
Abstract
Sorbus L. sensu lato (Rosaceae family) comprises over 250 trees and shrubs growing in the Northern Hemisphere. Several Sorbus species have found their way into traditonal materia medica, as different leaf, bark or fruit preparations have a long-standing folk use. Ethnomedicine recommends their use not only in respiratory and gastrointestinal system disorders, but also in rheumatism, cancer or diabetes. Chemical composition of the genus Sorbus has been studied since 1960s and until now more than 250 compounds have been identified from thirty-eight species. The most thoroughly investigated substances are phenolic compounds (flavonoids and phenolic acids). Other constituents include triterpenes, sterols, carboxylic acids, coumarins and cyanogenic glycosides. Biological activity studies performed on the representatives of the genus Sorbus have been predominantly conducted on extracts prepared with the use of different solvents, and more rarely on isolated constituents. The majority of these studies were devoted to determination of antioxidant potential of Sorbus extracts. In a few cases targeted studies were conducted to confirm specific folk usage. Two of the fairly well documented medicinal uses are the antidiabetic activity of Sorbus decora and anti-inflammatory activity of Sorbus commixta.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sorbus L. sensu lato (Rosaceae family) comprises over 250 trees and shrubs native to eastern Asia and distributed in the Northern Hemisphere (Aldasoro et al. 1998). Genus Sorbus is characterized by significant diversity what inflicts taxonomic difficulties. Recent studies provide data on its polyphyletic nature. Interspecific hybridization, apomixis and polyploidy, which have contributed to diversification process, seem to play a crucial role in plant evolution (Dickinson et al. 2007; Li et al. 2017). Nevertheless, the genus Sorbus is currently usually divided into six subgroups: Chamaemespilus Medikus, Torminalis Medikus, Aria (Pers.) Host, Micromeles Decaisne, Cormus Spach and Sorbus sensu stricto (Li et al. 2017). All simple-leaved species are included into the first four subgroups, whereas the two latter comprise the pinnate-leaved species (Sun et al. 2018). The complicated relationships between Sorbus species are the subject of many current studies in the field of molecular biology. The Sorbus sensu stricto (88 species) is one of the most profoundly analyzed. Recent findings confirmed the existence of two major lineages, namely core Sorbus and Albo-carmesinae, which correspond to morphological characters such as i.a. fruit color. As confirmed by phylogenetic analyzes, the core Sorbus includes two clades (Aucupariae and Commixtae), whereas Albo-carmesinae comprises three clades (Tianshanicae, Discolores, Multijugae) (Li et al. 2017). The greatest diversity of Sorbus in Europe is seen in Britain, where in the area of Avon Gorge, among fifteen native taxa, three endemic species and four novel hybrids have been reported (Robertson et al. 2010). This interesting problem of the diversity and relationship network between different Sorbus species has been exhaustively discussed in a number of surveys (Chester et al. 2007; Robertson et al. 2010; Pellicer et al. 2012; Hamston et al. 2018). It is also noteworthy that the European phylogenetic checklist of Sorbus s.l. has been recently published (Sennikov and Kurtto 2017).
Sorbus species are often cultivated for ornamental purposes mainly because of the colored fruits. The berries are consumed worldwide, as raw or processed into preserves and beverages. Moreover, various parts of these valuable plants have been used as remedies for healing common ailments. Traditional medicine recommends their use not only in respiratory and gastrointestinal system disorders, but also in rheumatism, cancer or diabetes. However, not all declared therapeutic properties seemed to be confirmed by recent phytochemical and biological studies.
Despite a large number of literature reports dedicated to phytochemistry and biological activity of various Sorbus species to date, to the best of our knowledge there is only one published review, which is available in Polish (Olczyk and Geszprych 2017). Thus, this paper is not accessible to a wider group of readers. Moreover, it is primarily focused on practical and nutritional value of Sorbus representatives. Even though chemical composition and biological activity is addressed as well, the authors had not assembled all existing data, citing only twenty-five papers devoted to phytochemical and/or biological studies on Sorbus species. Hence, the aim of this work was to provide the first comprehensive overview of the genus Sorbus in English, in which information on ethnobotanical, phytochemical and biological studies is summarized.
Methods
The search of scientific literature was performed in various databases including Google scholar, PubMed, Scopus and Embase—using „Sorbus” as the key word. All articles published in English to date (September 2019) were collected. Those that did not concern phytochemistry, biological activity and/or ethnopharmacology of Sorbus species were rejected. The chemical structures of the phytoconstituents were searched in PubChem database and ChemDraw 19.0 was used to redraw selected structures.
Ethnopharmacological studies
Medicinal use
In the majority of collected reports, their authors underlined ethnopharmacological significance of investigated Sorbus species referring to herbal books and traditional knowledge developed over generations. Some papers however were specially devoted to this issue and revealed outcomes of in-depth interviews with local community members especially in the regions of Turkey and Korea. The available literature refers to eleven different Sorbus species valued worldwide as medicinal products.
Asia
S. cashmiriana is a tree found in Kashmir and the western Himalayas, where its bark is used in folk medicine to treat nausea and heart diseases. The berries of this plant are a remedy for scurvy (Khan et al. 2015). Another Asian species—S. commixta—mainly distributed in Korea, Japan and China has also a long-standing folk use. Its bark is applied in bronchitis, asthma or cough (Sohn et al. 2005b), fruits are used as gargle for throat infections or as a laxative (Lee et al. 2017) whereas stem bark is an anti-atherosclerosis and expectorant agent (Yin et al. 2005). Moreover, during analysis of ethnomedicinal practices of local communities in southern Korea, it was revealed that leaves, stem and fruits of S. commixta are traditionally used as a decoction or tea in liver disorders (Kim and Song 2013). People of North Jeolla Province (Korea) recommend its stem decoction in asthma and pulmonary tuberculosis. Fruits are indicated in cough and common cold (Kim and Song 2012). Apart from S. commixta, the bark of another species growing in Korea, namely S. amurensis is also used in this region as an expectorant (Kang et al. 2003). In China, two species have been valued as medicinal agents especially in the disorders of the respiratory tract. S. tianschanica is a traditional medicine in the treatment of asthma, dyspnea, ventricular myocytes and gastritis (Ayupbek et al. 2012). Another species which is found growing wild in Northern China—S. pohuashanensis—is recommended in chronic tracheitis, edema and pulmonary tuberculosis (Li et al. 2014).
Europe
Four species typical of Europe have found their way into traditonal materia medica. One of them is S. domestica, with a well documented traditional medicinal usage in the Mediterranean Basin area. This species is also a popular medicinal plant of Turkey. Kültür (2007) interviewed local communities living in Kırklareli Province and revealed that decoction of S. domestica leaves is used to treat prostatitis, diabetes, nephritis (infusion of the leaves is used as well), gallbladder ailments, diarrhea and kidney stones. There are also records of its cholesterol lowering and diuretic properties. What is more, bark decoction is a remedy in stomach ache or ulcers and—finally—fresh fruits are applied as an anti-diarrheal agent. People of Central Anatolia drink a tea of S. domestica leaves to treat bradyuria and kidney stones (Sezik et al. 2001). Fruits are used as astringent, diarrheic and antidiabetic agents by the people of Xanthi (Rodopi) (Termentzi et al. 2006), while in Serbia they are also used as a tonic (Jarić et al. 2015).
Another species that is a very popular natural remedy against various ailments both in Europe and Asia is S. aucuparia, which is commonly called rowan tree. Rowanberries are recommended in folk medicine to treat hemorrhoids, hypertension (Kültür 2007; Polat and Satıl 2012), respiratory tract infections, rheumatism and gout (Vogl et al. 2013). In Lithuania (Kaišiadorys dictrict) preparations from rowanberries were administered orally in constipation and cough, whereas bark decoction was used as a wound wash (Pranskuniene et al. 2019). Sak et al. (2014) revealed that a tea prepared from bark of S. aucuparia has been used in Estonia to treat cancer diseases. Preparations made from the rowan tree leaves apart from being indicated in cancer, were also used in gastrointestinal problems or prostatitis (Kültür 2007; Neves et al. 2009). Finally, the inflorescences were used as diuretic and anti-inflammatory agents (Olszewska 2011a). Two other Sorbus species valued in traditional medicine of Europe are S. torminalis and S. aria (whitebeam). In both cases leaves are used as a medicinal product, the decoction made from the former species is traditionally used in Turkey to treat diabetes or stomach ache (Kültür 2007), whereas the latter are used as an anti-diarrhoeal agent (Olszewska and Michel 2012).
North America
The bark of S. decora (showy mountain ash), a species native to North America, is used to relieve symptoms related to diabetes by the Eeyou Istchee Cree First Nations of Quebec, Canada (Leduc et al. 2006; Guerrero-Analco et al. 2010). Also the bark of S. americana is a traditional remedy in the boreal regions of Canada, where it is recommended in the treatment of diabetes and its complications (McCune and Johns 2002, 2003, 2007).
Food use
Berries obtained from different Sorbus species, apart from their medicinal use, are known for their nutritional value which is also well documented. S. commixta and S. pohuashanensis fruits are eaten in Asia as a refreshment or in preserves, the former being also used to produce country wine (Li et al. 2014; Lee et al. 2017). Rowanberries (S. aucuparia) are very popular edible products especially in Europe. They are often consumed raw or processed into jams, wines etc. (Tardío et al. 2006; Kalle and Sõukand 2012; Łuczaj et al. 2013; Pranskuniene et al. 2019). Fruits of S. aucuparia, S. intermedia and S. rupicola are bread ingredients in Estonia (Kalle and Sõukand 2012). S. torminalis and S. intermedia berries are consumed in preserves or raw, after natural fermentation or storage (Tardío et al. 2006; Łuczaj et al. 2013). S. aria fruits are processed into alcoholic beverages, vinegar and jams (Tardío et al. 2006; Olszewska and Michel 2012). Finally, S. domestica has nutritional value in Spain (Tardío et al. 2006), southern Herzegovina (Łuczaj and Dolina 2015) and Xanthi (Rodopi) (Termentzi et al. 2006).
Phytochemical studies
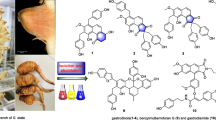
Chemical composition of the genus Sorbus has been studied since 1960s and until now more than 250 compounds have been identified from thirty-eight species. To the best of our knowledge, up to date Sorbus species have afforded twenty structurally new compounds, namely ten triterpenes, six phenols, two coumarins, one fatty acid and one dibenzofuran derivative. Examples of these are shown in Fig. 1. In general, the most thoroughly investigated substances are phenolic compounds. Other constituents include i.e. triterpenes, sterols, carboxylic acids, coumarins and cyanogenic glycosides.

Chemical structures of selected compounds isolated from the genus Sorbus
Phenolic compounds
As mentioned above, the highest number of papers devoted to phytochemical analysis of various Sorbus species concerns flavonoids and other phenols. Common sugar-free compounds (see Table 1) such as quercetin and kaempferol are the most frequent and were detected in leaves, inflorescences and fruits of many Sorbus species. A more rare flavonol, namely sexangularetin was found to be characteristic of inflorescences, but its small amounts were also detected in the leaves, e.g. of S. aria (about 14 mg/100 g of dw), S. domestica and S. intermedia (trace levels, < LOQ) (Olszewska 2008, 2012; Olszewska and Michel 2009). Moreover, in the study of Olszewska (2012) who applied a very sensitive RP-HPLC–PDA method to the analysis of different aerial parts of S. aria, S. torminalis, S. domestica and S. aucuparia, the presence of other aglycones, i.e. limocitrin, luteolin, chrysoeriol, apigenin, was revealed. Except for limocitrin, all remaining compounds were detected solely in S. torminalis. What is more, isorhamnetin was confirmed in S. aucuparia (inflorescence).
Apart from free flavonoids also their glycosides (see Table 1) have been reported in a number of representatives of the Sorbus genus. Rutin, hyperoside, isoquercitrin and other quercetin derivatives are definitely the most abundant. Tian et al. (2017) underlined that quercetin 3-O-(6”-malonyl)-glucoside constitutes over 50% of flavonoid glycosides present in the fruit extract of S. aucuparia (ethanol:water:acetic acid, 70:30:1, v/v/v).
Catechins and proanthocyanidins (mainly procyanidin B1 and procyanidin B2) were found in the leaves of several Sorbus species (see Table 2). Futhermore, procyanidin B2 was detected in S. aucuparia flowers (Olszewska et al. 2019) whereas procyanidin C1 in S. domestica leaves (Rutkowska et al. 2019a, b). Anthocyanins, including glycosides of cyanidin, pelargonidin and delphinidin, were reported in the fruits of S. aucuparia (Kylli et al. 2010; Boath et al. 2012; Klensporf-Pawlik and Przybylski 2015; Veberic et al. 2015; Tian et al. 2017), S. torminalis (Mikulic-Petkovsek et al. 2017), S. americana (Klensporf-Pawlik and Przybylski 2015) and S. discolor (Mikulic-Petkovsek et al. 2017).
Another common group of phenolic phytoconstituents are phenolic acids (see Table 3), of which hydroxycinnamic acids, primarily chlorogenic acid and its isomer neochlorogenic acid, are widely distributed. These two caffeoylquinic acid derivatives were reported in many species of genus Sorbus. In S. aucuparia fruits they constitute almost 80% of total phenolics (Tian et al. 2017). Nevertheless, hydroxybenzoic acids, such as protocatechuic acid and p-hydroxybenzoic acid, were also detected in Sorbus genus. Other phenolics present in Sorbus species are listed in Table 4.
Triterpenoids
Up till now, thirty-three triterpenes have been identified in seven Sorbus species (see Table 5). These included already known structures as well as rare compounds that were identified exclusively within this genus. The levels of triterpenoids, especially in fruits, constitute a substantial part of crude plant material. Yin et al. (2019) obtained a triterpenoid fraction after four times repeated recrystallization from ethyl acetate extract of S. pohuashanensis fruits. The Vanillin-glacial acetic acid-perchloric acid colorimetric method confirmed that triterpenes constitute 64.6% of the total. Moreover, ursolic acid was found to be the main component. This triterpene was also reported in S. aucuparia fruits together with squalene, β-amyrin, α-amyrin, cycloartenol, betulin and oleanolic acid (Klavins et al. 2016). It is noteworthy that apart from these fairly common triterpenes some Sorbus species provided structurally novel compounds. For instance, S. cashmiriana (the whole plant) afforded lanosta-5,22,24(28)-triene-1β,3β-diol, (22E,24E)-1β,3β-dihydroxylanosta-5,22,24-triene-27-oic acid, 5,12(13),15 ursatriene-1β,3β,23α-triol, 5,12,19 lupatriene-1α,3β,12α,28β-tetraol, 3β,23-dihydroxylupa-1,20(29)-dien-28-oic acid as well as 3β-hydroxy-23-methoxylupa-1,20(29)-dien-28-oic acid, which were named cashmirol A, cashmirol B, sorbinol A, sorbinol B, sorbicin A and sorbicin B, respectively (Kazmi et al. 2007, 2009, 2011). Another lupane-type triterpene—sorbanolic acid—identified as 2α,3β,23-trihydroxy-lup-20(29)en-28-oic acid-23-caffeate was obtained from the stem wood of S. lanata (Latif et al. 2014), while 23,28-dihydroxyursan-12-ene-3β-caffeate, 23,28-dihydroxylupan-20(29)-ene-3β-caffeate and 3β,23,28-trihydroxy-12-ursene were isolated from stem bark of S. decora (Guerrero-Analco et al. 2010).
Carboxylic acids, alcohols, sugars and other compounds
Some Sorbus species were analyzed for the presence of fatty acids. Johansson et al. (1997) identified these compounds in the seeds of S. aucuparia with the largest amount of linoleic acid. The presence of fatty acids was also confirmed in rowanberries, in which the dominant acids namely linoleic, oleic and palmitic amounted to 18,841.0, 11,201.0, 5149.7 mg/kg of air-dried raw material, respectively. Palmitic acid was most abundant in leaves (3793.5 mg/kg). In turn, the leaves of S. aria afforded mainly palmitic and linolenic acids (Krivoruchko et al. 2013). A new fatty acid named sorcomic acid was isolated from the bark of Sorbus commixta (Kim et al. 2016). Simple organic acids, primarily malic, citric and oxalic were reported in the leaves of S. aria and S. aucuparia (Krivoruchko et al. 2013). The fruits of the latter species as well as of S. torminalis and S. doscolor yielded citric, malic, tartaric and fumaric acids (Mikulic-Petkovsek et al. 2012, 2017).
Other phytoconstituents reported in the Sorbus genus include alkanes (heptacosane, nonacosane), fatty alcohols (1-docosanol, 1-tricosanol, 1-tetracosanol, 1-pentacosanol, 1-hexacosanol, 1-octacosanol) and some monoglycerides (α-monopalmitin, α-monostearin), all of which were detected in S. aucuparia fruits (Klavins et al. 2016) Among sugars, the most common were sorbitol, glucose and fructose. In S. aucuparia fruits the levels of sorbitol and glucose reached 134.1 g/kg fw and 52.9 g/kg fw, respectively (Mikulic-Petkovsek et al. 2012), whereas for S. torminalis fruits the highest amounts were seen for glucose and fructose (98.88 and 80.67 g/kg fw, respectively). In turn, the contents of glucose, fructose and sorbitol in the fruits of another species—S. discolor—were comparable. Sucrose generally was found in small amounts, and was detected in the fruits of S. discolor, wild S. aucuparia as well as its cultivars. These compounds are listed in Table 6.
Nutritional value of Sorbus berries
Apart from sugars, fatty acids and simple organic acids, which are characterized in detail above (and listed in Table 6), the fruits of different Sorbus species seem also to be a good source of vitamin C, minerals, pigments and tocopherols (Häkkinen et al. 1999; Aslantas et al. 2007; Egea et al. 2010; Mlcek et al. 2014; Klavins et al. 2016; Mrkonjić et al. 2017; Šavikin et al. 2017). Although, the presence of toxic parasorbic acid should be also underlined, this compound degrades during fruit processing and does not pose a health risk (Mlcek et al. 2014).
It is also noteworthy that various Sorbus cultivars have been bred mostly to intensify the sweet flavor and improve organoleptic properties of fruits. Indeed, a comparison of the sugars/organic acids ratios of twenty Sorbus clones indicated significant differences (Zymone et al. 2018). Other phytochemical studies revealed that also flavonoid, phenolic acid, anthocyanin and carotenoid contents of various Sorbus cultivars varied distinctly (Hukkanen et al. 2006; Kylli et al. 2010; Mikulic-Petkovsek et al. 2017; Zymone et al. 2018). Nevertheless, the fruits of both Sorbus species and their varieties constitute a valuable contribution to daily diet.
Biological studies
Biological activity studies performed on the representatives of the genus Sorbus have been predominantly conducted on extracts prepared with the use of different solvents, and more rarely on isolated constituents. Although extracts from all parts of the plant were investigated, the majority of data refer to fruits, leaves and bark. Taking into account the high number of phenolic compounds that have been detected in various Sorbus species, the majority of biological activity studies were focused on the determination of their antioxidant potential. Interestingly, a large number of data is however available on the cytotoxic activity of Sorbus extracts.
Antioxidant activity
It should be underlined that the vast majority of antioxidant activity studies were performed using chemical methods. In fact, the ability of different Sorbus extracts (or isolates) to scavenge radicals in vitro was predominantly determined. It however does not provide information on the real antioxidant effects in a human body. Although chemical models dominate, several studies using cellular systems have also been conducted.
Antiradical activity in chemical models
Antioxidant properties of various fruits were often studied because they constitute a substantial part of human diet. This activity was also confirmed for Sorbus berries, especially those derived from S. domestica, S. aucuparia and S. torminalis. Termentzi et al. (2006) examined methanol extracts from fruits of S. domestica which differed in the maturity stage and found that unripe yellow fruits and fruit pulp were most effective. Moreover, these methanol extracts were next partitioned with dichloromethane, diethyl ether, ethyl acetate, butanol and water. The first three fractions showed significant radical-scavenging activity in the chemiluminescence method (greater than trolox). In another study, fruits of S. domestica (extracted with phosphate buffer) showed poor ability to scavenge ABTS·− (TEAC value of 0.466 ± 0.14 μmol trolox/g fw) and H2O2 (3.63 ± 2.29% of inhibition at 5 mg/mL), however generation of OH· radicals was found to be suppressed. Nevertheless, in the absence of ascorbic acid in the reaction mixture, inhibition of hydroxyl radicals decreased from 68.30% to 59.05%. Thus, the authors underlined that the antioxidant activity of S. domestica may be in part related to direct scavenging of radicals as well as avoiding ascorbate oxidation (Egea et al. 2010). In turn, methanol extracts from the fruits of S. aucuparia as well as S. caucasica var. yaltirikii had SC50 values of 0.366 ± 0.009 mg/mL and 0.520 ± 0.023 mg/mL in DPPH test, respectively. The results were compared to these of gallic acid (SC50 = 0.002 ± 0.000 mg/mL), chlorogenic acid (SC50 = 0.006 ± 0.000 mg/mL) and quercetin (SC50 = 0.003 ± 0.000 mg/mL) (Turumtay et al. 2017). Antioxidant activity of rowanberries (S. aucuparia) was also confirmed by Kähkönen et al. (1999, 2001), Ganhão et al. (2010), Aladedunye and Matthäus (2014) and Aladedunye et al. (2015). Moreover, rapeseed oil fortified with polyphenolic fractions of Sorbus extract displayed lower thermooxidative degradation (Aladedunye and Matthäus 2014). The fruits of S. aucuparia exerted antiradical properties towards various particles. The methanol extract showed an ability to scavenge hydroxyl radicals (16.33 ± 0.96%, percentage of inhibition), nitric oxides (25.17 ± 1.72%) and superoxide anions (26.74 ± 1.75%) (Mlcek et al. 2014). These findings were confirmed by Mrkonjić et al. (2017), who also studied two forms of S. torminalis (torminalis and semitorminalis). In another study, water and ethyl acetate extracts from S. torminalis fruits showed higher antioxidant activity than adequate acetone and methanol (the lowest activity) extracts. For instance, EC50 values for water extract were 5.30 ± 0.166, 5.69 ± 0.364 and 9.01 ± 1.025 mg/mL in ABTS, DPPH and NBT assays, respectively. However, the results were still lower than those of reference standards, i.e. quercetin and α-tocopherol (Hasbal et al. 2015). Scientific literature reports on antioxidant potential of other Sorbus berries, like those of S. hajastana or S. sambusifgolia (Lapinskii and Gorbachev 2006; Manukyan et al. 2019). DPPH and hydrogen peroxide scavenging activity as well as metal-chelating activity were also measured for methanol–water extract from S. umbellata (Desf.) Fritsch var. umbellata (we can only assume that the fruit extract was examined as the plant material was not defined). Its antioxidant activity was significant but not so high as that observed for Rosa canina or Rosa hemisphaerica extracts (Serteser et al. 2008). Acetone extract from fruits of S. pohuashanensis reached EC50 values of 60.3 ± 2.11 and 23.8 ± 0.74 µM trolox/g of fw in ABTS and DPPH assays, respectively (Fan et al. 2011). Finally, ethyl acetate fraction of methanol extract from S. americana fruits had IC50 value of 113.96 ± 5.48 µg/mL in DPPH test and displayed low activity in comparison to other edible American plants (Acuña et al. 2002).
Apart from fruits also Sorbus leaves are a source of antioxidant compounds. Olszewska and Michel (2012) partitioned methanol–water extract of S. aria leaves (pre-extracted with chloroform) with diethyl ether, ethyl acetate, n-butanol and water, and observed the best radical-scavenging activity in the DPPH test for the ethyl acetate fraction (EC50 = 2.99 ± 0.11 mg/L). Further analysis revealed that its main constituents are isoquercitrin, astragalin and chlorogenic acid. In turn, S. domestica leaf extracts were potent antioxidants in DPPH, FRAP, TBARS tests, especially ethyl acetate and diethyl ether fractions. Moreover, antioxidant activity correlated with the total polyphenol content. As was confirmed by Matczak et al. (2018), the defatted S. domestica methanol leaf extract (0.427 g trolox/g dw) appeared to have stronger activity than methanol extracts from the fruits (0.009–0.122 g trolox/g dw) (Termentzi et al. 2006; Matczak et al. 2018). More in-depth analysis of the leaf extract revealed nine predominant polyphenols, namely (–)-epicatechin, procyanidin B2, procyanidin C1, rutin, quercitrin, quercetin, quercetin 3-O-(2”-O-β-d-glucopyranosyl)-α-l-rhamnopyranoside, quercetin 3-O-(2″-O-β-d-xylopyranosyl)-α-l-rhamnopyranoside and chlorogenic acid (Rutkowska et al. 2019b). Successively, the scavenging activity of these compounds (against O·−2, OH·, NO·, H2O2, ONOO−, HClO) was assessed using suitable fluorimetric and spectrophotometric methods. The most potent were quercetin, (–)-epicatechin and procyanidins (3.94–24.16 μmol ascorbic acid/mg). In the same study, antiradical activities of the diethyl ether and ethyl acetate fractions were confirmed what correlates with previous reports (Rutkowska et al. 2019a). Methanol extracts from the leaves as well as fruits of S. aucuparia and S. caucasica var. yaltirikii were tested for their DPPH radical scavenging activity. The most potent were leaf extracts (containing primarily rutin derivatives) with SC50 values of 0.036 ± 0.001 mg/mL and 0.045 ± 0.000 mg/mL, respectively (Turumtay et al. 2017). Finally, the differences in antioxidant capacity of extracts from S. aucuparia leaves collected during the whole vegetation season were confirmed (Olszewska 2011a).
Antioxidant activity of other plant organs was also documented. Ethanol extracts from bark (collected twice in 2003 and 2004) and stem of S. decora were assessed using DPPH, CD and TBARS assays. Strong scavenging activity was noted with IC50 values ranging from 19.63 ± 0.57 to 48.95 ± 2.49 ppm in the DPPH test (ascorbic acid with IC50 = 3.84 ± 0.01 ppm). What is noteworthy, bark collection from 2003 displayed weaker activity (Fraser et al. 2007). DPPH radical scavenging activity of S. decora inner bark (80% ethanol extract) was confirmed as well (Spoor et al. 2006). The bark of S. americana (methanol extract) had IC50 value of 15.80 ± 1.91 ppm in DPPH assay (McCune and Johns 2002). Methanol extract from S. amurensis bark was partitioned between n-hexane, ethyl acetate, butanol and water and next all fractions were examined for their scavenging effects on superoxide and hydroxyl radicals. Among the tested extracts, n-butanol extract exhibited the highest activity (Kang et al. 2003). Bae et al. (2007) assessed antioxidant activity of 70% ethanol extract from S. commixta cortex and revealed that the extract had IC50 = 14.2 ± 0.8 μg/mL and IC50 = 18.0 ± 0.2 μg/mL in NBT and DPPH assays, respectively. In a different study, DPPH radical scavenging activity of S. commixta (70% ethanol extract, however plant organ was not mentioned, thus we can only assume that the extract was derived from cortex as well) was confirmed (Yu et al. 2009). Also Liu et al. (2015) revealed that ethanol (30%) extract from stem of S. commixta had IC50 value of 23.4 μg/mL in DPPH test. Moreover, catechin-7-O-β-d-xylopyranoside and catechin-7-O-β-d-apiofuranoside, isolated from stem bark of S. commixta, showed significant antioxidant activity (Na et al. 2002a).
Several authors performed comparative analysis of different Sorbus extracts. 70% methanol extracts from various aerial parts (and in one case also their diethyl ether, ethyl acetate, n-butanol and water fractions) of S. aucuparia, S. aria, S. intermedia, S. commixta, S. decora, S. gracilis, S. koehneana, S. pogonopetala, S. wilfordii, S. pohuashanensis, S. scalaris, S. prattii var. prattii, S. americana, S. sambucifolia, S. sitchensis, S. cashmiriana, S. setschwanensis were screened for antioxidant activity using different models e.g. FRAP, DPPH, TEAC and AAPH-induced linoleic acid peroxidation tests. One of the most promising seems to be S. aucuparia inflorescence next to S. wilfordii leaves as well as S. decora and S. koehneana inflorescences (Olszewska and Michel 2009; Olszewska et al. 2010, 2012). Indeed, diethyl ether, ethyl acetate, and n-butanol fractions of water–methanol flower extract of S. aucuparia showed significant radical-scavenging activity (especially towards OH·, ONOO−, HClO, and O2·−) (Olszewska et al. 2019). Olszewska (2011b) compared the activity of 70% methanol extracts from the leaves, fruits and inflorescences of S. torminalis with those of S. aucuparia using DPPH, ABTS and the AAPH-induced linoleic acid peroxidation test. S. torminalis extracts expressed antiradical activity towards DPPH, with TEAA values from 62.0 to 244.1 μmol trolox/g dw. Still, plant extracts derived from this species possessed weaker ability to scavenge radicals than those obtained from S. aucuparia. Ekin et al. (2016) underlined that among all tested thirty-four Rosaceae samples (therein ten Sorbus species from different localities), the best values in FRAP and DMPD tests were obtained in case of 75% ethanol extract of S. umbellata and S. subfusca leaves, respectively. In turn, S. kusnetzovii leaf extract exerted the strongest metal-chelating capacity (27.13 ± 2.01% at 2000 µg/mL), however still not so high as EDTA (97.66 ± 0.12% at 2000 µg/mL).
Although, the antioxidant activities of S. lanata and S. cashmiriana have not been investigated, there are three papers referring to the activity of their constituents. The former afforded three new phenols (sorlanin, sorbanin and sorbalanin) and a new triterpene—sorbanolic acid. In the latter two new coumarins, cashmin A and cashmin B, were found. Sorbanolic acid showed significant activity in DPPH assay (IC50 = 24.2 µM), whereas sorlanin, sorbanin and sorbalanin had poor IC50 values of 192.2 ± 2.9, 90.7 ± 0.5 and 445.9 ± 4.0 µM, respectively (ascorbic acid, IC50 = 33.9 µM) (Uddin et al. 2013; Latif et al. 2014). Cashmin A and cashmin B showed significant antioxidant activity in DPPH and ABTS assays. Their Fe+3 reducing and hydrogen peroxide scavenging activities were confirmed as well (Khan et al. 2015).
Antioxidant activity in cellular models
Antioxidant activity of S. domestica, S. aucuparia and S. hajastana were assayed using human plasma model. The water–methanol flower extract of S. aucuparia and its fractions effectively protected human plasma exposed to oxidative/nitrative stress. Not only the levels of the oxidative stress biomarkers (3-nitrotyrosine, lipid hydroperoxides and thiobarbituric acid-reactive substances) were found to be decreased, but also the non-enzymatic antioxidant capacity of blood plasma was enhanced (Olszewska et al. 2019). Likewise, the defatted S. domestica leaf extract (methanol), at the concentration range of 1–50 µg/mL, decreased 3-nitrotyrosine, lipid hydroperoxides and thiobarbituric acid-reactive substances levels in human plasma (Matczak et al. 2018). Moreover, nine predominant polyphenols ((–)-epicatechin, procyanidin B2, procyanidin C1, rutin, quercitrin, quercetin, quercetin 3-O-(2″-O-β-d-glucopyranosyl)-α-l-rhamnopyranoside, quercetin 3-O-(2″-O-β-d-xylopyranosyl)-α-l-rhamnopyranoside and chlorogenic acid)—present in the leaf extract—significantly reduced lipid peroxidation and protein nitration as well as enhanced/normalized the non-enzymatic antioxidant capacity of blood plasma in the FRAP test (Rutkowska et al. 2019b). Finally, S. hajastana reduced lipid peroxidation in human plasma (Manukyan et al. 2019).
In another study, inhibitory effects of S. amurensis bark on lipid peroxidation in brain homogenates and erythrocyte hemolysis were assayed. The biological material, both erythrocytes and brains, were obtained from Sprague–Dawley rats. Here, n-butanol fraction of S. amurensis methanol extract was found to be active. Erythrocyte hemolysis decreased by 64.6 ± 2.0% and 78.9 ± 2.1% after incubation with the extract at the concentration of 100 and 200 μg/mL, respectively. In turn, lipid peroxide generation was reduced by 90.7 ± 0.7% at 200 μg/mL (Kang et al. 2003). In another study, methanol extract from the fruits of S. aucuparia moderately (8.21 ± 0.64%) prevented lipid peroxidation (homogenized rat liver) (Mlcek et al. 2014).
Antidiabetic activity
Among all Sorbus species, S. decora is most studied for potential antidiabetic activity as its bark was traditionally used to treat symptoms of diabetes. Therefore, surveys have appeared describing activity of the bark (or inner bark) extracts both in vitro and in vivo. Spoor et al. (2006) revealed that the ethanol extract from inner bark enhanced glucose uptake in C2C12 cells. Moreover, after 18–21 h incubation with the plant extract, the effect was comparable to that of a positive control (metformin in dose 400 μmol/L). What is noteworthy, no influence on basal or glucose-stimulated insulin secretion was observed in INS832/13 rat insulinoma cells, what suggests that antidiabetic effects were not related to affecting β-cells. Likewise, no glitazone-like activity was seen as triglyceride accumulation was not increased in differentiating 3T3-L1 adipocytes treated with the plant extract. Nevertheless, S. decora notably protected PC12-AC cells against glucose toxicity under high glucose conditions. Indeed, in another study, the ethanol extract at the concentration of 15 μg/mL increased glucose uptake in C2C12 cells after 18 h of incubation and inhibited glucose-6-phosphatase enzyme activity in H4IIE liver cells (Shang et al. 2015). Next, Martineau et al. (2010) studied mechanisms of glucose uptake improvement in C2C12 myoblasts by extracts from several Canadian species. The results indicate, that extracts, including that from the inner bark of S. decora, act through metformin-like mechanism, where the AMPK pathway is activated. We would like to underline, that also a pure compound isolated from stem bark of S. decora, namely 23,28-dihydroxylupan-20(29)-ene-3β-caffeate, significantly potentiated glucose uptake in C2C12 skeletal muscle cells with EC50 = 1.47 μM (Guerrero-Analco et al. 2010). Finally, this very promising plant was examined in vivo using genetic KK-Ay Type 2 diabetic mice, male Sprague–Dawley rats with streptozotocin-induced type 1 diabetes and insulin resistant rats treated for that purpose with 10% glucose water for 6 weeks. Ethanol extract from S. decora significantly decreased glycemia in model of Type 1 diabetes and the effect was observed after just a single dose (200 mg/kg, i.g.). Furthermore, the increasing intensity of peak decreases in glycemia observed during the experiment suggests a cumulative effect. Nevertheless, the effectiveness of the extract was not so high as that of the reference drug (metformin, 500 mg/kg, i.g.). On the other hand, modest hyperglycemia and hyperinsulinemia, observed in insulin-resistant model, were normalized when rats received S. decora extract (200 mg/kg/day, i.g.). Also, the HOMA insulin resistance parameter was reduced and the extract was as effective as metformin. In KK-Ay Type 2 diabetic mice, the glycemia decreased by 15% after 7 days of treatment with the extract in dose 200 mg/kg/day (incorporated into food to avoid stressing animals) (Vianna et al. 2011).
Reports on the antidiabetic potential of Sorbus berries can also be found in the scientific literature. The diethyl ether and ethyl acetate fractions derived from methanol extracts of S. domestica fruits (differing in maturity stage) showed strong aldose reductase activity ranging from 72 to 93% at the concentration of 50 μg/mL (Termentzi et al. 2008a). Also a component of S. domestica fruits—(1S,3R,4S,5R)5-O-caffeoylquinic acid—showed an ability to reduce glucose uptake by HepG2 cells. This isomer of chlorogenic acid exerted hypolipidemic properties as well (Forino et al. 2015). In turn, Boath et al. (2012) revealed that polyphenol-rich extract of S. aucuparia inhibited α-glucosidase with IC50 = 30 μg GAE/mL, which was comparable to acarbose.
It is probable that in the case of S. commixta bark and S. pohuashanensis fruits, triterpene constituents may at least in part contribute to the observed effects of extracts. Lupenone and lupeol (present in stem bark of S. commixta) inhibited noncompetitively PTP1B with IC50 = 13.7 ± 2.1 and 5.6 ± 0.9 μM (Na et al. 2009). Also, 3β-acetoxy-urs-12-ene-28-oic acid, pomolic acid-3β-acetate and betulinic acid from S. pohuashanensis fruits had IC50 values of 4.8 ± 0.5, 6.1 ± 0.3 and 3.5 ± 0.1 μM, respectively. Ursolic acid, a common triterpene in many Sorbus species (see Table 5) which was used as the positive control in this study, is a known PTP1B inhibitor (IC50 = 3.4 ± 0.1 μM) (Li et al. 2014).
Anti-inflammatory activity
Anti-inflammatory activity of S. commixta was confirmed both in vitro and in vivo. Regarding in vitro studies on LPS-stressed RAW 264.7 macrophages, 70% ethanol extract from S. commixta effectively inhibited NO and PGE2 production (no effect on TNF-α) with concomitant influence on iNOS and COX-2 mRNA levels (Yu et al. 2009). Dose-dependent suppression at the transcriptional level of NO and PGE2 production in LPS-induced RAW 264.7 cells was also observed for water extract from inner stem bark. Moreover, the extract was next tested in vivo and caused a significant reduction in arachidonic acid-induced ear edema in mice (pre-treated for 7 days by orally administered extract at the dose of 100 mg/kg or indomethacin 1 mg/kg) (Yu et al. 2011). What is noteworthy, β-sitosteryl-3-O-β-glucopyranoside, the main component of the bark of S. commixta, showed anti-inflammatory activity both in vitro and in vivo. The compound reduced NO, PGE2, TNF-α, IL-1β and IL-6 production (together with iNOS and COX-2 expressions) and suppressed NF-kB activation in LPS-induced RAW 264.7 cells. Regarding the results of in vivo study in a carrageenan-induced mouse paw edema model, the edema rate was decreased by 63.2% and 32.4% after 3 h of β-sitosteryl-3-O-β-glucopyranoside administration at the dose of 25 mg/kg (p.o) and 100 mg/kg (p.o), respectively. The results were compared to ibuprofen (100 mg/kg, p.o.), which was used as drug reference and at that time reduced the edema rate by 50.4% (Yang and An 2014). Another isolate of S. commixta bark, namely (9S,12R,13R)-(E)-9,12,13-trihydroxy-10-octadecaenoic acid, reduced NO production in LPS-stimulated BV-2 cells with an IC50 = 71.25 μM (Kim et al. 2016). Contrary to S. commixta, S. alnifolia (ethanol–water extract from leaves, branches and fruits) showed poor inhibitory effect on NO production in activated RAW 264.7 macrophages with IC50 value > 500 µg/mL (Kim et al. 2010).
Some authors determined inhibitory activity of Sorbus species on lipoxygenase and hyaluronidase. Matczak et al. (2018) examined defatted (pre-extracted with chloroform) water–methanol extract from S. domestica leaves. The influence of both the extract and its fractions was analyzed and compared. The results showed that the most potent lipoxygenase inhibitor was ethyl acetate fraction with IC50 value 115.54 ± 4.99 µg/mL (which was comparable to indomethacin, IC50 = 92.60 ± 3.71 µg/mL) and the most active towards hyaluronidase was n-butanol fraction with IC50 = 11.06 ± 0.30 µg/mL (indomethacin: IC50 = 12.77 ± 0.91 µg/mL). Indeed, subsequent studies confirmed the inhibitory activity of extract components against the tested enzymes (Rutkowska et al. 2019b). Also, n-butanol fraction of S. aucuparia water–methanol flower extract revealed strong anti-hyaluronidase activity (even stronger than indomethacin), whereas its ethyl acetate and diethyl ether fractions were potent lipoxygenase inhibitors. All extracts expressed no influence on xanthine oxidase (Olszewska et al. 2019). One paper refers to the activity of cashmirol A and cashmirol B, two new triterpenes from S. cashmiriana. These compounds were found to be moderate lipoxygenase inhibitors in in vitro assay with IC50 values 90.2 and 74.9 µM, respectively (baicalein was used as positive control, IC50 = 8.0 µM) (Kazmi et al. 2009).
Antimicrobial activity
The majority of papers referring to antimicrobial activity of Sorbus species provide data on the extracts from S. aucuparia fruits. Aqueous and 50% ethanol extracts inhibited the growth of Bacillus cereus MSCL 330, Staphylococcus aureus MSCL 334 and Pseudomonas aeruginosa MSCL 331 (against the latter, only extracts from fresh fruits were active) but no effects on Escherichia coli MSCL 332 and Candida albicans MSCL 378 were observed (Liepiņa et al. 2013). In another study, phenolic-rich fractions (SPE-purified acetone extracts) of wild S. aucuparia and its cultivars (Burka, Zoltaja, Titan, Granatnaja) showed a weak bacteriostatic activity against Staphylococcus aureus VTT E-70045. Zoltaja and Granatnaja inhibited weakly growth of Escherichia coli VTT E-94564T and Zoltaja slightly retarded Salmonella enterica sv. Typhimurium VTT E-981151 growth. In addition, extracts from wild S. aucuparia and the Burka cultivar affected M hemagglutinin-mediated hemagglutination of E. coli HB101 (pRR7) at concentrations of 1–2 μg and 0.5–1 μg of total phenolics/mL, respectively (Kylli et al. 2010). Krisch et al. (2008) screened antibacterial activity of juices as well as water and methanol pomace extracts from various berries against Bacillus subtilis ssp. subtilis BD 170, B. cereus var. mycoides ATCC 9634, Escherichia coli SZMC 0582 and Serratia marcescens SZMC 0567. In general, S. aucuparia next to Ribes nigrum and Cornus mas exhibited the best inhibition capacity. Nohynek et al. (2006) examined polyphenolic-rich fraction of S. aucuparia towards Clostridium perfringens E-861T, Campylobacter jejuni E-1008T, Bacillus cereus E-727, Staphylococcus aureus E-045 and Candida albicans NCPF 3179, however S. aucuparia exhibited poor antimicrobial activity towards all tested strains with the exception of Bacillus cereus E-727. In a different study S. aucuparia fruit extracts (water and methanol) moderately affected the growth of Escherichia coli ATCC 25922 and Staphylococcus aureus ATCC 25923. Slightly better effects were seen in case of adequate extracts from fruits of S. torminalis f. torminalis and S. torminalis f. semitorminalis (Mrkonjić et al. 2017). Despite all above-mentioned studies, the antimicrobial activity of rowan fruits is still poorly argued and seems to be quite weak.
Kavak and Akdeniz (2019) examined antimicrobial activity of the leaf extract of Sorbus umbellata (Desf.) Fritsch var. umbellata. The extract suppressed the growth of Staphylococcus aureus ATCC29213 and Escherichia coli ATCC25922, but displayed no effect on Bacillus cereus ATCC7064, Bacillus subtilis ATCC 6633, Pseudomonas aeruginosa ATCC27853 and Listeria monocytogen ATCC 7644 (ampicillin and gentamicin used as controls). Moreover, the extract inhibited the bacterial β-glucuronidase with IC50 = 117.9 μg/mL.
In turn, Turumtay et al. (2017) compared activity of fruit and leaf DMSO extracts from S. aucuparia as well as S. caucasica var. yaltiriki against Gram-negative (Escherichia coli ATCC25922, Pseudomonas aeruginosa ATCC 27853, Salmonella typhimurium ATTC14028) and Gram-positive bacteria (Staphylococcus aureus ATCC25923, Bacillus subtilis ATCC6633, Enterococcus faecalis ATTC29212). Although the activity of the extracts varied, they were all inactive against Gram-positive bacteria and active against P. aeruginosa. In the same study, the effects of methanol extracts on both non-replicative (Klenow Fragment-KF and Bacillus Large Fragment-BLF) and replicative (DnaE and PolC) bacterial DNA polymerases were measured. S. aucuparia fruit extract inhibited all replicative and non-replicative DNA polymerases, whereas leaf extract was able to affect only the two replicative ones. In turn, leaf extract of S. caucasica var. yaltirikii showed activity against KF (strong inhibitory effect), DnaE and polC, while fruit extract inhibited only KF and DnaE.
Cytotoxic activity
Many authors performed cytotoxicity studies on Sorbus species to confirm cellular safety or potential anticancer activity of investigated analytes. Scientific literature provides information on cytotoxicity of extracts derived from various Sorbus berries. Fairly well documented is the activity of S. aucuparia fruits, which were examined both in vitro and in vivo. Boncler et al. (2017) used a screening assay for the assessment of cytotoxicity of various agents towards HepG2, Caco-2, A549, HMEC-1 and 3T3 cells. Three key cell indicators were measured: cell membrane integrity, mitochondrial membrane potential and nuclear size. S. aucuparia fruit extract exerted relatively high toxicity especially where the nuclear area was concerned. In another study, a polyphenol-rich extract from rowanberries reduced the viability of HeLa cells to about 50% at 50 µg of GAE/mL (McDougall et al. 2008). Also methylene chloride fraction of ethanol extract from S. aucuparia reduced the viability of HeLa cells in MTT assay with IC50 = 15 ± 03 μg/mL (plant organ not mentioned) (Bozkurt-Guzel et al. 2018). Goun et al. (2002) tested not only methylene chloride, but also methanol extracts (extraction in sequence) towards mouse leukemia cells (ATCC L1210) and only the first one was active (likewise, plant organ not mentioned). Mrkonjić et al. (2017) observed poor cytotoxic activity of water and methanol extracts from fruits of S. aucuparia (fruit jam was studied as well, but it was totally inactive). Cytotoxicity studies were performed towards HeLa, HT-29, MCF7 and normal MRC-5 cells in comparison to podophyllotoxin using the SRB assay. The IC50 values of all extracts ranged from 414 ± 12.5 to 965 ± 27.9 µg/mL and no selectivity of action was noted (podophyllotoxin, IC50 values from (1.30 ± 0.20) × 10−3 to (4.70 ± 0.80) × 10−3). Razina et al. (2016) examined in vivo antitumor activity of acidified 95% ethanol extract from S. aucuparia fruits standardized on anthocyanins. This extract was given to female C57BL/6 mice with intramuscularly transplanted melanoma B-16 and Lewis lung carcinoma cells (5 × 106 cells/0.1 mL saline). Therapy with the extract alone or in combination with cyclophosphamide markedly decreased the tumor growth in both cases. Similarly, Isaikina et al. (2018) revealed a potential antitumor activity of acidified 95% ethanol extract from rowanberries in C57BL/6 female mice with transferred Lewis lung carcinoma.
As regards reports on cytotoxicity of berries of other Sorbus species, ethanol extract from S. sambucifolia fruits at the dose of 5 × 103 µg/mL decreased HL-60 leukemic cells proliferation by 51% in Alamar Blue method. Also differentiation-inducing effects were observed (Yoshizawa et al. 2000). S. pohuashanensis fruits were tested against HT-29 and HepG2 cells and significant dose-dependent inhibition of cell proliferation was observed (Fan et al. 2011). Cytotoxicity of water–methanol extract from S. commixta fruits was evaluated on A549, H1264, H1299 and Calu-6 cells differing in p53 gene status with the help of WST-1 assay. The extract induced apoptotic process and this effect was independent of cellular p53 status (Lee et al. 2017). Finally, percent of HepG2 cells viability was 78.13% after incubation with S. hajastana extract at the concentration of 200 μg/mL (Alamar Blue assay) (Manukyan et al. 2019).
Even though reports on cytotoxic activity of extracts from plant parts other than fruits are definitely less numerous, some of them provided interesting results. Water extract from S. commixta stem and bark showed significant effects in Hep3B (from 250 µg/mL of the extract), HepG2 HCC cells (from 200 µg/mL), Chang liver cells (at 500 µg/mL) and HCT116 colon cancer cells (from 100 µg/mL). Moreover, the anti-invasive and anti-migration potential of this extract was confirmed. In metastatic Hep3B cells the matrix metalloproteinase-9 activity and expression was decreased, likewise the expression of the chemokine receptors. What is more, actin fiber arrangement was found to be suppressed (Park et al. 2017). Results from another study showed that the water extract (S. commixta stem and bark) inhibited cell viability in several liver and colon cancer cells, i.e. Hep3B, HepG2 and HCT116 cells with IC50 values of 300.1, 237.8 and 77.7 µg/mL, respectively. Further analysis on HCT116 cells revealed that this extract caused apoptosis via the ROS-mediated mitochondrial pathway (Moon et al. 2018). What is noteworthy, not only S. commixta extracts were examined, but also its components. Kim et al. (2016, 2018) and Choi et al. (2018) tested several isolates from the bark of S. commixta, including four new structures, namely 1,2,4-trimethoxydibenzofuran-3,9-diol, sorcomic acid, sorcomisides A and sorcomisides B, however these compounds were found to be inactive.
Ethanol–water leaf extract from Sorbus umbellata (Desf.) Fritsch var. umbellata displayed cytotoxic activity towards A549 and MCF-7 cells in MTT assay with the highest inhibition of cell proliferation observed for A549 cells (71.8% at 150 μg/mL) (Kavak and Akdeniz 2019).
Neuroprotective activity
Ethanol extract from the stem of S. commixta showed neuroprotective activity against Aβ42 toxicity both in vitro and in vivo. In case of in vivo study, Drosophila models were employed, where Aβ42-expressing flies showed Alzheimer’s disease-like phenotypes, including decreased survival rate and motility, malformation of the eye as well as increased cell death in the larval brain and increased ROS levels in the eye imaginal discs. The extract ameliorated such phenotypes and its neuroprotective activity was confirmed in vitro using Aβ-treated SH-SY5Y cells (Liu et al. 2015). It is also interesting to note that sorcomic acid, a new fatty acid from the bark of S. commixta, was itself tested for potential neuroprotective activity, which was assessed in vitro using an ELISA development kit. Sorcomic acid significantly induced NGF secretion in C6 glioma cells (233.40 ± 12.82%, percentage of the untreated control group) in comparison to a positive control, 6-shogaol (168.58 ± 7.16%) (Kim et al. 2016).
Cheon et al. (2017) tested whether methanol extract from stem and twigs of S. alnifolia has protective effects on MPP+-induced dopaminergic neurodegeneration. It was shown that the extract protected MPP+-treated PC12 cells with the viability values 63.61 ± 11.76, 79.93 ± 9.22 and 85.83 ± 8.74% (% of control) at 125, 250 and 500 μg/mL, respectively. Furthermore, several in vivo tests were performed using Caenorhabditis elegans. S. alnifolia extract restored the loss of viability of MPP+-treated worms by 10.62%, 21.85%, and 54.93% at 62.5, 125, and 250 μg/mL, respectively. Moreover, both chemically- and genetically-induced dopaminergic neurodegeneration were reduced (BZ555 and UA57 strains were used). Food-sensing assay showed that dopaminergic specific behavioural deficit was rescued in worms fed with S. alnifolia extract. Also, the influence of the extract on α-synuclein aggregation in C. elegans (transgenic strain NL5901) was evaluated, but no significant effect was observed.
Some Sorbus species were found to have an ability to inhibit AChE and/or BuChE. Methanol extracts (75%) from the leaves of several Sorbus species, namely S. aucuparia, S. caucasica, S. caucasica var. yaltrikii, S. kusnetzovii, S. persica, S. roopiana, S. subfusca, S. torminalis and S. umbellata, were examined for possible AChE and BChE inhibitory activities. All samples were tested at the concentrations of 200 µg/mL and compared to the reference drug (galanthamine, 100 µg/mL). The strongest inhibition of AChE was observed for S. umbellata extract (58.18 ± 3.77%, IC50 = 177.8 µg/mL), however this was a weak effect when compared to the reference compound galanthamine (IC50 = 1.78 µg/mL). Among all tested Sorbus species, S. subfusca reached the highest percent of BChE inhibition with 43.11 ± 3.24% (Ekin et al. 2016). Previously, Hasbal et al. (2015) observed moderate antiacetylcholinesterase activity of S. torminalis fruits water extract. Interestingly, in the study of Mrkonjić et al. (2017), neither of extracts from S. torminalis f. torminalis and f. semitorminalis was active (the authors tested water and methanol extracts as well as fruit jams). Nevertheless, the anti-AChE activity of adequate S. aucuparia extracts was noted (IC50 values ranging from (2.02 ± 0.02) × 103 to (3.81 ± 0.21) × 103 µg/mL; galanthamine, IC50 = 0.39 ± 0.01 µg/mL).
Hepatoprotective activity
Triterpenoid fraction obtained from S. pohuashanensis fruits was tested for potential hepatoprotective effect in animal model of APAP-induced liver injury (male ICR mice). Not only the levels of AST, ALT, TNF-α, IL-1β, IL-6, MDA, SOD, GSH, CAT were normalized after treatment, but also hepatic tissues necrosis, hemorrhage and infiltration of inflammatory cell were inhibited. Moreover, western-blot and RT-PCR analysis revealed suppressive effect on iNOS and COX-2 over-expressions. Finally, triterpenoid fraction reduced APAP-induced phosphorylation of MAPK family signals (Yin et al. 2019).
Lee et al. (2006) extracted S. commixta bark with 80% aqueous methanol and then partitioned the extract with n-hexane, chloroform, ethyl acetate n-butanol and water. Methanol extract and its fractions were than administered to alcohol-treated Sprague–Dawley rats (alcohol intake 3.0 g/kg). Significant decrease in blood alcohol concentration was observed when rats were treated with methanol extract at doses higher than 200 mg/kg. The highest activity were observed for the ethyl acetate fraction (rich in phenolics), which caused about 46% decrease 2 h post alcohol administration at the dose of 150 mg/kg. Moreover, increased MDA levels and decreased catalase activity were reversed by the ethyl acetate fraction but no statistically significant influence on ADH, ALT, AST, GSH levels or SOD activity was observed (however the last two markers were not affected by the alcohol treatment itself).
Cardioprotective activity
S. pohuashanensis flavonoid fraction showed a protective effect against ATO-induced cardiotoxicity in male BALB/c mice and H9c2 cells. ATO-induced apoptosis and oxidative stress were suppressed both in vivo and in vitro. Moreover, alterations of cardiac tissue such as myofibrillar loss, cytoplasmic vacuolization and cardiomyocyte necrosis were observed in the hearts of ATO-treated mice. Pretreatment with S. pohuashanensis flavonoids (20 mg/kg, intraperitoneal injection) alleviated these abnormalities. Also, the levels of creatine kinase, creatine kinase-MB, lactate dehydrogenase and glutamic oxaloacetic transaminase were found to be significantly reduced. (Yu et al. 2017).
Vasorelaxant and anti-atherogenic activities
To the best of our knowledge, only S. commixta bark extracts were tested for potential vasorelaxant and anti-atherogenic activities. Anti-atherogenic effects of methanol extract were assessed by Sohn et al. (2005a, b) using two rat models. In one case, atherosclerosis in male Sprague–Dawley rats was induced by administration of NG-nitro-l-arginine methyl ester, which inhibited NO production and led to inflammation. Over-expressions of MCP-1, NF-κB p65, adhesion molecules (including ICAM-1, VCAM-1, E-selectin), ET-1 and ACE as well as decreased ecNOS expression in aorta were observed. All these abnormalities were ameliorated by chronic treatment with the extract in doses 100 mg/kg/day and 200 mg/kg/day (Sohn et al. 2005a). In the second model male Sprague–Dawley rats were fed an atherogenic-diet. Likewise, increases in the expression of ET-1, NF-kB p65, E-selectin, ICAM-1 and VCAM-1 were reversed. Moreover, the extract (in dose 100 mg/kg/day and 200 mg/kg/day) improved aortic NOS/NO system dysfunction (Sohn et al. 2005b).
Regarding a potential vasorelaxant activity, n-butanol fraction of S. commixta stem bark methanol extract was tested using aortic rings isolated from healthy male Sprague–Dawley rats. First, the aortic tissues were contracted with phenylephrine (3 × 10−6 M). The extract at the concentration of 100 μg/mL caused 92.9 ± 1.9% relaxation. What is more, when NG-nitro-l-arginine methyl ester (NOS inhibitor) was added or functional endothelium was removed, the activity disappeared suggesting that vasorelaxant effect may be associated with endothelial NO signaling pathway (Yin et al. 2005). These findings were also confirmed by Kang et al. (2005). Here, methanol extract induced relaxant response of the phenylephrine-contracted aorta obtained from male Sprague–Dawley rats. Likewise, the endothelium-denuded aortic tissues were insensitive. Pretreatment with NG-nitro-l-arginine methyl ester or soluble guanylyl cyclase inhibitors (methylene blue and 1H-[1,2,4]-oxadiazole-[4,3-α]-quinoxalin-1-one) blocked the extract-induced relaxation. Moreover, cGMP production in the carotid artery and human umbilical vein endothelial cells was found to be increased after incubation with S. commixta extract. These findings suggest involvement of endothelium-dependent NO-cGMP signaling pathway.
Other activities
Exposure of human dermal fibroblasts to UVA radiation (6.3 J/cm2) caused increase in MMP-1 expression by approximately 41%, which led to degradation of collagen, gelatin etc. Ethanol extract from S. commixta bark affected MMP-1 expression in UVA-irradiated cells and showed photoprotective activity. Moreover, enzymatic conversion of crude extract enhanced this inhibitory effect. Thus, β-glucanase-treated extract at concentrations of 2.5, 5, 10, and 20 μg/mL reduced the expression level of MMP-1 by 7.4, 16.5, 32.4, and 55.9% respectively (Bae et al. 2007). In turn, Xuan et al. (2018) investigated the effect of S. commixta twig ethanol extract on MMPs levels in UVB-irradiated human dermal fibroblasts (80 mJ/cm2). The extract at concentrations of 12.5–50 μg/mL notably decreased UVB-induced MMP-1, MMP-2, MMP-3 and c-Fos expressions (no effect on c-Jun phosphorylation) as well as suppressed the intracellular ROS generation. These results suggest involvement of MAPK pathway.
Ethanol extracts from stem and leaves of S. commixta showed anti-lipase and anti-phosphodiesterase activity (IC50 values of 29.6 μg/mL and 20.08 μg/mL, respectively), however it was not so effective as the reference drug orlistat (IC50 = 0.076 μg/mL). Phosphodiesterase activity was evaluated using the PDE-GloTM phosphodiesterase activity kit whereas pancreatic lipase inhibitory activity was assayed by measuring the hydrolysis of p-nitrophenyl butyrate to p-nitrophenol (Lee et al. 2012).
Goun et al. (2002) revealed that methanol extract of S. aucuparia showed weak antithrombin activity (plant organ not mentioned), whereas Tam et al. (2009) and Cieniak et al. (2013) have found that S. decora ethanol extract (we can only assume that the extract was derived from the bark) may affect CYP-mediated metabolism of other drugs.
It is noteworthy that sorbicin A and sorbicin B, new triterpenes isolated from S. cashmiriana, inhibited urease as well as α-chymotrypsin in vitro. In urease inhibitory assay both compounds displayed inhibitory potential with IC50 values 85.2 ± 0.28 and 17.8 ± 0.12 μM, whereas thiourea—positive control—had an IC50 value of 21.6 ± 0.18 μM. Also, both compounds inhibited α-chymotrypsin with IC50 values 23.2 ± 0.09 and 22.7 ± 0.12 μM (positive control—chymostatin, IC50 = 7.2 ± 0.26 μM) (Kazmi et al. 2011).
Conclusions
As presented in this work, many Sorbus species have an established position in folk medicine. However, not all ethnomedical applications have been confirmed by scientific studies. Even though many species showed antioxidant activity, which can support their value in the treatment of many diseases, only in a few cases more targeted studies were conducted to confirm specific folk usage. One of the fairly well documented medicinal uses is the antidiabetic activity of the bark of S. decora. It is noteworthy that in this case not only in vitro tests were performed, but also in vivo studies in three animal models of insulin resistance and diabetes. Although reports indicate that S. domestica berries were also applied as antidiabetic remedies, their therapeutic properties do not have much support in scientific data. However, their aldose reductase activity was observed, which could help to prevent retinopathy and neuropathy in people with diabetes. On the other hand, hypoglycemic potential of (1S,3R,4S,5R)5-O-caffeoylquinic acid, which is a component of S. domestica fruits, seems to be noteworthy. S. commixta is another Sorbus species, the traditional medicinal usage of which seems to be explained by recent scientific findings. Its bark preparations have been used to treat diseases related to inflammation like asthma or bronchitis. Indeed, the anti-inflammatory effect of aqueous extracts was confirmed both in vitro and in vivo. Moreover, one in vivo study proved the efficacy of the bark of S. commixta in atherosclerosis. Nevertheless, most folk applications of Sorbus preparations are not supported by scientific evidence thus attempts should be made to confirm their effectiveness.
As mentioned above, biological studies have been conducted primarily on plant extracts. However, only in some papers (mainly those published in recent years) their chemical constituents were determined. More often, the authors measured the total phenolic content, but it still does not explain which compounds are responsible for the activity of whole extracts. A small share of in vivo as well as safety studies is also noticeable, which extends the area of potential additional analyzes.
Regarding phytochemical studies, they all revolved around a few most commonly found species. Thus, a large number of Sorbus representatives have not been studied as thoroughly, or at all. Moreover, an overwhelming amount of papers was devoted to identification of phenolic compounds, which means, that Sorbus species are worthwhile to further investigate phytochemically.
Abbreviations
- AAPH:
-
2,2′-Azobis-(2-amidinopropane) dihydrochloride
- ABTS:
-
2,2′-Azino-bis(3-ethylbenzothioazoline-6-sulphonic acid) diammonium salt
- ACE:
-
Angiotensin converting enzyme
- AChE:
-
Acetylcholinesterase
- ADH:
-
Alcohol dehydrogenase
- ALT:
-
Alanine aminotransferase
- AMPK:
-
Adenosine-monophosphate-activated protein kinase
- AST:
-
Aspartate transaminase
- ATO:
-
Arsenic trioxide
- BChE:
-
Butyrylcholinesterase
- CAT:
-
Catalase
- CD:
-
Conjugated dienes
- cGMP:
-
Guanosine 3′,5′-cyclic monophosphate
- COX-2:
-
Cyclooxygenase-2
- CYP:
-
Cytochrome P450
- DMPD+ :
-
N,N-Dimethyl-p-phenylenediamine
- DPPH:
-
2,2′-Diphenyl-1-picrylhydrazyl
- dw:
-
Dry weight
- ecNOS:
-
Endothelial cell NO synthase
- EDTA:
-
Ethylenediamine tetraacetic acid
- ELISA:
-
Enzyme-linked immunosorbent assay
- ET-1:
-
Endothelin-1
- FRAP:
-
Ferric reducing antioxidant power
- fw:
-
Fresh weight
- GAE:
-
Gallic acid equivalents
- GSH:
-
Glutathione
- HOMA:
-
Homeostasis model assessment
- ICAM-1:
-
Intercellular adhesion molecule-1
- IL-1β :
-
Interleukin-1β
- IL-6:
-
Interleukin-6
- i.g.:
-
Intragastric
- iNOS:
-
Inducible NO synthase
- LPS:
-
Lipopolysaccharide
- MAPK:
-
Mitogen-activated protein kinase
- MCP-1:
-
Monocyte chemoattractant protein-1
- MDA:
-
Malondialdehyde
- MMP-1:
-
Matrix metalloproteinase-1
- MMP-2:
-
Matrix metalloproteinase-2
- MMP-3:
-
Matrix metalloproteinase-3
- MMPs:
-
Matrix metalloproteinases
- MPP+ :
-
1-Methyl-4-phenylpyridinium ion
- MTT:
-
Methylthiazolyldiphenyl-tetrazolium bromide
- NBT:
-
Nitroblue tetrazolium
- NF-κB:
-
Nuclear factor-κB
- NGF:
-
Nerve growth factor
- NO:
-
Nitric oxide
- NOS:
-
Nitric oxide synthase
- PGE2 :
-
Prostaglandin E2
- p.o.:
-
Per os
- PTP1B:
-
Protein tyrosine phosphatase 1B
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
- SPE:
-
Solid phase extraction
- SRB:
-
Sulforhodamine B
- TBARS:
-
Thiobarbituric acid-reactive substances
- TEAC:
-
Trolox equivalent antioxidant capacity
- TNF-α :
-
Tumor necrosis factor-α
- UVA:
-
Ultraviolet A
- UVB:
-
Ultraviolet B
- VCAM-1:
-
Vascular cell adhesion molecule-1
References
Acuña UM, Atha DE, Ma J et al (2002) Antioxidant capacities of ten edible North American plants. Phyther Res 16:63–65. https://doi.org/10.1002/ptr.1031
Aladedunye F, Matthäus B (2014) Phenolic extracts from Sorbus aucuparia (L.) and Malus baccata (L.) berries: antioxidant activity and performance in rapeseed oil during frying and storage. Food Chem 159:273–281. https://doi.org/10.1016/j.foodchem.2014.02.139
Aladedunye F, Niehaus K, Bednarz H et al (2015) Enzymatic lipophilization of phenolic extract from rowanberry (Sorbus aucuparia) and evaluation of antioxidative activity in edible oil. LWT Food Sci Technol 60:56–62. https://doi.org/10.1016/j.lwt.2014.08.008
Aldasoro JJ, Aedo C, Navarro C, Garmendia FM (1998) The genus Sorbus (Maloideae, Rosaceae) in Europe and in North Africa: morphological analysis and systematics. Syst Bot 23:189–212. https://doi.org/10.2307/2419588
Aslantas R, Pirlak L, Güleryüz M (2007) The nutritional value of wild fruits from the North Eastern Anatolia Region of Turkey. Asian J Chem 19:3072–3078
Ayupbek A, Ke-lin H, Aisa HA (2012) Chemical constituents from the leaves of Sorbus tianschanica. Chem Nat Compd 48:133–134. https://doi.org/10.1007/s10600-012-0180-x
Bae J-T, Sim G-S, Kim J-H et al (2007) Antioxidative activity of the hydrolytic enzyme treated Sorbus commixta Hedl. and its inhibitory effect on matrix metalloproteinase-1 in UV irradiated human dermal fibroblasts. Arch Pharm Res 30:1116–1123. https://doi.org/10.1007/BF02980246
Becerra-Herrera M, Lazzoi MR, Sayago A et al (2015) Extraction and determination of phenolic compounds in the berries of Sorbus americana Marsh and Lonicera oblongifolia (Goldie) Hook. Food Anal Methods 8:2554–2559. https://doi.org/10.1007/s12161-015-0151-5
Bhatt LR, Bae MS, Kim BM et al (2009) A chalcone glycoside from the fruits of Sorbus commixta Hedl. Molecules 14:5323–5327. https://doi.org/10.3390/molecules14125323
Boath AS, Stewart D, McDougall GJ (2012) Berry components inhibit α-glucosidase in vitro: synergies between acarbose and polyphenols from black currant and rowanberry. Food Chem 135:929–936. https://doi.org/10.1016/j.foodchem.2012.06.065
Boncler M, Golanski J, Lukasiak M et al (2017) A new approach for the assessment of the toxicity of polyphenol-rich compounds with the use of high content screening analysis. PLoS ONE 12:1–26. https://doi.org/10.1371/journal.pone.0180022
Bozkurt-Guzel C, Serbetci T, Kultur S (2018) Cytotoxic activities of some Turkish medicinal plants against HeLa cells in vitro. Indian J Tradit Knowl 17:43–49
Cheon S-M, Jang I, Lee M-H et al (2017) Sorbus alnifolia protects dopaminergic neurodegeneration in Caenorhabditis elegans. Pharm Biol 55:481–486. https://doi.org/10.1080/13880209.2016.1251468
Chester M, Cowan RS, Fay MF, Rich TCG (2007) Parentage of endemic Sorbus L. (Rosaceae) species in the British Isles: evidence from plastid DNA. Bot J Linn Soc 154:291–304. https://doi.org/10.1111/j.1095-8339.2007.00669.x
Chizzali C, Beerhues L (2012) Phytoalexins of the Pyrinae: biphenyls and dibenzofurans. Beilstein J Org Chem 8:613–620. https://doi.org/10.3762/bjoc.8.68
Choi H-J (2017) In vitro antiviral activity of sakuranetin against human rhinovirus 3. Osong Public Heal Res Perspect 8:415–420. https://doi.org/10.24171/j.phrp.2017.8.6.09
Choi SY, Jeong B, Jang HS et al (2018) A new dibenzofuran from the barks of Sorbus commixta. Rec Nat Prod 12:179–183. https://doi.org/10.25135/rnp.20.17.06.112
Cieniak C, Liu R, Fottinger A et al (2013) In vitro inhibition of metabolism but not transport of gliclazide and repaglinide by Cree medicinal plant extracts. J Ethnopharmacol 150:1087–1095. https://doi.org/10.1016/j.jep.2013.10.029
Cobzac S, Cimpan G, Olah N, Gocan S (1999) The quantitative determination of rutin in different glycerinic plant extracts by solid-phase extraction and thin-layer chromatography with densitometry. J Planar Chromatogr 12:26–29
Dehaen W, Mashentseva AA, Seitembetov TS (2011) Allobetulin and its derivatives: synthesis and biological activity. Molecules 16:2443–2466. https://doi.org/10.3390/molecules16032443
Deren’ko SA, Suprunov NI (1979) Ursolic acid from the fruit of Sorbus aucuparia. Chem Nat Compd 15:363. https://doi.org/10.1007/BF00566103
Dickinson TA, Lo E, Talent N (2007) Polyploidy, reproductive biology, and Rosaceae: understanding evolution and making classifications. Plant Syst Evol 266:59–78. https://doi.org/10.1007/s00606-007-0541-2
Egea I, Sánchez-Bel P, Romojaro F, Pretel MT (2010) Six edible wild fruits as potential antioxidant additives or nutritional supplements. Plant Foods Hum Nutr 65:121–129. https://doi.org/10.1007/s11130-010-0159-3
Ekin HN, Gokbulut A, Aydin ZU et al (2016) Insight into anticholinesterase and antioxidant potential of thirty-four Rosaceae samples and phenolic characterization of the active extracts by HPLC. Ind Crops Prod 91:104–113. https://doi.org/10.1016/j.indcrop.2016.06.029
Erdtman H, Eriksson G, Norin T (1963) Aucuparin and methoxyaucuparin, two phenolic biphenyl derivatives from the heartwood of Sorbus aucuparia (L.). Acta Chem Scand 17:1151–1156
Fan Z-L, Wang Z-Y, Liu J-R (2011) Cold-field fruit extracts exert different antioxidant and antiproliferative activities in vitro. Food Chem 129:402–407. https://doi.org/10.1016/j.foodchem.2011.04.091
Forino M, Tenore GC, Tartaglione L et al (2015) (1S,3R,4S,5R)5-O-caffeoylquinic acid: isolation, stereo-structure characterization and biological activity. Food Chem 178:306–310. https://doi.org/10.1016/j.foodchem.2015.01.109
Fraser M-H, Cuerrier A, Haddad PS et al (2007) Medicinal plants of Cree communities (Québec, Canada): antioxidant activity of plants used to treat type 2 diabetes symptoms. Can J Physiol Pharmacol 85:1200–1214. https://doi.org/10.1139/Y07-108
Gaivelyte K, Jakstas V, Razukas A, Janulis V (2013) Variation in the contents of neochlorogenic acid, chlorogenic acid and three quercetin glycosides in leaves and fruits of rowan (Sorbus) species and varieties from collections in Lithuania. Nat Prod Commun 8:1105–1110. https://doi.org/10.1177/1934578X1300800819
Gaivelyte K, Jakstas V, Razukas A, Janulis V (2014) Variation of quantitative composition of phenolic compounds in rowan (Sorbus aucuparia L.) leaves during the growth season. Nat Prod Res 28:1018–1020
Ganhão R, Estévez M, Kylli P et al (2010) Characterization of selected wild mediterranean fruits and comparative efficacy as inhibitors of oxidative reactions in emulsified raw pork burger patties. J Agric Food Chem 58:8854–8861. https://doi.org/10.1021/jf101646y
Gil-Izquierdo A, Mellenthin A (2001) Identification and quantitation of flavonols in rowanberry (Sorbus aucuparia L.) juice. Eur Food Res Technol 213:12–17. https://doi.org/10.1007/s002170100328
Goun EA, Petrichenko VM, Solodnikov SU et al (2002) Anticancer and antithrombin activity of Russian plants. J Ethnopharmacol 81:337–342. https://doi.org/10.1016/S0378-8741(02)00116-2
Gu H, Chen F, Zhang Q, Zang J (2016) Application of ionic liquids in vacuum microwave-assisted extraction followed by macroporous resin isolation of three flavonoids rutin, hyperoside and hesperidin from Sorbus tianschanica leaves. J Chromatogr B 1014:45–55. https://doi.org/10.1016/j.jchromb.2016.01.045
Guerrero-Analco JA, Martineau L, Saleem A et al (2010) Bioassay-guided isolation of the antidiabetic principle from Sorbus decora (Rosaceae) used traditionally by the eeyou istchee Cree first nations. J Nat Prod 73:1519–1523. https://doi.org/10.1021/np1003005
Häkkinen SH, Kärenlampi SO, Heinonen IM et al (1999) Content of the flavonols quercetin, myricetin, and kaempferol in 25 edible berries. J Agric Food Chem 47:2274–2279. https://doi.org/10.1021/jf9811065
Hamston TJ, de Vere N, King RA et al (2018) Apomixis and hybridization drives reticulate evolution and phyletic differentiation in Sorbus L.: implications for conservation. Front Plant Sci 871:1–13. https://doi.org/10.3389/fpls.2018.01796
Hasbal G, Yilmaz-Ozden T, Can A (2015) Antioxidant and antiacetylcholinesterase activities of Sorbus torminalis (L.) Crantz (wild service tree) fruits. J Food Drug Anal 23:57–62. https://doi.org/10.1016/j.jfda.2014.06.006
Hukkanen AT, Pölönen SS, Kärenlampi SO, Kokko HI (2006) Antioxidant capacity and phenolic content of sweet rowanberries. J Agric Food Chem 54:112–119. https://doi.org/10.1021/jf051697g
Isaikina NV, Kalinkina GI, Razina TG et al (2018) Sorbus aucuparia L. fruit is a source of the drug for increasing the efficiency of tumor chemotherapy. Russ J Bioorgan Chem 44:899–905. https://doi.org/10.1134/S1068162018070038
Jarić S, Mačukanović-Jocić M, Djurdjević L et al (2015) An ethnobotanical survey of traditionally used plants on Suva planina mountain (south-eastern Serbia). J Ethnopharmacol 175:93–108. https://doi.org/10.1016/j.jep.2015.09.002
Johansson A, Laakso P, Kallio H (1997) Characterization of seed oils of wild, edible Finnish berries. Lebensm Unters Forsch 204:300–307. https://doi.org/10.1007/s002170050081
Kähkönen MP, Hopia AI, Vuorela HJ et al (1999) Antioxidant activity of plant extracts containing phenolic compounds. J Agric Food Chem 47:3954–3962. https://doi.org/10.1021/jf990146l
Kähkönen MP, Hopia AI, Heinonen M (2001) Berry phenolics and their antioxidant activity. J Agric Food Chem 49:4076–4082. https://doi.org/10.1021/jf010152t
Kalle R, Sõukand R (2012) Historical ethnobotanical review of wild edible plants of Estonia (1770s–1960s). Acta Soc Bot Pol 81:271–281
Kang DG, Keun Yun C, Lee HS (2003) Screening and comparison of antioxidant activity of solvent extracts of herbal medicines used in Korea. J Ethnopharmacol 87:231–236. https://doi.org/10.1016/S0378-8741(03)00142-9
Kang DG, Lee JK, Choi DH et al (2005) Vascular Relaxation by the Methanol Extract of Sorbus Cortex via NO-cGMP Pathway. Biol Pharm Bull 28:860–864. https://doi.org/10.1248/bpb.28.860
Kavak DD, Akdeniz B (2019) Sorbus umbellata (Desf.) Fritsch var. umbellata leaves: optimization of extraction conditions and investigation antimicrobial, cytotoxic, and β-glucuronidase inhibitory potential. Plant Foods Hum Nutr 74:364–369. https://doi.org/10.1007/s11130-019-00743-9
Kazmi MH, Ahmed E, Hameed S et al (2007) Isolation and structural determination of sorbinols A and B, new triterpenes from Sorbus cashmariana, by 1D and 2D NMR spectroscopy. Magn Reson Chem 45:416–419. https://doi.org/10.1002/mrc.1961
Kazmi MH, Ahmed E, Hameed S et al (2009) Cashmirols A and B, new Lipoxygenase Inhibiting Triterpenes from Sorbus cashmiriana. Chem Biodivers 6:1471–1476. https://doi.org/10.1002/cbdv.200800213
Kazmi MH, Fatima I, Malik A et al (2011) Sorbicins A and B, new urease and serine protease inhibitory triterpenes from Sorbus cashmiriana. J Asian Nat Prod Res 13:1081–1086. https://doi.org/10.1080/10286020.2011.626405
Khan S, Fatima I, Kazmi MH et al (2015) Cashmins A and B, Potent Antioxidant Coumarins from Sorbus cashmiriana. Chem Nat Compd 51:626–629. https://doi.org/10.1007/s10600-015-1370-0
Kim H, Song M-J (2012) Traditional plant-based therapies for respiratory diseases found in North Jeolla Province, Korea. J Altern Complement Med 18:287–293. https://doi.org/10.1089/acm.2010.0848
Kim H, Song M-J (2013) Ethnomedicinal practices for treating liver disorders of local communities in the Southern Regions of Korea. Evid Based Complement Altern Med 2013:1–11. https://doi.org/10.1155/2013/869176
Kim M-B, Park J-S, Lim S-B (2010) Antioxidant activity and cell toxicity of pressurised liquid extracts from 20 selected plant species in Jeju, Korea. Food Chem 122:546–552. https://doi.org/10.1016/j.foodchem.2010.03.007
Kim CS, Suh WS, Subedi L et al (2016) Neuroprotective fatty acids from the stem bark of Sorbus commixta. Lipids 51:989–995. https://doi.org/10.1007/s11745-016-4172-7
Kim CS, Oh J, Subedi L et al (2018) Two new phenolic glycosides from Sorbus commixta. Chem Pharm Bull 66:839–842. https://doi.org/10.1248/cpb.c18-00280
Klavins L, Kviesis J, Steinberga I et al (2016) Gas chromatography-mass spectrometry study of lipids in northern berries. Agron Res 14:1328–1346
Klensporf-Pawlik D, Przybylski R (2015) Antioxidant activity of selected wild Canadian prairie fruits. Acta Sci Pol Technol Aliment 14:357–366. https://doi.org/10.17306/J.AFS.2015.4.35
Krisch J, Galgóczy L, Tölgyesì M et al (2008) Effect of fruit juices and pomace extracts on the growth of Gram-positive and Gram-negative bacteria. Acta Biol Szeged 52:267–270
Krivoruchko EV, Andrushchenko OA, Kononenko AV (2013) Carboxylic acids from Sorbus aucuparia and S. aria. Chem Nat Compd 49:742–743. https://doi.org/10.1007/s10600-013-0725-7
Kültür Ş (2007) Medicinal plants used in Kırklareli Province (Turkey). J Ethnopharmacol 111:341–364. https://doi.org/10.1016/j.jep.2006.11.035
Kylli P, Nohynek L, Puupponen-Pimiä R et al (2010) Rowanberry phenolics: compositional analysis and bioactivities. J Agric Food Chem 58:11985–11992. https://doi.org/10.1021/jf102739v
Lapinskii AG, Gorbachev VV (2006) The antiradical activity of extracts from some wild-growing plants of the Okhotsk Sea northern coastal region. Pharm Chem J 40:317–319. https://doi.org/10.1007/s11094-006-0118-3
Latif A, Hussain SH, Ali M et al (2014) A New Antioxidant Triterpenoid from the Stem Wood of Sorbus lanata. Rec Nat Prod 8:19–24
Leduc C, Coonishish J, Haddad P, Cuerrier A (2006) Plants used by the Cree Nation of Eeyou Istchee (Quebec, Canada) for the treatment of diabetes: a novel approach in quantitative ethnobotany. J Ethnopharmacol 105:55–63. https://doi.org/10.1016/j.jep.2005.09.038
Lee S-O, Lee HW, Lee I-S, Im HG (2006) The pharmacological potential of Sorbus commixta cortex on blood alcohol concentration and hepatic lipid peroxidation in acute alcohol-treated rats. J Pharm Pharmacol 58:685–693. https://doi.org/10.1211/jpp.58.5.0014
Lee YM, Kim YS, Lee Y et al (2012) Inhibitory activities of pancreatic lipase and phosphodiesterase from Korean medicinal plant extracts. Phyther Res 26:778–782. https://doi.org/10.1002/ptr.3644
Lee TK, Roh H-S, Yu JS et al (2017) A novel cytotoxic activity of the fruit of Sorbus commixta against human lung cancer cells and isolation of the major constituents. J Funct Foods 30:1–7. https://doi.org/10.1016/j.jff.2017.01.003
Li H, Matsuura M, Li W et al (2012) Chemical constituents from the fruits of Sorbus pohuashanensis. Biochem Syst Ecol 43:166–168. https://doi.org/10.1016/j.bse.2012.03.011
Li D, Li W, Higai K, Koike K (2014) Protein tyrosine phosphatase 1B inhibitory activities of ursane- and lupane-type triterpenes from Sorbus pohuashanensis. J Nat Med 68:427–431. https://doi.org/10.1007/s11418-013-0804-x
Li M, Ohi-Toma T, Gao Y-D et al (2017) Molecular phylogenetics and historical biogeography of Sorbus sensu stricto (Rosaceae). Mol Phylogenet Evol 111:76–86. https://doi.org/10.1016/j.ympev.2017.03.018
Liepiņa I, Nikolajeva V, Jākobsone I (2013) Antimicrobial activity of extracts from fruits of Aronia melanocarpa and Sorbus aucuparia. Environ Exp Biol 11:195–199. https://doi.org/10.22364/eeb
Liu QF, Lee JH, Kim Y-M et al (2015) In vivo screening of traditional medicinal plants for neuroprotective activity against Aβ42 cytotoxicity by using Drosophila models of Alzheimer’s disease. Biol Pharm Bull 38:1891–1901. https://doi.org/10.1248/bpb.b15-00459
Łuczaj Ł, Dolina K (2015) A hundred years of change in wild vegetable use in southern Herzegovina. J Ethnopharmacol 166:297–304. https://doi.org/10.1016/j.jep.2015.02.033
Łuczaj Ł, Köhler P, Pirożnikow E et al (2013) Wild edible plants of Belarus: from Rostafiński’s questionnaire of 1883 to the present. J Ethnobiol Ethnomed 9:1–21. https://doi.org/10.1186/1746-4269-9-21
Manukyan A, Lumlerdkij N, Heinrich M (2019) Caucasian endemic medicinal and nutraceutical plants: in vitro antioxidant and cytotoxic activities and bioactive compounds. J Pharm Pharmacol 71:1152–1161. https://doi.org/10.1111/jphp.13093
Martineau LC, Adeyiwola-Spoor DCA, Vallerand D et al (2010) Enhancement of muscle cell glucose uptake by medicinal plant species of Canada’s native populations is mediated by a common, Metformin-like mechanism. J Ethnopharmacol 127:396–406. https://doi.org/10.1016/j.jep.2009.10.026
Matczak M, Marchelak A, Michel P et al (2018) Sorbus domestica L. leaf extracts as functional products: phytochemical profiling, cellular safety, pro-inflammatory enzymes inhibition and protective effects against oxidative stress in vitro. J Funct Foods 40:207–218. https://doi.org/10.1016/j.jff.2017.10.046
McCune LM, Johns T (2002) Antioxidant activity in medicinal plants associated with the symptoms of diabetes mellitus used by the Indigenous Peoples of the North American boreal forest. J Ethnopharmacol 82:197–205. https://doi.org/10.1016/S0378-8741(02)00180-0
McCune LM, Johns T (2003) Symptom-specific antioxidant activity of boreal diabetes treatments. Pharm Biol 41:362–370. https://doi.org/10.1076/phbi.41.5.362.15942
McCune LM, Johns T (2007) Antioxidant activity relates to plant part, life form and growing condition in some diabetes remedies. J Ethnopharmacol 112:461–469. https://doi.org/10.1016/j.jep.2007.04.006
McDougall GJ, Ross HA, Ikeji M, Stewart D (2008) Berry extracts exert different antiproliferative effects against cervical and colon cancer cells grown in vitro. J Agric Food Chem 56:3016–3023. https://doi.org/10.1021/jf073469n
Mikulic-Petkovsek M, Schmitzer V, Slatnar A et al (2012) Composition of sugars, organic acids, and total phenolics in 25 wild or cultivated berry species. J Food Sci 77:C1064–C1070. https://doi.org/10.1111/j.1750-3841.2012.02896.x
Mikulic-Petkovsek M, Krska B, Kiprovski B, Veberic R (2017) Bioactive components and antioxidant capacity of fruits from nine Sorbus genotypes. J Food Sci 82:647–658. https://doi.org/10.1111/1750-3841.13643
Mlcek J, Rop O, Jurikova T et al (2014) Bioactive compounds in sweet rowanberry fruits of interspecific Rowan crosses. Cent Eur J Biol 9:1078–1086. https://doi.org/10.2478/s11535-014-0336-8
Moon S-C, Choi H-J, Chung T-W et al (2018) Sorbus commixta water extract induces apoptotic cell death via a ROS-dependent pathway. Oncol Lett 16:4193–4200. https://doi.org/10.3892/ol.2018.9217
Mrkonjić ZO, Nađpal JD, Beara IN et al (2017) Phenolic profiling and bioactivities of fresh fruits and jam of Sorbus species. J Serbian Chem Soc 82:651–664. https://doi.org/10.2298/JSC170202049M
Na M, An R, Lee S et al (2002a) Antioxidant compounds from the stem bark of Sorbus commixta. Nat Prod Sci 8:26–29
Na M, An R, Min BS et al (2002b) Chemical Constituents from Sorbus commixta. Nat Prod Sci 8:62–65
Na M, Kim BY, Osada H, Ahn JS (2009) Inhibition of protein tyrosine phosphatase 1B by lupeol and lupenone isolated from Sorbus commixta. J Enzyme Inhib Med Chem 24:1056–1059. https://doi.org/10.1080/14756360802693312
Narasimhachari N, von Rudloff E (1962) The chemical composition of the wood extractives of Sorbus decora (Sarg.) Schneid. Can J Chem 40:1118–1122. https://doi.org/10.1139/v62-170
Neves JM, Matos C, Moutinho C et al (2009) Ethnopharmacological notes about ancient uses of medicinal plants in Trás-os-Montes (northern of Portugal). J Ethnopharmacol 124:270–283. https://doi.org/10.1016/j.jep.2009.04.041
Nohynek LJ, Alakomi H-L, Kähkönen MP et al (2006) Berry phenolics: antimicrobial properties and mechanisms of action against severe human pathogens. Nutr Cancer 54:18–32. https://doi.org/10.1207/s15327914nc5401_4
Olczyk M, Geszprych A (2017) Edible and medicinal plants of the genus Sorbus L. Postępy Fitoter 18:278–285. https://doi.org/10.25121/PF.2017.18.4.278
Olszewska M (2008) Separation of quercetin, sexangularetin, kaempferol and isorhamnetin for simultaneous HPLC determination of flavonoid aglycones in inflorescences, leaves and fruits of three Sorbus species. J Pharm Biomed Anal 48:629–635. https://doi.org/10.1016/j.jpba.2008.06.004
Olszewska MA (2009) Flavonoid profile of Sorbus intermedia. Chem Nat Compd 45:722–724. https://doi.org/10.1007/s10600-009-9409-8
Olszewska MA (2011a) Variation in the phenolic content and in vitro antioxidant activity of Sorbus aucuparia leaf extracts during vegetation. Acta Pol Pharm Drug Res 68:937–944
Olszewska MA (2011b) In vitro antioxidant activity and total phenolic content of the inflorescence, leaves and fruits of Sorbus torminalis (L.) Crantz. Acta Pol Pharm Drug Res 68:945–953
Olszewska MA (2012) New validated high-performance liquid chromatographic method for simultaneous analysis of ten flavonoid aglycones in plant extracts using a C18 fused-core column and acetonitrile-tetrahydrofuran gradient. J Sep Sci 35:2174–2183. https://doi.org/10.1002/jssc.201200287
Olszewska MA, Michel P (2009) Antioxidant activity of inflorescences, leaves and fruits of three Sorbus species in relation to their polyphenolic composition. Nat Prod Res 23:1507–1521. https://doi.org/10.1080/14786410802636177
Olszewska MA, Michel P (2012) Activity-guided isolation and identification of free radical-scavenging components from various leaf extracts of Sorbus aria (L.) Crantz. Nat Prod Res 26:243–254. https://doi.org/10.1080/14786419.2010.537271
Olszewska MA, Roj JM (2011) Phenolic constituents of the inflorescences of Sorbus torminalis (L.) Crantz. Phytochem Lett 4:151–157. https://doi.org/10.1016/j.phytol.2011.02.005
Olszewska MA, Nowak S, Michel P et al (2010) Assessment of the content of phenolics and antioxidant action of inflorescences and leaves of selected species from the Genus Sorbus Sensu Stricto. Molecules 15:8769–8783. https://doi.org/10.3390/molecules15128769
Olszewska MA, Presler A, Michel P (2012) Profiling of phenolic compounds and antioxidant activity of dry extracts from the selected Sorbus species. Molecules 17:3093–3113. https://doi.org/10.3390/molecules17033093
Olszewska MA, Kolodziejczyk-Czepas J, Rutkowska M et al (2019) The effect of standardised flower extracts of Sorbus aucuparia L. on proinflammatory enzymes, multiple oxidants, and oxidative/nitrative damage of human plasma components in vitro. Oxid Med Cell Longev 2019:1–18. https://doi.org/10.1155/2019/9746358
Park H, Park H, Chung T-W et al (2017) Effect of Sorbus commixta on the invasion and migration of human hepatocellular carcinoma Hep3B cells. Int J Mol Med 40:483–490. https://doi.org/10.3892/ijmm.2017.3010
Pavlii AI, Makarova GV (1970) Polyphenols of Sorbus quercifolia. Chem Nat Compd 6:773. https://doi.org/10.1007/BF00565361
Pellicer J, Clermont S, Houston L et al (2012) Cytotype diversity in the Sorbus complex (Rosaceae) in Britain: sorting out the puzzle. Ann Bot 110:1185–1193. https://doi.org/10.1093/aob/mcs185
Polat R, Satıl F (2012) An ethnobotanical survey of medicinal plants in Edremit Gulf (Balikesir–Turkey). J Ethnopharmacol 139:626–641. https://doi.org/10.1016/j.jep.2011.12.004
Pranskuniene Z, Ratkeviciute K, Simaitiene Z et al (2019) Ethnobotanical study of cultivated plants in Kaišiadorys District, Lithuania: possible trends for new herbal based medicines. Evid Based Complement Altern Med 2019:1–15. https://doi.org/10.1155/2019/3940397
Raudonė L, Raudonis R, Gaivelytė K et al (2015) Phytochemical and antioxidant profiles of leaves from different Sorbus L. species. Nat Prod Res 29:281–285. https://doi.org/10.1080/14786419.2014.950577
Raudonis R, Raudonė L, Gaivelytė K et al (2014) Phenolic and antioxidant profiles of rowan (Sorbus L.) fruits. Nat Prod Res 28:1231–1240. https://doi.org/10.1080/14786419.2014.895727
Razina TG, Zueva EP, Ulrich AV et al (2016) Antitumor Effects of Sorbus aucuparia L. Extract Highly Saturated with Anthocyans and Their Mechanisms. Bull Exp Biol Med 162:93–97. https://doi.org/10.1007/s10517-016-3554-4
Robertson A, Rich TCG, Allen AM et al (2010) Hybridization and polyploidy as drivers of continuing evolution and speciation in Sorbus. Mol Ecol 19:1675–1690. https://doi.org/10.1111/j.1365-294X.2010.04585.x
Rutkowska M, Olszewska MA, Kolodziejczyk-Czepas J et al (2019a) Sorbus domestica leaf extracts and their activity markers: antioxidant potential and synergy effects in scavenging assays of multiple oxidants. Molecules 24:2289. https://doi.org/10.3390/molecules24122289
Rutkowska M, Owczarek A, Kolodziejczyk-Czepas J et al (2019b) Identification of bioactivity markers of Sorbus domestica leaves in chromatographic, spectroscopic and biological capacity tests: application for the quality control. Phytochem Lett 30:278–287. https://doi.org/10.1016/j.phytol.2019.02.004
Sak K, Jürisoo K, Raal A (2014) Estonian folk traditional experiences on natural anticancer remedies: from past to the future. Pharm Biol 52:855–866. https://doi.org/10.3109/13880209.2013.871641
Šavikin KP, Zdunić GM, Krstić-Milošević DB et al (2017) Sorbus aucuparia and Sorbus aria as a source of antioxidant phenolics, tocopherols, and pigments. Chem Biodivers 14:e1700329. https://doi.org/10.1002/cbdv.201700329
Sennikov AN, Kurtto A (2017) A phylogenetic checklist of Sorbus s.l. (Rosaceae) in Europe. Memo Soc pro Fauna Flora Fenn 93:1–78
Serteser A, Kargioǧlu M, Gök V et al (2008) Determination of antioxidant effects of some plant species wild growing in Turkey. Int J Food Sci Nutr 59:643–651. https://doi.org/10.1080/09637480701602530
Sezik E, Yeşilada E, Honda G et al (2001) Traditional medicine in Turkey X. Folk medicine in Central Anatolia. J Ethnopharmacol 75:95–115. https://doi.org/10.1016/S0378-8741(00)00399-8
Shang N, Saleem A, Musallam L et al (2015) Novel approach to identify potential bioactive plant metabolites: pharmacological and metabolomics analyses of ethanol and hot water extracts of several canadian medicinal plants of the Cree of Eeyou Istchee. PLoS ONE 10:e0135721. https://doi.org/10.1371/journal.pone.0135721
Shavva A, Matyukhina L, Saltykova I (1969) Ursolic acid in Sorbus aucuparia. Chem Nat Compd 5:372. https://doi.org/10.1007/BF00595098
Sohn EJ, Kang DG, Choi DH et al (2005a) Effect of methanol extract of Sorbus cortex in a rat model of L-NAME-induced atherosclerosis. Biol Pharm Bull 28:1239–1243. https://doi.org/10.1248/bpb.28.1239
Sohn EJ, Kang DG, Mun YJ et al (2005b) Anti-atherogenic Effects of the Methanol Extract of Sorbus Cortex in Atherogenic-Diet Rats. Biol Pharm Bull 28:1444–1449. https://doi.org/10.1248/bpb.28.1444
Spoor DCA, Martineau LC, Leduc C et al (2006) Selected plant species from the Cree pharmacopoeia of northern Quebec possess anti-diabetic potential. Can J Physiol Pharmacol 84:847–858. https://doi.org/10.1139/y06-018
Sun J, Shi S, Li J et al (2018) Phylogeny of maleae (Rosaceae) based on multiple chloroplast regions: implications to genera circumscription. Biomed Res Int 2018:6–9. https://doi.org/10.1155/2018/7627191
Takaishi K, Kuwajima H, Hatano H, Go H (1977) Cyanogenic glycosides and glycosidase of Sorbus species. Chem Pharm Bull 25:3075–3077. https://doi.org/10.1248/cpb.25.3075
Tam TW, Liu R, Arnason JT et al (2009) Actions of ethnobotanically selected Cree anti-diabetic plants on human cytochrome P450 isoforms and flavin-containing monooxygenase 3. J Ethnopharmacol 126:119–126. https://doi.org/10.1016/j.jep.2009.07.036
Tardío J, Pardo-de-Santayana M, Morales R (2006) Ethnobotanical review of wild edible plants in Spain. Bot J Linn Soc 152:27–71. https://doi.org/10.1111/j.1095-8339.2006.00549.x
Termentzi A, Kefalas P, Kokkalou E (2006) Antioxidant activities of various extracts and fractions of Sorbus domestica fruits at different maturity stages. Food Chem 98:599–608. https://doi.org/10.1016/j.foodchem.2005.06.025
Termentzi A, Alexiou P, Demopoulos VJ, Kokkalou E (2008a) The aldose reductase inhibitory capacity of Sorbus domestica fruit extracts depends on their phenolic content and may be useful for the control of diabetic complications. Pharmazie 63:693–696. https://doi.org/10.1691/ph.2008.8567
Termentzi A, Kefalas P, Kokkalou E (2008b) LC-DAD-MS (ESI+) analysis of the phenolic content of Sorbus domestica fruits in relation to their maturity stage. Food Chem 106:1234–1245. https://doi.org/10.1016/j.foodchem.2007.07.021
Termentzi A, Zervou M, Kokkalou E (2009) Isolation and structure elucidation of novel phenolic constituents from Sorbus domestica fruits. Food Chem 116:371–381. https://doi.org/10.1016/j.foodchem.2009.02.019
Tian Y, Liimatainen J, Alanne AL et al (2017) Phenolic compounds extracted by acidic aqueous ethanol from berries and leaves of different berry plants. Food Chem 220:266–281. https://doi.org/10.1016/j.foodchem.2016.09.145
Turumtay H, Midilli A, Turumtay EA, et al (2017) Gram (−) microorganisms DNA polymerase inhibition, antibacterial and chemical properties of fruit and leaf extracts of Sorbus acuparia and Sorbus caucasica var. yaltirikii. Biomed Chromatogr 31:e3901. https://doi.org/10.1002/bmc.3901
Uddin G, Latif A, Arfan M et al (2013) Phytochemicals from the stem wood of Sorbus lanata (D. Don.) Schauer. Phytochem Lett 6:84–89. https://doi.org/10.1016/j.phytol.2012.10.002
Veberic R, Slatnar A, Bizjak J et al (2015) Anthocyanin composition of different wild and cultivated berry species. LWT Food Sci Technol 60:509–517. https://doi.org/10.1016/j.lwt.2014.08.033
Vianna R, Brault A, Martineau LC et al (2011) In vivo anti-diabetic activity of the ethanolic crude extract of Sorbus decora C.K.Schneid. (Rosacea): a medicinal plant used by Canadian james bay cree nations to treat symptoms related to diabetes. Evid Based Complement Altern Med 2011:1–7. https://doi.org/10.1093/ecam/nep158
Vogl S, Picker P, Mihaly-Bison J et al (2013) Ethnopharmacological in vitro studies on Austria’s folk medicine—an unexplored lore in vitro anti-inflammatory activities of 71 Austrian traditional herbal drugs. J Ethnopharmacol 149:750–771. https://doi.org/10.1016/j.jep.2013.06.007
Xuan SH, Park YM, Park SH et al (2018) Suppression of ultraviolet B-mediated matrix metalloproteinase generation by Sorbus commixta Twig extract in human dermal fibroblasts. Photochem Photobiol 94:370–377. https://doi.org/10.1111/php.12868
Yang G, An H-J (2014) β-sitosteryl-3-O-β-glucopyranoside isolated from the bark of Sorbus commixta ameliorates pro-inflammatory mediators in RAW 264.7 macrophages. Immunopharmacol Immunotoxicol 36:70–77. https://doi.org/10.3109/08923973.2013.866956
Yin MH, Kang DG, Choi DH et al (2005) Screening of vasorelaxant activity of some medicinal plants used in Oriental medicines. J Ethnopharmacol 99:113–117. https://doi.org/10.1016/j.jep.2005.02.013
Yin Y, Zhang Y, Li H et al (2019) Triterpenoids from fruits of Sorbus pohuashanensis inhibit acetaminophen-induced acute liver injury in mice. Biomed Pharmacother 109:493–502. https://doi.org/10.1016/j.biopha.2018.10.160
Yoshizawa Y, Kawaii S, Urashima M et al (2000) Differentiation-Inducing Effects of Small Fruit Juices on HL-60 Leukemic Cells. J Agric Food Chem 48:3177–3182. https://doi.org/10.1021/jf9908650
Yu T, Lee YJ, Cho JY (2009) Inhibitory effect of Sorbus commixta extract on lipopolysaccharide-induced pro-inflammatory events in macrophages. J Med Plants Res 3:600–607
Yu T, Lee YJ, Jang H-J et al (2011) Anti-inflammatory activity of Sorbus commixta water extract and its molecular inhibitory mechanism. J Ethnopharmacol 134:493–500. https://doi.org/10.1016/j.jep.2010.12.032
Yu C, Tang H, Zheng Y, Li L (2013) Flavonoids from the leaves of Sorbus tianschanica. Chem Nat Compd 49:115–116. https://doi.org/10.1007/s10600-013-0524-1
Yu X, Wang Z, Shu Z et al (2017) Effect and mechanism of Sorbus pohuashanensis (Hante) Hedl. flavonoids protect against arsenic trioxide-induced cardiotoxicity. Biomed Pharmacother 88:1–10. https://doi.org/10.1016/j.biopha.2016.12.130
Zapesochnaya GG, Ban’kovskii AI, Gubanov IA (1969) Hyperin from Sorbus tianschanica. Chem Nat Compd 5:104. https://doi.org/10.1007/BF00633293
Zapesochnaya GG, Aitbaeva RK, Ban’kovskii A (1973) Flavonoids and chlorogenic acid from Sorbus tianschanica. II. Chem Nat Compd 9:112. https://doi.org/10.1007/BF00580914
Zymone K, Raudone L, Raudonis R et al (2018) Phytochemical profiling of fruit powders of twenty Sorbus L. cultivars. Molecules 23:2593. https://doi.org/10.3390/molecules23102593
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sołtys, A., Galanty, A. & Podolak, I. Ethnopharmacologically important but underestimated genus Sorbus: a comprehensive review. Phytochem Rev 19, 491–526 (2020). https://doi.org/10.1007/s11101-020-09674-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11101-020-09674-9