
Abstract
Background and aims
Soil heterogeneity has been found to promote plant invasion, as many naturalized alien species benefit more from nutrient-rich patches than native species do. However, it remains unclear whether naturalized alien species also benefit more from growing on patch boundaries than from growing in the patch centres of heterogeneous soils.
Methods
We experimentally tested whether the planting position changes the effect of soil heterogeneity on the growth of native and naturalized alien plants. We created pots with heterogeneous soils consisting of patches of sand, peat-based potting substrate (peat) and a 1:1 sand-peat mixture, and homogenous soils consisting of the mixture only. In each pot, we planted three plants of one of five native or five naturalized alien species, either on the boundaries between or in the centres of the soil patches.
Results
Individual plant growth increased when the plant had direct access to the peat patches. This positive effect of peat on plant growth tended to be more pronounced for the naturalized alien species than for the natives. The populations of naturalized alien species also tended to benefit more, in terms of total biomass, from soil heterogeneity than the natives did. Belowground biomass of the plant populations, was not significantly affected by soil heterogeneity, but was larger when plants grew on the patch boundaries instead of in the centres.
Conclusion
Our study suggests that soil heterogeneity promotes the growth of naturalized alien plants over native plants. Moreover, being positioned on the boundary of strongly contrasting patches might provide benefits to plant growth.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Soil heterogeneity refers to the uneven and patchy distribution of soil types, and soil resources (e.g. nutrients, water). As resources are naturally heterogeneously distributed in the environment, soil heterogeneity is ubiquitous across spatial and temporal scales, especially in terrestrial ecosystems. Patch size and patch contrast are two important aspects of soil heterogeneity, and they can affect the root systems and growth performance of individual plants directly and indirectly (Stuefer 1996; Hutchings et al. 2003; Huang et al. 2023). When the patch size is larger than the root system and the plant is in the centre of the patch, the plant may fail to detect its heterogeneous surroundings, and therefore will not respond to the soil heterogeneity. However, when the patch size is smaller than the root system or when the plant is at the edge of a large patch or at the boundary between patches, it will likely perceive the heterogeneous surroundings (Hutchings et al. 2003).
Whether a plant will perceive soil heterogeneity and exert plastic responses will also depend on the contrast between patches, defined as the degree of difference between patches in a spatially heterogeneous environment (Kotliar and Wiens 1990; Stuefer 1996; Wijesinghe and Hutchings 1999; Hutchings et al. 2003). For example, the stoloniferous herb Glechoma hederacea selectively placed its roots into nutrient-rich patches when the contrast between patches was high, whereas this was not the case when patch contrast was low (Wijesinghe and Hutchings 1999). As patch contrast operates at the boundary between patches, these boundaries might provide unique niches for plants as they are interfaces where plants experience the properties of the different neighbouring patches (Stover and Henry 2018; Kowalski and Henry 2020). However, the potential effect of growing on patch boundaries of heterogeneous soils on plant growth, and how this might vary among species remains to be further explored.
Physical boundaries in nature are everywhere and occur across scales (Cadenasso et al. 2003). At large spatial scales, boundaries, as the border or interface of two different habitats, are clearly visible, such as roads in cities and mountains, the riparian zone between river and land, and the paths between agricultural fields. At smaller spatial scales, within the same habitat there are patches and boundaries in soil caused by animal activities. For example, many mammals dig in the soil for food and water, thereby creating small patches of soil which may differ in nutrient and water contents from the undisturbed matrix (Davies et al. 2019; Lundgren et al. 2021). In addition, large grazers were found to create spatial variation in N availability by their excreta (Mikola et al. 2009). Either way, when patches are created, heterogeneity increases, and there is a higher chance that plants will grow at the boundaries of the patches.
The direction and magnitude of a plant’s response to soil heterogeneity may depend on its life-history stage. For example, with increasing soil heterogeneity, seed germination of the grass species Lolium perenne and Elymus nutans was delayed and reduced (Liu and Hou 2021). On the other hand, spatial heterogeneity of surface soil properties was found to increase early seedling survival and establishment of the shrub species Pistacia lentiscus in semiarid field sites (Maestre et al. 2003). Plastic responses like root foraging allow plants to selectively place their roots and to proliferate in patches with high levels of resource, such as nutrients (Hodge 2004; Mommer et al. 2012) and water (Bauerle et al. 2008), but also to grow away from patches with pollutants such as microplastics (Speißer and van Kleunen 2023). Soil heterogeneity could affect growth performance as well as biomass allocation of plants (Day et al. 2003; Maestre et al. 2006; Prietzel 2020; Liu et al. 2021). In addition, soil heterogeneity may also contribute to species coexistence and biodiversity patterns (Wijesinghe et al. 2005; Gazol et al. 2013; Liu et al. 2019; Baer et al. 2020). Nevertheless, responses of plants to soil heterogeneity are species-specific (Keser et al. 2015; Tamme et al. 2016; Speißer and van Kleunen 2023). Therefore, whether soil-heterogeneity effects on plants are generally positive or negative, and whether the effects vary between certain categories of species still remains unclear.
In the last decades, studies have started to focus on the role of soil heterogeneity in plant invasion, because patchily distributed resources might increase niche availability, and thereby increase the invasibility of communities (Davis et al. 2000; Parepa et al. 2013). Furthermore, as many invasive plants are thought to have a high phenotypic plasticity and a high environmental tolerance (Richards et al. 2006; van Kleunen et al. 2010), they may benefit more from environmental heterogeneity than native plants. Indeed, Skálová et al. (2012) found that three invasive Impatiens species in Central Europe were more plastic than the native Impatiens noli-tangere L. Furthermore, naturalized clonal plants were found to benefit more from heterogeneous resource conditions than closely related native clonal species (Keser et al. 2014; Wang et al. 2017). However, even within a heterogeneous environment, plants at the patch boundaries are likely to experience more heterogeneous conditions than plants in the patch centres. Previous studies have found that roads might provide favourable disturbed habitats and dispersal routes for non-native species to spread along the roads (Christen and Matlack 2006; Iseli et al. 2023; Son et al. 2024). Agricultural field boundaries may act as refugia for grassland species and have an effect on biodiversity in managed agricultural landscapes (Smart et al. 2002), thus might also provide opportunities for non-native species to establish. Whether naturalized alien plants, compared to native plants, also benefit more from growing at small scale patch boundaries than from growing in the centres of patches remains unknown.
Here, we experimentally tested whether planting position modifies the effect of soil heterogeneity on the growth of individual plants and small populations of naturalized and native species. We grew plants of ten species that are either native or naturalized in Germany in homogeneous and heterogeneous soils, and we planted them either in the centres of the patches or at the boundaries between patches. As we had three patches per pot, we planted three individuals per pot (Fig. 1). With this experimental setting, we tested the following hypotheses:
-
(1) In heterogeneous soils, individual plants benefit from growing on the patch boundaries as the roots have direct access to both patches, which may improve the precision of selective root placement. (2) Naturalized alien plant individuals in general exert high phenotypic plasticity and fast growth, and therefore will benefit more from growing on the patch boundaries than native plant individuals. (3) Overall, soil heterogeneity may increase the resource variability and niche availability, and thereby promote plant growth. (4) The beneficial soil-heterogeneity effect is stronger for naturalized alien than for native plants as the naturalized alien plants may take more advantage of variable environments.

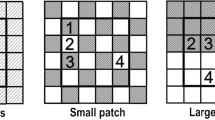
Overview of the experiment. Illustrations of soil-heterogeneity treatments, planting positions of the plants (red crosses) with the specified neighbouring soil patch types written in orange colour (a). Photos of heterogeneous soil treatment (b) and the homogeneous soil treatment (c) applied in 3-L pots (upper Ø: 19 cm, lower Ø: 14 cm, height: 15 cm). Plants after one week of growth at the centre positions (d) and at the boundary positions (e) in the heterogeneous soil treatment
Materials and methods
Study species
In order to increase the generalizability of our study (van Kleunen et al. 2014), we selected 10 species that are either native to or naturalized in Germany (Table S1). These species represented eight families; the Plantaginaceae were represented by one native and one naturalized species, and the Asteraceae, which includes many naturalized species (Pysek et al. 2017), was represented by two naturalized species. All species are widely distributed in Germany according to the FloraWeb database (www.floraweb.de). Seeds of the 10 study species came from the seed collection of the Botanical Garden of the University of Konstanz or from commercial seed suppliers (Table S1).
Plant materials, experimental design and measurements
We conducted the experiment in a greenhouse of the Botanical Garden of the University of Konstanz, Germany (47°41′32″N, 9°10′41″E).
From the 23rd to the 30th of May 2022, we germinated seeds of the ten plant species in a greenhouse. To assure that the seedlings of the different species would be at a similar developmental stage at the start of the experiment, we sowed the seeds on different dates based on knowledge about the time to germination from previous experiments (Table S1). Seeds of each species were sown separately in plastic trays (18.5 cm × 14 cm × 5 cm, length × width × height) filled with a peat-based potting soil (Pikiererde®, Einheitserdewerke Patzer PATZER ERDEN GmbH, Sinntal—Altengronau, Germany; pH 5.8; 1.5 g/L KCl; 180 mg/L N; 200 mg/L P2O5; 240 mg/L K2O; 130 mg/L S; and 150 mg/L Mg).
We created spatially homogeneous and spatially heterogeneous soils in 3-L pots (upper Ø: 19 cm, bottom Ø: 14 cm, height: 15 cm). With a specifically designed plastic divider, we divided each pot into three equal-sized patches. In pots with heterogeneous soil, we filled one patch with sand (commercial river sand), one patch with peat-based potting substrate that contains 55% white peat (Cyclamen®, HAWITA GRUPPE GmbH, Vechta, Germany; pH 6.0; 150 mg/L N; 150 mg/L P2O5; 250 mg/L K2O; and 130 mg/L Mg), hereafter referred to as ‘peat’, and one patch with a 1:1 (v:v) mixture of sand and peat (Fig. S1 b). In pots with homogeneous soil, all three patches were filled with the 1:1 mixture of sand and peat (Fig. S1 c). The boundaries between the patches, both in the heterogeneous and homogeneous soil treatments, were marked on the pot rim with a white permanent marker. So, even though the pots in the homogeneous soil treatment did not have true patches, we still marked, for comparative reasons, the locations of ‘pseudo-patch’ boundaries based on the position of the divider. The divider was removed from each pot after filling in the soil (Fig. 1b, c).
On the 20th of June 2022, we started to transplant seedlings of the five naturalized alien and five native plant species. Each pot received three individuals of one of the ten species. In order to test the effect of planting position, we either transplanted the seedling in the centres of the three soil patches in each pot (centre position, Fig. 1 d) or at the boundaries between the three soil patches (boundary position, Fig. 1 e). In total, our factorial experiment (Fig. 1 a) consisted of two soil heterogeneity treatments × two planting position treatments × (five naturalized alien species + five native species) × five replicates = 200 pots, and we had a total of 600 seedlings. Each replicate set of all species-by-treatment combinations consisted of 40 pots that were placed together as a block in the greenhouse.
In the week after transplanting, dead seedlings were replaced, and then initial leaf areas of all seedlings were estimated in order to be able to account for initial size differences in the analyses. We counted the number of true leaves and cotyledons and measured the length and width of the largest true leaf and cotyledon of each seedling. As it was difficult to separate the cotyledon from the true leaves of the grass species Anthoxanthum odoratum, we only measured the length and width of the largest leaf and counted the total number of leaves for this grass. Then, we randomly selected for each species 20 spare seedlings from the germination trays, and we measured the lengths and widths of all their leaves and cotyledons. Then, we scanned all these leaves and cotyledons using a flatbed scanner, and determined the individual leaf and cotyledon areas using the software ImageJ (Schneider et al. 2012). We then used linear regression for each species separately to establish the relationships of the single leaf area and the single cotyledon area with the length and width measurements. Subsequently, we used the species-specific regression equations to estimate the areas of the largest leaf and cotyledon on each seedling at the beginning of the experiment. We multiplied these values with the leaf and cotyledon numbers, respectively, to obtain an estimate of the total initial leaf area of each seedling in the experiment (Table S2).
The experiment ran for ten weeks. The air temperature in the greenhouse during the experiment was between 16.2 and 29.2°C. During the experimental period, we watered each pot regularly to keep the soil moist. On the 28th of August 2022, we harvested the aboveground plant parts of each of the three individuals in a pot separately. After that, we randomly chose three of the five blocks and washed the roots free from substrate. As the root washing was very labour intensive, we did not do this for the other two blocks. Moreover, root washing was done at the pot level as the roots of the three individual plants in a pot were intertwined and therefore difficult to separate. All the plant materials were dried in an oven at 70℃ for at least 72 hours before weighing.
Statistical analyses
In addition to the aboveground biomass data of each of the three plant individuals per pot, we also calculated the pot-level aboveground biomass data as a performance estimate of the small experimental population of three plants in each pot. This way, we were able to test our different research questions at the level of individual plants and at the level of populations (pots).
Firstly, to test whether the different soil-patch types, planting position, and the origin of the plant species affected the aboveground biomass of individual plants, we analysed the subset of individual-plant data collected from pots in the heterogeneous treatment. To test whether individual plants planted on the boundary position between patches produced more or less biomass than expected, we calculated for the pots in the patch-centre treatment for each neighbouring soil-patch type (sand|mixture, sand|peat, mixture|peat) the average aboveground biomass of the two plants in the centre positions of the respective neighbouring patches. In other words, the aboveground biomass value of the sand|peat neighbouring patches in the centre position treatment was the average of the individual in the centre of the sand patch and the individual in the centre of the peat patch. We then fitted the model with the explanatory variables in a specified order according to our experimental design. We first added the initial leaf area of each plant as a covariate to account for initial size variation. For the centre position treatment, this was the average initial leaf area of the two plants in the centres of the respective neighbouring patches. We then added block as a fixed factor to account for variation caused by differences in environmental conditions in the greenhouse. After that, we included neighbouring soil-patch type (sand|mixture, sand|peat, mixture|peat), planting position (centre and boundary position), the origin of species (naturalized alien or native species), and their interactions as fixed effects in the model. To account for non-independence of the three aboveground biomass values per pot, we included pot identity as a random effect. As we were interested in how species with different origins responded to neighbouring soil-patch type and planting position rather than in species specific differences, we included species identity as a random factor. In addition, we added for each of the interactions associated with species origin in the fixed terms of the model, the corresponding interaction associated with species identity as a random term to account for species specific differences in their responses to the treatments.
Secondly, we tested how soil heterogeneity, planting position, and the origin of plant species affected the total biomass of the plants in each pot (i.e. the small populations of three individuals). As we collected the belowground biomass per pot for only three of the five blocks, we also restricted the analysis of the aboveground biomass per pot to those three blocks. However, the results were qualitatively the same when we analysed the full dataset of aboveground biomass per pot (Table S4, Figure S1). As the roots of Veronica persica were extremely fine and fragile, we lost most roots of this species during root washing; hence we excluded this species from the pot-level analyses. We calculated the total biomass as the sum of the above- and belowground biomass per pot, and the root weight ratio as the proportion of belowground biomass relative to the total biomass per pot. These traits thus were also only available for the pots in the three blocks for which we had collected belowground biomass. Similar to the model described for the analyses of aboveground biomass of individual plants, we first fitted initial leaf area (the sum of the initial leaf areas of each of the three individual plants) and block. After these two fixed terms, we included soil heterogeneity (heterogeneous and homogeneous), planting position (centre and boundary position), the origin of species (naturalized alien or native species), and their interactions as further fixed effects in the model. We included species identity and its interactions with soil heterogeneity and planting position as random terms to account for species specific differences and responses to the treatments.
For all models, we graphically checked whether the residuals were normally distributed using histograms and quantile–quantile plots. Homoscedasticity was checked by plotting the residuals against the fitted values and against each explanatory variable. Whenever needed, data were transformed to improve normality and homoscedasticity of the residuals (Table S3). All the linear mixed effects models were fitted using the aov function of the R stats package, the corrected F-test for each model was calculated using the aov.ftest function of the pascal package (Niklaus 2023), and figures were made using the ggplot function of the ggplot2 package (Wickham 2016) in R 4.3.0 (R Core Team 2023). When there was at least a marginally significant interaction of fixed variables, we performed post-hoc pairwise comparisons of estimated marginal means among the different treatment combinations using the emmeans function of the emmeans package (Lenth 2023). We then obtained the compact letter display of all pair-wise comparisons using the cld function of the multcomp package (Hothorn et al. 2008).
Results
Growth responses of individual plants to different neighbouring soil patches
Aboveground biomass was highest for plants in the mixture|peat neighbouring soil-patches (mean ± se: 2.87 ± 0.38 g), followed by plants in the sand|peat neighbouring soil-patches (mean ± se: 2.42 ± 0.32 g), and lowest for plants in the sand|mixture neighbouring soil-patches (mean ± se: 1.78 ± 0. 21 g) (Table 1, Fig. 2a, b). The effect of the planting position on aboveground biomass depended on the neighbouring soil-patch types (significant patch type × position interaction in Table 1, Fig. 2a). In the mixture|peat neighbouring soil-patches, plants produced on average more biomass when they were planted in the centres rather than directly on the boundary position of the neighbouring soil-patches, whereas the opposite was true for the sand|mixture neighbouring soil-patches, and the planting position did not matter for the sand|mixture neighbouring soil-patches (Fig. 2a). Moreover, naturalized alien plants tended to respond more strongly to the different neighbouring soil patches than native plants (marginally significant patch type × origin interaction in Table 1, Fig. 2b). The aboveground biomass of naturalized alien plants was on average 31.2% higher than that of native plants in the sand|mixture neighbouring soil-patches, and this was even larger in the sand|peat (66.9%) and mixture|peat (71.6%) neighbouring soil-patches (Fig. 2b).

a Effects of neighbouring soil-patch types (sand|mixture, sand|peat and mixture|peat) and planting position (centre and boundary position) on aboveground biomass of individual plant; b effects of neighbouring soil-patch types (sand|mixture, sand|peat and mixture|peat) and origin of plants (native and naturalized alien) on aboveground biomass of individual plants per neighbouring soil-patches. Data shown are mean species group values (± SEs) based on the means of the individual species. The significant and marginally significant two-way interactions (see Table 2) are indicated in the figures (P < 0.1: †, P < 0.01: **). Different letters on top of the bars indicate significant differences between treatments (P < 0.05)
Growth responses of plant populations in heterogeneous and homogeneous soils
The populations of three plants per pot planted on the boundary positions of the neighbouring patches produced overall 10.9% more belowground biomass than the populations of plants planted in the centres of the soil patches (significant position effect in Table 2, Fig. 3b). Soil heterogeneity did not significantly affect belowground biomass, but the aboveground biomass of the plant populations (i.e. the joint aboveground biomass of the three plants in a pot) and the total biomass in the heterogeneous soils was significantly higher (+ 16.3% and + 11.9% for aboveground and total biomass, respectively) than in the homogeneous soil (significant soil-heterogeneity effects in Table 2, Fig. 3a). Moreover, the naturalized alien plant populations tended to benefit more strongly from the soil heterogeneity (+ 18.1% increase in total biomass) than the native plant populations did (+ 6.1%; marginally significant soil heterogeneity × origin interaction in Table 2, Fig. 3c). Planting position, however, did not significantly affect the total biomass per pot (Table 2, Fig. 3c). Despite different responses of aboveground and belowground biomass, the root weight ratio of the plant populations was not significantly affected by soil heterogeneity, planting position, the origin of plants and their interactions.

Population-level or pot-level effects of soil heterogeneity (H: heterogeneous and homogeneous), planting position (centre and boundary positions) and origin of plant species (native and naturalized alien) on aboveground biomass (a) belowground biomass (b), total biomass (c) and root weight ratio (d) of the plant populations. Data shown are mean species group values (± SEs) based on the means of the individual species. The significant and marginally significant effects (see Table 1) are indicated in the figures. P < 0.1: †, P < 0.05: *, P < 0.001: ***. Different letters in orange colour on top of the bars indicate significant differences between soil heterogeneity treatments averaged across position treatments (P < 0.05). Different letters in blue colour on top of the bars in the box indicate significant differences between position treatments averaged across soil heterogeneity treatments (P < 0.05)
Discussion
Our study showed that plants on the heterogeneous soils grew best when they were in the centre or on the boundary of peat patches, and this positive effect of peat tended to be stronger for the naturalized alien species than for the native species. The effect of growing on the boundary between patches instead of in the centres of the respective patches was positive for the sand|peat neighbouring soil-patches, neutral for the sand|mixture neighbouring soil-patches and negative for the mixture|peat neighbouring soil-patches. This suggest that the effect of growing on the boundary between patches depends on patch contrast. The aboveground and total biomass of the small plant populations in each pot was higher in the heterogeneous than in the homogeneous treatment, and this benefit of soil heterogeneity also tended to be stronger for naturalized alien than for native species. Belowground biomass of the plant populations, however, was not affected by soil heterogeneity per se but was higher when plants grew on the boundary position between patches instead of in the patch centres. So, in line with the results of previous studies (Wei and van Kleunen 2022), we found that naturalized alien plants benefited more from heterogeneous soils than native plants do. Moreover, we found evidence that the growing position within heterogeneous environments might also matter for biomass production.
Individual plants benefited from access to peat patches
Individual plants in the heterogeneous pots produced significantly more aboveground biomass when they had direct access to peat patches (Fig. 2a). As peat-based planting soil is rich in nutrients (i.e. has fertilizer added) and has a high water-holding capacity, it is not surprising that individual plants took advantage of the peat patches, as has also been found in other studies (Kitir et al. 2018; Wei and van Kleunen 2022). A previous study found that through selective biomass allocation to roots and root foraging), plants on boundary positions can access and benefit from nutrient-rich patches (Kowalski and Henry 2020). As the patches within each pot were not physically separated in our study, roots of plants in the patch centres could explore other patches and grow into the more resource-rich patches. As we observed during root washing that the roots of the three plants per pot were intertwined, they indeed grew into the different patches. Nevertheless, being positioned straight on the boundary of a nutrient-rich patch rather than at a larger distance from it, may provide it with a priority effect as it can rapidly take up the resources and even take up space that help to exclude later arriving roots of plants that are more distant from the boundary (Mommer et al. 2012; Craine and Dybzinski 2013). Although we could not investigate the roots of the individual plants and soil patches, we found that the populations of plants positioned on the boundary positions of the patches produced more root biomass than those positioned in the centres of the patches (Fig. 3a). Interestingly, the position effect on belowground biomass did not differ significantly between the heterogeneous and homogeneous soil treatments. This suggests that even in pots without true patches, there was a position effect. A possible explanation for this could be that the removal of the divider used during infilling of the pots resulted in improved aeration of the soil at the positions where the divider had been. Nevertheless, the position effect appears to be mainly present in the pots with heterogeneous soil (Fig. 2b). Therefore, we conclude that patch boundaries might provide the plants with unique opportunities to proliferate their roots and to have access to more soil resources.
Interestingly, the effect of the planting position on aboveground biomass of the individual plants depended on the neighbouring soil-patch type. Being directly on the boundary position was advantageous for the plants in sand|peat neighbouring soil-patches, but disadvantageous in mixture|peat neighbouring soil-patches and neutral in sand|mixture neighbouring soil-patches. One possible explanation could be that the contrast between neighbouring patches mattered. In our study, the contrast between sand and peat patches was larger than those between the other patches. However, although based on the difference in the amount of peat-based planting soil, the contrasts of sand|mixture and mixture|peat are the same, based on the biomass differences between individual plants in the centres of the patches in the heterogeneous treatment, the contrast was lower for the mixed|peat neighbouring soil-patches than for the sand|mixture neighbouring soil-patches (Figure S2). In line with our results, a previous study also found that the aboveground biomass of plants was higher under heterogeneous nutrient conditions with high-contrast patches than with low-contrast patches (Wang et al. 2018). So, overall, these findings suggest that with increasing patch contrast the effect of being on the boundary position changes from negative to positive.
A limitation of our study is that we used only one patch size, but the effect of planting position (i.e. centre vs boundary position) is likely to depend on the patch size. As in our study the size of each patch was relatively small (283 cm2), roots of the plants positioned in the patch centres could still access the other patches. Therefore, even the plants in the centres of the sand patches still had access to the higher resource levels in the neighbouring patches. And it is thus likely that the benefit of the boundary position for sand|peat was mainly due to a priority effect. However, if the patches would have been larger, the plants at the centre of the sand patches would have been even more limited by resources, while the plants at the centre of the peat patches would have experienced even less competition from plants in the neighbouring patches. So, whether the beneficial boundary-position effect for high-contrast patches holds for larger patches remains to be tested.
Naturalized alien plants take more advantage from soil heterogeneity than natives
Overall, the aboveground biomass as well as the total biomass of the small plant populations were significantly higher in pots with heterogeneous soils than in pots with homogeneous soils. Furthermore, we found that this overall beneficial effect of soil heterogeneity could mainly be attributed to the naturalized aliens and less so to the natives, although this was only marginally significant (Fig. 3c). Even though the total amounts of sand and peat, and thus nutrients, were the same in the heterogeneous and homogeneous pots, the aggregation of peat in patches of the heterogeneous soils, is likely to have enhanced the access to the available nutrients. The naturalized alien plants may have benefited more strongly from this than the native plants. Specifically, irrespective of the planting position, naturalized alien plant individuals produced 79% more aboveground biomass on the nutrient rich mixture|peat than on the nutrient poor sand|mixture, whereas native plant individuals only achieved 37% more aboveground biomass (Fig. 2b). According to FloraWeb (the online database providing information on wild plants in Germany), all naturalized alien plant species included in our study have wide geographic ranges in Germany, based on the number of grid cell occurrences (Table S1). Moreover, many of them can be locally abundant based on the dominance indicator value in FloraWeb (Table S1). This means that all the naturalized alien species in our study are invasive in at least one or two of the four invasiveness dimensions in the framework proposed by Catford et al. (2016). It has been reported that naturalized aliens, and especially those that spread widely and are considered invasive or weedy, tend to capitalize more intensively on high resource access due to their more acquisitive growth strategies (van Kleunen et al. 2010; Dawson et al. 2012). Another recent study found that the invasive species Spartina alterniflora exerted a high level of root plasticity to nitrogen addition, and that this allowed the species to take more advantage of the increased nutrient availability than the co-occurring native species Phragmites australis (Liu et al. 2023). The over-proportional growth increase of naturalized aliens due to higher resource access in the heterogeneous soils at the individual plant level, added up and resulted in an overall benefit for the populations of naturalized alien plants over the native ones.
The stronger effect of soil heterogeneity on naturalized alien than on native species may in nature affect the interactions between them and thus the invasion process. From the perspective of the roots, alien plants with a high phenotypic plasticity could pre-empt nutrient-rich patches in the heterogeneous soil leaving few nutrients for the later arriving species (Craine and Dybzinski 2013; Schwinning et al. 2017). From the perspective of the shoots, the fast growth and tall stature of many naturalized alien plants may provide an advantage when competing for light and might simultaneously impose shade stress to smaller neighbours (Craine and Dybzinski 2013; Schwinning et al. 2017). Indeed, in a previous study it was found that populations of naturalized alien plants benefited from heterogeneous soil conditions by capitalizing more strongly on nutrient-rich patches, especially when a native community was present (Wei and van Kleunen 2022). Especially in highly grazed habitats, while large grazers potentially create more nutrient rich patches and increase the N availability with their excreta (Mikola et al. 2009), at the same time grazing creates open patches in the plant communities. According to our findings, naturalized alien species are more likely to take advantage of the heterogeneous environments and take up the open space in grazed ecosystems than native species. Nevertheless, other studies suggest that the effect of soil heterogeneity on plant species might not always be beneficial, can be species specific, and is dependent on other factors (Mommer et al. 2012). Therefore, more studies are needed to understand the effect of soil heterogeneity on the invasion process.
In our current study, we could only include ten species, which is a small proportion of the local species pool, and the naturalized alien and native species were not phylogenetically paired. Therefore, future studies should aim to include more species and to create more phylogenetically balanced species selection when doing multispecies experiments. It should also be considered that some of the native species used in our study are naturalized in other regions or continents. Thus, these native species could be strong competitors, like the naturalized alien species in our study, possibly having similar traits and responses to soil heterogeneity. So, differences between naturalized alien and native species might have been larger, if we had selected native species that themselves are not naturalized anywhere in the world. Therefore, it might be important that future study should also test the effects of soil heterogeneity on both naturalized alien and native species with different global naturalization success status.
Conclusion and future directions
Our study suggests that soil heterogeneity promotes the growth of naturalized alien plants over native plants. Moreover, growing on the boundary of patches might provide benefits to individual plant growth and belowground biomass production of plant populations, at least when the patch contrast is sufficiently high. Therefore, boundaries of soil patches may offer specific ecological niches in heterogeneous environments. Although the beneficial growth effects of boundary position did not differ between the native and naturalized plant individuals and populations in our study, whether or not the boundaries will affect the interactions between these native and naturalized plants remain unclear. Furthermore, it is generally known that soil heterogeneity can influence soil microbial processes. Yet our understanding of how soil heterogeneity interacts with microbial processes and how the soil microbiome affects plant communities is still limited. It might thus be a good starting point to explore the microbial communities on the soil patch boundaries and link them to the plant communities in future studies.
Data availability
Data related to this paper can be accessed through the figshare repository: https://doi.org/10.6084/m9.figshare.26053192.v1.
References
Baer SG, Adams T, Scott DA, Blair JM, Collins SL (2020) Soil heterogeneity increases plant diversity after 20 years of manipulation during grassland restoration. Ecol Appl 30(1):e02014
Bauerle TL, Smart DR, Bauerle WL, Stockert C, Eissenstat DM (2008) Root foraging in response to heterogeneous soil moisture in two grapevines that differ in potential growth rate. New Phytol 179:857–866
Cadenasso ML, Pickett STA, Weathers KC, Jones CG (2003) A framework for a theory of ecological boundaries. Bioscience 53:750–758
Catford JA, Baumgartner JB, Vesk PA, White M, Buckley YM, McCarthy MA (2016) Disentangling the four demographic dimensions of species invasiveness. J Ecol 104:1745–1758
Christen D, Matlack G (2006) The role of roadsides in plant invasions: a demographic approach. Conserv Biol 20:385–391
Craine JM, Dybzinski R (2013) Mechanisms of plant competition for nutrients, water and light. Funct Ecol 27:833–840
Davies GTO, Kirkpatrick JB, Cameron EZ, Carver S, Johnson CN (2019) Ecosystem engineering by digging mammals: effects on soil fertility and condition in Tasmanian temperate woodland. Royal Society Open Science 6:180621
Davis MA, Grime JP, Thompson K (2000) Fluctuating resources in plant communities: a general theory of invasibility. J Ecol 88:528–534
Dawson W, Rohr RP, van Kleunen M, Fischer M (2012) Alien plant species with a wider global distribution are better able to capitalize on increased resource availability. New Phytol 194:859–867
Day KJ, Hutchings MJ, John EA (2003) The effects of spatial pattern of nutrient supply on the early stages of growth in plant populations. J Ecol 91:305–315
Gazol A, Tamme R, Price JN, Hiiesalu I, Laanisto L, Partel M (2013) A negative heterogeneity-diversity relationship found in experimental grassland communities. Oecologia 173:545–555
Hodge A (2004) The plastic plant: root responses to heterogeneous supplies of nutrients. New Phytol 162:9–24
Hothorn T, Bretz F, Westfall P (2008) Simultaneous Inference in General Parametric Models. Biom J 50:346–363
Huang L, Chen R-Y, Xue W, Yu F-H (2023) Effects of scale and contrast of spatial heterogeneity in plant-soil feedbacks on plant growth. Sci Total Environ 878:163159
Hutchings MJ, John EA, Wijesinghe DK (2003) Toward understanding the consequences of soil heterogeneity for plant populations and communities. Ecology 84:2322–2334
Iseli E, Chisholm C, Lenoir J, Haider S, Seipel T, Barros A, Hargreaves AL, Kardol P, Lembrechts JJ, McDougall K, Rashid I, Rumpf SB, Arévalo JR, Cavieres L, Daehler C, Dar PA, Endress B, Jakobs G, Jiménez A, Küffer C, Mihoc M, Milbau A, Morgan JW, Naylor BJ, Pauchard A, Ratier Backes A, Reshi ZA, Rew LJ, Righetti D, Shannon JM, Valencia G, Walsh N, Wright GT, Alexander JM (2023) Rapid upwards spread of non-native plants in mountains across continents. Nature Ecology & Evolution 7:405–413
Keser LH, Dawson W, Song Y-B, Yu F-H, Fischer M, Dong M, van Kleunen M (2014) Invasive clonal plant species have a greater root-foraging plasticity than non-invasive ones. Oecologia 174:1055–1064
Keser LH, Visser EJW, Dawson W, Song YB, Yu FH, Fischer M, Dong M, van Kleunen M (2015) Herbaceous plant species invading natural areas tend to have stronger adaptive root foraging than other naturalized species. Front Plant Sci 6
Kitir N, Yildirim E, Şahin Ü, Turan M, Ekinci M, Ors S, Kul R, Ünlü H, Ünlü H (2018) Peat use in horticulture. In: Topcuoğlu B, Turan M (eds) Peat (pp. ch. 5). IntechOpen. Rijeka
Kotliar NB, Wiens JA (1990) Multiple scales of patchiness and patch structure: a hierarchical framework for the study of heterogeneity. Oikos 59:253–260
Kowalski JJ, Henry HAL (2020) Soil microedges provide an ecological niche for Desmodium canadense. Plant Ecol 221:15–24
Lenth RV (2023) _emmeans: estimated marginal means, aka least-squares means_. R package version 1.8.9. https://CRAN.R-project.org/package=emmeans. Accessed 18 Oct 2023
Liu H, Zhang Y, Xu X, Li S, Wu J, Li B, Nie M (2023) Root plasticity benefits a global invasive species in eutrophic coastal wetlands. Funct Ecol 38:165–178
Liu YJ, De Boeck HJ, Li ZQ, Nijs I (2019) Unimodal relationship between three-dimensional soil heterogeneity and plant species diversity in experimental mesocosms. Plant Soil 436:397–411
Liu YJ, Hou FJ (2021) Effects of three-dimensional soil heterogeneity on seed germination in controlled experiments. J Plant Ecol 14:1–9
Liu YJ, Li GE, Wang MX, Yan WJ, Hou FJ (2021) Effects of three-dimensional soil heterogeneity and species composition on plant biomass and biomass allocation of grass-mixtures. Aob Plants 13:8
Lundgren EJ, Ramp D, Stromberg JC, Wu J, Nieto NC, Sluk M, Moeller KT, Wallach AD (2021) Equids engineer desert water availability. Science 372:491–495
Maestre FT, Bradford MA, Reynolds JF (2006) Soil heterogeneity and community composition jointly influence grassland biomass. J Veg Sci 17:261–270
Maestre FT, Cortina J, Bautista S, Bellot J, Vallejo R (2003) Small-scale environmental heterogeneity and spatiotemporal dynamics of seedling establishment in a semiarid degraded ecosystem. Ecosystems 6:630–643
Mikola J, Setälä H, Virkajärvi P, Saarijärvi K, Ilmarinen K, Voigt W, Vestberg M (2009) Defoliation and patchy nutrient return drive grazing effects on plant and soil properties in a dairy cow pasture. Ecol Monogr 79:221–244
Mommer L, van Ruijven J, Jansen C, van de Steeg HM, de Kroon H (2012) Interactive effects of nutrient heterogeneity and competition: implications for root foraging theory? Funct Ecol 26:66–73
Niklaus PA (2023) _pascal: Pascal’s library_. R package version 1.17.1
Parepa M, Fischer M, Bossdorf O (2013) Environmental variability promotes plant invasion. Nat Commun 4:1604
Prietzel J (2020) Soil phosphorus heterogeneity improves growth and P nutrition of Norway spruce seedlings. Front For Glob Chang 3:17
Pysek P, Pergl J, Essl F, Lenzner B, Dawson W, Kreft H, Weigelt P, Winter M, Kartesz J, Nishino M, Antonova LA, Barcelona JF, Cabezas FJ, Cardenas D, Cardenas-Toro J, Castano N, Chacon E, Chatelain C, Dullinger S, Ebel AL, Figueiredo E, Fuentes N, Genovesi P, Groom QJ, Henderson L, Inderjit, Kupriyanov A, Masciadri S, Maurel N, Meerman J, Morozova O, Moser D, Nickrent D, Nowak PM, Pagad S, Patzelt A, Pelser PB, Seebens H, Shu WS, Thomas J, Velayos M, Weber E, Wieringa JJ, Baptiste MP, van Kleunen M (2017) Naturalized alien flora of the world: species diversity, taxonomic and phylogenetic patterns, geographic distribution and global hotspots of plant invasion. Preslia 89:203–274
R Core Team (2023) R: a language and environment for statistical computing. R foundation for statistical computing, Vienna. https://www.R-project.org/. Accessed 18 Oct 2023
Richards CL, Bossdorf O, Muth NZ, Gurevitch J, Pigliucci M (2006) Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecol Lett 9:981–993
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9:671–675
Schwinning S, Meckel H, Reichmann LG, Polley HW, Fay PA (2017) Accelerated development in Johnsongrass seedlings (Sorghum halepense) suppresses the growth of native grasses through size-asymmetric competition. PLoS ONE 12:18
Skálová H, Havlícková V, Pysek P (2012) Seedling traits, plasticity and local differentiation as strategies of invasive species of Impatiens in central Europe. Ann Bot 110:1429–1438
Smart SM, Bunce RGH, Firbank LG, Coward P (2002) Do field boundaries act as refugia for grassland plant species diversity in intensively managed agricultural landscapes in Britain? Agr Ecosyst Environ 91:73–87
Son D, Chu Y, Lee H (2024) Roads as conduits for alien plant introduction and dispersal: the amplifying role of road construction in Ambrosia trifida dispersal. Sci Total Environ 912:169109
Speißer B, van Kleunen M (2023) Plants forage for soil patches free of plastic pollution but cannot bag the profits. Sci Rep 13:18506
Stover HJ, Henry HAL (2018) Soil homogenization and microedges: perspectives on soil-based drivers of plant diversity and ecosystem processes. Ecosphere 9:e02289
Stuefer JF (1996) Potential and limitations of current concepts regarding the response of clonal plants to environmental heterogeneity. Vegetatio 127:55–70
Tamme R, Gazol A, Price JN, Hiiesalu I, Partel M (2016) Co-occurring grassland species vary in their responses to fine-scale soil heterogeneity. J Veg Sci 27:1012–1022
van Kleunen M, Dawson W, Bossdorf O, Fischer M (2014) The more the merrier: multi-species experiments in ecology. Basic Appl Ecol 15:1–9
van Kleunen M, Weber E, Fischer M (2010) A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol Lett 13:235–245
Wang P, Yang Y, Mou P, Zhao QZ, Li YB (2018) Local root growth and death are mediated by contrasts in nutrient availability and root quantity between soil patches. Proc R Soc B-Biol Sci 285:7
Wang YJ, Muller-Scharer H, van Kleunen M, Cai AM, Zhang P, Yan R, Dong BC, Yu FH (2017) Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol 216:1072–1078
Wei G-W, van Kleunen M (2022) Soil heterogeneity tends to promote the growth of naturalized aliens when competing with native plant communities. J Ecol 110:1161–1173
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag, New York
Wijesinghe DK, Hutchings MJ (1999) The effects of environmental heterogeneity on the performance of Glechoma hederacea: the interactions between patch contrast and patch scale. J Ecol 87:860–872
Wijesinghe DK, John EA, Hutchings MJ (2005) Does pattern of soil resource heterogeneity determine plant community structure? An experimental investigation. J Ecol 93:99–112
Acknowledgements
We thank Otmar Ficht, Kristina Schlötter, Beate Rüter, Ekaterina Mamonova, Fang-Wei Lin, Benedikt Speißer, Weihan Zhao, Bernadette Pham, Louisa Neubauer, Anna Rankl, and Shila Teichmann for practical assistance. We thank Pascal A. Niklaus for providing valuable suggestions on statistical analyses. Guan-Wen Wei acknowledges funding from the China Scholarship Council (201906510047) and support from the International Max Planck Research School for Quantitative Behaviour, Ecology and Evolution.
Funding
Open Access funding enabled and organized by Projekt DEAL. Author Guan-Wen Wei has received funding from the China Scholarship Council (201906510047).
Author information
Authors and Affiliations
Contributions
G.-W.W. conceived the idea and designed the experiment; G.-W.W. performed the experiment; G.-W.W. and M.v.K. conducted the data analyses; G.-W.W. wrote the first draft of the manuscript; G.-W.W. and M.v.K. revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no conflict of interest to declare.
Additional information
Responsible Editor: Luca Bragazza.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wei, GW., van Kleunen, M. Growing on patch boundaries of heterogeneous soils promotes root growth but not the total biomass of naturalized alien and native plants. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06796-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06796-3