
Abstract
Purpose
We sought to identify plant traits important for initial establishment of a globally invasive shrub, Prosopis spp. (mesquite), by quantifying morphological and ecophysiological responses of seedlings to contrasting moisture regimes. Our model plant was Prosopis velutina, a prominent invader of North American semi-desert grasslands.
Methods
Seedlings received one of three watering levels corresponding to regional ‘ambient’ (100%), ‘dry’ (−65%; 65% reduction of ambient), and ‘wet’ (+165%; 65% increase above ambient) growing season precipitation regimes. A total of 32 plant performance (e.g., biomass and growth) and functional (e.g., morphological and physiological) traits were assessed when seedlings reached 11- and 22-days old.
Results
Germination under dry conditions was high (72%) and only slightly reduced compared to ambient and wet conditions with subsequent seedling survival unaffected. High trait variation enabled early recruitment over the range of soil moisture conditions, including extreme drought. Root volume, surface area, tips, tap root length, and leaf length functional traits were top predictors of seedling performance, with fresh weight, absolute growth rate, and root mass secondarily contributing to establishment.
Conclusion
No physiological functional traits were significant in predicting performance or discriminating between moisture regimes; suggesting morphological traits for maintaining metabolic and hydraulic function are key to early establishment. Dry conditions reduced germination and slowed P. velutina seedling growth but did not prevent early establishment. Thus, recruitment of certain dryland shrubs may not be episodic with respect to above-average rainfall. Indeed, early recruitment of P. velutina and functionally similar species can potentially occur even in moisture conditions well below-average.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Woody plant encroachment is a global phenomenon whereby native and nonnative shrubs and trees displace grasses (Archer 1996; Eldridge et al. 2011; Stevens et al. 2017). When disturbances that limit the abundance of woody plants are eliminated (e.g. fire), shrub or tree species having the potential to access water and nutrient resources at deeper soil depths than shallow, adventitious-rooted grass species may be able to establish (Ward et al. 2013). This resource partitioning between herbaceous and woody plant community constituents may be achieved early in the life cycle of woody plants (Brown and Archer 1989; Weltzin and McPherson 1997). The primary determinates of increased woody cover in grasslands are debated, but are likely a combination of altered woody-grass competition and disturbance regimes (e.g., wildfire and livestock grazing/land-use) magnified or dampened by edaphic properties and climatic variables (Archer et al. 2017; Kulmatiski and Beard 2013). Barring biotic (Hale et al. 2020) and disturbance-related constraints on shrub establishment (e.g., fire), growing season precipitation may be a primary constraint on shrub recruitment in arid/semi-arid grasslands (Case et al. 2020), wherein soil moisture conditions during the first weeks the shrub life cycle impact recruitment probabilities (Woods et al. 2014).
Plant functional traits are influenced by growing season precipitation, but knowledge of which specific trait(s) are key for determining performance at a given life-history stage is limited. Inferences emerging from studies of plants at adult stages are suspect in that they are not necessarily germane in weeks following seedling emergence (Grubb 1977). Identification of a set of “key traits” governing plant performance at the seedling stage positions us to achieve a more precise understanding of factors controlling plant recruitment. Such knowledge would also enable us to prioritize which, among the myriad of possible measurements, to focus on in controlled environment and field studies. Furthermore, elucidation of the key traits germane to seedling establishment would strengthen inter-comparisons among studies of invasive and weedy species of global interest for which information is lacking. For example, invasive alien plant species may or may not exhibit higher performance-related trait values (e.g., growth rate, above/belowground biomass, size, and fitness) compared to non-invasives, depending on the ecosystem (temperate vs tropical) and growth form (Van Kleunen et al. 2010).
Fast-growing, long-lived species that escape their native ranges can become global invaders with lasting negative consequences on economies, agricultural production, and biodiversity (Shackleton et al. 2014). Such species in grasslands are often woody plants (e.g., shrubs, trees) that alter ecosystem structure and function and are difficult and costly to manage once established (Archer et al. 2017). A global review of woody plant flora estimated 0.5–0.7% of tree and shrub species are current invaders outside their native ranges, but a question still remains regarding the potential unique traits that make for an alien woody invader (Richardson and Rejmánek 2011). Identifying the leading morphological and physiological traits as they relate to either function and/or performance of an aggressive, successful grassland shrub invader could provide mechanistic insights into future invasion potentials. Common garden studies suggest traits such as rapid above- and belowground growth and high leaf area are positively correlated with invasion potential (Van Kleunen et al. 2010; Reichmann et al. 2016). However, knowledge of an invasive’s trait response to fluctuating environmental conditions may be more informative for evaluating invasion probabilities under variable, non-optimal growing conditions that typify real-world settings (Milbau and Stout 2008; Leffler et al. 2014). Here, we examine 32 shrub seedling traits of a known grassland invader to assess their relative importance in determining seedling success during the earliest phases of establishment under contrasting watering regimes.
Germination, emergence, and early establishment are typically “bottlenecks” in the plant lifecycle (Bowers et al. 2004; Leck et al. 2008). Shrub recruitment in arid and semi-arid environments has often been presumed to be “episodic” (Crisp 1978). Episodic, or rare but recurring, recruitment events are characterized by periods of above-average amounts and frequencies of rainfall that produce a large number of recruits, especially of long-lived dryland species with low fecundity (Walker 1993). There is, however, debate as to the importance of episodic recruitment (Brown and Archer 1999; Meyer and Pendleton 2005; Staver et al. 2007), with models and field data suggesting that both continuous and episodic recruitment may be at play (Watson et al. 1997a, b; Wiegand et al. 2004). Narrowing our focus to the traits most critical to shrub establishment will allow for improved modeling of community dynamics (Gherardi and Sala 2015). In addition, knowledge of the functional trait responses to abiotic conditions as they relate to performance can be used to predict which species and under what conditions shrub recruitment is likely to occur. This is an important contrast to examining woody encroachment or plant community dynamics via structural and functional traits ex post facto (a la D’Odorico et al. 2012), since plant attributes conferring success during the seedling establishment phase may differ substantively from those associated with the success of adult plants (Grubb 1977; Butterfield and Briggs 2011). Seedling establishment studies have historically focused on aboveground morphological functional traits (e.g., height), because physiological functional (e.g., water potential) and performance traits (e.g., total biomass) are more difficult to quantify and often involve destructive sampling. However, inclusion of such traits may provide a more complete understanding of the relative importance of the plant functional traits most germane to plant performance and recruitment in water limited systems (Harrison and LaForgia 2019). Indeed, recent work evaluating both above- and belowground functional trait variation with time, seedling age (e.g., ontogeny), and performance under altered environments on grasses and herbaceous dicots (Garbowski et al. 2021; Larson et al. 2021; Havrilla et al. 2021), suggests this would be a productive pursuit with shrub lifeforms.
Here, we quantify changes in the morphological and physiological responses of above- and belowground shrub seedling traits under contrasting watering regimes while controlling for factors such as herbivory, competition, etc. that might otherwise confound interpretations and obscure mechanistic insights (Hairston 1989). Our aim was to identify shrub seedling traits most critical to establishment and performance, and experimentally assess the potential role of soil moisture availability in constraining or promoting shrub establishment. Specifically, we asked:
-
Which traits of an aggressive, successful shrub invader of grasslands are most critical to early establishment?
-
Is shrub establishment confined to conditions of unusually high soil moisture or can it potentially occur in moderate or even extremely dry moisture regimes?
We addressed these questions using a member of the shrub genus Prosopis (commonly known as ‘mesquite’), which is among the world’s most notorious woody invasive plant taxa (Shackleton et al. 2014). Evaluation of functional and performance trait responses under contrasting moisture regimes additionally informs the probability of shrub recruitment beyond Prosopis’ native ranges. Introductions and weedy expansions of Prosopis species have been documented across the southern Great Plains of North American and Southwestern desert grasslands, in the drylands of South America (e.g., Argentina), in Africa (e.g., South Africa, Ethiopia, Kenya, and Sudan), Australia, and India (Fisher 1977; van Klinken et al. 2006; Fernández et al. 2009; Kumar and Mathur 2014). Purposely introduced for erosion control in some countries, Prosopis has also been found to provide beneficial services, such as forage for browsers, nectar for pollinators, flour from seeds/pods for bread, and fuelwood/charcoal for heating homes (Patnaik et al. 2017). Despite these services to global communities, Prosopis species are infamously difficult and costly to control/eradicate once established (Archer et al. 2011; Ding and Eldridge 2019), and their rapid growth, high seed production, and ready dispersal by rodents and livestock allows Prosopis to expand rapidly from initial introduction sites (Ayanu et al. 2015). Proliferation of Prosopis has been linked to losses of grassland endemic plants and animals as well as reductions in streamflow, groundwater recharge, and forage/habitat for wildlife and domesticated livestock (Archer and Predick 2014).
The ability for Prosopis to gain a foothold in climatically diverse regions may, in part, reflect selection for traits enabling seedling establishment across a wide range of soil moisture conditions. Functional traits such as rapid tap root elongation and increasing root surface area during early development could improve the ability of seedlings to garner scarce soil water and nutrient resources and reduce competition with fibrous, shallow-rooted grasses (Ward et al. 2013), ostensibly allowing Prosopis to successfully establish and maintain performance under relatively stressful conditions. Recent work by Larson et al. (2020) found annual species to exhibit high root trait variation in response to drought and noted the importance of growth rate and biomass allocation (e.g., performance) trade-offs for survival. Conversely, under benign or resource-abundant conditions, Prosopis seedlings may alter resource allocation to support rapid aboveground development to more quickly escape disturbances such as fire and browsing (Archer 1993). Combining multiple performance (e.g., growth rate, biomass) and functional (e.g., specific leaf area, plant height, etc.) traits across tissue and whole-plant scales may help to elucidate specific strategies for Prosopis to cope with the substantial variation in soil water availability occurring in drylands (see Fig. 1 in Volaire 2018). With this mind, our experiment was conducted using a model Prosopis species, P. velutina Woot. (velvet mesquite), a prominent native woody invader of North American desert grasslands whose abundance has increased markedly since the early 1900s (McClaran 2003). Insights into how contrasting moisture regimes influence shrub seedling trait expression would allow for the assessment of the growing season conditions underpinning the historic proliferation of this invasive shrub, while providing mechanistic insights into the circumstances and/or functional traits needed for recruitment and maintaining performance under future climates (e.g., McGill et al. 2006).
Materials and methods
Experimental design
Greenhouse trials were conducted from May 14 – June 5, 2018 at the University of Arizona Biochemistry Greenhouse A Facility in Tucson, AZ USA. Mean (± SE) daily maximum (45 ± 0.26 °C) and minimum (22 ± 0.13 °C) room temperatures were controlled to approximate the 2017 July weather conditions and the nearby Santa Rita Experimental Range (SRER), where rain events >0.5 mm occurred on 22 of 25 days (Fig. S1). Hourly temperature (o C) and light intensity (lux) measurements (ambient/incoming levels and at opposite ends of the greenhouse room) were monitored with HOBO Pendant® sensors. Solar noon photosynthetically active radiation (PAR; Line Quantum Sensor LI-191R, LI-COR Inc., Lincoln, NE) inside the greenhouse on clear days averaged 1438 ± 17 W/m2 and did not significantly differ from incoming PAR (t (18) = −0.54, p = 0.598) over the 3-week trial.
P. velutina seed pods were hand-collected from ~10 adult trees within a 5-ha area in October 2017 on the SRER (see Soils subsection) and dried at 50 °C. Seeds were subsequently manually extracted from pods, scarified using sandpaper (Tschirley and Martin 1960), and stored at room temperature. Germination was 100% (n = 200 seeds: 4 trials of 50 seeds each on moist filter paper at room temperature). P. velutina seedlings were grown from these seeds in 35.6 cm by 6.9 cm containers (Deepots™ D60L, Stuewe and Sons, Inc.) filled with homogenized field-collected soil from the 5–20 cm depth then topped with surface (0–5 cm) soil (see Soils section below). Seedling containers (total n = 60; 20 pots per moisture treatment) were organized in a completely randomized design (CRD) to accommodate the degree of replication and low number of treatments (Gomez and Gomez 1984). CRD also allowed for adjacency of pots receiving the same treatments and minimized potential effects of within-greenhouse variation in climate, lighting, etc. that might influence seedling performance. Three scarified seeds were pressed into the surface soil of each pot. Germination was recorded and seedlings were then thinned to one per pot. Additional pots (2 per moisture treatment) were instrumented to quantify soil moisture and temperature at 5 cm and 15 cm depths (HOBO Onset EC-5 Soil Moisture Smart Sensors and HOBO Pendant® Temperature sensors).
Treatments consisted of control/ambient (8 mm/day), dry (3 mm/day), and wet (11 mm/day) moisture regimes (hereafter “PPTx” for expediency). The 3 mm/day (hereafter ‘dry’) and 11 mm/day (hereafter ‘wet’) PPTx treatments are representative growing season precipitation (PPT) amounts of extreme drought and extremely wet years (≤ 10th percentile and ≥ 90th percentile of 100-year record, respectively) (Knapp et al. 2015). The PPTx’s aimed to simulate the July start of the 2017 monsoon season rainfall total (190 mm from July 9th – August 3rd) and frequency of 22 rain days with events >0.5 mm over a 25 day period measured at the ~3-ha SRER Desert Grassland Enclosure where complimentary field seedling establishment trials were being conducted (Fig. S1). Rainfall at this location was quantified with two field-based HOBO Onset RG-2 M Rain Gauge systems (Onset Computer Corporation, Bourne, MA). July PPT was chosen, as it corresponds with the historical timing for the start of the southern Arizona monsoon season (Grantz et al. 2007).
To put our greenhouse watering regime into a broader spatiotemporal context, landscape-scale patterns of PPT were evaluated using daily measurements (1975–2020) from seven rain gauges distributed across the ~21,000-ha SRER (dataset history provided in Goodrich et al. 2008). The July 2017 frequency of consecutive wet days (CWD) with at least one event ≥0.5 mm ranked in the 95th percentile of these records with 20 CWD (out of 31 possible). The frequency of CWD occurring in July 2017 also occurred in 1990. Only 1999 and 2001 had a higher frequency of rain days (21/31 days each year). Total PPT in July 2017 across the SRER (106 mm) ranked in the top 85th percentile with six years since 1975 (1981, 1984, 1988, 1990, 1998, and 2008) receiving more PPT. The July 2017 PPT and CWD at our field-based Desert Grassland Enclosure were elevated compared to the entire SRER (190 mm vs 106 mm; CWD = 22/25 vs 20/31) in the 1975–2020 record. Thus, while our greenhouse moisture treatments represented an extreme wet year (≥ 90th percentile), similar rainfall amount and event frequency conditions have occurred historically. Yearly July rainfall percentiles were calculated from data provided by the USDA Agricultural Research Service Southwest Watershed Resource Center: Data Access Project (https://www.tucson.ars.ag.gov/dap/).
Soils
Soils used in greenhouse trials were obtained from the Santa Rita Experimental Range (SRER) Desert Grassland Enclosure (31° 47′ 2.77“N, -110° 51’ 33.08”W) located 45 km south of Tucson, AZ USA. Field-collected soil was used to try and best match the soil conditions (e.g., nutrient content and texture) experienced by P. velutina seedlings in Sonoran grasslands. The Enclosure has a mean (1895–2017) annual precipitation of 420 mm (Standard Precipitation Index Explorer Tool v2.0 [https://uaclimateextension.shinyapps.io/SPItool/]) and has been excluded from livestock grazing for ≥80 years (Pease et al. 2006). The alluvial, Holocene-age soils of the Enclosure have been characterized as fine, mixed, superactive, thermic Typic Haplargids, bordering on Ustic Haplargids (Breckenfeld and Robinett 1998; Pease et al. 2006). Surface (0–5 cm) soil texture is loamy sand with 15–20% gravels (A horizon) giving way to subsurface (30–70 cm) very gravelly clay loams with a distinct brown-red argillic (Bt) horizon. See McClaran et al. 2003 for additional details on climate, vegetation, soil, and management history of the SRER.
Soils from the 0–5 cm and 5–20 cm depths were characterized and analyzed for texture, pH, and chemical constituents (Motzz Laboratory Inc., Phoenix, AZ). The 0–5 cm and 5–20 cm depth ranges were used based on field-observed differences in texture between the surface and subsurface soils at 5 cm. Bulk soils from each depth were homogenized, air-dried, and stored indoors. Residual soil volumetric water content was estimated using a soil water retention model based on soil sand, silt, and clay content with field-collected bulk density (van Genuchten 1980).
Seedling traits
Seedlings were grown in their respective PPTx for 11 days (e.g., halfway point of 22 rain days simulated from the 2017 monsoon growing season), at which time 10 seedlings/PPTx were randomly selected, assessed for functional and performance trait measurements and then harvested. Remaining seedlings were grown for an additional 11 days, at which time their traits were assessed.
Specific functional and performance traits (Violle et al. 2007) are delineated in Table 1. The mid-point (11-day) and final (22-day) seedling functional and performance traits were measured according to Perez-Harguindeguy et al. (2013). P. velutina has pinnately compound leaves, thus individual leaf measurements included leaflets, the rachis, and petiole. The aboveground traits quantified included seedling height; number of leaves; leaflets/leaf; leaf length; number of thorns; stem lignification height (as indicated by visual inspection of stem browning and toughening [Aoyama et al. 2001; Malavasi et al. 2016]); cotyledon presence/absence; leaf area; seedling/shoot fresh weight and dry mass; and dry leaf mass. Harvested pots were cut open and roots carefully separated from soil by gentle handwashing. Belowground measurements included coarse (≥ 0.5 mm diameter) and fine (< 0.5 mm diameter) root length and number, coarse root diameter, and total dry mass. Leaf area and additional root metrics (e.g., total volume and length, surface area, crossings, forks, and tips) were obtained from high resolution scanner (Model STD4800) images and WinFolia™ and WinRhizo™ image analysis software (Regent Inst. Quebec City, Canada). Mass data are based on material oven-dried 48 h at 70 °C. Root to shoot ratios (R:S) were based on dry mass, and specific leaf area was computed as total leaf area/leaf dry mass (m2/g). Absolute growth rate (AGR) was calculated as:
(Swanborough and Westoby 1996; Paine et al. 2012), where mean (± SE) seed mass (seed coat removed) was 27.0 ± 0.65 mg (n = 30). Equation 1 ensured treatment influences on seedling growth rates were not confounded by differences in seed mass (Rees et al. 2010). These metrics were chosen to enable potential comparisons with other experiments in the literature (Vilela and Ravetta 2001; Resco et al. 2009; Woods et al. 2014).
Seedling physiology
Physiological functional traits quantified at the mid-point (11th) and final (22nd) day of the experiment included midday (1100–1300) net photosynthesis/assimilation (Anet; μmol CO2 m−2 s−1), transpiration (E; mmol H2O m−2 s−1), and stomatal conductance (gs; mol H2O m−2 s−1) using a portable gas exchange system (LI-6400, LI-COR Inc., Lincoln, NE). True leaves used for Anet determinations were marked and pressed for leaf area determination (WinFolia™), and a pressure chamber (PMS Model 1505D) was used to quantify mid-day xylem water potential (Ψp; -MPa) on seedling shoots (Turner 1988) prior to drying and weighing. Leaves, shoots, and roots used in physiological measurements were retained for the mass-based calculations (e.g., AGR, above- and belowground biomass, etc.).
Statistical analysis
All analyses were performed in R v4.0.0 (R Core Team 2020). Soil chemistry, texture, and classification variables for the 0–5 cm and 5–20 cm depths were compared via simple paired t-tests at the α ≤ 0.05 level. Plant traits were preliminarily evaluated for normality (Shapiro-Wilk tests and visualization) and multicollinearity (based on high Pearson correlations; e.g., r ≤ −0.90 and/or r ≥ 0.90), as many of the measured traits were ostensibly correlated (e.g., root surface area and total root mass; Fig. S2) (Graham 2003). Since a goal of this work was to highlight the “key” traits out of all those measured that were most critical to P. velutina early establishment, all traits measured were retained for additional analyses following our initial evaluation. Shapiro-Wilk multivariate normality (W = 0.942, p = 0.006) was tested using ‘mshapiro.test’ in the mvnormtest package (Jarek 2012). We initially evaluated PPTx and seedling age/sampling period (Day 11 or Day 22) as dependent variables for all seedling functional and performance traits by a two-way multivariate ANOVA (MANOVA), where individual seedling variables (Table S2) were assessed with two-way ANOVAs using R ‘manova’ and ‘aov’ functions, respectively. Due to the large number of variables (n = 32), we calculated Pearson correlations between performance (n = 6; Table 1) and functional traits (n = 26) for each sampling period to ascertain the functional traits most related to performance for each age class. Traits were standardized prior to analysis to allow measurements to be on comparable scales, and correlations were calculated using the “corr.test” and “corr.p” functions with Holm adjustment for multiple comparisons in the R ‘psych’ package (Revelle 2015). Functional traits significantly (p ≤ 0.05) correlated with ≥4 performance traits on Day 11 and Day 22 were then used for multiple linear regression modeling. Using a composite variable for performance (Table 1) as the dependent variable and watering treatment and seedling age as covariates, we sought to additionally ascertain the combination of functional traits that best predicted (e.g., highest coefficient of determination, R2) seedling performance. The top five (out of 26) functional traits used in the regression analysis are highlighted in the Results and Discussion.
MANOVA results were assessed using the Pillai’s Trace statistic because of its lack of sensitivity to multivariate non-normality (Adeleke et al. 2015). Parametric and nonparametric MANOVA (‘adonis2’ in the vegan package) tests exhibited similar trends and seedling metrics remained non-normal following transformation. Parametric statistics are reported, as they are deemed more robust and better preserve statistical power than nonparametric tests (Finch 2005). Significant (p ≤ 0.05) ANOVAs were followed by post-hoc comparisons using Tukey’s HSD tests with ‘HSD.test’ in the agricolae R package (Mendiburu 2020). Two-way ANOVA analyses were performed on unscaled biomass (g), but descriptive statistics and figures present scaled biomass values (mg).
We also evaluated seedling trait responses to watering regime using a multivariate Linear Discriminant Analysis (LDA) to explore seedling trait variation in response to our watering treatments. A supervised LDA technique was chosen over an unsupervised Principal Components Analysis (PCA) due to (i) our a priori knowledge of greenhouse watering treatments and (ii) the large number of predictor variables (e.g., traits) compared to classes (watering treatments) (James et al. 2013). Furthermore, LDA helped to maximize treatment separation (Anowar et al. 2021) between our watering treatment categories (e.g., ambient, drought, and wet) to reveal the subset of traits, be they related to performance or functional, that contributed to the class separation. Results from a PCA are presented in the Supplemental section (Fig. S3-S6), but we focus on the LDA in the Results and Discussion. Briefly, the grouping of seedlings by watering treatment in the PCA were not maximized as with an LDA approach, where an understanding of which traits contribute the most to survival under altered water regimes may inform future work on Prosopis seedling establishment probabilities under different climate regimes. The top five (out of 32 measured) traits are highlighted in the Results and Discussion. Each variable was scaled and mean-centered before analysis to meet LDA assumptions. Variable preprocessing was completed with the R caret package (Kuhn et al. 2019); LDA was performed using MASS (Venables and Ripley 2002).
Results
Soil moisture and characterization
Soil volumetric water content (VWC) was 3.6 ± 0.47% when watering treatments began. VWC increased rapidly in the ambient and wet PPTx’s (e.g., within 1 day at 5 cm and 3 days at 20 cm depths) and reach higher maximum levels (5 cm: ambient = 22.3 ± 0.86%, wet = 25.0 ± 0.66%; 20 cm: ambient = 26.9 ± 1.08%, wet = 24.6 ± 0.76%) compared to soils in the dry PPTx (5 cm: 17.5 ± 0.99%; 20 cm: 8.0 ± 1.77%) (Fig. 1). Residual soil water content (θr) was highest in the sub-surface (4.7 ± 0.07%) compared to the surface (4.0 ± 0.12%) soils (Table S1; p ≤ 0.05). Gravimetric soil moisture at both depths was consistently lowest in the dry PPTx and comparable in ambient and wet PPTxs (Table S2).

Mean soil volumetric water content (%) at 5 cm and 20 cm depths over the experimental period for contrasting watering regimes (solid lines; ambient = grey; dry = dark orange = −65% of ambient, wet = blue = 165% of ambient). Dashed lines bounding shading denote ± standard error of the mean. Boxes on the x-axis denote 11- and 22-day samplings
The classification (Sandy Loam) and clay content of 0–5 cm and 5–20 cm soils were comparable across PPTx’s (Table S1). Surface soil (0–5 cm) had elevated fertility attributes (e.g., K, nitrate, and phosphate) compared to that of the deeper soil, whereas pH, cation exchange capacity, Ca and Mg were highest in the sub-surface soils.
Seed germination and seedling survival
Germination differed by watering treatment (F2,57 = 9.65, p = 0.000246) with dry PPTx seedlings experiencing a ~ 25% reduction compared to ambient and wet seedlings (71.7 ± 7.0% vs. 96.7 ± 2.3% and 96.7 ± 3.3%, respectively). All emerging seedlings survived for the duration of the study. Multivariate (two-way MANOVA) trait space was significantly affected by PPTx (Pillai’s Trace = 1.92, F18,60 = 6.68, p < 0.0001), seedling age (11- vs. 22-day) (Pillai’s Trace = 0.998; F8,30 = 116.95, p < 0.0001), and their interaction (Pillai’s Trace = 1.85; F18,60 = 3.55, p = 0.002) (Table 2). Trait-specific two-way ANOVA results are enumerated in Table 3 following removal of highly correlated (Pearson’s r ≤ −0.90 or ≥ 0.90) traits.
Functional and performance trait responses
A comprehensive list of all 32 traits and descriptive statistics are provided in Table S2. The primary traits identified by the LDA for discriminating between PPTx’s and notable functional and performance traits (Table 1) are described below and shown in Figs. 2, 3 and 4.

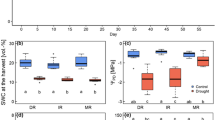
Boxplots of watering regime effects (Ambient = grey, dry = dark orange = −65% of ambient, wet = blue = 165% of ambient) on 11- (solid-lined boxes) and 22- (dashed-lined boxes) day old seedlings. Traits include seedling fresh weight (mg) (a), absolute growth rate (mg d−1) (b), root:shoot ratios (dry mass basis; unitless) (c), and dried leaf biomass (mg) (d). Central bold lines in each box represent median values, box boundaries denote the 1st (lower) and 3rd (upper) quartiles, and whiskers the 1.5 inter-quartile range; outliers are represented by solid points. Different lower-case letters denote significant differences between days and treatments (p < 0.05; Tukey’s Honest Significant Difference test)

Boxplots of 11- and 22-day old seedling responses to contrasting watering treatments (Ambient = grey, dry = dark orange = −65% of ambient, wet = blue = 165% of ambient). Variables include Anet (net CO2 assimilation) (μmol CO2 m−2 s−1) (a), transpiration (E) (mmol H2O m−2 s−1) (b), stomatal conductance (gs) (mol H2O m−2 s−1) (c), and xylem water potential (MPa) (d). See Fig. 2 legend for boxplot formatting. Different lower-case letters denote significant differences between days and treatments (p < 0.05; Tukey’s Honest Significant Difference test)

Bar plot (a) and boxplots (b-d) of seedling root traits for ambient (= grey), dry (= dark orange = −65% of ambient) and wet (= blue = 165% of ambient) watering regimes as a function of seedling age (11 vs. 22 days). Variables include number of fine roots (a), root biomass (mg) (b), tap root length (mm) (c), and root surface area (mm2) (d). Bars in (a) are the mean + SE; letters indicate TukeyHSD post-hoc significance (p < 0.05). See Fig. 2 legend for boxplot formatting
Performance traits
Shrub seedling fresh weight was significantly lower in the dry PPTxs compared to ambient on Day 11 with dry and wet PPTx seedlings being comparable (Fig. 2a). Wet and ambient PPTx seedling fresh weights were comparable by Day 22, while dry PPTx seedling fresh weight remained statistically lower. Seedling biomass (oven-dried basis) was also lower in the dry PPTx on Day 11 and Day 22 (Table S2). Absolute growth rates (AGRs) on Day 22 of the experiment were comparable (p > 0.05) for seedlings developing under ambient (8.8 ± 0.53 mg d−1) and wet (7.5 ± 0.56 mg d−1) PPTx’s and more than double that of seedlings in the dry PPTx (3.2 ± 0.88 mg d−1; Fig. 2b). R:S (0.33 ± 0.050 to 0.37 ± 0.020) were statistically comparable across PPTx’s (Table 3; F2,42 = 0.38, p = 0.69) but increased with age (Fig. 2c). A reduction in AGRs in the dry PPTx seedlings was not reflected in the R:S. Leaf biomass (oven-dried basis) was comparable across PPTx’s on Day 11, but by Day 22 dry PPTx seedlings had significantly lower leaf mass compared to ambient, but not wet PPTx seedlings (Fig. 2d; wet: 12.4 ± 1.64 mg; ambient: 13.9 ± 0.91 mg; dry: 8.2 ± 1.1 mg). Root mass of ambient (58.4 ± 3.32 mg) and wet seedlings (49.5 ± 3.25 mg) was comparable on Day 22 and higher than that of dry PPTx seedlings (26.3 ± 6.06 mg; Table S2).
Functional traits
Midday Anet, E and gs functional traits were comparable for seedlings in the ambient and wet PPTx regimes (Fig. 3a-d and Table S2) and reduced two orders of magnitude in dry seedlings on Day 11 (dry seedling Anet: 0.46 ± 1.43 μmol CO2 m−2 s−1 vs ambient: 17.30 ± 1.46 vs wet: 15.12 ± 2.24). Dry PPTx seedling Anet capacity lagged behind that of ambient and wet seedlings, but by Day 22 Anet capacities were comparable in all treatments (Fig. 3a, p > 0.05). E was significantly lower in dry PPTx seedlings at both sampling ages (Fig. 3b, p < 0.05). Mid-day xylem water potentials (Ψp) were comparable across the three watering regimes among Day 11 seedlings (Fig. 3d, p > 0.05). By Day 22, mid-day Ψp was highest in wet PPTx seedlings and lower and comparable in dry and ambient PPTx seedlings.
The number of fine roots, tap root length, and root surface area were highest under ambient watering conditions (Fig. 4a-d). Root volume, however, was suppressed in both the wet and dry PPTx compared to ambient (Table S2). Root surface area was similar between wet and ambient PPTx seedlings (Fig. 4d), but volume was significantly different across all PPTx’s and ages (Table 3). Ambient PPTx seedlings increased both root surface area and volume between Day 11 and 22, while wet PPTx seedlings accumulated relatively more surface area compared to volume (Fig. 4d, Table S2). Dry PPTx seedlings had the highest root surface area to volume ratio (dry PPTx = 5.8; ambient PPTx = 3.5; wet PPTx = 5.2; calculated from means in Table S2). Tap roots were shortest in the dry PPTx (Fig. 4c), with treatment differences in tap root length (F1,57 = 27.58, p < 0.0001), not root diameter (F1,57 = 7.17, p = 0.01) accounting for most of the variation in root volume (Table S3). Dry and wet PPTx seedlings each had less fine root development compared to ambient (Fig. 4a), but dry PPTx seedlings allocated more to coarse roots (0.85 ± 0.05 mm by Day 22) than seedlings in ambient (0.72 ± 0.03 mm) and wet (0.81 ± 0.04 mm) PPTx’s (Table S2). Overall, seedling belowground responses in the dry PPTx were much more variable by Day 22 (CV for number of fine roots: 63.5%; for tap root length: 39.4%; for root surface area: 35.9%; for root volume: 57.5%) compared to other watering regimes (Table S2; Fig. 4a-d).
Regression analysis
Twelve (out of 26) functional traits were significantly correlated (Pearson’s r; p ≤ 0.05) with the majority (e.g., >4) of performance traits on both Day 11 and Day 22 (Fig. S2), whereas the number of thorns, specific leaf area, and 5–20 cm soil GWC did not emerge as significant until Day 22. E, gs, and Anet were significantly correlated with fresh weight on Day 11 (Fig. S2), but no physiological variables significantly correlated with seedling performance at Day 22. Root volume (F11,46 = 37.69, p < 0.0001, R2 = 0.88), root surface area (F11,46 = 27.68, p < 0.0001, R2 = 0.84), root tips (F11,46 = 25.91, p < 0.0001, R2 = 0.83), leaf length (F11,46 = 25.70, p < 0.0001, R2 = 0.83), and tap root length (F11,46 = 22.52, p < 0.0001, R2 = 0.81) functional traits were the best predictors of performance in our linear models (using seedling age and watering treatment as covariates).
Linear discriminate analysis
The 32 traits in Table 1 were reduced to two discriminant axes (Fig. 5) with LD1 accounting for 85.2% of between-class (watering treatment) separation and LD2 an additional 14.8%. Full LDA model accuracy was 85.7%. Loadings for above- and belowground seedling traits on LD1 and LD2 are summarized in Fig. 6a,b. Aboveground mass had the highest absolute value loading scores (seedling fresh weight: −33.2; seedling dry mass: 22.4) followed by absolute growth rate (7.7), tap root length (−5.8), and root surface area (5.5) for LD1. Highest loading scores on PCA Principal Component 1 were similar, but also contained some belowground traits (Fig. S3b). Top discriminant variables for LD2 included fresh weight (−19.8), total root length (−14.7), root biomass (14.1), thorns (11.6), and seedling water content (9.9). Since fresh weight was the top performance trait (Table 1, Fig. 6a, b) on both LD1 and LD2 and was highly correlated with the majority of functional traits on Day 11 and Day 22 (Fig. S2), we focus largely on fresh weight and its relation to functional traits in the Discussion.

Linear Discriminant Analysis score plot of 32 seedling trait and physiological variables for watering regimes representing ambient (gray, n = 15), dry (−65% of ambient; dark orange, n = 8) and wet (165% of ambient; blue, n = 15) watering regimes. The variance explained by each axis is given in parentheses. Ellipses denote 95% confidence intervals. Smaller sample sizes in seedlings in the dry treatment reflect lower germination rates

Loading scores (unitless) of the seedling traits in Table 1 for Linear Discriminate axes 1 (a) and 2 (b) ordered by absolute value. All traits are presented with the top loading score attributed to seedling fresh weights and dried mass, followed by absolute growth rate (a) and belowground variables (e.g., tap root length, root surface area, total root length and weight) (a and b). Performance metrics itemized in Table 1 are denoted with highlighting and asterisks. Bar color symbolizes positive (black) and negative (dark orange) loadings. GWC = Gravimetric Water Content
Discussion
Precipitation influences the timing and amount of soil moisture necessary for seed germination, a fundamental first step in recruitment. We saw no evidence that germination of scarified P. velutina seeds would be episodic and confined to periods of high rainfall. Indeed, although germination was reduced in our dry PPTx, it was notably high at ~72%. Accordingly, germination bottlenecks, should they occur under field conditions for P. velutina, would likely depend on the preponderance of factors required to overcome seed coat dormancy rather than moisture availability per se. Emergence of seedlings in our dry PPTx (3 mm d−1) was delayed compared to ambient (8 mm d−1) and wet treatments (11 mm d−1) but only by ~2 days (data not shown). This short delay was, however, of little consequence for subsequent seedling performance. This is in contrast to scarified Prosopis juliflora seeds that do not produce seedlings until receiving 12 mm of water (Yoda et al. 2015) and suggests a reduced soil moisture requirement for P. velutina emergence compared to other Prosopis species.
Few greenhouse studies have combined both Prosopis seedling morphometrics and ecophysiology. Woods et al. (2014) examined shrub seedling growth rates and other performance traits under differing watering regimes but did not examine the ecophysiological functional trait responses. In contrast, Fravolini et al. (2003, 2005) focused heavily on Prosopis seedling ecophysiological sensitives to soil moisture availability, but did not document morphological adjustments. Here, we aimed to assess how morphological and physiological functional traits relate to seedling performance and identify the suite of traits most germane to Prosopis establishment.
Soil texture is well-known to influence water infiltration, percolation, and its availability to plants. Despite the high sand content of the surface soils in our experiment (78.7%), surface soil (0–5 cm) moisture content in our experiment was sufficient for maintaining seedling performance even in the dry PPTx regardless of age (Day 11 vs Day 22). Furthermore, the low levels of subsurface (20 cm) moisture in the dry PPTx was, unexpectedly, of no appreciable consequence for seedling performance (Fig. 1), as well as maintaining physiological productivity (Fig. 3a) between Day 11 and Day 22. This may be a consequence of watering frequency overriding watering amount. Although the total amount of water delivered in the ‘dry’ regime of our experiment was low, it was applied at high frequency (daily) that was based on the growing season PPT received during a concomitant field trial (Rutherford and Archer, in prep.). As it turns out, that frequency of watering ranks in the 95th percentile of records dating to 1975 (see Materials and Methods Experimental design). Projected increases in monsoon rainfall event frequency in our region (Demaria et al. 2019) bode well for bolstering future germination and early establishment potentials for P. velutina. Although the elevated subsurface soil moisture in the ambient and wet PPTx’s did promote performance with increased seedling growth rates (Fig. 2b) and root biomass (Fig. 4b) compared to dry PPTx seedlings, those enhancements did not influence survival, which was 100% across treatments. Functional trait metrics were generally comparable for ambient and wet PPTx seedlings. All-in-all, we saw little evidence that early establishment would be episodic and confined to years with above-average PPT amounts. However, the frequency of watering in our experiment was high and did not vary. Future studies should focus on quantifying the amount of PPT needed to trigger germination and elucidate the timing and frequency (i.e., temporal spacing) of subsequent small PPT events needed to sustain seedling root/shoot development and survival (e.g., performance) over longer time frames.
Root biomass of dryland shrub seedlings is highly responsive to soil water availability (Woods et al. 2011, 2014), where seedlings can allocate resources among a variety of structures (e.g., coarse, fine, tap, and lateral roots). P. velutina root traits were highly variable overall and varied by age/ontogeny in response to water availability, as has also been observed in monocots (Garbowski et al. 2021). However, whereas Garbowski et al. (2021) found little variability in root diameter, our P. velutina seedlings exhibited high PPTx average root diameter variation at Day 11 (ambient CV = 21%; dry = 8%; wet = 13%), but not Day 22 (ambient CV = 15%; dry = 14%; wet = 12%), suggesting early seedling root functional trait development may be more important for shrub establishment compared to herbaceous species. Tap root length and surface area were two of the most important belowground functional traits in determining seedling performance (e.g., growth and biomass) responses to water availability (Fig. 6a,b) and both were significantly reduced under dry conditions compared to ambient and wet (Fig. 4b-d). Seedlings in the ambient PPTx allocated more to fine roots (Fig. 4a) than did dry and wet PPTx seedlings (Table S2). Fewer fine roots in the dry and wet PPTx seedlings could ostensibly hinder performance or, in extreme cases, lead to mortality if seedlings do not compensate via tap root elongation to acquire deeper water sources (Ward et al. 2013; Priyadarshini et al. 2016). Furthermore, high root surface area to volume ratios in dry and wet PPTx seedlings compared to ambient potentially increases the efficiency of absorption of minerals and water, particularly if accompanied by root hair development (e.g., Gilroy and Jones 2000). These patterns provide a basis for further investigation with high-resolution image analysis into shrub seedling fine root development and their functional contributions to seedling establishment and survival.
Compared to three other Prosopis species, P. velutina has been found to preferentially allocate biomass to roots at the expense of stem and leaf development (Vilela and Ravetta 2001). Increasing water availably has also been found to promote lateral root development over tap root elongation in Prosopis (Yoda et al. 2012). Due to pot size constraints and emphasis on early P. velutina seedling age-states in our experiment, lateral versus non-lateral roots were difficult to differentiate. With that said, tap root length did contribute the most to PPTx total root volume (Table S3) and tap root length was also highly predictive of performance (R2 = 0.81, see Results Regression analysis). A reduction in tap root elongation coupled with a decrease in root surface area and volume may increase the risk of seedling mortality if drought conditions develop in upper soils before roots can extend to into deeper soils where moisture may be more abundant. However, the young seedlings in our experiment were well-suited to cope with the extreme range of water inputs they experienced in our PPTx regimes.
Out of 32 total traits examined, aboveground seedling fresh weight was the primary performance trait in separating and classifying the seedlings by watering regime (Fig. 6a,b). Seedlings in the dry PPTx maintained a stable internal water content (Day 11: 75% vs Day 22: 74%; Table S2), while that of seedlings in the ambient and wet treatments was less stable and decreased over time (Ambient Day 11: 78% vs Day 22: 71%; Wet Day 11: 79% vs Day 22: 70%; Table S2). This was also reflected in seedling transpiration (E), which remained low under dry conditions with large reductions by Day 22 in ambient (%∆E = −30%) and wet (%∆E = −49%) seedlings. Ambient and wet seedlings also experienced lower stomatal conductance by Day 22, unlike the dry PPTx seedlings (Fig. 3c). A similar pattern occurred with the photosynthetic rates, whereby dry seedlings increased mean Anet with age, while wet and ambient PPTx seedlings decreased (Fig. 3a). Seedling photosynthetic rates in all PPTx were comparable by Day 22 (Fig. 3a), also reflected by the lack of a significant relationship between Anet and fresh weight on Day 22 (Fig. S2). Despite seedling physiological trait influences on performance, none were amongst the top discriminant variables (Fig. 6a,b), and none emerged as significant in our linear predictive models. This suggests functional physiological trait measurements (e.g., Anet, E, and gs), which are often used as proxies for performance, may be secondary to size-based trait measurements directly related to seedling structure and morphological attributes (e.g., root volume, root surface area, root tips, tap root length, and leaf length) essential for maintaining metabolic and hydraulic functions critical for seedling establishment (e.g., von Arx et al. 2012). It should be noted that physiology- and size-based trait measurements are inherently related, wherein seedling size-based traits are an outcome of physiological activity and growth, creating a ‘chicken vs egg’ conundrum interpreting certain trait responses. Both physiology- and size-based measurements are also highly sensitive to the passage of time during the establishment phase of the plant lifecycle (Henn and Damschen 2021). The said, our data suggest that functional structural and morphological traits critical for maintenance of seedling metabolic and hydraulic function may be most reflected by the performance trait of seedling fresh weight, which has been linked to plant metabolite and enzyme concentrations (Çakmakçı et al. 2007), organ function (Llamas and Sanz 2008), and leaf photosynthetic capacity (Braun and Wild 1984) more so than is dry mass. Given the importance of Prosopis fresh weight in our analysis, future assessments of shrub traits should consider biomass on both a fresh and dry mass basis when examining seedling responses to abiotic stress.
Drylands are characterized by high levels of spatiotemporal variability in rainfall (Ahlström et al. 2015), as also seen by the site- vs landscape-level rainfall frequency differences at the SRER (see Methods Experimental design). Trait variation in response to altered abiotic conditions may be a key factor in seedling establishment in these systems, particularly if it enhances the seedling ability to regulate water use (e.g., Reynolds et al. 2018). The ability of dry PPTx seedlings to retain tissue moisture via low midday E may reflect a beneficial physiological/functional adjustment for environmental stress mediation to maintain performance. Conservation of above−/belowground biomass (e.g., performance) and physiological function (e.g., photosynthetic rate, transpiration rate, and stomatal conductance, collectively) may have enabled the dry PPTx seedlings to continue development comparable to that seen with seedlings in the ambient and wet PPTx. This is in accordance with Xu et al. (2021) who found that physiological functional trait variation, not genetic adaptation, of the cosmopolitan shrub, Dodonaea viscosa, was key to mitigating drought stress. While prolonged drought can impose metabolic and hydraulic damage and constrain seedling photosynthetic responses to subsequent PPT pulses (Resco et al. 2009), our work suggests young P. velutina seedlings have the short-term capacity to limit physiological activity (e.g., decreased Anet, E, and gs; Fig. 3a-c) and thereby maintain hydraulic function (e.g., comparable xylem water potentials in ambient and drought seedlings; Fig. 3d) and avert potential damage that might otherwise constrain physiological response to a subsequent PPT event(s).
When water is not limiting, as with seedlings in our ambient and wet PPTx treatments, above−/belowground biomass accumulation was comparable. Conversely, when water was extremely limiting (e.g., reduction of 65% compared to ambient), above−/belowground development and growth (e.g., performance) was reduced (Fig. 3b) as a possible stress tolerance strategy for maintaining an internal balance between water content and physiological activity (e.g., function). Optimal partitioning theory (Bloom et al. 1985) suggests that droughted seedlings may forego aboveground growth to prioritize belowground growth and tap root elongation to reach deeper soils where water content may be higher or more consistent (Resco et al. 2009; Zhang et al. 2019). In our study, droughted P. velutina seedlings developed above- and belowground biomass in accordance with available soil water instead of disproportionately developing longer tap roots or additional root biomass compared to aboveground biomass as optimal partitioning theory would suggest. Therefore, drought tolerant shrub seedlings may be positioned to withstand short or longer-term periods of abiotic stress by reducing the need for water resources that would be required to support increased performance/biomass. The ability to endure periods of abiotic stress, like drought, may be a functional mechanism underlying P. velutina’s ability to invade and establish in grasslands. We note that our experiments were conducted on seedlings grown in isolation. Successful recruitment in nature would also have to balance additional trade-offs that come into play when competition is a factor.
Our assessment of the performance and functional trait responses of P. velutina with respect to soil water variability provides a basis for comparative investigations into the world’s Prosopis species (e.g., P. gladulosa, P. pallida, P. chilensis, just to name a few), and would be highly beneficial for future application of this work across grasslands currently experiencing Prosopis proliferation. Additional comparative and controlled experimental studies may also elucidate more nuanced performance and functional trait relationships as in the contribution of root hairs to seedling survival during drought. In all, P. velutina seedlings were found to establish and maintain productivity even when subjected to severe drought-like conditions. Thus, Prosopis germination and early establishment can potentially occur across a wide-range of moisture regimes with recruitment not limited to periods of above-average soil moisture availability.
Conclusions
Shrub proliferation in arid/semi-arid grasslands is a global phenomenon with broad ramifications for ecosystem structure and function. The abiotic conditions and mechanisms that constrain and/or drive shrub recruitment are not well understood. Prosopis is one of the leading shrub genera of global concern due to its aggressive establishment and the management challenges it presents. In this study, the early seedling establishment phase in the life cycle of P. velultina, was assessed as a model Prosopis species. A suite of 32 performance and functional traits were examined under contrasting watering regimes to identify those most informative in identifying abiotic moisture constraints on initial establishment. We found that P. velutina seedlings have sufficient trait variation to enable recruitment over a wide range of moisture conditions, including extreme drought. The “key traits” conferring this capability that should be prioritized in future Prosopis spp. studies include the performance traits of seedling fresh weight, absolute growth rate, and root biomass and functional traits of tap root length, root surface area, root volume, root tips, and leaf length. Structural more so than physiological functional trait adjustments may enable seedlings to maintaining key metabolic and hydraulic functions needed for establishment over a wide range of moisture regimes. Germination and early establishment in our trials was found to readily occur even in moisture regimes mimicking average and well below-average rainfall. High performance and functional trait variation in response to available soil moisture may help to explain why P. velutina has readily established in grasslands over the past decades while the many other shrub species in the regional flora have not. Future investigations should consider dry day interval/drought duration effects in combination with varying rainfall event frequency/amount combinations comparing across multiple Prosopis species.
Data availability
All datasets used in the current study are accessible in an archived Zenodo repository (https://zenodo.org/badge/latestdoi/386100148).
Code availability
Code for data cleaning and analysis is provided in an archived Zenodo repository (https://zenodo.org/badge/latestdoi/386100148).
References
Adeleke BL, Yahya WB, Usman A (2015) A comparison of some test statistics for multivariate analysis of variance model with non-normal responses. J Nat Sci Res 5:1–9
Ahlström A, Raupach MR, Schurgers G et al (2015) The dominant role of semi-arid ecosystems in the trend and variability of the land CO2 sink. Science (80-) 348:895–899
Anowar F, Sadaoui S, Selim B (2021) Conceptual and empirical comparison of dimensionality reduction algorithms (PCA, KPCA, LDA, MDS, SVD, LLE, ISOMAP, LE, ICA, t-SNE). Comput Sci Rev 40:100378. https://doi.org/10.1016/j.cosrev.2021.100378
Aoyama W, Matsumura A, Tsutsumi Y, Nishida T (2001) Lignification and peroxidase in tension wood of Eucalyptus viminalis seedlings. J Wood Sci 47:419–424
Archer S (1993) Vegetation dynamics in changing environments. Rangel J 15:104–116
Archer SR (1996) Assessing and interpreting grass-woody plant dynamics. In: Hodgson J, Illius AW (eds) The ecology and management of grazing systems. CAB International, Wallingford, pp 101–134
Archer SR, Davies KW, Fulbright TE, McDaniel KC, Wilcox BP, Predick KI, and Briske DD (2011) Brush management as a rangeland conservation strategy: a critical evaluation. Conservation benefits of rangeland practices 105–170
Archer SR, Predick KI (2014) An ecosystem services perspective on brush management: Research priorities for competing land use objectives. J Ecol 102:1394–1407
Archer SR, Andersen EM, Predick KI, Schwinning S, Steidl RJ, Woods SR (2017) Woody plant encroachment: Causes and consequences. In: Briske D (eds) Rangeland systems. Springer series on environmental management. Springer, Cham. https://doi.org/10.1007/978-3-319-46709-2_2
Ayanu Y, Jentsch A, Müller-Mahn D et al (2015) Ecosystem engineer unleashed: Prosopis juliflora threatening ecosystem services? Reg Environ Chang 15:155–167
Bloom AJ, Chapin FS III, Mooney HA (1985) Resource limitation in plants-an economic analogy. Annu Rev Ecol Syst 16:363–392
Bowers JE, Turner RM, Burgess TL (2004) Temporal and spatial patterns in emergence and early survival of perennial plants in the Sonoran Desert. Plant Ecol 172:107
Braun P, Wild A (1984) The influence of brassinosteroid on growth and parameters of photosynthesis of wheat and mustard plants. J Plant Physiol 116:189–196. https://doi.org/10.1016/S0176-1617(84)80088-7
Breckenfeld DJ, Robinett D (1998) Soil and ecological sites of the Santa Rita experimental range. In: McClaran MP, Ffolliott PF, Edminster CB, tech. coords (eds) Santa Rita experimental range: 100 years (1903 to 2003) of accomplishments and contributions. USA Department of Agriculture, Forest Service, Rocky Mountain Research Station, Tucson, pp 157–165
Brown JR, Archer S (1989) Woody plant invasion of grasslands: establishment of honey mesquite (Prosopis glandulosa var. glandulosa) on sites differing in herbaceous biomass and grazing history. Oecologia 80:19–26
Brown JR, Archer S (1999) Shrub invasion of grassland: recruitment is continuous and not regulated by herbaceous biomass or density. Ecology 80:2385–2396
Butterfield BJ, Briggs JM (2011) Regeneration niche differentiates functional strategies of desert woody plant species. Oecologia 165:477–487
Çakmakçı R, Erat M, Erdoğan Ü, Dönmez MF (2007) The influence of plant growth–promoting rhizobacteria on growth and enzyme activities in wheat and spinach plants. J Plant Nutr Soil Sci 170:288–295. https://doi.org/10.1002/jpln.200625105
Case MF, Wigley BJ, Wigley-Coetsee C, Carla Staver A (2020) Could drought constrain woody encroachers in savannas? African J Range Forage Sci 37:19–29
Crisp MD (1978) Demography and survival under grazing of three Australian semi-desert shrubs. Oikos:520–528
D’Odorico P, Okin GS, Bestelmeyer BT (2012) A synthetic review of feedbacks and drivers of shrub encroachment in arid grasslands. Ecohydrology 5:520–530
Demaria EMC, Hazenberg P, Scott RL, Meles MB, Nichols M, Goodrich D (2019) Intensification of the North American Monsoon rainfall as observed from a long-term high-density gauge network. Geophys Res Lett 46:6839–6847. https://doi.org/10.1029/2019GL082461
Ding J, Eldridge DJ (2019) Contrasting global effects of woody plant removal on ecosystem structure, function and composition. Perspect Plant Ecol, Evolution and Systematics 39:125460. https://doi.org/10.1016/j.ppees.2019.125460
Eldridge DJ, Bowker MA, Maestre FT et al (2011) Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol Lett 14:709–722
Fernández OA, Gil ME, Distel RA (2009) The challenge of rangeland degradation in a temperate semiarid region of Argentina: the Caldenal. L Degrad Dev 20:431–440
Finch H (2005) Comparison of the performance of nonparametric and parametric MANOVA test statistics when assumptions are violated. Methodology 1:27–38. https://doi.org/10.1027/1614-1881.1.1.27
Fisher CE (1977) Mesquite and modern man in southwestern North America. In: Mesquite: its biology in two desert ecosystems. Dowden, Hutchinson and Ross, Stroudsburg, pp 177–188
Fravolini A, Hultine KR, Koepke DF, Williams DG (2003) The role of soil texture on mesquite water relations and response to summer precipitation. In: McClaran MP, Ffolliott PF, Edminster CB, tech. coords (eds) Santa Rita experimental range: 100 years (1903 to 2003) of accomplishments and contributions. USA Department of Agriculture, Forest Service, Rocky Mountain Research Station, Tucson, pp 125–129
Fravolini A, Hultine KR, Brugnoli E et al (2005) Precipitation pulse use by an invasive woody legume: the role of soil texture and pulse size. Oecologia 144:618–627
Garbowski M, Johnston DB, Brown CS (2021) Leaf and root traits, but not relationships among traits, vary with ontogeny in seedlings. Plant Soil 460:247–261. https://doi.org/10.1007/s11104-020-04790-z
Geber MA, Griffen LR (2003) Inheritance and natural selection on functional traits. Int J Plant Sci 164(S3):S21–S42. https://doi.org/10.1086/368233
Gherardi LA, Sala OE (2015) Enhanced precipitation variability decreases grass- and increases shrub-productivity. Proc Natl Acad Sci 112:12735–12740
Gilroy S, Jones DL (2000) Through form to function: root hair development and nutrient uptake. Trends Plant Sci 5:56–60
Gomez KA, Gomez AA (1984) Statistical procedures for agricultural research. Wiley, Hoboken
Goodrich DC, Keefer TO, Unkrich CL et al (2008) Long-term precipitation database, walnut gulch experimental watershed, Arizona, United States. Water Resour Res 44. https://doi.org/10.1029/2006WR005782
Graham MH (2003) Confronting multicollinearity in ecological multiple regression. Ecology 84:2809–2815. https://doi.org/10.1890/02-3114
Grantz K, Rajagopalan B, Clark M, Zagona E (2007) Seasonal shifts in the north American monsoon. J Clim 20:1923–1935
Grubb PJ (1977) The maintenance of species-richness in plant communities: the importance of the regeneration niche. Biol Rev 52:107–145
Hairston NG (1989) Ecological experiments: purpose, design and execution. Cambridge University Press, Cambridge
Hale SL, Koprowski JL, Archer SR (2020) Black-tailed prairie dog (Cynomys ludovicianus) reintroduction can limit woody plant proliferation in grasslands. Front Ecol Evol 8:233
Harrison S, LaForgia M (2019) Seedling traits predict drought-induced mortality linked to diversity loss. Proc Natl Acad Sci 116:5576–5581
Havrilla CA, Munson SM, Yackulic EO, Butterfield BJ (2021) Ontogenetic trait shifts: seedlings display high trait variability during early stages of development. Funct Ecol 35:2409–2423. https://doi.org/10.1111/1365-2435.13897
Henn JJ, Damschen EI (2021) Plant age affects intraspecific variation in functional traits. Plant Ecol 222:669–680. https://doi.org/10.1007/s11258-021-01136-2
James G, Witten D, Hastie T, Tibshirani R (2013) An introduction to statistical learning: with applications in R. Springer, Berlin
Jarek S (2012) Mvnormtest: normality test for multivariate variables. R Packag version:01–09
Knapp AK, Hoover DL, Wilcox KR et al (2015) Characterizing differences in precipitation regimes of extreme wet and dry years: implications for climate change experiments. Glob Chang Biol 21:2624–2633
Kuhn M, Wing J, Weston S, et al (2019) caret: Classification and Regression Training. R package version 6.0–84
Kulmatiski A, Beard KH (2013) Woody plant encroachment facilitated by increased precipitation intensity. Nat Clim Chang 3:833
Kumar S, Mathur M (2014) Impact of invasion by Prosopis juliflora on plant communities in arid grazing lands. Trop Ecol 55:33–46
Larson JE, Anacker BL, Wanous S, Funk JL (2020) Ecological strategies begin at germination: traits, plasticity and survival in the first 4 days of plant life. Funct Ecol 34:968–979. https://doi.org/10.1111/1365-2435.13543
Larson JE, Ebinger KR, Suding KN (2021) Water the odds? Spring rainfall and emergence-related seed traits drive plant recruitment. Oikos 130:1665–1678. https://doi.org/10.1111/oik.08638
Leck MA, Parker VT, Simpson RL, Simpson RS (2008) Seedling ecology and evolution. Cambridge University Press, Cambridge
Leffler AJ, James JJ, Monaco TA, Sheley RL (2014) A new perspective on trait differences between native and invasive exotic plants. Ecology 95:298–305. https://doi.org/10.1890/13-0102.1
Llamas A, Sanz A (2008) Organ-distinctive changes in respiration rates of rice plants under nickel stress. Plant Growth Regul 54:63–69. https://doi.org/10.1007/s10725-007-9229-4
Malavasi UC, Davis AS, de Matos Malavasi M (2016) Lignin in woody plants under water stress: a review. Floresta e Ambient 23:589–597
McClaran MP (2003) A century of vegetation change on the Santa Rita experimental range. In: McClaran MP, Ffolliott PF, Edminster CB, tech. coords. (eds) Santa Rita experimental range: 100 years (1903 to 2003) of accomplishments and contributions. US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Tucson
McClaran MP, Ffolliott PF, Edminster CB (2003) Santa Rita experimental range: 100 years (1903 to 2003) of accomplishments and contributions. US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Tucson
McGill BJ, Enquist BJ, Weiher E, Westoby M (2006) Rebuilding community ecology from functional traits. Trends Ecol Evol 21:178–185
Mendiburu F (2020) Agricolae: Statistical procedures for agricultural research. R package Ver. 1.3–2
Meyer SE, Pendleton BK (2005) Factors affecting seed germination and seedling establishment of a long-lived desert shrub (Coleogyne ramosissima: Rosaceae). Plant Ecol 178:171–187
Milbau A, Stout JC (2008) Factors associated with alien plants transitioning from casual, to naturalized, to invasive. Conserv Biol 22:308–317
Paine CET, Marthews TR, Vogt DR et al (2012) How to fit nonlinear plant growth models and calculate growth rates: an update for ecologists. Methods Ecol Evol 3:245–256
Patnaik P, Abbasi T, Abbasi SA (2017) Prosopis (Prosopis juliflora): blessing and bane. Trop Ecol 58:455–483
Pease S, Ffolliott PF, Gottfried GJ, DeBano LF (2006) Mesquite removal and mulching impacts on herbage production on a semidesert grass-shrub rangeland. US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Tucson
Perez-Harguindeguy N, Diaz S, Garnier E et al (2013) New handbook for standardised measurement of plant functional traits worldwide. Aust J Bot 61:167–234
Priyadarshini KVR, Prins HHT, de Bie S et al (2016) Seasonality of hydraulic redistribution by trees to grasses and changes in their water-source use that change tree–grass interactions. Ecohydrology 9:218–228. https://doi.org/10.1002/eco.1624
R Core Team (2020) R: A language and environment for statistical ## computing. R foundation for statistical computing, Vienna, Austria. https://www.R-project.org/
Rees M, Osborne CP, Woodward FI et al (2010) Partitioning the components of relative growth rate: how important is plant size variation? Am Nat 176:E152–E161
Reichmann LG, Schwinning S, Polley HW, Fay PA (2016) Traits of an invasive grass conferring an early growth advantage over native grasses. J Plant Ecol 9:672–681. https://doi.org/10.1093/jpe/rtw014
Resco V, Ewers BE, Sun W et al (2009) Drought-induced hydraulic limitations constrain leaf gas exchange recovery after precipitation pulses in the C3 woody legume, Prosopis velutina. New Phytol 181:672–682
Revelle W (2015) Package ‘psych.’. Compr R Arch Netw 337:338
Reynolds VA, Anderegg LDL, Loy X et al (2018) Unexpected drought resistance strategies in seedlings of four Brachychiton species. Tree Physiol 38:664–677
Richardson DM, Rejmánek M (2011) Trees and shrubs as invasive alien species – a global review. Divers Distrib 17:788–809. https://doi.org/10.1111/j.1472-4642.2011.00782.x
Schwinning S, Ehleringer JR (2001) Water use trade-offs and optimal adaptations to pulse-driven arid ecosystems. J Ecol 89:464–480. https://doi.org/10.1046/j.1365-2745.2001.00576.x
Shackleton RT, Le Maitre DC, Pasiecznik NM, Richardson DM (2014) Prosopis: A global assessment of the biogeography, benefits, impacts and management of one of the world’s worst woody invasive plant taxa. AoB plants 6. https://doi.org/10.1093/aobpla/plu027
Staver AC, Bond WJ, February EC (2007) Continuous v. episodic recruitment in Acacia karroo in Hluhluwe-Imfolozi Park: implications for understanding savanna structure and dynamics. South African J Bot 73:314
Stevens N, Lehmann CER, Murphy BP, Durigan G (2017) Savanna woody encroachment is widespread across three continents. Glob Chang Biol 23:235–244
Swanborough P, Westoby M (1996) Seedling relative growth rate and its components in relation to seed size: Phylogenetically independent contrasts. Funct Ecol 10(2):176–184. https://doi.org/10.2307/2389841
Tschirley FH, Martin SC (1960) Germination and longevity of velvet mesquite seed in soil. J Range Manag 13:94–97
Turner NC (1988) Measurement of plant water status by the pressure chamber technique. Irrig Sci 9:289–308
van Genuchten M (1980) A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci Soc Am J 44:892–898
Van Kleunen M, Weber E, Fischer M (2010) A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol Lett 13:235–245. https://doi.org/10.1111/j.1461-0248.2009.01418.x
van Klinken RD, Graham J, Flack LK (2006) Population ecology of hybrid mesquite (Prosopis species) in Western Australia: how does it differ from native range invasions and what are the implications for impacts and management? Biol Invasions 8:727–741
Venables WN, Ripley BD (2002) Modern applied statistics with S, Fourth. Springer, New York
Vilela AE, Ravetta DA (2001) The effect of seed scarification and soil-media on germination, growth, storage, and survival of seedlings of five species of Prosopis L. (Mimosaceae). J Arid Environ 48:171–184
Violle C, Navas M-L, Vile D et al (2007) Let the concept of trait be functional! Oikos 116:882–892. https://doi.org/10.1111/j.0030-1299.2007.15559.x
Volaire F (2018) A unified framework of plant adaptive strategies to drought: crossing scales and disciplines. Glob Chang Biol 24:2929–2938. https://doi.org/10.1111/gcb.14062
von Arx G, Archer SR, Hughes MK (2012) Long-term functional plasticity in plant hydraulic architecture in response to supplemental moisture. Ann Bot 109:1091–1100. https://doi.org/10.1093/aob/mcs030
Walker BH (1993) Rangeland ecology: understanding and managing change. Ambio 22:80–87
Ward D, Wiegand K, Getzin S (2013) Walter’s two-layer hypothesis revisited: back to the roots! Oecologia 172:617–630
Watson IW, Westoby M, Holm AM (1997a) Demography of two shrub species from an arid grazed ecosystem in Western Australia 1983-93. J Ecol 85:815–832
Watson IW, Westoby M, Holm AM (1997b) Continuous and episodic components of demographic change in arid zone shrubs: models of two Eremophila species from western Australia compared with published data on other species. J Ecol 85:833–846
Weltzin JF, McPherson GR (1997) Spatial and temporal soil moisture resource partitioning by trees and grasses in a temperate savanna, Arizona, USA. Oecologia 112:156–164
Wiegand K, Jeltsch F, Ward D (2004) Minimum recruitment frequency in plants with episodic recruitment. Oecologia 141:363
Woods SR, Archer SR, Schwinning S (2011) Early taproot development of a xeric shrub (Larrea tridentata) is optimized within a narrow range of soil moisture. Plant Ecol 212:507–517
Woods SR, Archer SR, Schwinning S (2014) Seedling responses to water pulses in shrubs with contrasting histories of grassland encroachment. PLoS One 9:e87278
Xu GQ, Farrell C, Arndt SK (2022) Climate of origin has no influence on drought adaptive traits and the drought responses of a widely distributed polymorphic shrub. Tree Physiology 42(1):86–98. https://doi.org/10.1093/treephys/tpab085
Yoda K, ELbasit MA, Hoshino B et al (2012) Root system development of Prosopis seedlings under different soil moisture conditions. J Arid L Stud 22:13–16
Yoda K, Tsuji W, Inoue T et al (2015) Evaluation of the effect of a rain pulse on the initial growth of Prosopis seedlings. Arid L Res Manag 29:210–221
Zhang B, Cadotte MW, Chen S et al (2019) Plants alter their vertical root distribution rather than biomass allocation in response to changing precipitation. Ecology 100:e02828
Acknowledgements
We extend thanks to two anonymous reviewers, section editor François Teste, Dr. David L. Hoover and Dr. Katherine I. Predick for their critical review and constructive comments on earlier versions of this manuscript, and to Sara Amiot for assistance with data collection.
Funding
This study was supported by the US Department of Agriculture National Institute of Food and Agriculture (USDA-NIFA Award 2016–67013-24932) and Arizona Agricultural Experiment Station ARZT-1361610-H12–223.
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study conception and design. WAR performed the data collection, statistical analysis, and initial material preparation. The manuscript was written and edited by WAR and SRA. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflicts of interest/competing interests
The authors declare no conflicts of interests/competing interests.
Additional information
Responsible Editor: François Teste.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 1.60 mb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rutherford, W.A., Archer, S.R. Trait responses of a grassland shrub invader to altered moisture regimes. Plant Soil 482, 141–161 (2023). https://doi.org/10.1007/s11104-022-05678-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-022-05678-w