
Abstract
Glioblastoma (GBM) is one of the most frequent primary brain tumors. Limited therapeutic options and high recurrency rates lead to a dismal prognosis. One frequent, putative driver mutation is the genomic amplification of the oncogenic receptor tyrosine kinase EGFR. Often accompanied by variants like EGFRvIII, heterogenous expression and ligand independent signaling render this tumor subtype even more difficult to treat, as EGFR-directed therapeutics show only weak effects at best. So EGFR-amplified GBM is considered to have an even worse prognosis, and therefore, deeper understanding of molecular mechanisms and detection of potential targets for novel therapeutic strategies is urgently needed. In this study, we looked at the level of microRNAs (miRs), small non-coding RNAs frequently deregulated in cancer, both acting as oncogenes and tumor suppressors. Comparative analysis of GBM with and without EGFR amplification should give insight into the expression profiles of miRs, which are considered both as potential targets for directed therapies or as therapeutic reagents. Comparison of miR profiles of EGFR-amplified and EGFR-normal GBM revealed an upregulation of the miR-183/96/182 cluster, which is associated with oncogenic properties in several tumor entities. One prominent target of this miR cluster is FOXO1, a pro-apoptotic factor. By observing FOXO1 downregulation in EGFR-amplified tumors, we can see a significant correlation of EGFR amplification, miR-183/96/182 cluster upregulation, and repression of FOXO1. Although no significant difference in overall survival is shown, these data may contribute to the molecular understanding of this tumor subtype and offer potential targets for miR-based therapies.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Glioblastoma (GBM) is one of the most frequent and fatal primary malignancy of the central nervous system. Despite advances in therapy, the prognosis remains dismal with a median survival of about 12–14.6 months after diagnosis and full treatment [1], which standardly consists of surgical resection, radiotherapy, and chemotherapy with temozolomide (TMZ) [1, 2]. However, the recurrence rate is very high and diffuse infiltrative growth makes complete tumor removal nearly impossible. Given the high inter- and intratumoral heterogeneity of GBM [3], increased efforts in molecular profiling are necessary to facilitate development of more targeted therapy options to overcome the limitations of the current standard GBM treatment. One frequent mutation is the amplification of the receptor tyrosine kinase EGFR, which occurs in almost 60% of primary GBM [4] and is associated with a worse prognosis [5, 6] Increased EGFR activity leads to enhanced signaling in several tumor associated downstream pathways like RAS or PI3K (reviewed in [7] and [8]). In around 50% of tumors carrying an EGFR amplification, accompanying expression of additional mutational variants is seen [8], the most frequent being EGFRvIII, a deletion variant lacking exons 2–7 leading to a loss of the extracellular ligand-binding domain. The ability to heterodimerize with wild-type EGFR and other receptors of the ERBB family leads to ligand independent signaling [9] and, therefore, to ectopic activation of downstream targets. Furthermore, the existence of different forms of aberrant EGFR leads to a heterogenous expression pattern and, therefore, heterogenous signaling abilities [10], making this tumor entity even more difficult to target with specific therapeutic approaches. Indeed, in contrast to several other tumor entities (lung, colon), targeting EGFR and/or EGFRvIII with inhibitors, or antibodies showed hardly any significant therapeutic response in GBM [7, 11,12,13]. Possible reasons for resistance to EGFR/EGFRvIII-targeted therapy are blood–brain barrier, tumor heterogeneity, extrachromosomal localization of EGFR, and EGFRvIII amplicons in double minutes, and mutation of genes in downstream pathways [7]. Furthermore, tyrosine kinase inhibitors are rather inefficient, as they specifically target sites of the kinase domain which are mutated for instance in lung cancer, but not in GBM [7]. EGFR-directed antibodies seem not to be able to inactivate all mutant forms like EGFRvIII, as they rather lead to protein recycling than to degradation [13]. As direct targeting of EGFR and EGFRvIII proved difficult and, so far, inefficient, other therapeutic strategies are urgently need. One potential level of targeted therapy approaches could be the level of microRNAs (miRs). MiRs are small non-coding RNAs, which bind to their target mRNAs to inhibit protein expression. They are frequently deregulated in multiple cancers. Deregulation can occur at any point of miR-biogenesis and has, depending on the miR and tissue or tumor type involved, oncogenic or tumor suppressive effects [14]. Importantly, a given miR can be oncogenic in one tumor type, and tumor suppressive in another [15]. In GBM, several miRs, like miR-21 with oncogenic or miR-34a with tumor suppressive properties, are involved in tumorigenesis, impacting diverse cancer signatures therein [16]. Furthermore, miRs are deemed therapeutic targets and can be inhibited or re-expressed in tumors [17]. Therefore, application of anti-miRs coupled to nanoparticles, shown to be functional in vitro, is hypothesized to successfully cross the blood–brain barrier to deliver the therapeutic agents directly to the tumor [18, 19]. Additionally, several miRs have been identified to repress MGMT expression and increase sensitivity to alkylating agents [20].
Information about correlations of EGFR alterations and changes of miR expression is scarce. One study showed a change of miR-9 expression in vitro for cells expressing EGFRvIII [9, 21]. Another study comparing miR profiles showed downregulation of miR-200c correlating with EGFR amplification [22]. To assess whether EGFR amplification has an influence on miR expression, we compared the miR expression profiles of EGFR amplified to those of EGFR-normal GBM. The aim of this study is the identification of commonly differentially expressed miRs and/or their target genes, which could give hints to potential targets for miR-associated or otherwise targeted therapeutic strategies. Our results show a significant upregulation of the miR-183/96/182 cluster in EGFR-amplified tumors. This cluster, consisting of miR-183, miR-96, and miR-182, is deregulated in different tumor entities and mostly considered as oncogenic [23]. Although members of the cluster are known to play a role in GBM [24, 25], no correlation of EGFR amplification and miR-183/96/182 cluster upregulation has been reported to date. Additionally, FOXO1, a prominent target of the miR-183/96/182 cluster [23] and associated with pro-apoptotic and tumor suppressive properties [26,27,28] is downregulated in EGFR-amplified GBM. Our findings should contribute to a better understanding of the role of microRNAs in EGFR-amplified GBM and help facilitate the development of new therapeutic strategies, such as those combining miR-based approaches with, so far widely inefficient, EGFR inhibition.
Materials and methods
Tumor samples
Formalin-fixed paraffin-embedded (FFPE) tumor material from glioblastoma patients was obtained from the archive of the Institute for Pathology, University Medicine, Rostock. Specimen collection was conducted in accordance with the ethics guidelines for the use of human material, approved by the Ethics Committee of the University of Rostock (Reference number: A 2009/34) and with informed written consent from all patients prior to surgery. Hematoxylin and eosin (H&E)-stained sections were examined by an experienced pathologist to ensure sample sufficiency and quality. Samples with high content of necrotic tissue were excluded from the study. Patient data are listed in Supplementary Table 1.
Tissue microarrays
Tissue microarrays (TMA) were created using a Manual Tissue Arrayer MTA-1 (Beecher Instruments, Sun Prairie, WI, USA) with 1-mm diameter punches. From the donor blocks, three punches per sample were taken from tumor regions suitable for analysis (high tumor cell content, no necrosis) and transferred to an empty acceptor block. Afterwards, blocks were heated to 50 °C, and the correct placement of the punches was confirmed by microscopic examination. A final quality check of the TMA was done by H&E staining to confirm that all punches contained the desired amount of tumor tissue for further analysis.
Chromogenic in situ hybridization
For determination of EGFR amplification status, two-colored chromogenic in situ hybridization (2C CISH) was performed. Microtome sections of 4 µm thickness of samples or TMA were mounted on coated slides. EGFR-specific 2C CISH was performed using the ZytoDot 2C CISH implementation Kit (Zytomed Systems, Berlin, Germany) and the ZytoDot 2C SPEC EGFR/CEN 7 Probe (Zytomed Systems) according to the manufacturer’s protocol. Analysis of the processed samples was performed with bright-field microscopy using a 40× objective. Each red signal specifically represented the centromere of chromosome 7 for reference and ploidy determination. Green signals were specific for the EGFR gene. Cells were considered to carry EGFR amplification if the ratio of green signals to red signals was greater than 2 or if green signals occurred in clusters.
microRNA extraction
For microRNA extraction, the miRNeasy FFPE-Kit (Qiagen, Hilden, Germany) was used for FFPE sections and the Isolate II miRNA Kit (Bioline, Luckenwalde, Germany) for cell pellets, following the manufacturers’ protocols. Concentration of extracted RNA was determined with a Nanodrop spectrometer (Peqlab, Erlangen, Germany).
miR screening arrays
Analysis of microRNA expression was performed using the Nanostring nCounter System with the Human v3 miRNA assay (Nanostring, Seattle, WA, USA). The analysis procedure was performed with 250 ng per sample by a service provider lab (Transcriptome and Genome Analysis Laboratory (TAL), Microarray and Deep-Sequencing Facility, University Medicine Göttingen) and analyzed by the authors with the nSolver 2.6 software (Nanostring) using standard settings for background subtraction and normalization. Group-wise comparison (EGFR-amp vs. EGFR-norm) was performed, delivering fold change and p values.
microRNA-specific quantitative PCR
Expression analysis of miR-183-5p, miR-96-5p, and miR-182-5p was performed by qPCR using miR-specific TaqMan Assays (Applied Biosystems, Darmstadt, Germany), containing RT-primers specific for the mature miR and corresponding primer/probe mixes with RNU6B as endogenous control (Applied Biosystems). The specific reverse transcription of miR-183-5p, miR-96-5p, miR-182-5p, and RNU6B was performed with the TaqMan MicroRNA Reverse Transcription Kit (Applied Biosystems) according to the manufacturer’s protocol modified as follows: the reaction volume was scaled up to 30 µl and 20 ng RNA were used as template. The reverse transcription was carried out as a multiplex reaction, containing the RT-primer for all three miRs and the control in one reaction. The subsequent qPCR reactions were set up according to the manufacturer’s protocol using TaqMan Universal PCR Master Mix II, No UNG (Applied Biosystems) and the miR-specific primer/probe mixes. The runs were performed on a StepOne Plus Real-time PCR system (Applied Biosystems) and the data analyzed using the StepOne Software v2.1 (Applied Biosystems). Relative expression against RNU6B (fold change) was calculated using the ΔCt-algorithm.
Identification of target genes
Potential target genes of the miR-183 cluster for further analyses were selected by consulting online databases miRWalk 2.0 (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/) and miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/) and the review by Dambal et al. [23] concerning the miR-183/96/182 cluster.
Immunohistochemistry
As target-specific primary antibodies, a monoclonal mouse-anti-FOXO1, clone 3B6, dilution 1:200 (Biozol, Hamburg, Germany), and a monoclonal mouse-anti-EGFR, clone 3G143, dilution 1:200 (Zytomed Systems) were used. Slides were processed on an automatic IHC system, AutostainerLink48 (Dako, Hamburg, Germany) according to routine protocols. Expression was assessed by scoring the staining of three representative areas with 1 (negative / weak), 2 (moderate), or 3 (strong). A final score was built of the mean values of the triplicates. Samples were classified as FOXO1-low, medium, and high with final scores of < 1.5, 1.5–2.5, and > 2.5, respectively.
GBM in vitro model
The GBM cell line HROG33 was established from a patient-derived xenograft with experimentally validated EGFR amplification. The EGFR amplification status of HROG33 is adjustable by adaptation of culture conditions. It was cultivated in DMEM/Ham’s F12 with 2 mM L-glutamine and B-27 in an incubator (37 °C, 5% CO2, 95% relative humidity), either without EGF to maintain EGFR amplification or supplemented with 30 ng/ml EGF to reduce EGFR amplification to normal levels [29, 30].
For CISH and IHC analyses, cell pellets were fixed and embedded according to following procedure:
Cells were harvested, washed twice with PBS, and fixed immediately by resuspending pellets in 4% buffered formalin (Formafix; Grimm, Torgelow, Germany). Cells were subsequently processed following standard procedures [31] to form a conglomerate and then embedded in paraffin using the automated Excelsior AS system (Thermo Scientific, Dreieich, Germany).
For miR expression analysis, freshly harvested pellets were used, and miR extraction and miR-specific qPCR were performed as described above.
For mRNA expression analysis, freshly harvested pellets were used, RNA was extracted using the RNeasy Mini Kit (Qiagen) according to the manufacturer’s protocol. cDNA synthesis was performed using SuperScript II Reverse Transcriptase (Invitrogen, Thermo Fisher). Quantitative PCR was performed using the SensiFast Probe Kit (Bioline, Luckenwalde, Germany) and Primer/Probe Sets for EGFR, EGFRvIII (both TIB Molbiol, Berlin, Germany), FOXO1, and TBP (both AppliedBiosystems, Darmstadt, Germany) on an ABI StepOne Plus System (Applied Biosystems) according to the manufacturers’ instructions.
Statistics and box plots
Statistical analyses for the significance of Nanostring array results were embedded within the nSolver Software and p values provided in the results table. Further statistical analysis for expression values obtained by qPCR or IHC, Wilcoxon tests were applied using RStudio v1.2.5019 (RStudio PBC, Boston, USA). Boxplots were generated with the help of the online tool BoxPlotR (URL: http://shiny.chemgrid.org/boxplotr/). Statistical analyses involving patients’ characteristics and survival analyses were performed using SPSS Statistics, version 28 (IBM, Ehningen, Germany).
Results
Classification of GBM tumors by EGFR amplification status
Initially, for the miR-array analysis, tumor samples from 24 GBM patients were available (Median age 62.2 years, 12 male and 12 female, IDH1/2 mutation in 1/24 patients).
These samples were analyzed for EGFR amplification by EGFR-specific 2C CISH. In 13 samples’ clusters of green signals were observed indicating EGFR amplification (EGFR-amp), whereas an even distribution of green and red signals was seen in the remaining 11 samples, indicating EGFR-normal status (EGFR-norm; Fig. 1, Supplementary Fig. 1).

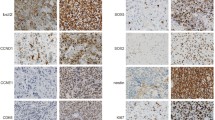
Examples for CISH analysis of glioblastomas with normal EGFR (top) and EGFR amplification (bottom). Red dots represent centromeres of chromosome 7, green dots are specific for EGFR. ×400 magnification. Inlays show, digitally zoomed, single cells representing EGFR-normal and EGFR-amplified status
For further analyses, samples from another 56 patients were obtained (median age 66.4 years, 32 male and 24 female; IDH status unknown), whereof 20 showed EGFR amplification and 36 had an EGFR-normal status (Supplementary Table 1). None of the samples showed an aneuploidy for chromosome 7.
Considering patients’ characteristics like sex and age of diagnosis, none of them showed significant correlations with EGFR amplification state.
miR screening revealed upregulation of the miR-183/96/182 cluster in EGFR-amp GBMs
The group-wise comparison of the miR expression profiles obtained by the Nanostring nCounter assays revealed only moderate changes in expression between the EGFR-amp and EGFR-normal tumors. The differential expression of miRs downregulated in EGFR-amp barely exceeded a factor of 3.5× (Supplementary Table 1) and were, therefore, not considered further in this study. Similarly, most miRs upregulated in EGFR-amp tumors did not show pronounced changes in expression levels compared to EGFR-norm group with factors up to 3.2× (Supplementary Table 2). Only three miRs—miR-182-5p, miR-96-5p, and miR-183-5p—clearly and significantly outperformed this expression range with upregulation factors of 10.7× (p = 0.0015), 6.9× (p = 0.0032), and 6.4× (p = 0.0019), respectively (Supplementary Table 2).
Verification of these array data was performed by quantitative PCR of mature miRs 182-5p, 96-5p, and 183-5p and confirmed their significant upregulation in EGFR-amp tumors showing a median overexpression of 5.56× (p = 0.003), 7.29× (p = 0.003), and 6.23× (p = 0.006), respectively (Fig. 2).

Verification of miR-array data for the initial 24 cases. Boxplot representation of miR-specific qPCR for EGFR-amplified samples (gray) and EGFR-normal samples (white) showing relative expression normalized against endogenous control RNU6B. Bars represent median values, boxes contain 50% of data points for each group, whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles, empty circles: outliers. Asterisks indicate significance (p < 0.05)
To evaluate the significance of the co-occurrence of EGFR amplification and miR-183/96/182 cluster expression, the analysis was extended to a total of 80 cases (including the samples used for initial array screening).
The miR-specific qPCR analyses revealed again significant higher expression of miR-182-5p, miR-96-5p, and miR-183-5p showing median fold changes of 4.33× (p = 0.003), 4× (p = 0.003), and 4× (p = 0.006), respectively (Fig. 3). On the miR cluster expression, neither patients’ age at diagnosis nor gender had a significant influence.

Expression analysis by miR-specific qPCR for all 80 cases. Boxplot representation of miR-specific qPCR for EGFR-amplified samples (gray) and EGFR-normal samples (white) showing relative expression normalized against endogenous control RNU6B. Bars represent median values, boxes contain 50% of data points for each group, whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles, empty circles: outliers. Asterisks indicate significance (p < 0.05)
Expression of miR-183/96/182 cluster target FOXO1 is decreased in EGFR-amp tumors
Combined search of miR-target databases and literature (based on the review of Dambal et al. [23]) revealed FOXO1 as the most prominent target of all three members of the miR-183/96/182 cluster.
FOXO1 immunohistochemistry was successful in 74 cases. Expression level scoring classified 31 (40.8%) samples as FOXO1-low, 33 (43.4%) samples as FOXO1-medium, and 12 (15.8%) samples as FOXO1-high.
EGFR-amplified tumors showed a significant (p = 0.004) lower expression of FOXO1 then EGFR-normal samples: in EGFR-normal samples, 11/42 (26.2%) showed low, 24/42 (57.1%) showed medium, and 7/42 (16.7%) showed high FOXO1 expression. In EGFR-amplified samples, 20/32 (62.5%) showed low, 7/32 (21.9%) showed medium, and 5/32 (15.6%) showed high expression. (Fig. 4a, b).

Expression analysis of miR-183/96/182 cluster target FOXO1. a Examples for FOXO1 IHC of glioblastomas with EGFR amplification (left) and normal EGFR (right) showing weak and strong FOXO1 staining, respectively. ×200 magnification. b Bar chart showing number of EGFR-amplified (top) and EGFR-normal (bottom) cases for each FOXO1 expression status (white = low; light gray = medium; and dark gray = high) (c) boxplot representation of miR-specific qPCR for samples with low (white), medium (light gray), and high (dark gray) FOXO1 expression, showing relative expression normalized against control RNU6B. Bars represent median values, boxes contain 50% of data points for each group, whiskers extend 1.5 times the interquartile range for the 25th and 75th percentiles, empty circles: outliers. Asterisks indicate significance (p < 0.05)
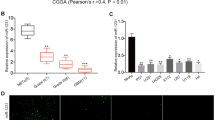
Correlation analysis of FOXO1 expression and miR expression revealed a significant higher expression of miR-183-5p, miR-96-5p, and miR-182-5p in FOXO1-low samples than in FOXO1-med and FOXO1-high samples (Fig. 4c).
miR-183-5p showed in FOXO1-low samples a medium fold change of 6.5× compared to FOXO1-medium samples (p = 0.0009), and of 13× compared to FOXO1-high samples (p = 0.0045).
miR-96-5p showed in FOXO1-low samples a medium fold change of 5× compared FOXO1-medium samples (p = 0.0021) and of 5× compared to FOXO1-high samples (p = 0.0037).
miR-182-5p showed in FOXO1-low samples a median fold change of 7.4× compared to FOXO1-medium samples (p = 0.0004) and 12.33× compared to FOXO1-high samples (p = 0.0013).
Neither patients’ age at diagnosis nor gender had a significant influence to FOXO1 expression.
Inverse correlation of EGFR and FOXO1 expression in vitro
Cultured HROG33 cells, either cultured under standard condition with 30 ng EGF/ml medium (termed “33–30”) or without EGF supplement (termed “33–0”), were comparatively analyzed with regard to EGFR amplification and expression of EGFR, EGFRvIII, and FOXO1. 33–30 cells lost their EGFR amplification, whereas 33–0 cells retained the amplified status, as shown by CISH in Fig. 5a, top row. For EGFR and EGFRvIII, a stronger protein expression, analyzed by IHC, was seen in 33–0 cells, EGFRvIII showing an even stronger staining than EGFR (Fig. 5a, second and third row). For FOXO1, an inverse correlation with EGFR amplification and EGFR and EGFRvIII expression was observable, as 33–0 cells show a weaker stain in FOXO1 IHC.

a Analysis of HROG33 cells grown with 30 ng (left, 33–30) and without (right, 33–0) EGF Representative pictures showing, from top to bottom, EGFR amplification by CISH analysis; EGFR expression, EGFRvIII expression, and FOXO1 expression by IHC. Magnification ×400. b Relative quantification of EGFR expression, EGFRvIII expression, and FOXO1 expression by qPCR. c Relative expression analysis of miRs 183-5p, 96-5p, and 182-5p by miR-specific qPCR. Expression values of 33–30 cells are used as calibrators and set to 1
Interestingly, looking at the mRNA level, the expression of EGFR (wt) is in 33–0 cells surprisingly weaker than in 33–30 cells (Fig. 5b, left), indicating a post-transcriptional control of EGFR expression. For EGFRvIII, the increase of expression in 33–0 cells is immense, which reflects the IHC data, a fold change of more than 45× compared to 33–30 cells (Fig. 5b, middle). FOXO1 shows on the mRNA level a slight decrease of expression in 33–0 cells compared to 33–30 cells (Fig. 5b, right). This is concordant with the IHC data, although the effect on the protein level seems to be stronger, also indicating possible post-transcriptional regulatory mechanisms. The expression levels of miR-183-5p, miR-96-5p, and miR-182-5p remain virtually unchanged (Fig. 5c), unlike the observed upregulated miR expression in EGFR-amplified primary tumors.
Discussion
MicroArray-profiling analysis revealed upregulation of the miR-183/96/182 cluster in EGFR-amplified glioblastoma, comprising the mature miRs 183-5p, 96-5p, and 182-5p. Regarding individual miR expression values, these partly overlapped between the groups, EGFR-amp and EGFR-norm. Nevertheless, in the group-wise comparison, the upregulation of these three miRs was significant. The initially mentioned EGFR amplification-dependent upregulated miR-9 [21] showed in our study a fold change of 1.99, whereas miR-200c, significant target in the study of Serna [22] revealed a fold change of − 2.59, both not reaching top level, yet significant, deregulation (Suppl. Table 1). It must be mentioned that for miR-200 (a, b, and c), only the mature -3p miRs are included in the assay.
The three members of the miR cluster are considered to play multiple roles in several types of cancer, influencing many cellular functions and targeting a multitude of mRNAs, partly in common, partly unique for each of these miRs [23].
In glioblastoma and other gliomas, the miR-183/96/182 cluster is mostly considered to be oncogenic. MiR-183 expression denotes a worse prognosis [32], increased proliferation, and invasion by inhibiting NEFL leading to upregulation of mTOR [25] and promoting angiogenesis by HIF1A upregulation via IDH2 repression [33]. Although not much is known about the role of miR-96 in glioblastoma, it is associated with oncogenic functions by influencing the Wnt-pathway, combined with worse prognosis [34] and enhanced angiogenesis [35]. Nevertheless, in some other cancers, e.g., nasopharyngeal or pancreatic cancer, it is considered to have tumor suppressor functions targeting MTAI, thus, inhibiting proliferation and invasion [36, 37].
The role of miR-182 in glioblastoma is regarded as controversial. In several studies, miR-182 expression is correlated with prolonged survival [38] and enhanced sensitivity to chemotherapy and may oppose tumorigenesis [39]. Additionally, pro-apoptotic effects have been described [40, 41], and this miR downregulates growth and migration of GBM cells [39, 42]. In contrast, other studies have shown an association of miR-182 expression with a worse prognosis [43, 44] and oncogenic features, like growth promotion [45], sustained NF-kB activation [46] and enhanced invasion [47]. miR-182 is described to be activated by TGFb, which in turn can be stimulated by EGFR/EGFRvIII signaling via miR-524 silencing [48]. However, our Nanostring based miR-profiling showed no differential expression of miR-524.
Furthermore, it is shown that the knockdown of the whole cluster increases apoptosis and sensitivity to temozolomide in glioblastoma [24]. Also in other tumor entities, the miR-183/96/182 cluster is widely considered to be oncogenic [23].
FOXO1 is described as a prominent target, of all three members of the miR-183/96/182 cluster [23]. In our study, the expression of FOXO1 was lower in EGFR-amplified tumors. Although IHC is not considered a quantitative method, the differences of FOXO1 expression between EGFR-amplified and EGFR-normal tumors were obvious. In lung cancer, FOXO1 repression by miR-183 is described as an anti-apoptotic mechanism [26]. FOXO1 is also considered as a pro-apoptotic factor in GBM [27] and even as a potential target of directed therapy [28].
The reportedly worse prognosis of EGFR-amplified tumors [6] is consistent with a mechanistic scenario whereby EGFR amplification leads to repression of FOXO1, either via the miR-183/96/182 cluster, direct targeting by EGFR and/or downstream factors, or a synergistic effect of both. Loss of its anti-apoptotic activity may render these tumors more aggressive.
The in vitro data obtained in this study contradict the direct influence of EGFR to the miR cluster, at least in the single-cell line model available. Modulation of EGFR amplification and expression did not lead to a differential miR-183/96/182 cluster expression, but, identical to the majority of the patients’ samples, the FOXO1 expression was inversely correlated with EGFR amplification and expression. This supports other, or additional, miR-183/96/182 cluster-independent effects of EGFR expression to FOXO1 regulation. In accordance with this it is known that EGFR signaling can promote downregulation of FOXO-genes via the AKT pathway activation leading to inhibitory phosphorylation [49].
Interestingly, for EGFR and FOXO1 expression, the EGFR modulation showed effects rather on the protein level. The changes on the RNA level are rather minor (FOXO1) or even contrary (EGFR), indicating post-transcriptional effects. For EGFRvIII, the high increase of RNA expression is accordant to the observed increased protein expression, maybe rendering EGFRvIII as a principal oncogenic component for this cell line model.
The reason for the lack of influence to the miR cluster expression remains speculative. Maybe other control mechanisms of miR expression predominate in this cell line model as several transcription factors are known to play a role in regulating the whole cluster or single miRs thereof (reviewed in [23]). In contrast to primary tumors analyzed, the in vitro model lacks a tumor microenvironment, which could also have an effect to differential miR expression. Not all the primary tumors with EGFR amplification show miR cluster upregulation and FOXO1 downregulation, demonstrating heterogeneity. Thus, the results of only a single-cell line model, especially in consideration of this heterogeneity, are not eligible to draw common conclusions. Therefore, intensifying studies with more models, maybe in vivo PDX tumors with a microenvironment, and functional assays with overexpression as well as silencing of miR-183-5p, miR-96-5p, and miR-182-5p would be necessary in the future to give more inside into these complex mechanisms.
Despite yet lacking evidence for a functional relationship, we have shown that there exist significant correlations between EGFR amplification, upregulation of the miR-183/96/182 cluster and FOXO1 downregulation, rendering this miR cluster an interesting subject for further analysis concerning its influence on tumorigenicity and its potential as target for directed therapy by silencing approaches.
In summary, this study shows an upregulation of the miR-183/96/182 cluster in EGFR-amplified glioblastoma, accompanied by reduced expression of FOXO1. Although the functional context remains to be identified, these data provide a solid molecular basis for further functional in vitro or in vivo assays to analyze the effects of miR-183/96/182 cluster and FOXO1 deregulation on proliferation, tumorigenicity, or chemoresistance, and to investigate whether they offer potential targets for new directed therapy approaches.
Data availability
Raw data and images can be obtained from the authors.
Code availability
Not applicable.
References
Stupp R, Mason WP, van den Bent MJ et al (2005) Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med 352:987–996. https://doi.org/10.1056/NEJMoa043330
Stupp R, Hegi ME, Mason WP et al (2009) Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol 10:459–466. https://doi.org/10.1016/S1470-2045(09)70025-7
Cloughesy TF, Cavenee WK, Mischel PS (2014) Glioblastoma: from molecular pathology to targeted treatment. Annu Rev Pathol 9:1–25. https://doi.org/10.1146/annurev-pathol-011110-130324
Brennan CW, Verhaak RGW, McKenna A et al (2013) The somatic genomic landscape of glioblastoma. Cell 155:462–477. https://doi.org/10.1016/j.cell.2013.09.034
Furnari FB, Fenton T, Bachoo RM et al (2007) Malignant astrocytic glioma: genetics, biology, and paths to treatment. Genes Dev 21:2683–2710. https://doi.org/10.1101/gad.1596707
Shinojima N, Tada K, Shiraishi S et al (2003) Prognostic value of epidermal growth factor receptor in patients with glioblastoma multiforme. Cancer Res 63:6962–6970
An Z, Aksoy O, Zheng T et al (2018) Epidermal growth factor receptor and EGFRvIII in glioblastoma: signaling pathways and targeted therapies. Oncogene 37:1561–1575. https://doi.org/10.1038/s41388-017-0045-7
Oprita A, Baloi S-C, Staicu G-A et al (2021) Updated insights on EGFR signaling pathways in glioma. Int J Mol Sci. https://doi.org/10.3390/ijms22020587
Guo G, Gong K, Wohlfeld B et al (2015) Ligand-independent EGFR signaling. Cancer Res 75:3436–3441. https://doi.org/10.1158/0008-5472.CAN-15-0989
Furnari FB, Cloughesy TF, Cavenee WK et al (2015) Heterogeneity of epidermal growth factor receptor signalling networks in glioblastoma. Nat Rev Cancer 15:302–310. https://doi.org/10.1038/nrc3918
Padfield E, Ellis HP, Kurian KM (2015) Current therapeutic advances targeting EGFR and EGFRvIII in glioblastoma. Front Oncol 5:5. https://doi.org/10.3389/fonc.2015.00005
Bagley SJ, Kothari S, Rahman R et al (2022) Glioblastoma clinical trials: current landscape and opportunities for improvement. Clin Cancer Res 28:594–602. https://doi.org/10.1158/1078-0432.CCR-21-2750
Greenall SA, McKenzie M, Seminova E et al (2019) Most clinical anti-EGFR antibodies do not neutralize both wtEGFR and EGFRvIII activation in glioma. Neuro Oncol. https://doi.org/10.1093/neuonc/noz073
Di Leva G, Garofalo M, Croce CM (2014) MicroRNAs in cancer. Annu Rev Pathol 9:287–314. https://doi.org/10.1146/annurev-pathol-012513-104715
Svoronos AA, Engelman DM, Slack FJ (2016) OncomiR or tumor suppressor? the duplicity of microRNAs in cancer. Cancer Res 76:3666–3670. https://doi.org/10.1158/0008-5472.CAN-16-0359
Rolle K (2015) miRNA Multiplayers in glioma. From bench to bedside. Acta Biochim Pol 62:353–365. https://doi.org/10.18388/abp.2015_1072
Chen L, Kang C (2015) miRNA interventions serve as “magic bullets” in the reversal of glioblastoma hallmarks. Oncotarget 6(1):38628–38642. https://doi.org/10.18632/oncotarget.5926
Ananta JS, Paulmurugan R, Massoud TF (2015) Nanoparticle-delivered antisense microRNA-21 enhances the effects of temozolomide on glioblastoma cells. Mol Pharm 12:4509–4517. https://doi.org/10.1021/acs.molpharmaceut.5b00694
Ananta JS, Paulmurugan R, Massoud TF (2016) Tailored nanoparticle codelivery of antimiR-21 and antimiR-10b augments glioblastoma cell kill by temozolomide: toward a “Personalized” anti-microRNA therapy. Mol Pharm 13:3164–3175. https://doi.org/10.1021/acs.molpharmaceut.6b00388
Kirstein A, Schmid TE, Combs SE (2020) The role of miRNA for the treatment of MGMT unmethylated glioblastoma multiforme. Cancers (Basel). https://doi.org/10.3390/cancers12051099
Gomez GG, Volinia S, Croce CM et al (2014) Suppression of microRNA-9 by mutant EGFR signaling upregulates FOXP1 to enhance glioblastoma tumorigenicity. Cancer Res 74:1429–1439. https://doi.org/10.1158/0008-5472.CAN-13-2117
Serna E, Lopez-Gines C, Monleon D et al (2014) Correlation between EGFR amplification and the expression of microRNA-200c in primary glioblastoma multiforme. PLoS ONE 9:e102927. https://doi.org/10.1371/journal.pone.0102927
Dambal S, Shah M, Mihelich B et al (2015) The microRNA-183 cluster: the family that plays together stays together. Nucleic Acids Res 43:7173–7188. https://doi.org/10.1093/nar/gkv703
Tang H, Bian Y, Tu C et al (2013) The miR-183/96/182 cluster regulates oxidative apoptosis and sensitizes cells to chemotherapy in gliomas. CCDT 13:221–231. https://doi.org/10.2174/1568009611313020010
Wang Z-Y, Xiong J, Zhang S-S et al (2016) Up-regulation of microRNA-183 promotes cell proliferation and invasion in glioma by directly targeting NEFL. Cell Mol Neurobiol 36:1303–1310. https://doi.org/10.1007/s10571-016-0328-5
Zhang L, Quan H, Wang S et al (2015) MiR-183 promotes growth of non-small cell lung cancer cells through FoxO1 inhibition. Tumour Biol 36:8121–8126. https://doi.org/10.1007/s13277-015-3550-8
Tang G, Liu D, Xiao G et al (2016) Transcriptional repression of FOXO1 by KLF4 contributes to glioma progression. Oncotarget 7:81757–81767. https://doi.org/10.18632/oncotarget.13184
Lau CJ, Koty Z, Nalbantoglu J (2009) Differential response of glioma cells to FOXO1-directed therapy. Cancer Res 69:5433–5440. https://doi.org/10.1158/0008-5472.CAN-08-4540
William D, Mokri P, Lamp N et al (2017) Amplification of the EGFR gene can be maintained and modulated by variation of EGF concentrations in in vitro models of glioblastoma multiforme. PLoS ONE 12:e0185208. https://doi.org/10.1371/journal.pone.0185208
William D, Mullins CS, Schneider B et al (2017) Optimized creation of glioblastoma patient derived xenografts for use in preclinical studies. J Transl Med 15:27. https://doi.org/10.1186/s12967-017-1128-5
Dijkstra JR, Opdam FJM, Boonyaratanakornkit J et al (2012) Implementation of formalin-fixed, paraffin-embedded cell line pellets as high-quality process controls in quality assessment programs for KRAS mutation analysis. J Mol Diagn 14:187–191. https://doi.org/10.1016/j.jmoldx.2012.01.002
Ye Z, Zhang Z, Wu L et al (2016) Upregulation of miR-183 expression and its clinical significance in human brain glioma. Neurol Sci 37:1341–1347. https://doi.org/10.1007/s10072-016-2599-5
Tanaka H, Sasayama T, Tanaka K et al (2013) MicroRNA-183 upregulates HIF-1α by targeting isocitrate dehydrogenase 2 (IDH2) in glioma cells. J Neurooncol 111:273–283. https://doi.org/10.1007/s11060-012-1027-9
Yan Z, Wang J, Wang C et al (2014) miR-96/HBP1/Wnt/β-catenin regulatory circuitry promotes glioma growth. FEBS Lett 588:3038–3046. https://doi.org/10.1016/j.febslet.2014.06.017
Li Z, Ma J, Liu L et al (2017) Endothelial-monocyte activating polypeptide II suppresses the In Vitro Glioblastoma-Induced Angiogenesis by Inducing Autophagy. Front Mol Neurosci 10:208. https://doi.org/10.3389/fnmol.2017.00208
Wang G, Wang S, Li C (2017) MiR-183 overexpression inhibits tumorigenesis and enhances DDP-induced cytotoxicity by targeting MTA1 in nasopharyngeal carcinoma. Tumour Biol. https://doi.org/10.1177/1010428317703825
Lin X, Zheng L, Song H et al (2017) Effects of microRNA-183 on epithelial-mesenchymal transition, proliferation, migration, invasion and apoptosis in human pancreatic cancer SW1900 cells by targeting MTA1. Exp Mol Pathol 102:522–532. https://doi.org/10.1016/j.yexmp.2017.05.009
Zhao H, Shen J, Hodges TR et al (2017) Serum microRNA profiling in patients with glioblastoma: a survival analysis. Mol Cancer 16:59. https://doi.org/10.1186/s12943-017-0628-5
Kouri FM, Ritner C, Stegh AH (2015) miRNA-182 and the regulation of the glioblastoma phenotype - toward miRNA-based precision therapeutics. Cell Cycle 14:3794–3800. https://doi.org/10.1080/15384101.2015.1093711
He Z, Wang Y, Huang G et al (2017) The lncRNA UCA1 interacts with miR-182 to modulate glioma proliferation and migration by targeting iASPP. Arch Biochem Biophys 623–624:1–8. https://doi.org/10.1016/j.abb.2017.01.013
Kouri FM, Hurley LA, Daniel WL et al (2015) miR-182 integrates apoptosis, growth, and differentiation programs in glioblastoma. Genes Dev 29:732–745. https://doi.org/10.1101/gad.257394.114
Feng YA, Liu TE, Wu Y (2015) microRNA-182 inhibits the proliferation and migration of glioma cells through the induction of neuritin expression. Oncol Lett 10:1197–1203. https://doi.org/10.3892/ol.2015.3365
Xiao Y, Zhang L, Song Z et al (2016) Potential diagnostic and prognostic value of plasma circulating microRNA-182 in human glioma. Med Sci Monit 22:855–862. https://doi.org/10.12659/MSM.897164
Jiang L, Mao P, Song L et al (2010) miR-182 as a prognostic marker for glioma progression and patient survival. Am J Pathol 177:29–38. https://doi.org/10.2353/ajpath.2010.090812
Tang H, Wang Z, Liu Q et al (2014) Disturbing miR-182 and -381 inhibits BRD7 transcription and glioma growth by directly targeting LRRC4. PLoS ONE 9:e84146. https://doi.org/10.1371/journal.pone.0084146
Song L, Liu L, Wu Z et al (2012) TGF-β induces miR-182 to sustain NF-κB activation in glioma subsets. J Clin Investig 122:3563–3578. https://doi.org/10.1172/JCI62339
Xue J, Zhou A, Wu Y et al (2016) miR-182-5p induced by STAT3 activation promotes glioma tumorigenesis. Cancer Res 76:4293–4304. https://doi.org/10.1158/0008-5472.CAN-15-3073
Zhao K, Wang Q, Wang Y et al (2017) EGFR/c-myc axis regulates TGFβ/Hippo/Notch pathway via epigenetic silencing miR-524 in gliomas. Cancer Lett 406:12–21. https://doi.org/10.1016/j.canlet.2017.07.022
Wee P, Wang Z (2017) Epidermal growth factor receptor cell proliferation signaling pathways. Cancers (Basel). https://doi.org/10.3390/cancers9050052
Acknowledgements
The authors want to thank the technicians of the Institute of Pathology for excellent assistance.
Funding
Open Access funding enabled and organized by Projekt DEAL. None.
Author information
Authors and Affiliations
Contributions
BS: study design, all molecular analyses, statistics, writing and editing the manuscript; DW: in vitro experiments, statistics, editing the manuscript; NL: IHC interpretation; AZ: IHC interpretation, statistics, editing the manuscript; CH: patient data/clinical background, editing the manuscript; CFC: in vitro data, editing the manuscript; AE: providing patient FFPE material, editing the manuscript, supervising.
Corresponding author
Ethics declarations
Conflict of interest
The authors have not disclosed any competing interests.
Ethical approval
Specimen collection was conducted in accordance with the ethics guidelines for the use of human material, approved by the Ethics Committee of the University of Rostock (Reference number: A 2009/34) and with informed written consent from all patients prior to surgery.
Consent to participate
All authors stated consent for participation.
Consent for Publication
For all authors the final version of the manuscript was available and consent for publication was stated.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
11010_2022_4435_MOESM1_ESM.jpg
Supplementary file1 (JPG 4760 kb) Fig.1 EGFR specific CISH analysis of the 24 glioblastoma tissues used for miR-screening. Red dots represent centromeres of chromosome 7, green dots are specific for EGFR. #1 - #13 show EGFR amplified tumors, #14 - #24 EGFR normal tumors. ×400 magnification
11010_2022_4435_MOESM2_ESM.xlsx
Supplementary file2 (XLSX 10 kb) Table 1 Patiens' data showing their gender, age at dignosis and EGFR amplification status
11010_2022_4435_MOESM3_ESM.xlsx
Supplementary file3 (XLSX 31 kb) Table 2 Results of Nanostring Human v3 miRNA assays showing fold changes and their p values of EGFR amplified vs EGFR normal glioblastomas
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visithttp://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Schneider, B., William, D., Lamp, N. et al. The miR-183/96/182 cluster is upregulated in glioblastoma carrying EGFR amplification. Mol Cell Biochem 477, 2297–2307 (2022). https://doi.org/10.1007/s11010-022-04435-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11010-022-04435-y