
Abstract
Between the nineteenth and early twentieth centuries, large-scale afforestation projects were undertaken in the Swiss Prealps. The Swiss stone pine (Pinus cembra L.), an emblematic alpine tree with important economic potential, conservation value and ecological relevance, was one of the species used in these afforestation efforts. We investigated the distribution and site characteristics of all known natural populations and planted stands in the canton of Fribourg (Switzerland). Between 1885 and 1952, artificial afforestation was carried out in the mountainous regions of the canton of Fribourg. Nearly 450,000 seedlings of P. cembra were planted. One century later, only approximately 650 trees survived (0.15% of all planted trees). Moreover, no natural regeneration can be observed in the afforested stands. Since no known harvest activities were undertaken in these planted stands, the whole afforestation campaign of P. cembra in the canton of Fribourg can be described as a failure. Possible causes are investigated and discussed. In this region, P. cembra occurs naturally only in particular microrefugia with markedly rough topography. We argue that the lack of knowledge or consideration of the ecology of the species at a local scale can be an important factor leading to this failure. In the actual context of large-scale afforestation projects anticipating and mitigating the negative effects of global warming, our study highlights the importance of a precise understanding of species ecology and regional conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Since the establishment of the first agricultural societies in Europe, the landscape has been significantly modified, particularly by the clearance of forests for cropland and pasture and by the overexploitation of forests as a source of energy and construction materials (Lillie 2000). Consequently, the forest cover decreased almost continuously, reaching a minimum around the time of the industrial revolution. In Switzerland, forest cover in so-called usable land (land that can be used for pasture) was estimated at 41.8% in 1000 B.C. but reached only 1.3% in 1850 (Kaplan et al. 2009). The preindustrial period was often referred to as the age of wood, as wood was the main source of energy for industries and households, leading to an important overexploitation of the forests (Neet and Bolliger 2004; Hürlimann 2008). In Switzerland, wood was transported in large quantities by rivers, notably to smelting plants and shipyards in neighboring countries. In the mountainous regions of the Western Swiss Prealps, especially in the canton of Fribourg, a large quantity of wood was also necessary for the flourishing cheese industry. The fear of wood penury was widespread in the eighteenth and nineteenth centuries and had political repercussions by legitimizing forest laws. Many Swiss cantons began to enact forest laws in the first half of the nineteenth century, allowing the transition to a regulated forest economy (Schuler 2008). The aim was to guarantee long-term forest exploitation and to ensure the conservation and extension of protective forests.
In this context, major reforestation and afforestation projects emerged in the canton of Fribourg in the nineteenth and early twentieth centuries, mainly in the Prealps (Müller 1990; Stalder et al. 1990). Afforestation and reforestation are two closely related silvicultural practices. Reforestation can be defined as the “re-creation of forest on a previously forested area” and afforestation as the “creation of forest on areas not naturally forested in recent times” (Di Sacco et al., 2020). The objective was not only to re-establish forests for their economic value but also to avoid problems that arose with deforestation, e.g., to diminish the negative effects and frequencies of avalanches, landslides and floods (Fankhauser 1898; Rikli 1909). These afforestation projects were ambitious and considerably laborious, as all the work was handmade, and the access to and transport of the material to remote locations at high elevations were difficult. Many local people were often engaged, and job creation was an additional important argument to finance such complex projects (Müller 1990; Stalder et al. 1990).
Several conifer species were particularly popular in afforestation projects, including Swiss stone pine (Pinus cembra L.). The species is distributed in timberline ecotones in the Alps and in the Carpathian Mountains (Ulber et al. 2004; Boden et al. 2010) and is often considered a glacial relict of high conservation value (Zięba et al. 2020). Moreover, it is the most well adapted European tree species to harsh upper subalpine climates (Ulber et al. 2004). Pinus cembra rarely exceeds a height of 25 m and is characterized by a very slow growth rate. It has wingless seeds and is thus dependent on birds for seed dispersal, which is a result of coevolution with corvid birds; in Europe this occurs mainly with the Eurasian nutcracker (Nucifraga caryocatactes) (Tomback and Linhart 1990; Caudullo and De Rigo 2016). One of the most important consequences of this nearly obligate bird–pine mutualism is the clustering and a population structure that is observed as a consequence of the seed-burying behavior of the nutcracker (Rikli 1909; Tomback et al. 1993).
Furthermore, P. cembra belongs to the five needle pines (section Quinquefoliae) and is closely related to the Siberian and East Asiatic P. sibirica and P. pumila (Hao et al. 2015; Gernandt et al. 2018). Currently, there is a very large disjunction between P. cembra and its nearest relative, P. sibirica. The fragmentation of the formerly united range of these closely related species may have occurred only recently in the Holocene (Gugerli et al. 2009). Pinus cembra was thus widespread in the Alps during the early and mid-Holocene (Lotter and Birks 2003), but its distribution was largely reduced when the climate became warmer and as a result of competition with Norway spruce (Picea abies) (Höhn et al. 2009). In Switzerland, the core distribution is situated in the central Alps, where the climate is continental, giving a competitive advantage to P. cembra over Picea abies, which otherwise dominates the subalpine vegetation zone of the Alps and the Prealps (Gugerli et al. 2009; Info Flora 2021). In the canton of Fribourg, P. cembra occurs at the margin of its distribution and is restricted to particular microrefugia (Rikli 1909). The species is consequently relatively rare and has been legally protected in the canton of Fribourg since 1973 (Conseil d’Etat du canton de Fribourg 1973); thus, no harvest is currently allowed.
There are several plausible reasons why P. cembra was selected for afforestation in Switzerland and, more specifically, in the Western Prealps. First, it has always been appreciated as a very attractive tree. Second, it was seen to be a perfect tree to prevent avalanches and to stabilize the terrain (Ulber et al. 2004) since it grows naturally at high elevations, above other trees, frequently in areas with steep topography and shallow soils. Third, trees have many uses and therefore have important economic potential (Caudullo and De Rigo 2016). Its wood possesses many prized properties, since it is soft, resistant, and visually aesthetic and has a highly appreciated resin smell. The bark and needles of P. cembra contain useful antioxidant and antimicrobial compounds (Apetrei et al. 2011), and its large seeds are edible and have a high lipid content (Schütt 2014). In his very popular book Der Lehrer im Walde, published at the beginning of the nineteenth century, Karl Kasthofer, forest inspector and silviculture professor in Switzerland, complained that P. cembra was never planted (Kasthofer 1828). He declared that if a forest of P. cembra had been planted, “from the yield of this forest alone, all the poor could be paid for, all roads and bridges could be maintained, and good, skilled schoolmasters could be paid for the youth in our villages” (Kasthofer 1828). This example illustrates that the reputation of P. cembra was very good, and all these arguments may explain why the choice of P. cembra for large afforestation was very logical and an extremely attractive idea at the end of the nineteenth century.
Today, afforestation is again a very popular topic, both locally and at the global scale. Numerous countries and international organizations have chosen afforestation or reforestation as one of the main goals of the twenty-first century, not only to expand or restore forests but also as a key instrument in mitigating climate change (Van Kooten et al. 2002; Naudts et al. 2016; Zheng et al. 2016; Di Sacco et al. 2020).
The aim of our study was to investigate the past and present status and characteristics of P. cembra forests in the Western Prealps of Switzerland. More specifically, we aimed to address the following questions: (1) What are the distribution and population sizes of natural populations and planted stands of P. cembra in the canton of Fribourg? (2) How intensive were afforestation efforts with P. cembra in the past two centuries? (3) What is the success of these artificial plantations at the beginning of the twenty-first century? (4) What are the differences between the population structures and between the topographic characteristics of the sites where P. cembra is growing naturally and where it was artificially planted? The synthesis of our results should deliver valuable conclusions for future afforestation projects.
Material and methods
Actual distribution
To map the occurrences of P. cembra in the canton of Fribourg (both natural populations and planted stands), we first collected information from different sources: (1) notes or personal communications of local botanists and foresters, (2) data from the Forest and Nature Service of the State of Fribourg, (3) data from the national center on the Swiss flora (Info Flora 2021), (4) information found in the available literature (Cottet and Castella 1891; Rikli 1909), (5) historical data of afforestation in the Archives of the State of Fribourg (see below) and (6) our own field observations. Specifically, all areas where historical data indicated artificial afforestation projects were surveyed for the presence of P. cembra. Furthermore, we visited and surveyed all the natural populations in either known locations or favorable potential areas. Occurrences with P. cembra were defined as planted artificial stands when we detected that historical afforestation took place in the same area (most cases) or sometimes because it was evident that the population was not natural (based on the age of the trees, the environment, the absence of seedlings and rejuvenation, among other factors).
Historical data
The majority of historical data were found in the Archives of the State of Fribourg (Etat de Fribourg 2021a). Information about afforestation projects and the number of trees planted were summarized in the annual reports of the cantonal authorities (“Compte rendu de l’administration du Conseil d’Etat”). The list of all references is given in the Online Resource 1. We rarely found information about P. cembra in other documents (e.g., in the archives of the cantonal Forest and Nature Service (Etat de Fribourg 2021b), in municipality archives or by local foresters). To obtain information about the possible ancient harvest of P. cembra in the study area, we carried out surveys by long-term experienced foresters and local long-established sawmills.
Field surveys
The field surveys and investigations were carried out from late spring until the beginning of autumn 2020. For each small stand or small population, the number of trees (height > 1 m) was directly counted in the field. In a few cases, due to the steep, dangerous topography, the individual number was estimated at a certain distance with binoculars. For large populations, we used the point-centered-quarter method (Cottam and Curtis 1956; Mitchell 2010); in the large natural population of Forêt du Lapé, we selected 41 random points (= 164 trees, one per quarter). The distance between the random points and the four closest trees of P. cembra (height > 1 m) was measured with a laser telemeter (LEICA Disto D510). The diameter at breast height (DBH) of these random trees was also recorded. We used our field observations and data to map large natural populations using high-resolution (25 cm) orthophotos (aerial photography), provided by the Swiss Federal Office of Topography (Swisstopo 2021), and canopy height models derived from LiDAR (light detection and ranging; data acquisition 2016), provided by the Cantonal Forest and Nature Service (Etat de Fribourg 2021b). The number of P. cembra trees in large natural populations was extrapolated (mapped area multiplied by the tree density calculated with the point-centered-quarter method). Other data were also collected during the field work, e.g., the presence of seedlings and young trees, the DBH in planted stands (20 randomly selected trees in 6 stands = 120 trees), and general information about the site characteristics and surrounding vegetation.
Data analyses
Geographic information system (GIS) software (QGIS Development Team 2021) was used to georeference and map the natural populations and planted stands. Microtopography was analyzed using a high-resolution (25 cm) LiDAR-derived digital elevation model (DEM) provided by the Cantonal Forest and Nature Service (Brubaker et al. 2013; Etat de Fribourg 2021b). All populations and planted stands were divided and intersected by a grid with cells of 1 hectare (100 m × 100 m). Slope, aspect, elevation and terrain ruggedness index (TRI) were calculated from the DEM for each cell (mean, median or range per cell). The TRI is an index that quantifies topographic heterogeneity (Riley et al. 1999; Graser 2013). It corresponds to the mean difference between a central pixel and its surrounding cells. We used the nonparametric Wilcoxon signed-rank test (Wilcoxon et al. 1970) to compare some variables between the planted stands and natural populations. Since the data are not truly independent (due to spatial autocorrelation), we used populations or stands as units rather than single 1-hectare cells. The calculations were performed and the graphs were constructed in R (R Core Team 2018). A LOESS (locally estimated scatterplot smoothing) curve (Jacoby 2000) was fitted on histograms to give a smooth approximation of the results.
Results
Distribution of natural populations and planted stands
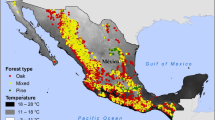
The distribution of P. cembra in the canton of Fribourg was divided into 12 populations (Fig. 1 and Table 1). Several natural populations are very small and isolated (for example Dent de Broc with 10 trees). Other populations cover large areas, especially along cliffs and in the vicinity of summits in the Gastlosen chain and on old and stabilized screes with large blocks (ancient cliff falls). Our division into 12 natural populations is somewhat arbitrary and used for practical reasons since the most adjacent populations are likely connected. However, we decided to use the term population as “a group of individuals of the same species living together in a particular place” (Berryman 2002). We calculated the density of P. cembra in natural populations at 53.24 trees/ha, with a 95% confidence interval of [45.60; 61.99]. Thus, we estimated a total number of 7954 trees for natural populations in the study area, with a 95% confidence interval of [6849; 9218]. The total area mapped for natural populations is approximately 147.9 ha.

Situation and overall distribution of Pinus cembra in the canton of Fribourg (Switzerland), indicating natural populations (orange) and planted stands (red). The numbers refer to the estimated numbers of trees. (Color figure online)
We found a total of 11 occurrences where the origin of P. cembra trees was likely artificial (Fig. 1 and Table 1). The largest planted and still existing stand (Dent du Bourgo) consisted of approximately 300 trees. However, the majority of planted stands are much smaller, and three of them are composed of only 1–7 individuals (Stockberg, La Goille au Cerf, La Berra). For the entire study area, we found a total of 652 trees in planted stands on a total surface of approximately 17 ha. Although it is not impossible that isolated trees in afforested areas may have been missed, we are confident that we have gathered detailed knowledge of the distribution of P. cembra in the canton of Fribourg.
Historical afforestation
Historical data of past afforestation efforts were found for the years between 1885 and 1952, with the majority of data being from before 1914. We recorded a total of 23 areas where plantations were undertaken (Fig. 2 and Online Resource 1). In general, the projects ran over several consecutive years. Pinus cembra was usually planted in the study area together with other species, mainly Picea abies, Larix decidua and Abies alba. Exotic species were also used, in particular Pinus strobus and Pseudotsuga menziesii. More rarely, Pinus mugo or broadleaf species, such as Fagus sylvatica, were also planted.

Numbers of planted Pinus cembra trees in all afforestation projects for the period 1885–1952 (white bars and left numbers) and numbers of remaining trees found today in the same area (red bars and right numbers). The black dots indicate the approximate location where the projects took place. (Color figure online)
During this period, we found that at least 448,025 seedlings of P. cembra were planted in the canton of Fribourg. This number should be considered as a minimum (see Discussion). Some of the projects were relatively modest, with less than 1000 seedlings planted during a single year. In contrast, several afforestation projects lasted for several years, and thousands of seedlings were planted. This was the case, for example, in Höllbach-Gérine, with 136,750 seedlings planted between 1896 and 1952, and in Plan de Tissiniva, with 74,300 seedlings planted between 1905 and 1910 (Fig. 2 and Online Resource 1). For the entire canton, only 0.15% (652 individuals) of the afforested P. cembra trees are alive today.
The last indication that P. cembra was still sometimes planted was found for 1952, but the species was no longer commonly used during this period. We found reliable information for several large afforestation projects that took place between 1930 and 1950, where P. cembra was no longer used.
No information was obtained from local foresters, local sawmills, museums or archives showing that P. cembra has been harvested in the past in the canton of Fribourg. It is therefore very likely that the planted stands were never harvested, or if so, only marginally.
Differences between natural populations and existing planted stands
Our study detected notable differences between remaining planted stands and natural populations. Natural populations occur mainly between 1500 and 2000 m a.s.l. (Fig. 3a). Trees are nearly exclusively restricted to northwest-exposed limestone cliffs, cliff edges and crests and at the edges of large blocks (fallen rocks) (Fig. 3c). The microtopography is particularly steep, rugged and irregular (Fig. 3e). In natural populations, the vegetation is sparse and low in height, and the soil is very shallow. The only tree species that usually occur together with P. cembra are P. mugo and small individuals of Picea abies.

Topographic analysis of the natural populations and planted stands of Pinus cembra, with relative frequency histograms and boxplots showing: a mean elevation per hectare for natural populations, b mean elevation per hectare for planted stands, c median slope aspect per hectare for natural populations, d median slope aspect per hectare for planted stands, e topographic ruggedness index (TRI), range per hectare for natural populations, f topographic ruggedness index (TRI), range per hectare for planted stands. The curves above the histograms give a smooth approximation (LOESS curve)
Planted stands, where the remaining trees were present, were located mainly between 1500 and 1800 m a.s.l. (Fig. 3b). Although this is on average slightly lower in elevation than natural populations, there is a large overlap, and the difference is only marginally significant (Wilcoxon signed-rank test p-value: 0.069). Similar to natural populations, planted stands are also mainly exposed to the northwest (Fig. 3d). The main difference between natural populations and planted stands concerned the microtopography (Fig. 3f). Where large afforestation projects took place, trees were often planted on steep slopes but with a rather homogenous microtopography. Accordingly, the TRI was significantly lower in planted stands than in natural populations (Wilcoxon signed-rank test p-value: < 0.001). This difference in microtopography seemed evident in the field (Fig. 4). The vegetation developing in the planted stands is rather dense and competitive, of the type Adenostylion, Alnenion viridis, Abieti-Piceion or Vaccinio-Piceion. Only a few planted stands (for example, Teysachaux) were located in areas with similar characteristics as natural populations, in the proximity of cliffs and with the presence of large blocks.

Example of a topographic comparison (3-dimensional views) between two sites with Pinus cembra: a a site with a natural population of Forêt du Lapé, the species occurs in the ancient pile of fallen rocks (foreground, bottom orange arrow) and on cliffs (background, top orange arrow); and b a site with a planted stand at Plan de Tissiniva, where trees were planted on the steep slope (foreground, red arrow). The color gradient indicates the terrain ruggedness index (TRI), from low (dark blue) to high ruggedness (yellow). (Color figure online)
Moreover, in planted stands, no seedlings or young trees of P. cembra were observed during our surveys. Although several trees with small DBHs were observed, they were previously broken and only recently produced new shoots. The trees in planted stands are able to reproduce because cones with seeds inside were regularly observed. The tree size in planted stands was rather homogenous, with a DBH mainly between 20 and 40 cm (Fig. 5b). In natural populations, young trees and seedlings were always observed, mainly in rock cracks, but were not extremely abundant. Furthermore, trees of different ages were mixed (Fig. 5a), and the DBH of approximately 20% of all trees was ≤ 5 cm (seedlings and small trees > 1 m height not included). The DBH of the largest trees was rarely > 80 cm.

Distribution of diameter at breast height (DBH) of the natural population and the planted stand of Pinus cembra: a relative frequency histogram for a natural population in Forêt du Lapé, 164 random trees, b relative frequency histogram for planted stands, 120 random trees (20 trees in 6 different stands). The curves above histograms give a smooth approximation (LOESS curve)
Discussion
Pinus cembra is one of the most characteristic woody species of the alpine treeline (Körner 2012; Tranquillini 2012). It is considered an emblematic species of the European tree flora and has attracted the attention of local people, foresters and researchers for centuries (Kasthofer 1828; Caudullo and De Rigo 2016). As a consequence, there is a large amount of research on various aspects of the biology, ecology, distribution, biogeography and genetics of P. cembra on a large geographical scale (Gugerli et al. 2001; Höhn et al. 2009; Casalegno et al. 2010; Neuschulz et al. 2018; Dauphin et al. 2020, 2021; Zięba et al. 2020). Far fewer studies have been performed at the local level, especially on the peripheral populations of this species (Lendvay et al. 2014; Tóth et al. 2019). The information on P. cembra from the Western Prealps, for example, is either very old (Rikli 1909) or based on only a few individuals collected exclusively from the largest and best known populations (Gugerli et al. 2009). Our work is the first detailed synthesis of the distribution, population sizes, site topography and sylvicultural history of P. cembra, covering the entire territory of the canton of Fribourg in Switzerland.
Natural distribution
Our results confirm the peripheral situation of P. cembra in the canton of Fribourg, attaining its northwestern distribution and climatic limits in this region (Dauphin et al. 2020). Other than in the Central Alps, the species rarely forms monospecific populations in this region and grows exclusively in very specific and spatially restricted microhabitats (Doutaz et al. 2006). According to our detailed estimations, there were fewer than 8000 individuals of P. cembra in the study area. The largest concentration of trees and populations occurs along the chain of Gastlosen, on the border with the cantons of Vaud and Bern, with approximately 6850 individuals (86%). Furthermore, much smaller populations occur in the Vallon des Morteys, Hochmatt and Ziebegg. The smallest and most isolated natural population occurs on a very steep part of the Dent de Broc (ca. 10 individuals). Some of these populations or their fragments are growing in very steep and inaccessible cliffs and are certainly primary forest patches. There is no doubt that those populations could not be planted, and their structure, with a mix of ages, also confirms this assumption (Tranquillini 2012; Nussbaumer 2016). Although many trees in these natural forests may be very old (P. cembra can live up to 1000 years, Caudullo and De Rigo 2016), the trees in the canton of Fribourg are rather small (mainly 8–12 m high), and only an extremely small proportion attain a diameter > 80 cm (Fig. 5).
According to Gugerli et al. (2009), P. cembra migrated into the Western Prealps during postglacial colonization, similar to all other alpine regions, from peripheral refugia. Although the species reached our study area many thousands years ago (Burga et al. 1998), it was not able to colonize other neighboring high mountain chains of the canton of Fribourg, which are situated further north or west, such as the Moléson or Kaiseregg region (Rikli 1909; Info Flora 2021). The mutualistic seed disperser of P. cembra, the Eurasian nutcracker, is very common in the canton of Fribourg. The distances between the main concentration of P. cembra along the Gastlosen to the other uncolonized high-altitude mountain chains are very short. Thus, the distance was certainly not an obstacle for the nutcracker, which is able to cover distances up to 15 km (Caudullo and De Rigo 2016).
Historical afforestation
We decided to use the term afforestation for P. cembra in our study region because the species was probably not present before in the planted area, and we do not know when the forest became absent from these areas before plantations. With only 0.15% of the remaining P. cembra trees planted (out of ca. 450,000), and with no known wood harvest, one century after the afforestation, we can conclude that this impressive campaign in the canton of Fribourg was a failure. Moreover, P. cembra was unable to regenerate where it was artificially introduced since no seedlings and no juvenile trees were found in the remaining planted sites, and thus it will probably completely disappear from these locations in the near future. In comparison, in natural populations, also at very high altitudes, the natural regeneration and recruitment is very common (Tranquillini 2012). In the natural P. cembra forests in our study area, the proportion of juvenile trees is often higher than 30% (Doutaz et al. 2006), attaining ca. 10 juvenile trees per hectare (Nussbaumer 2016).
The number of planted trees (448,025) must be considered as a minimum estimate. First, we did not find all of the information for every afforestation project in the archives. We discovered several planted stands without any historical records and, thus, without numbers of seedlings used in these sites, for example, for Moléson (MOL) and Combe d’Allière (ALL; Figs. 1 and 2). Second, some information was contradictory between various sources, mainly for the large afforestation projects in Höllbach-Gérine (Online Resource 1). In such cases, we always selected the most prudent numbers. Independent of the exact numbers, the magnitude of this past afforestation campaign is enormous in terms of both time and costs.
Without having studied it in detail, we also observed the poor success of afforestation of some other species planted together with P. cembra (in particular P. mugo, P. strobus and Larix decidua).
Possible causes of afforestation failure
There are several possible reasons why this huge afforestation effort in the canton of Fribourg turned out to fail in the mid-term and we discuss some of them bellow. Unfortunately, historical data for such causes does not exist, thus we can only formulate hypotheses according to the present state and available knowledge. We miss information about the historical afforested area, where no tree is remaining today. As these locations are not precisely known, we could only consider in our analysis places where planted trees are remaining. We also miss information about early twentieth century silvicultural practices and if management activities where undertaken following the plantations.
Unsuitable climatic conditions and topography. As explained above, P. cembra reaches the margin of its natural distribution in the canton of Fribourg. The climate of this region is more oceanic than in the Central Alps, i.e., in the core distribution area of the species. Under less continental conditions, P. cembra is generally outcompeted by other species, mainly Picea abies (Ettlinger 1975; Höhn et al. 2009). Thus, P. cembra is able to occur naturally only in microrefugia with harsh continental conditions: on cliffs and cliff edges, at the edges of large blocks and sometimes on crests. These conditions are present in the canton of Fribourg mainly along the chain of Gastlosen and especially in Forêt du Lapé, the largest population, with a large number of huge blocks. The unique topography of this area results in site characteristics that reduce competition (mainly of spruce) and help the establishment of P. cembra: (1) temporarily strong aridity; (2) low accumulation of snow and early melting, which exposes the soil, young trees and vegetation layer to extreme cold in winter conditions (Schönenberger 2001; Doutaz et al. 2006). For example, Schönenberger (1975) showed that planted Picea abies in locations with early snow melting were more frequently damaged by frost than P. cembra. The latter is one of the most cold-hardy trees known and is well adapted to frost and supporting winter temperatures below -43 °C without any damage (Caudullo and De Rigo 2016); (3) high wind exposure and air dryness; and (4) shallow soils with low fertility. Therefore, according to our results, the rugged topography and microtopography seem to be key factors for P. cembra survival in this region. Thus, this could also be one of the main problems in artificially planted stands outside of the natural distribution area of P. cembra. The afforested sites where the species still persist possess microtopography that is too homogeneous (see Fig. 4), favorable to other more competitive species.
We did not find any important difference between the remaining planted stands and natural populations (see Fig. 5) in regard to the elevational range or slope aspect. Therefore, it seems that neither altitude nor exposure of sites selected for afforestation was a relevant cause of the failure. However, it cannot be completely ruled out that some planted stands where P. cembra completely disappeared (see Fig. 2) were located at altitudes too low and/or on unsuitable south-exposed slopes. Unfortunately, the lack of information on the exact location of the historically afforested areas do not allow us to carry out any comparative analysis.
Competition. The vegetation that exists around planted seedlings can either be considered as competition and cause mortality, or as facilitation and protect seedlings from stressful environmental conditions, thereby improve their establishment. The effect of herbaceous alpine vegetation on P. cembra seedling emergence and first-year performance were studied in a field experiment in the French Alps (2100 m a.s.l.). Total emergence and locally-germinated seedling survival were not affected, but for seedlings planted at 2 months of age, negative vegetation impacts dominated for all response parameters: first-year survival, growth and carbohydrate accumulation (Loranger et al. 2017). The possible absence of management of vegetation following plantations could be an important factor of failure. Pinus cembra is a light-demanding species that can tolerate only moderate shadows (Fourchy 1968). Today, more than a hundred years after plantations, the access to light in planted stands is low, with well-established and competitive vegetation, which probably hinder the natural regeneration of P. cembra. In contrast in natural populations, a rugged topography and microtopography allow more light to enter the populations, especially at the edges of cliffs and large blocks. The vegetation is also less competitive, lower and sparse in these places.
Pathogens. It has long been demonstrated that pathogens can cause high mortality among planted seedlings of P. cembra (Rikli 1909; Fourchy 1968), mainly due to snow blight (Phacidium infestans) and brunchorstia disease (Gremmeniella abietina). In an afforestation project that took place in Davos (Switzerland) in the 1950s and 1960s, approximately 47.6% of all P. cembra trees were infected by P. infestans approximately 20 years later (Schönenberger 1975). In the same region, for another plantation performed in 1975, only 5% of P. cembra trees survived 30 years later due to pathogen attack (Barbeito et al. 2013). How newly planted stands, sometimes isolated, can be infected by pathogens is difficult to know, but long-distance dispersal seems to be possible, for example for Gremmeniella abietina. It occurs through wind-borne ascospores (but reported as absent in the European strain, EPPO 2021), or conidia rain-splash dispersal could also play a role for long-distance dispersal with the help of wind (Petäistö and Heinonen 2003). Gremmeniella abietina is more successful in trees that are stressed by adverse environmental conditions (Senn 1999).
It was demonstrated that in planted stands, persistent snow cover and low wind speed were two factors favoring the development of pathogens, and P. cembra survived mostly at sites where the snow melted early (Schönenberger 1975; Senn and Schönenberger 2001). This corroborates our results since natural populations occur where snow does not persist for a long time due to topography, thus reducing the impact of pathogens on seedlings and young trees. Accordingly, the microrefugia of P. cembra in this region may be considered as not only climatic and topographic refugia but also as “refugia from pathogens”.
Wildlife damage. Browsing can generate important damage in forest plantations and P. cembra can be impacted (Ulber et al. 2004; Barbeito et al. 2013; Oberhuber et al. 2019). Here, again, the topography and microtopography of natural populations may largely protect juvenile individuals of P. cembra from browsing by ungulate herbivores. This impact of browsing could be larger in planted stands, although we have no data to support this assumption. Moreover, practically all large herbivores were either regionally extinct or present at very low density in the nineteenth century due to intensive hunting (Breitenmoser 1998). Thus, wildlife damage is certainly very marginal as an explanation of afforestation failure.
Planting practices. It is well documented, that the afforestation at high altitudes, and especially at timberline, is an extremely difficult endeavor (Schönenberger 2001; Senn and Schönenberger 2001). In the treeline ecotone, the environmental conditions are very harsh (Körner 2012). Kronfuss and Havranek (1999) for example, studying artificial afforestation of P. cembra in Tyrol (Austria), demonstrated that the height growth of young artificially planted trees decreased with altitude by about 5% per 100 m. The ecological conditions encountered near the tree line should be given the greatest consideration for successful afforestation (Mullenbach 1982, 2000). However, the context of the afforestation activities in the canton of Fribourg is different. Plantations were carried out in our study area mainly between 1500 and 1900 m a.s.l. (probably even lower). It means that the main plantations were not close to the timberline but in much lower altitudes, and thus in better conditions for afforestation.
However, even at these lower altitudes, laborious methods of cultivation and planting are generally advised, e.g. to use potted rather than bare-rooted plants. Cluster arrangement must be preferred to a regular planting pattern, since it allows consideration of micro-site conditions, the establishment of a good surface structure, and the preclusion of uniform stands. The growth conditions in mountainous regions impose appropriate methods of tending for young growth and thickets (Schönenberger et al. 1990, 1995). It is advised that for plantation near the tree limit, the planting material should lie within a 100 m altitude range relative to the planting site. It is also suggested to breed the plants in montane altitudes and to transplant them into a nursery located above 1500 m to ensure acclimatization (Ulber et al. 2004).
The planting practices used for P. cembra afforestation in the canton of Fribourg are largely unknown. Considering the period of plantations and some historical pictures, seedlings used were almost certainly bare-root. Pinus cembra often has difficulties with rooting in this case (Felix Gugerli, personal communication). In high-altitude forestry, potted plants are preferred to bare rooted plants in order to reduce the stress to which trees are exposed when transferred from the nursery to a high altitude afforestation site (Senn 1999). Moreover, P. cembra trees are usually strongly mycorrhized. Three basidiomycete species of the genus Suillus form a strictly host-specific symbiosis with it, as well as a number of other generalist ectomycorrhizae (Rainer et al. 2015). It has been demonstrated that ectomycorrhizal fungi are favorable or even necessary for its establishment (Mullenbach 2000). Failure of afforestation in the Austrian Alps have been reported and linked to the absence of ectomycorrhizal fungi (Moser 1967). Inoculation of the mycelium in the nursery and cultivation of the plants in large buckets with inoculated substrate seems to be a factor of success for planting trees on unforested areas (Mullenbach 2000).
Inappropriate methods for plantings could be one of the main problem for historical afforestation. However, plantations occurred during many years. If high mortality occurred right after plantation, it would have probably been detected and avoided. Forestry was already a well-developed discipline at this time. Secondly, plantation occurred in the canton of Fribourg mainly between 1500 and 1900 m a.s.l., probably even lower, but no trees are remaining. It means that the main plantations are not close to the tree line, where the condition are less difficult for afforestation.
Seed production and dispersal. According to our surveys, trees in planted stands produce cones with seeds. Seed cones appear in P. cembra after 40–60 years (Caudullo and De Rigo 2016); thus, all artificial plantations in our study area are potentially able to reproduce via seeds for more than half a century. A sound cone yield an average of about 40 seeds, but it was estimated that a high proportion (> 80%) are aborted or attacked by pathogens (Dormont et al. 1996). Large wingless seeds of P. cembra are dispersed mainly by the corvid N. caryocatactes, which caches them as food reserves over winter but does not retrieve all seeds (Fourchy 1968). This bird is present more or less everywhere in the Prealps where P. cembra occurs, notably also where planted stands are located. Thus, neither seed production nor dispersal seem to be limiting factors that could prevent planted stands from regenerating. However, further study should be conducted to determine whether the seeds were viable, e.g., due to possible inbreeding effects (Salzer and Gugerli 2012).
Ecotypes. We did not find any information about the origin of the seeds used in the afforestation with P. cembra. We only know that plant nurseries were mainly regional. According to the selected ecotype and the region of origin, it may be possible that certain plants were not well adapted to the local conditions, which additionally decreased the chances of success. It was demonstrated experimentally for P. cembra that significant differences (e.g. height growth, diameter at root-collar and number of buds) can be observed between provenances (Blada 1997). Preliminary results of our parallel genetic investigation, performed with plant material collected in both natural populations and planted stands of the canton of Fribourg, indicate significant genetic differentiation between natural and planted stands (Sonnenwyl 2021). Thus, the seeds used for the production of seedlings were probably not collected from the natural populations in the canton of Fribourg. However, the use of seeds of very remote origin for local afforestation was not exceptional in the nineteenth century (Lendvay et al. 2014; Raffl et al. 2018). Because of the dangerous topography of natural populations in the canton of Fribourg, seed collection was likely easier in better accessible regions.
Conclusions
Many reasons could explain the low success of afforestation with P. cembra in the Western Prealps. Most probably not a single cause but a combination of different factors led to this unsatisfactory result. In the particular context of the canton of Fribourg, topography seems however to be one of the main points.
Forests are complex ecosystems, and it is nearly impossible to obtain an overview of all the interacting factors. Worldwide, experiences show that even traditional forms of timber plantations can be risky operations (Corlett 1999; Lamb et al. 2005; Cao et al. 2009). Large-scale and well-documented failures, such as those presented in our study, have rarely been reported in European forests, and must serve as lessons. Recommendations proposed by Di Sacco et al. (2020) seems particularly relevant in this context.
Availability of data and material
All data available in supplementary material or by the Zenodo open-access repository (Shapefiles, https://doi.org/10.5281/zenodo.4603114).
References
Apetrei CL, Tuchilus C, Aprotosoaie AC et al (2011) Chemical, antioxidant and antimicrobial investigations of Pinus cembra L. Bark Needles Molecules 16:7773–7788. https://doi.org/10.3390/molecules16097773
Barbeito I, Brücker RL, Rixen C, Bebi P (2013) Snow fungi induced mortality of Pinus cembra at the alpine treeline: evidence from plantations. Arct Antarct Alp Res 45:455–470
Berryman AA (2002) Population: a central concept for ecology? Oikos 97:439–442
Blada I (1997) Stone pine (Pinus cembra L.) provenance experiment in Romania. Silvae Genetica 46:197
Boden S, Pyttel P, Eastaugh CS (2010) Impacts of climate change on the establishment, distribution, growth and mortality of Swiss stone pine (Pinus cembra L.). iForest - Biogeosciences and Forestry 3:82. https://doi.org/10.3832/ifor0537-003
Breitenmoser U (1998) Large predators in the Alps: the fall and rise of man’s competitors. Biol Cons 83:279–289. https://doi.org/10.1016/S0006-3207(97)00084-0
Brubaker KM, Myers WL, Drohan PJ et al (2013) The use of LiDAR terrain data in characterizing surface roughness and microtopography. Appl Environ Soil Sci. https://doi.org/10.1155/2013/891534
Burga CA, Perret R, Vonarburg C (1998) Vegetation und Klima der Schweiz seit dem jüngeren Eiszeitalter. Ott, Thun
Cao S, Chen L, Yu X (2009) Impact of China’s Grain for Green Project on the landscape of vulnerable arid and semi-arid agricultural regions: a case study in northern Shaanxi Province. J Appl Ecol 46:536–543. https://doi.org/10.1111/j.1365-2664.2008.01605.x
Casalegno S, Amatulli G, Camia A et al (2010) Vulnerability of Pinus cembra L. in the Alps and the Carpathian mountains under present and future climates. For Ecol Manage 259:750–761
Caudullo G, De Rigo D (2016) Pinus cembra in Europe: distribution, habitat, usage, and threats. In: San-Miguel-Ayanz J, De Rigo D, Caudullo G et al. (eds) European Atlas of Forest Tree Species, Off. EU. Publ. Off. EU Luxembourg, Luxembourg
Conseil d’Etat du canton de Fribourg (1973) Arrêté concernant la protection de la faune et de la flore fribourgeoise
Corlett RT (1999) Environmental forestry in Hong Kong: 1871–1997. For Ecol Manage 116:93–105. https://doi.org/10.1016/S0378-1127(98)00443-5
Cottam G, Curtis JT (1956) The use of distance measures in phytosociological sampling. Ecology 37:451–460
Cottet M, Castella F (1891) Guide du botaniste dans le canton de Fribourg. Fribourg
Dauphin B, Rellstab C, Schmid M et al (2021) Genomic vulnerability to rapid climate warming in a tree species with a long generation time. Glob Change Biol 27:1181–1195. https://doi.org/10.1111/gcb.15469
Dauphin B, Wüest RO, Brodbeck S et al (2020) Disentangling the effects of geographic peripherality and habitat suitability on neutral and adaptive genetic variation in Swiss stone pine. Mol Ecol 29:1972–1989. https://doi.org/10.1111/mec.15467
Di Sacco A, Hardwick K, Blakesley D, et al (2020) Ten Golden Rules for Reforestation to Optimise Carbon Sequestration, Biodiversity Recovery and Livelihood Benefits. Global Change Biology. https://doi.org/10.1111/gcb.15498
Dormont L, Roques A, Trosset L (1996) Insect damage to cones and other mortality factors limiting natural regeneration potential of Swiss stone pine (Pinus cembra L) in the northern French Alps. In: Annales des sciences forestières. EDP Sciences, pp 153–158
Doutaz J, Bugmann H, Frey H-U (2006) Arolle et climat océanique: l’importance des facteurs micro-stationnels| Swiss stone pine and oceanic climate: the importance of microsites. Schweizerische Zeitschrift Fur Forstwesen 157:196–207
EPPO (2021) Data sheet on Gremmeniella abietina,. European and Mediterranean Plant Protection Organization EPPO
Etat de Fribourg (2021a) Archives de l’Etat de Fribourg (AEF)/Staatsarchiv Freiburg (StAF). https://www.fr.ch/cha/aef. Accessed 1 Oct 2020
Etat de Fribourg (2021b) Service des forêts et de la nature (SFN)/Amt für Wald und Natur (WNA). https://www.fr.ch/diaf/sfn. Accessed 1 Oct 2020
Ettlinger P (1975) Untersuchungen über den Erfolg früherer Flyschaufforstungen. ETH Zurich
Fankhauser F (1898) Les inondations et le reboisement de nos montagnes : la Société des forestiers suisses aux autorités fédérales, cantonales et communales. Bern
Fourchy P (1968) Notes sur le Pin cembro (Pinus cembra L.) dans les Alpes françaises. Revue forestière française
Gernandt DS, López GG, García SO, Liston A (2018) Phylogeny and classification of Pinus. Taxon 54:29–42. https://doi.org/10.2307/25065300
Graser A (2013) Learning QGIS 2.0. Packt Publishing, Birmingham
Gugerli F, Rüegg M, Vendramin G (2009) Gradual decline in genetic diversity in Swiss stone pine populations (Pinus cembra) across Switzerland suggests postglacial re-colonization into the Alps from a common eastern glacial refugium. Bot Helv 119:13
Gugerli F, Senn J, Anzidei M et al (2001) Chloroplast microsatellites and mitochondrial nad1 intron 2 sequences indicate congruent phylogenetic relationships among Swiss stone pine (Pinus cembra), Siberian stone pine (Pinus sibirica), and Siberian dwarf pine (Pinus pumila). Mol Ecol 10:1489–1497. https://doi.org/10.1046/j.1365-294X.2001.01285.x
Hao Z-Z, Liu Y-Y, Nazaire M et al (2015) Molecular phylogenetics and evolutionary history of sect. Quinquefoliae (Pinus): Implications for Northern Hemisphere biogeography. Mol Phylogenet Evol 87:65–79
Höhn M, Gugerli F, Abran P et al (2009) Variation in the chloroplast DNA of Swiss stone pine (Pinus cembra L.) reflects contrasting post-glacial history of populations from the Carpathians and the Alps. J Biogeogr 36:1798–1806. https://doi.org/10.1111/j.1365-2699.2009.02122.x
Hürlimann K (2008) Industrie du bois. In: Dictionnaire historique de la Suisse (DHS). Bern
Info Flora (2021) The National Data and Information Center on the Swiss Flora. https://www.infoflora.ch/en/. Accessed 1 Oct 2020
Jacoby WG (2000) Loess: a nonparametric, graphical tool for depicting relationships between variables. Elect Stud 19:577–613
Kaplan JO, Krumhardt KM, Zimmermann N (2009) The prehistoric and preindustrial deforestation of Europe. Quatern Sci Rev 28:3016–3034. https://doi.org/10.1016/j.quascirev.2009.09.028
Kasthofer K (1828) Der Lehrer im Walde: ein Lesebuch für schweizerische Landschulen, Landleute und Gemeindsverwalter, welche über die Waldungen zu gebieten haben
Körner C (2012) Alpine treelines: functional ecology of the global high elevation tree limits. Springer Science & Business Media, Cham
Kronfuss H, Havranek WM (1999) Effects of elevation and wind on the growth of Pinus cembra L. in a subalpine afforestation. Phyton 39:99–106
Lamb D, Erskine PD, Parrotta JA (2005) Restoration of degraded tropical forest landscapes. Science 310:1628–1632. https://doi.org/10.1126/science.1111773
Lendvay B, Höhn M, Brodbeck S et al (2014) Genetic structure in Pinus cembra from the Carpathian Mountains inferred from nuclear and chloroplast microsatellites confirms post-glacial range contraction and identifies introduced individuals. Tree Genet Genomes 10:1419–1433
Lillie M (2000) Europe’s first farmers. Cambridge University Press, Cambridge
Loranger H, Zotz G, Bader MY (2017) Competitor or facilitator? The ambiguous role of alpine grassland for the early establishment of tree seedlings at treeline. Oikos 126:1625–1636. https://doi.org/10.1111/oik.04377
Lotter AF, Birks HJB (2003) The Holocene palaeolimnology of Sägistalsee and its environmental history–a synthesis. J Paleolimnol 30:333–342
Mitchell K (2010) Quantitative analysis by the point-centered quarter method. arXiv preprint arXiv:10103303
Moser M (1967) Die ektotrophe Ernährungsweise an der Waldgrenze. Mitt Forstl Bundesversuchsanst Wien 75:357–380
Mullenbach P (1982) Les reboisements au voisinage de la limite altitudinale de la végétation forestière (limite sylvestre). L’exemple de la station du Chazelet. Premiers résultats. Revue Forestière Française
Mullenbach P (2000) Reboisements d’altitude. Editions Quae
Müller U (1990) Schutzwaldaufforstungen des Staates Freiburg im Senseoberland : forstpolitische Massnahmen des Staates Freiburg seit 1850 am Beispiel der Schutzwaldaufforstungen im Flyschgebiet des Senseoberlandes. ETH Zürich
Naudts K, Chen Y, McGrath MJ et al (2016) Europe’s forest management did not mitigate climate warming. Science 351:597–600. https://doi.org/10.1126/science.aad7270
Neet CR, Bolliger M (2004) Biodiversity management in Swiss mountain forests. Ecol Bullet, pp 101–108
Neuschulz EL, Merges D, Bollmann K et al (2018) Biotic interactions and seed deposition rather than abiotic factors determine recruitment at elevational range limits of an alpine tree. J Ecol 106:948–959
Nussbaumer C (2016) L’arolle rattrapé par l’épicéa ? Analyse des conditions de régénération et de développement de l’arolle (Pinus cembra) dans les Préalpes romandes (Suisse). Master thesis, Berner Fachhochschule
Oberhuber W, Geisler TA, Bernich F, Wieser G (2019) Weak Apical Control of Swiss Stone Pine (Pinus cembra L.) May Serve as a Protection against Environmental Stress above Treeline in the Central European Alps. Forests 10:744. https://doi.org/10.3390/f10090744
Petäistö R-L, Heinonen J (2003) Conidial dispersal of Gremmeniella abietina: climatic and microclimatic factors. Forest Pathol 33:363–373. https://doi.org/10.1111/j.1439-0329.2003.00335.x
QGIS Development Team (2021) QGIS Geographic Information System. QGIS Association
R Core Team (2018) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria
Raffl H, Konrad H, Curtu LA, Geburek T (2018) Genetic evidence of human mediated, historical seed transfer from the Tyrolean Alps to the Romanian Carpathians in Larix decidua (Mill.) forests. Ann for Sci 75:1–13
Rainer G, Kuhnert R, Unterholzer M et al (2015) Host-Specialist Dominated Ectomycorrhizal Communities of Pinus cembra are not Affected by Temperature Manipulation. Journal of Fungi 1:55–75. https://doi.org/10.3390/jof1010055
Rikli M (1909) Die Arve in der Schweiz. Georg & Cie, Basel, Genf, Lyon
Riley SJ, DeGloria SD, Elliot R (1999) Index that quantifies topographic heterogeneity. intermountain Journal of sciences 5:23–27
Salzer K, Gugerli F (2012) Reduced fitness at early life stages in peripheral versus core populations of Swiss stone pine (Pinus cembra) is not reflected by levels of inbreeding in seed families. Alp Bot,pp 75–85. https://doi.org/10.1007/s00035-012-0106-z
Schönenberger W (2001) Cluster afforestation for creating diverse mountain forest structures—a review. For Ecol Manage 145:121–128
Schönenberger W (1975) Standortseinflusse auf Versuchsaufforstungen an der alpinen Waldgrenze (Stillberg, Davos). Beer & Co., Zürich
Schönenberger W, Frey W, Leuenberger F (1990) Ecology and techniques of afforestation in the mountains. Practical recommendations. Ecology and techniques of afforestation in the mountains Practical recommendations
Schönenberger W, Senn J, Wasem U (1995) Factors affecting establishment of planted trees, including European larch, near the alpine timberline. Ecology and Management of Forests: a Look Ahead’(Eds WC Schmidt, KJ McDonald) USDA Forest Service, Intermountain Research Station General Technical Report INT-GTR-319 170–175
Schuler A (2008) Lois sur les forêts. In: Dictionnaire historique de la Suisse (DHS). Bern
Schütt P (2014) Pinus cembra. Enzyklopädie der Holzgewächse: Handbuch und Atlas der Dendrologie. Wiley-VCH, Weinheim, pp 1–12
Senn J (1999) Tree mortality caused by Gremmeniella abietina in a subalpine afforestation in the central Alps and its relationship with duration of snow cover. Eur J for Pathol 29:65–74. https://doi.org/10.1046/j.1439-0329.1999.00131.x
Senn J, Schönenberger W (2001) Zwanzig Jahre Versuchsaufforstung Stillberg: Überleben und Wachstum einer subalpinen Aufforstung in Abhängigkeit vom Standort| Twenty years experimental afforestation Stillberg: Survival and growth in a subalpine afforestation in relation to site conditions. Schweizerische Zeitschrift Fur Forstwesen 152:226–246
Sonnenwyl V (2021) History, distribution and genetic diversity of natural populations and planted stands of Pinus cembra (Pinaceae) in Western Prealps: conservation and sylvicultural implications. Master thesis, University of Fribourg
Stalder J-M, Pfister-Ritter M, Pfister-Ritter F (1990) Geschichte einer Wechselvollen Beziehung : Aufforstungen im Sense-Oberland/FR : 100 Jahre. Département cantonal des forêts/Direction de l’intérieur et de l’agriculture, Fribourg
Swisstopo (2021) Federal Office of Topography swisstopo. https://www.swisstopo.admin.ch/fr/home.html. Accessed 15 Jan 2021
Tomback DF, Holtmeier F-K, Mattes H et al (1993) Tree clusters and growth form distribution in Pinus cembra, a bird-dispersed pine. Arct Alp Res 25:374–381
Tomback DF, Linhart YB (1990) The evolution of bird-dispersed pines. Evol Ecol 4:185–219
Tóth EG, Tremblay F, Housset JM et al (2019) Geographic isolation and climatic variability contribute to genetic differentiation in fragmented populations of the long-lived subalpine conifer Pinus cembra L. in the western Alps. BMC Evol Biol 19:1–17
Tranquillini W (2012) Physiological ecology of the alpine timberline: tree existence at high altitudes with special reference to the European Alps. Springer Science & Business Media
Ulber M, Gugerli F, Bozic G (2004) EUFORGEN technical guidelines for genetic conservation and use for swiss stone pine (Pinus cembra). International Plant Genetic Resources Institute, Rome, Italy 6:
Van Kooten GC, Shaikh SL, Suchánek P (2002) Mitigating climate change by planting trees: the transaction costs trap. Land Econ 78:559–572
Wilcoxon F, Katti SK, Wilcox RA (1970) Critical values and probability levels for the Wilcoxon rank sum test and the Wilcoxon signed rank test. Selected Tables in Mathematical Statistics 1:171–259
Zheng H, Wang Y, Chen Y, Zhao T (2016) Effects of large-scale afforestation project on the ecosystem water balance in humid areas: An example for southern China. Ecol Eng 89:103–108. https://doi.org/10.1016/j.ecoleng.2016.01.013
Zięba A, Różański W, Szwagrzyk J (2020) Structure of Dominance among Tree Species in Relic Swiss Stone Pine (Pinus cembra L.) Forests in Tatra Mountains. pjoe 68:159–171. https://doi.org/10.3161/15052249PJE2020.68.2.005
Acknowledgements
We would like to thank all the persons who helped in the field work and in the archives, notably, Dominique Schaller and Pascal Sonnenwyl, numerous foresters and forest engineers from the Forest and Nature Service of the State of Fribourg, and the collaborators from the Archives of the State of Fribourg. We would also like to thank Felix Gugerli from the Swiss Federal Institute for Forest, Snow and Landscape Research (WSL) and Christian Parisod from the University of Bern for reviewing the manuscript and their valuable input. Finally, we would like to thank Peter Wandeler, Emanuel Gerber, Henri Descombes, Gilles Hauser and Sophie Giriens from the Natural History Museum of Fribourg for their valuable support.
Funding
Open Access funding provided by Université de Fribourg. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
YF and GK developed the research idea; VS, BC, YF and GK conducted the field work; VS led the research in the archives; YF analyzed the data and realized the graphs and maps; YF and GK led the writing of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Summary of actual and historical data
Supplementary file1 (PDF 216 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fragnière, Y., Sonnenwyl, V., Clément, B. et al. Large-scale historical afforestation failure with Pinus cembra in the Swiss Prealps. New Forests 53, 533–553 (2022). https://doi.org/10.1007/s11056-021-09871-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-021-09871-0