci Biotechnol 83:435–440. https://doi.org/10.1080/14620316.2008.11512403
Acknowledgements
We would like to thank Professors Piotr Karolewski and Marcin Zadworny for their critical review of the original manuscript.
Funding
This work was financially supported by the Institute of Dendrology PAS, Poland.
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by [Marian J. Giertych. The first draft of the manuscript was written by [Marian J. Giertych]. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Hans Lambers.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Giertych, M.J., Leski, T. Split-root system as a useful tool to study woody plant biology. Plant Soil 489, 89–105 (2023). https://doi.org/10.1007/s11104-023-06025-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-023-06025-3