
Abstract
Context
Human presence and land-use activities influence habitat use and activity of species. It is crucial to study the ecological and anthropogenic determinants that drive these relationships.
Objectives
We investigated the effects of land-use change and human presence on the habitat use of seven mammalian species and on their activity patterns.
Methods
We conducted a camera-trapping survey in Ballari district, India. We deployed camera-traps at 1457 sampling grid cells across Protected Areas (PAs), Reserved Forests (RFs), and adjoining fringe mosaic farmlands (FMFLs). We assessed the influence of 19 covariates related to habitat features, landscape-level human presence, climate, and local-level human presence on species habitat use. We examined the diel activity patterns and activity levels of mammals within different land-use types.
Results
Four species exhibited a net negative response to increasing human presence, while three species exhibited positive responses. Respectively, proximity to farmlands, and settlements positively influenced habitat use by Panthera pardus, and Lepus nigricollis,. Increasing direct human presence negatively affected Sus scrofa habitat use. Large body-sized species exhibited higher diurnal activity in PA/RF compared to FMFL while smaller-sized (≤ 2.2 kg) species maintained similar activity patterns across land-use types. Generalist meso-predators exhibited more diurnal activity in RF/FMFL.
Conclusions
Ecological and life history traits were strong predictors of species responses to increasing human presence which favoured smaller and generalist species. These insights into the complex mechanisms through which human activities shape mammal communities are important landscape level perspectives on conservation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Land use change and increased human activities are the prominent drivers of ongoing biodiversity loss (Powers and Jetz 2019; Suraci et al. 2021). These forces have a strong impact on the distribution of mammalian species (Polaina et al. 2019; Ramesh and Downs 2015) and underlie the declining population size of many mammals (Baisero et al. 2020; Rondinini and Visconti 2015). The ability of wild mammals to coexist with human in human-dominated landscapes may be strongly influenced by a species, ecological and life history traits (Gascon et al. 1999; Samia et al. 2015; Suraci et al. 2021). The resulting differential responses of species to human disturbance have a filtering effect at the community level, favoring species that can adapt to modified landscapes (Bellón et al. 2022; Drouilly et al. 2018; Ehlers Smith et al. 2018; Penjor et al. 2021). Consequently, human disturbance reshapes mammal communities in a way that can be predicted from suites of species traits (Estes et al. 2011; Schmitz et al. 2018). Therefore, understanding the effects of human-induced modifications on wild mammals has become a key aspect in biodiversity conservation at many scales ranging from local to global.
Species survival and success in disturbed and modified landscapes are heavily influenced by their ecological and life history traits (Galán-Acedo et al. 2019; Parsons et al. 2018). These traits interact with multiple anthropogenic stressors changing species behavior and distribution (Doherty et al. 2021; Nickel et al. 2020). Specifically, human presence can induce fear responses that alter habitat use and suppress activity (Gaynor et al. 2018; Oberosler et al. 2017). Such changes are often influenced by body size, home range size and trophic position (Clinchy et al. 2016; Suraci et al. 2021). Species with large home ranges are most vulnerable to habitat loss due to land conversion (Crooks et al. 2017; Milda et al. 2023a; Ripple et al. 2014). Carnivores are particularly sensitive to human activity and habitat alteration (Di Minin et al. 2016; Milda et al. 2023a). In contrast, species that habituate to the disturbance of human activity and exploiting anthropogenic food subsidies can thrive in human modified areas (Athreya and Karanth 2016; Wang et al. 2015). Species with traits like wide habitat type tolerance, nocturnal activity, and higher dietary flexibility can more easily adapt to modified landscapes (Larson et al. 2015; Newsome and Van Eeden 2017; Šálek et al. 2015). Furthermore, prey species and meso-carnivores can benefit when human presence reduces local predation pressure (Crooks 2002; Ordenana et al. 2010). Thus, understanding how functional traits of wild mammals influence their response to human disturbance can provide insights for conservation planning.
The importance of conserving biodiversity by creating Protected Areas (PAs) has been recognized by authorities around the globe (Dudley 2008; Hockings 2003). In India, PAs are wildlife sanctuaries that fall into IUCN category IV and include a very high degree of protection from poaching, grazing, and Non-timber Forest Product (NTFP) collection (Badola 1999; Ghosh-Harihar et al. 2019). However, PAs only encompass a small proportion of the landscape, less than 5% of the total landmass (Ghosh-Harihar et al. 2019). Consequently, PAs are only efficient in safeguarding a small proportion of diversity, as majority of species persist outside PAs (Schmitt et al. 2009). The non-protected forested areas in India are recognized as Reserved Forests, Community Reserves, and unclassed forests (Macura et al. 2011). Reserved forests (RFs), owned by state governments, cover 13.2% of India’s landmass and often contribute to buffers and corridors for PAs (Ghosh-Harihar et al. 2019; Shahabuddin and Thadani 2018). RFs have weaker safeguards against wildlife harvesting and livestock grazing by local people compared to PAs (Milda et al. 2020). Natural resource extraction is allowed in many RFs, putting additional pressure on these landscapes (Forest Survey of India 2019). The degradation of wildlife habitats (PAs and RFs), together with the increasing expansion of agricultural land, has a strong negative impact on biodiversity. Many mammalian species thrive in farmlands and forest remnants outside of PAs and RFs (Schmitt et al. 2009), which we will refer to as Fringe Mosaic Farmlands (FMFLs) in this study. FMFLs represents the anthropogenic land-use mosaic of agricultural lands. Thus, FMFLs are prone to more human disturbances and has lower habitat heterogeneity than PAs or RFs. Assessing anthropogenic landscape variables related to land-use types within the multi-use landscape mosaic and their impact on species distribution and behaviour at different scales and time frames is critical for biodiversity conservation (Noon et al. 2012; Roland et al. 2013; McDonald et al. 2015).
The need for research investigating factors shaping diverse mammalian community assemblages in less studied parts of the world is great (Pardikar 2020; Rowan et al. 2020). The landscape of the Ballari district situated in the Deccan plateau of India is one of such less studied area which has undergone extensive land-use change during the last century. Specifically, mining activities within forested habitats including more traffic from heavy vehicles have increased (Intercultural Resources 2014). Encroachment into forests from agrarian activities by locals, including heavy overgrazing, and collection of Non-timber Forest Product (NTFP) collection, as well as poaching have increased (Behera et al. 2022; Meena 2013; Rudramuniyappa 1997). All these activities exert intense pressure on the remnant forest patches contributing to the decline of many mammalian species. These activities also increase the frequency of negative human-wildlife interactions in the form of livestock predation, human injury or death, wild animal mortality, and crop damage (Martins-Oliveira et al. 2021; Meena 2013; Sharp et al. 2020). Resolving increasing anthropogenic pressure and the associated decline of mammalian species in such disturbed ecosystems requires broad landscape-level studies. Such studies can help discern the primary determinants of species extinction at the local level and identify solutions to help forest managers ensure effective wildlife conservation (Jones 2011). We address a critical knowledge gap regarding the influence of anthropogenic and ecological factors on specific species in our study system to understand forces shaping the mammalian assemblage.
We examined the effect of anthropogenic disturbances on seven mammal species found within Ballari district using landscape-scale variables from four broad categories (habitat features, landscape-level human presence, climate, and local-level human presence). We report habitat use of black napped hare (Lepus nigricollis), golden jackal (Canis aureus), Indian fox (Vulpes bengalensis), jungle cat (Felis chaus), leopard (Panthera pardus), sloth bear (Melursus ursinus) and wild pig (Sus scrofa). This group of species includes varied functional traits like body sizes, diets, trophic positions, and home range sizes. All of these species are native to the study site and details about their functional traits (Table 1) and IUCN red list statuses are provided in Supplementary Table S1. Our main objective was to understand how the combination of a species’ functional traits influences its response to human activities. Specifically, we predicted that species with; larger body sizes, larger home range sizes, specialized diets and top carnivores, will be negatively affected by variables associated with landscape-level or local level human presence (Ripple et al. 2014). We predicted that habitat use, and activity of small and generalist carnivores will respond positively to increased human presence and availability of farmlands (Katna et al. 2022) and that the hare habitat use will be positively associated with PA. We predicted that habitat use by large mammals would be positively associated with natural habitat with dense canopy coverage (Andrade-Núñez and Aide 2010). We also compared activity patterns and activity levels between the study species across three land-use types. We predicted that mammalian species sensitive to human activities would exhibit highest nocturnal activity in FMFL, followed by RFs and PAs. We also predicted an increase in the concentration of activity i.e., a shorter activity window in disturbed land use types (FMFL and RF).
Methods
Study area
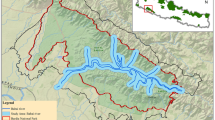
We conducted our study within the jurisdictional area of the Ballari territorial forest division that forms a part of the Deccan Peninsula of India (between 14° 30′ and 15° 50′ north latitude and 75° 40′ and 77° 11′ east longitude; Fig. 1). The region lies between 550 and 750 m AMSL with the terrain varying from open plains to undulating areas with some patches of rugged hills (Meena 2013). The average annual temperature varies from 20 to 40 °C. Our study area consists of a mostly dry landscape with an average annual rainfall of around 574.9 mm. The central part of the district contains tropical dry deciduous forests predominately composed of species like Albizzia amara, Chloroxylon swietenia, Hardwickia binata, Grewia tiliaefolia, and Azadirachta indica). The other forested patches represent dry thorn forests with sparse vegetation (Champion and Seth 1968), that are dominated by Albizzia amara, Acacia catechu, Cassia auriculata, Euphorbia spp., and Zizyphus spp.).

Camera trap locations of the survey area in Ballari district, Karnataka, across Protected Areas (PAs; n = 251), Reserved Forests (RFs; n = 756) and the adjoining fringe mosaic farmlands (FMFLs; n = 450). Camera traps located in farmlands i.e., outside forested landscape (PAs/RFs; within 1 km from forest boundaries) are the traps located in FMFLs.
Ballari is one of the most economically vulnerable districts in the country and its economy is primarily based on mining and agrarian activities. The local people maintain large cattle populations for sustenance/economic prosperity and are highly dependent on NTFP collection. The district contains rich deposits of minerals of economic importance like iron and manganese (Meena 2013). More than 5000 ha of land within the study area is actively mined, with most of the mines situated within RFs.
Data collection and analyses
Camera trapping data were collected within the forested areas of Ballari district including PAs [Daroji (est. 1994) and Gudekote (est. 2013) Sloth bear sanctuaries], RFs, and FMFL (a 1 km buffer surrounding forest boundaries). We established a camera trapping grid composed of 1 km2 cells as sampling units by overlaying a 1 × 1 km2 grid across the study areas using ArcGIS 10.1 (ESRI, Redlands, CA, USA). We used a land cover layer that we obtained from Karnataka Forest Department to identify the land cover types in the study area.
We conducted camera-trapping surveys to to collect occurrence data during a period of 21 months between January 2019 and October 2021. Within each 1 km2 grid cell, we walked for at least 1 km around the centroid of the grid cell and identifed the best potential camera sites based on mammalian sign evidences (sightings, scats/pellets, and tracks of mammals) (Jhala et al. 2008; Kalle et al. 2013; Karanth and Nichols 2002; Ramesh et al. 2012). We installed a single passive infrared camera trap (Cuddeback digital 20MP Blue Series, USA) at each sample site within each grid cell. Camera traps were fitted 30 cm above the ground level and placed along dirt roads, animal trails, rivers, and stream beds, near water holes, etc. that are commonly used by study species. We maintained a minimum distance of approximately 500 m between any two cameras to insure spatial independence of our samples. In total, we sampled 1457 grid cells including 251 grid cells within PAs, 756 grid cells within RFs, and 450 grid cells within FMFL. The number of sampling sites per land-use type was proportional to to the occurrence of those land-use types within in the study area. We deployed 142–145 camera traps simultaneously and shifted those cameras to other grid cells throughout the study period. We set camera traps (30s motion triggered delay setting) to run continuously for 22 days within each grid cell and collected data weekly. In each camera trap location, we visually estimated the percentage of canopy coverage, tree density, percentage of rockiness, and shrub coverage within a circular plot with a radius of 20 m (Table 2). After retrieving photos from the camera traps, we measured local-level human presence at each site using four proxies: relative abundance index (RAI) of human (on foot or in vehicles), cattle, goats-sheep, and domestic dogs. We calculated RAI of a species as the total number of independent (one photograph with 30 min intervals) photographs of that species divided by the total number of camera trap-nights at that location (Palmer et al. 2018; O’Brien et al. 2003). We estimated proximity variables like proximity to road, settlements, farmlands, etc. using verified spatial layers like GRIP global roads database, and Roy et al. 2015 (after converting these rasters into shapefiles) and the Euclidean distance tool in ArcMap 10.3 (ArcMap 10.3, Software. Redlands, CA: Esri Inc, 2016). Details for the covariates are provided in Table 2.
Habitat use and detection probability
We considered 21 predictor variables estimated from camera sites to be important to the study species. We categorised these variables into- (1) Habitat features, comprising naturally occurring features or habitat characteristics; (2) Landscape-level human presence, comprising features associated with varying degrees of human footprint and landscape management; (3) Climate, comprising two climatic features and (4) Local-level human presence, comprising the RAI of humans, dogs, and livestock at camera stations (Table-2).
Considering the average home range size of the study species [hare: 0.01–0.02 km2 (Kirk and Bathe 1994), jackal: 3.1–23.7 km2 (Katna et al. 2022), fox: 1.16–10.71 km2 (Katna et al. 2022), jungle cat: 1.01–26.48 km2 (Katna et al. 2022), leopard: 8–65 km2 (Odden et al. 2014), bear: 12–85 km2 (Yoganand et al. 2005), and wild pig: 0.62–48.3 km2 (Garza et al. 2018), we interpreted occupancy estimates as habitat use because the home ranges of some of our study species are larger than the grid cell size used. This interpretation of our results as probability of habitat use, and not probability of occupancy at each sampling unit is done to in part minimize the impact of our likely violating the assumption of closure (Burton et al. 2015). We treated each camera site as an independent sample and sampling occasion as a temporal repeat of the survey. In our analyses, we assumed the habitat use of a species to be independent of other species. We developed ‘0’-‘1’matrices for each species. Rows represented sample locations and columns daily sampling occasions. Detection and non-detection during successive sampling occasion were respectively indicated with ‘1’ and ‘0’ in this matrixd (Otis et al. 1978). We used a single-season occupancy model to estimate site occupancy/habitat use (Ψ) and detection probability (p) of study species (MacKenzie et al. 2017) using the package “unmarked” (Fiske and Chandler 2011) in program R (R Core Team 2020). Prior to modeling, we standardized all continuous site covariates to z-scores which allows model coefficients to be interpreted as the change in the log-odds ratio of habitat use relative to a 1-standard deviation change in a covariate from its mean (Cooch and White 2005). To avoid multi-collinearity problems, we tested the correlations among independent variables (Graham 2003) using Pearson’s correlation coefficient (Supplementary Figure S1) using the package “corrplot” in program R and didn’t use highly correlated variables (r > 0.60) in the same model (Wei and Simko 2021). We removed a few highly correlated independent variables (decidist: proximity to deciduous forests and FMFL: fringe mosaic farmlands) before modelling. Details on the selection of relevant variables to estimate habitat use and detection probability in our models are provided on Table 2. We followed stepwise model selection procedures and the goodness of fit for model selection as described in Burnham and Anderson (2002) and ranked the models using AIC (Akaike information criterion). We considered the models with the lowest AIC values (ΔAIC ≤ 2) as the best descriptors of species habitat use and detection probability among candidate models (Burnham and Anderson 2002).
Activity patterns and activity levels
We investigated the diel activity patterns for the seven study species across three land-use types. The activity patterns of a species are the temporal data that provide information on the diel trends of its actions (Gerber et al. 2012). Studying activity patterns provides insight into the ecological processes that shape the space use by an animal community (Houngbégnon et al. 2020). We subsampled the detection data considering consecutive detections within a 30 min interval as a single event (Meek et al. 2014; Zimmermann et al. 2016). We segregated all independent events of a species into three categories for activity pattern analysis based on land use type (Table 1). Then we performed pairwise comparisons of activity patterns of species between all three land use types (PA vs. RF, RF vs. FMFL, and FMFL vs. PA) by estimating the overlap coefficient Δ (ranging from 0 to 1 i.e., no overlap to complete overlap) and its confidence interval (Ridout and Linkie 2009). We performed statistical analyses and produced Kernel density estimation curves using the ‘Overlap’ package (Meredith and Ridout 2016) in Program R 4.0.5 (R Core Team 2020). Kernel density estimation is a non-parametric method for estimating the probability density function of a distribution of records (Linkie and Ridout 2011). Activity level of a species is the proportion of time that species spend being active during an entire day (Rowcliffe et al. 2014). This method assumes that all individuals in the sampled population are active at the peak of the daily activity cycle. We used the function compareAct() in the package “Activity” Program R 4.0.5 (R Core Team 2020) to conduct Wald test for the statistical difference between two or more activity level estimates. We quantified the activity level estimates of study species in the different land-use types.
Results
Camera trapping survey at 1457 sampling sites, n = 31,702 trap-nights yielded photographs of 24 wild mammalian species, livestock, dogs, and humans (Supplementary Table S1). We conducted occupancy analysis for seven wild mammalian species that are widely distributed across our study site and, for which we had sufficient data for occupancy analysis (MacKenzie et al. 2017). We didn’t include some species for analysis as they occurred in low densities (Neilson et al. 2018). Hare had the highest naïve occupancy 0.528, followed by wild pig = 0.318, jungle cat = 0.217, jackal = 0.203, fox = 0.189, leopard = 0.173, and bear = 0.15. We determined the best-fit models for each species and their log-transformed parameter estimates of habitat use Ψ and detection probabilities p with detailed results listed in Table 3; Fig. 3, respectively. The summary of AIC model selection and parameter estimates of habitat use and detection for study species are listed in supplementary tables S2-S8. The number of variables influencing patterns of habitat use and detection probability in well-supported models (∆AIC ≤ 2), ranged from 4 to 8 across the study species (Table 3). Estimates of habitat use Ψ with standard errors varied from 0.239 ± 0.023 to 0.538 ± 0.028 across study species and models (Fig. 2). Hares had the highest estimates for habitat use. Wild pigs had the second highest habitat use after hares (Fig. 2). Jackals, foxes, and sloth bears had lower habitat use compared to other study species (Fig. 2). Hares had the highest detection probability, followed by jackals and foxes (Fig. 2). The residuals of the top models for jackal, fox, jungle cat, and hare did not exhibit any spatial autocorrelation [Moran’s I for jackal = 0.0006 (P = 0.38); fox = − 0.0026 (P = 0.82); jungle cat = 0.0009 (P = 0.17); and hare = − 0.0007 (P = 0.84)]. However, top model residuals for leopard, bear, and wild pig documented some spatial autocorrelation [Moran’s I for leopard = 0.0066 (P = 0.00); bear = 0.0108 (P = 0.00); and pig = 0.0022 (P = 0.03)].

Habitat use and detection probabilities of study species present in Ballari district, India
We found that variables related to human presence whether at landscape level or local level, were associated with study species habitat use. Proximity measures like distance to human settlement and agricultural farmlands were positively associated with the habitat use of hares and leopards, respectively. Distance to major water bodies was positively associated with the habitat use of leopards. Variables associated with local-level human presence were negatively associated with wild pig habitat use while they were positively associated with habitat use by jackals and jungle cats. Habitat use of bears and hares was highest in PAs while the habitat use of jackals and foxes was highest in RFs. Unexpectedly, relative cattle abundance emerged as an important predictor for increased habitat use of bears. Canopy coverage and mean annual precipitation were the two common predictors across top models for three and five species, respectively (Fig. 3a and b). The dense canopy coverage positively influenced the habitat use of leopards, bears, and wild pigs. Mean annual precipitation positively influenced the habitat use of hares, leopards, and wild pigs while it had a negative impact on the habitat use of jackals and foxes. The availability of plains positively influenced the habitat use of hares and jungle cats. The most common covariate for detection probability was proximity to the farmlands. Higher detection probabilities for jackals and jungle cats were associated to close proximity to farmlands while this significant trend was reversed for hares.

Log-transformed parameter estimates for explanatory variables from the top (a) habitat use and (b) detection probability model for mammalian species in Ballari, Karnataka, India
Activity patterns of species varied across the three land use types (PA vs. RF, RF vs. FMFL, and FMFL vs. PA) (Fig. 4). Activity patterns of all species showed bimodal peaks with first peaks occurred between late evening and midnight while second peaks occurred during early mornings. Hares and jungle cats exhibited similar activity patterns across three land use types. Activity patterns of hares in RF and FMFL were very similar to each other with the mean kernel temporal overlap coefficient estimate of 0.94 (Table 4). Jackals and foxes had higher diurnal activities in RF and FMFL than in PA (Fig. 4; Table 4). While leopards, sloth bears, and wild pigs had higher diurnal activity patterns in wildlife habitats like PA/RF than in FMFL (Fig. 4; Table 4). Activity level estimates of all study species varied across land use types. The activity level of sloth bears and hares in PAs and RFs were significantly higher than in FMFL (Table 5). Leopards, jackals, wild pigs, and hares had significantly higher activity levels in PAs/RFs, relative to FMFL (Table 5). Foxes had significantly higher activity levels in RFs, relative to PAs/FMFLs while there was no significant difference in activity level estimates for jungle cats across land-use types.

Comparison of Kernal density activity patterns of focal species in different land use types [Protected Areas (PAs) vs. Reserved Forests (RFs) vs. adjoining fringe mosaic farmlands (FMFL) in Ballari, Karnataka, India
Discussion
Intensification of human disturbance has driven local extinction for most mammals during recent centuries (Wan et al. 2019). Landscape-level studies describing the effects of human disturbance on the distribution of large mammals are essential to understand how to protect their populations from further extinction. Our investigation of habitat uses and activity of seven mammalian species provides a thorough understanding of mammalian responses to anthropogenic disturbances in a highly fragmented landscape. We found that all species’ habitat use, and activity patterns are not affected equally by human presence and land use activities. Our landscape-level spatial analysis results confirmed that accelerated anthropogenic activities have more detrimental effect on species with larger body size, larger home range size, higher trophic level, specialist diet, and less nocturnal habit. The results of our analysis of temporal trends of larger mammals showed adjustment in diel activity pattern towards more nocturnality is strongly influenced by increasing human activity and reduction of habitat heterogeneity across land-use types. Some of our study species were highly sensitive to land conversion and reduction of natural habitat while others showed varying tolerance to human-modified habitats. Our findings are consistent with previous studies demonstrating that pristine natural habitats like Protected Areas are the most favourable habitats for large mammals (Pacifici et al. 2020). Thus, our study provided an informative perspective on the decline of several species by identifying the important land use changes that altered their habitat use and behavior within our poorly understudy study area.
In landscapes that are prone to heavy anthropogenic disturbances, it’s important to identify how land use changes impact different species (Ramesh and Downs 2015; Sodhi et al. 2010). Ballari district is a landscape that experiences heavy human pressure in form of mining activities, livestock over-grazing, and illegal wildlife harvesting. Our analysis revealed that each study species was distributed throughout the entire study area, but they used land types differently. The difference in the distribution of these species indicates varying levels of tolerance to human disturbances. The two the most widely distributed species within this landscape were hare and wild pig which may be important as these are two major prey species in this system. Leopards, sloth bears, wild pigs, and hares preferred intact undisturbed forests (Ripple et al. 2014). While meso-predators like jackals, foxes, and jungle cats used more modified landscapes, this is consistent with many prior studies on meso-predators (Beasley et al. 2011; Torre et al. 2022; Zamuda et al. 2022).
Species occurrence can be strongly influenced by the quantity and quality of available habitat (Ceballos and Brown 1995; Ramesh and Downs 2015; White et al. 1997) and our results supported these findings. Agricultural lands negatively influenced the leopards’ habitat use suggesting the importance of natural forested habitats for this species as shown by Gubbi et al. (2020). On the other hand, the abundance of small prey species for meso-carnivores like jackals, foxes and jungle cats is higher in agricultural lands than forest habitats (Alain et al. 2006). High food availability, protection against large predators, and their ability to use human resources undoubtedly all contribute to increased habitat use and detection of these meso-predators within RF and FMFL (Carricondo-Sanchez et al. 2019; Šálek et al. 2014; Vanak and Gompper 2010). The preference of hares for PAs within our study area may be associated with restricted poaching activities within this land use because hares are heavily poached across our study area (Behera et al. 2022). The positive association of hare habitat use with proximity to human settlement supports this hypothesis. Our finding that leopard habitat use was positively associated with proximity to water is likely specific to our study area. Ballari district is a dry landscape and all the major water sources are either situated near human settlement or associated with higher human presence (Meena 2013). Habitat use of sloth bears was higher in PA. Thus, our results that two existing sloth bear sanctuaries in the district were placed in locations that are highly valuable for bear conservation. Relative abundance of humans was positively associated with the habitat use of jungle cats which could be due to their strong dependence on the availability of small prey like rodents that are mostly found in human-modified landscapes (Mukherjee et al. 2004). The habitat use of foxes was negatively associated with proximity to scrub forest and landscape rockiness. Higher habitat use of foxes and jackals in RFs relative to PAs maybe be explained by two factors. First, the moderately disturbed landscape of RFs might provide additional food resources (Jaegar et al. 2007) as well as reducing the frequency of encounters with large predators. Second, the RFs in our study area are associated with the plain landscapes and that feature could provide suitable sites for denning (Carricondo-Sanchez et al. 2019; Kumara and Singh 2012). Wild pigs are heavily poached throughout our study site (Behera et al. 2022) which likely explains their negative response to increased RAI human. The habitat use of two large mammals in our study i.e. leopard, and sloth bear, were positively influenced by the canopy coverage, which concurs with the findings of prior studies (Ngoprasert et al. 2007; Whitworth et al. 2019). According to Gould et al. (2001), high termite densities are associated with cattle dung patches, which explains our identification of RAI cattle as a strong positive predicting factor for sloth bear habitat use.
We found the mean annual precipitation was one of the important predictor variables for habitat use by five mammals. Precipitation positively influenced the habitat use of leopards, wild pigs, and hares but it was negatively associated with the habitat use of jackals and foxes. This affirms strong relationship of precipitation with the distribution of these mammals in dry landscapes (Ogutu and Owen-Smith 2005; Yusefi et al. 2021). With increased human activities, more species inhabit modified landscapes consisting of a mosaic of both natural and anthropic land covers. The ability of species to adapt and use anthropic land covers is a key determinant of their persistence in human-modified landscapes (Galán-Acedo et al. 2019).
Intensification of human disturbances causes wild mammals to avoid direct confrontation with humans to survive (Ditchkoff et al. 2006). Human disturbances and presence have negative effects on many mammals causing them to avoid disturbed landscapes, suppress their activity levels, or alter their activity patterns (Ikeda et al. 2022; Oriol-Cotterill et al. 2015; Suraci et al. 2021). With human activities being primarily diurnal in our study area, all species showed nocturnal activity patterns. We found that species with larger body sizes like leopards, bears, and wild pigs increased nocturnal activity pattern in more disturbed land-use type like FMFL relative to PAs/RFs, which is consistent with earlier studies (Carter et al. 2015; Johann et al. 2020; Ohashi et al. 2013; Ramesh et al. 2013; Van Cleave et al. 2018;). Meso-predators like jackals and foxes increased diurnal activity in RF or FMFL relative to PA. Sunquist and Sunquist (2002) found that jungle cats are typically diurnal. However, in our study jungle cat exhibited nocturnal activity patterns in all land use types. This difference in our data maybe a response to the high intensity of human disturbance across our study area. Activity patterns of jackals, foxes, and jungle cats across land-use types may relate to their tolerance of human presence. Hare had similar activity patterns across land-use types, but we observed a spike in their activity patterns in RFs and FMFL. Hares’ activity pattern varies from crepuscular to nocturnal, and their increased activity in the twilight period suggests an increased concentration of activity in human modified landscapes. We also observed a shorter window in their activity patterns within human-dominated landscapes (Shamoon et al. 2018). Estimates of activity level of four out of seven of our study species showed higher activity in PAs or RFs than in FMFL suggesting wild mammalian species prefer spending more time in less-disturbed landscape. Thus, our mammalian activity-related results supported our prediction that mammalian species would show increased nocturnal activity patterns in FMFL relative to PAs/RFs and increased concentration of activity i.e., a shorter activity of window with increasing habitat disturbance.
Our study provided novel insights into the complex relationship between anthropogenic disturbance and mammalian assemblage by quantifying the filtering effect of human land use change on seven mammal species with diverse traits (Belote et al. 2020; Hill et al. 2020). Our consideration of several land use variables provides quantitative insights to inform strategic decision-making in the conservation of mammals. Even though our analyses focused on the Deccan plateau of India, we strongly advocate that the patterns observed are likely applicable to medium-to-large mammal species globally. For example, a shift in mammal community composition towards smaller and generalist species with reduced space requirements can be a consequence of the increasing the human footprint (Suraci et al. 2021). The structure and diversity of mammal communities are the key determinants of ecosystem processes (Estes et al. 2011; Schmitz et al. 2018). However, more studies need to investigate the links between functional traits and habitat use of species in different land use types. Similar studies which also incorporate inter-specific interaction along environmental gradients will provide more robust information about ecosystem function. Conservationists and wildlife managers are typically focused on the protection of wildlife species in PAs. Given that the majority of species reside outside PAs, our results highlight the shortcomings of the forest PA network. Despite being detrimental to many species, semi-disturbed non-protected areas like RFs, provide opportunities for some species to prosper while also connecting PAs (Berger 2007). Expanding PAs will certainly help the populations of large mammals (Milda et al. 2023), but for species that thrive outside PAs, we recommend new studies determining their habitat requirements to provide effective refuge areas for conservation (Li et al. 2021). Conservation objectives require approaches for managing landscapes holistically by incorporating anthropogenic habitats like RFs, and farmlands with PA networks.
Ongoing landscape modification and increasing human use of remaining natural areas make it important to identify species that will thrive as well as those that will perish under anthropogenic pressure. Such inference is critical to conserving mammal communities and to maintaining mammalian diversity and associated ecological services. Natural habitats in a mosaic of anthropic-modified habitats can reduce negative effects on wildlife (Ehlers Smith et al. 2018; Galán-Acedo et al. 2019). Therefore, forest patch protection should be encouraged through continued reforestation and restoration, which will benefit a diverse range of mammalian species. Regulation of mining activities and mining-vehicular movements inside forests along with a strict enforcement of rules related to illegal resource extraction such as overgrazing and poaching activities could further reduce adverse effects on wildlife. So, we advocate for new studies that analyze the effects of mining related activities, overgrazing, and poaching on wildlife conservation in Ballari district We urge conservation agencies to increase awareness by organizing training programs for local communities. Such programs should improve understanding of human-wildlife interactions while providing suitable incentives and compensation to people who suffer wildlife damage. Our findings illustrate the need for conservationists, forest managers, and local people to work together to create landscape-level planning that integrates local livelihoods and wildlife conservation.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Alain B, Gilles P, Yannick D (2006) Factors driving small rodents assemblages from field boundaries in agricultural landscapes of western France. Landsc Ecol 21:449–461
Andrade-Núñez MJ, Aide TM (2010) Effects of habitat and landscape characteristics on medium and large mammal species richness and composition in northern Uruguay. Zoologia (Curitiba) 27:909–917
Athreya V, Odden M, Linnell JD, Krishnaswamy J, Karanth KU (2016) A cat among the dogs: leopard Panthera pardus diet in a human-dominated landscape in western Maharashtra, India. Oryx 50:156–162
Badola R (1999) People and protected areas in India. UNASYLVA-FAO-, 12–14
Baisero D, Visconti P, Pacifici M, Cimatti M, Rondinini C (2020) Projected global loss of mammal habitat due to land-use and climate change. One Earth 2:578–585
Beasley JC, Olson ZH, Dharmarajan G, Eagan TS, Rhodes OE (2011) Spatio-temporal variation in the demographic attributes of a generalist mesopredator. Landsc Ecol 26:937–950
Behera AK, Kumar PR, Priya MM, Ramesh T, Kalle R (2022) The impacts of COVID-19 lockdown on wildlife in Deccan Plateau, India. Sci Total Environ 822:153268
Bellón B, Henry DA, Renaud PC, Roque FDO, Santos CC et al (2022) Landscape drivers of mammal habitat use and richness in a protected area and its surrounding agricultural lands. Agric Ecosyst Environ 334:107989
Belote RT, Faurby S, Brennan A, Carter NH, Dietz MS et al (2020) Mammal species composition reveals new insights into Earth’s remaining wilderness. Front Ecol Environ 18:376–383
Berger J (2007) Fear, human shields and the redistribution of prey and predators in protected areas. Biol Lett 3:620–623
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a theoretic approach, 2nd edn. Springer, New York
Burton AC, Neilson E, Moreira D, Ladle A, Steenweg R et al (2015) Wildlife camera trapping: a review and recommendations for linking surveys to ecological processes. J Appl Ecol 52:675–685
Carricondo-Sanchez D, Odden M, Kulkarni A, Vanak AT (2019) Scale‐dependent strategies for coexistence of mesocarnivores in human‐dominated landscapes. Biotropica 51:781–791
Carter N, Jasny M, Gurung B, Liu J (2015) Impacts of people and tigers on leopard spatiotemporal activity patterns in a global biodiversity hotspot. Glob Ecol Conserv 3:149–162
Ceballos G, Brown JH (1995) Global patterns of mammalian diversity, endemism, and endangerment. Conserv Biol 9:559–568
Champion HG, Seth SK (1968) A revised survey of the forest types of India. Manager of Publications
Clinchy M, Zanette LY, Roberts D, Suraci JP, Buesching CD, Newman C, Macdonald DW (2016) Fear of the human super predator far exceeds the fear of large carnivores in a model mesocarnivore. Behav Ecol 27:1826–1832
Cooch E, White GM (2005) Program Mark: A Gentle Introduction. http://www.phidot.org/software/mark/docs/book
Crooks KR (2002) Relative sensitivities of mammalian carnivores to habitat fragmentation. Conserv Biol 16:488–502
Crooks KR, Burdett CL, Theobald DM, King SR, Di Marco M, Rondinini C, Boitani L (2017) Quantification of habitat fragmentation reveals extinction risk in terrestrial mammals. Proc Natl Acad Sci 114:7635–7640
Di Minin E, Slotow R, Hunter LT, Montesino Pouzols F, Toivonen T et al (2016) Global priorities for national Carnivore conservation under land use change. Sci Rep 6:1–9
Ditchkoff SS, Saalfeld ST, Gibson CJ (2006) Animal behavior in urban ecosystems: modifications due to human-induced stress. Urban Ecosyst 9:5–12
Doherty TS, Hays GC, Driscoll DA (2021) Human disturbance causes widespread disruption of animal movement. Nat Ecol Evol 5:513–519
Drouilly M, Clark A, O’Riain MJ (2018) Multi-species occupancy modelling of mammal and ground bird communities in rangeland in the Karoo: a case for dryland systems globally. Biol Conserv 224:16–25
Dudley N (2008) Guidelines for applying protected area management categories. IUCN, Gland
Ehlers Smith YC, Ehlers Smith DA, Ramesh T, Downs CT (2018) Forest habitats in a mixed urban-agriculture mosaic landscape: patterns of mammal occupancy. Landsc Ecol 33:59–76
Ekka P, Patra S, Upreti M, Kumar G, Kumar A, Saikia P (2023) Land Degradation and its impacts on Biodiversity and Ecosystem services. Land and Environmental Management through Forestry, pp 77–101
Estes JA, Terborgh J, Brashares JS, Power ME, Berger J et al (2011) Trophic downgrading of planet earth. Science 333:301–306
Fiske I, Chandler R (2011) Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J Stat Softw 43:1–23
Forest Survey of India (2019) State of Forest Report. Ministry of Environment and Forests, Government of India, Dehradun, India
Galán-Acedo C, Arroyo-Rodríguez V, Andresen E, Verde Arregoitia L, Vega E, Peres CA, Ewers RM (2019) The conservation value of human-modified landscapes for the world’s primates. Nat Commun 10:1–8
Garza SJ, Tabak MA, Miller RS, Farnsworth ML, Burdett CL (2018) Abiotic and biotic influences on home-range size of wild pigs (Sus scrofa). J Mammal 99:97–107
Gascon C, Lovejoy TE, Bierregaard RO Jr, Malcolm JR, Stouffer PC et al (1999) Matrix habitat and species richness in tropical forest remnants. Biol Conserv 91:223–229
Gaynor KM, Hojnowski CE, Carter NH, Brashares JS (2018) The influence of human disturbance on wildlife nocturnality. Science 360:1232–1235
Gerber BD, Karpanty SM, Randrianantenaina J (2012) Activity patterns of carnivores in the rain forests of Madagascar: implications for species coexistence. J Mammal 93:667–676
Ghosh-Harihar M, An R, Athreya R, Borthakur U, Chanchani P et al (2019) Protected areas and biodiversity conservation in India. Biol Conserv 237:114–124
Gould KA, Herrick JE, Lezama H (2001) Refuse to Refuge: Dry Season Use and Modification of Cattle Dung by Subterranean Termites in Guanacaste, Costa Rica 1. Biotropica 33:121–130
Graham MH (2003) Confronting multicollinearity in ecological multiple regression. Ecology 84:2809–2815
Gubbi S, Sharma K, Kumara V (2020) Every hill has its leopard: patterns of space use by leopards (Panthera pardus) in a mixed-use landscape in India. PeerJ 8:e10072
Hill JE, DeVault TL, Wang G, Belant JL (2020) Anthropogenic mortality in mammals increases with the human footprint. Front Ecol Environ 18:13–18
Hockings M (2003) Systems for assessing the effectiveness of management in protected areas. Bioscience 53:823–832
Houngbégnon FG, Cornelis D, Vermeulen C, Sonké B, Ntie S et al (2020) Daily activity patterns and co-occurrence of duikers revealed by an intensive camera trap survey across Central African rainforests. Animals 10:2200
Ikeda T, Higashide D, Shichijo T (2022) Impact of human disturbance in Japan on the distribution and diel activity pattern of terrestrial mammals. J Nat Conserv 70:126293
Intercultural Resources (2014) The Horrors of Bellary. Climate change, corporate accountability, indigenous struggles of land, mining scams and urban displacement. https://www.ritimo.org/The-Horrors-of-Bellary (Accessed 22 September 2022)
Jaeger MM, Haque E, Sultana P, Bruggers RL (2007) Daytime cover, diet and space-use of golden jackals (Canis aureus) in agro-ecosystems of Bangladesh. Mammalia 71:1–10
Jhala YV, Gopal R, Qureshi Q (2008) Status of tigers, co-predators, and prey in India by National Tiger Conservation Authority and Wildlife Institute of India. TR08/001, p. 164
Johann F, Handschuh M, Linderoth P, Dormann CF, Arnold J (2020) Adaptation of wild boar (Sus scrofa) activity in a human-dominated landscape. BMC Ecol 20:1–14
Jones JP (2011) Monitoring species abundance and distribution at the landscape scale. J Appl Ecol 48:9–13
Kalle R, Ramesh T, Qureshi Q, Sankar K (2013) Predicting the distribution pattern of small carnivores in response to environmental factors in the western ghats. PLoS ONE 8:e79295
Karanth KU, Nichols JD (2002) Monitoring Tigers and their prey: a manual for researchers, managers and conservationists in Tropical Asia. Centre for Wildlife Studies, Bangalore, India, pp 139–152
Katna A, Kulkarni A, Thaker M, Vanak AT (2022) Habitat specificity drives differences in space-use patterns of multiple mesocarnivores in an agroecosystem. J Zool 316:92–103
Kirk DA, Bathe GM (1994) Population size and home range of black-naped hares Lepus nigricollis nigricollis on Cousin Island (Seychelles, Indian Ocean)
Kumara HN, Singh M (2012) Distribution, den characteristics and diet of the Indian Fox Vulpes bengalensis (Mammalia: Canidae) in Karnataka, India: preliminary observations. J Threat Taxa 4:3349–3354
Larson RN, Morin DJ, Wierzbowska IA, Crooks KR (2015) Food habits of coyotes, gray foxes, and bobcats in a coastal southern California urban landscape. West N Am Nat 75:339–347
Li W, Yang P, Li B, Liu C, Sun L et al (2021) Habitat characteristics or protected area size: what is more important for the composition and diversity of mammals in nonprotected areas? Ecol Evol 11:7250–7263
Linkie M, Ridout MS (2011) Assessing tiger–prey interactions in Sumatran rainforests. J Zool 284:224–229
MacKenzie DI, Nichols JD, Royle JA, Pollock KH, Bailey L, Hines JE (2017) Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Elsevier
Macura B, Zorondo-Rodríguez F, Grau-Satorras M, Demps K, Laval M et al (2011) Local community attitudes toward forests outside protected areas in India. Impact of legal awareness, trust, and participation. Ecol Soc 16
Martins-Oliveira AT, Zanin M, Canale GR, da Costa CA, Eisenlohr PV et al (2021) A global review of the threats of mining on mid-sized and large mammals. J Nat Conserv 62:126025
McDonald PJ, Griffiths AD, Nano CE, Dickman CR, Ward SJ, Luck GW (2015) Landscape-scale factors determine occupancy of the critically endangered central rock-rat in arid Australia: the utility of camera trapping. Biol Conserv 191:93–100
Meek PD, Ballard G, Claridge A, Kays R, Moseby K et al (2014) Recommended guiding principles for reporting on camera trapping research. Biodivers Conserv 23:2321–2343
Meena VL (2013) Working Plan of Ballari Forest Division. Government of Karnataka, Forest Department, Karnataka. 2013-14 to 2022-23
Menon V (2023) Indian mammals: a field guide. Hachette India
Meredith M, Ridout M (2016) Overlap: Estimates of Coefficient of Overlapping for Animal Activity Patterns. R Package Version 0.2.6. http://CRAN.R-project.org/ package = overlap
Milda D, Ramesh T, Kalle R, Gayathri V, Thanikodi M (2020) Ranger survey reveals conservation issues across Protected and outside Protected Areas in southern India. Glob Ecol Conserv 24:e01256
Milda D, Ashish K, Ramesh T, Kalle R, Thanikodi M (2023a) Evaluation of anthropogenic pressure on the occupancy patterns of large mammals in the Western and Eastern Ghats. Landsc Ecol 38(2):1–14
Mukherjee S, Goyal SP, Johnsingh AJT, Pitman ML (2004) The importance of rodents in the diet of jungle cat (Felis chaus), caracal (Caracal caracal) and golden jackal (Canis aureus) in Sariska Tiger Reserve, Rajasthan, India. J Zool 262:405–411
Neilson EW, Avgar T, Burton AC, Broadley K, Boutin S (2018) Animal movement affects interpretation of occupancy models from camera-trap surveys of unmarked animals. Ecosphere 9:e02092
Newsome TM, Van Eeden LM (2017) The effects of food waste on wildlife and humans. Sustainability 9:1269
Ngoprasert D, Lynam AJ, Gale GA (2007) Human disturbance affects habitat use and behaviour of Asiatic leopard Panthera pardus in Kaeng Krachan National Park, Thailand. Oryx 41:343–351
Nickel BA, Suraci JP, Allen ML, Wilmers CC (2020) Human presence and human footprint have non-equivalent effects on wildlife spatiotemporal habitat use. Biol Conserv 241:108383
Noon BR, Bailey LL, Sisk TD, McKelvey KS (2012) Efficient species-level monitoring at the landscape scale. Conserv Biol 26:432–441
O’Brien TG, Kinnaird MF, Wibisono HT (2003) Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. In: Animal Conservation Forum, Cambridge University Press, pp 131–139
Oberosler V, Groff C, Iemma A, Pedrini P, Rovero F (2017) The influence of human disturbance on occupancy and activity patterns of mammals in the Italian alps from systematic camera trapping. Mamm Biol 87:50–61
Odden M, Athreya V, Rattan S, Linnell JD (2014) Adaptable neighbours: movement patterns of GPS-collared leopards in human dominated landscapes in India. PLoS ONE 9:e112044
Ogutu JO, Owen-Smith N (2005) Oscillations in large mammal populations: are they related to predation or rainfall? Afr J Ecol 43:332–339
Ohashi H, Saito M, Horie R, Tsunoda H, Noba H et al (2013) Differences in the activity pattern of the wild boar Sus scrofa related to human disturbance. Eur J Wildl Res 59:167–177
Ordeñana MA, Crooks KR, Boydston EE, Fisher RN, Lyren LM et al (2010) Effects of urbanization on Carnivore species distribution and richness. J Mammal 91:1322–1331
Oriol-Cotterill A, Valeix M, Frank LG, Riginos C, Macdonald DW (2015) Landscapes of coexistence for terrestrial carnivores: the ecological consequences of being downgraded from ultimate to penultimate predator by humans. Oikos 124:1263–1273
Otis DL, Burnham KP, White GC, Anderson DR (1978) Statistical inference from capture data on closed animal populations. Wildl Monogr 62:3–135
Pacifici M, Di Marco M, Watson JE (2020) Protected areas are now the last strongholds for many imperiled mammal species. Conserv Lett 13:e12748
Palmer MS, Swanson A, Kosmala M, Arnold T, Packer C (2018) Evaluating relative abundance indices for terrestrial herbivores from large-scale camera trap surveys. Afr J Ecol 56:791–803
Pardikar R (2020) Small and lesser studied mammals of the Western Ghats need more attention. Mongabay. https://india.mongabay.com/2020/11/small-mammals-of-the-western-ghats-need-more-attention/. Accessed 22 March 2023
Parsons AW, Forrester T, Baker-Whatton MC, McShea WJ, Rota CT et al (2018) Mammal communities are larger and more diverse in moderately developed areas. ELife 7:e38012
Penjor U, Wangdi S, Tandin T, Macdonald DW (2021) Vulnerability of mammal communities to the combined impacts of anthropic land-use and climate change in the himalayan conservation landscape of Bhutan. Ecol Indic 121:107085
Polaina E, González-Suárez M, Revilla E (2019) The legacy of past human land use in current patterns of mammal distribution. Ecography 42:1623–1635
Powers RP, Jetz W (2019) Global habitat loss and extinction risk of terrestrial vertebrates under future land-use-change scenarios. Nat Clim Change 9:323–329
Ramesh T, Downs CT (2015) Impact of land use on occupancy and abundance of terrestrial mammals in the Drakensberg Midlands, South Africa. J Nat Conserv 23:9–18
Ramesh T, Kalle R, Sankar K, Qureshi Q (2012) Spatio-temporal partitioning among large carnivores in relation to major prey species in western ghats. J Zool 287:269–275
Ramesh T, Kalle R, Sankar K, Qureshi Q (2013) Activity pattern of sloth bear Melursus ursinus (Mammalia: Ursidae) in Mudumalai Tiger Reserve, Western Ghats, India. J Threat Taxa 5:3989–3992
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. J Agric Biol Environ Stat 14:322–337
Ripple WJ, Estes JA, Beschta RL, Wilmers CC, Ritchie EG et al (2014) Status and ecological effects of the world’s largest carnivores. Science 343:1241484
Roland CA, Schmidt JH, Nicklen EF (2013) Landscape-scale patterns in tree occupancy and abundance in subarctic Alaska. Ecol Monogr 83:19–48
R Core Team (2020) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rondinini C, Visconti P (2015) Scenarios of large mammal loss in Europe for the 21st century. Conserv Biol 29:1028–1036
Rowan J, Beaudrot L, Franklin J, Reed KE, Smail IE, Zamora A, Kamilar JM (2020) Geographically divergent evolutionary and ecological legacies shape mammal biodiversity in the global tropics and subtropics. Proc Nat Acad Sci 117:1559–1565
Rowcliffe JM, Kays R, Kranstauber B, Carbone C, Jansen PA (2014) Quantifying levels of animal activity using camera trap data. Methods Ecol Evol 5:1170–1179
Roy PS, Behera MD, Murthy MSR, Roy A, Singh S et al (2015) New vegetation type map of India prepared using satellite remote sensing: comparison with global vegetation maps and utilities. Int J Appl Earth Obs Geoinf 39:142–159
Rudramuniyappa MV (1997) Iron ore fines and their impact on environment in Sandur-Hospet region, Bellary district, Karnataka, India
Šálek M, Červinka J, Banea OC, Krofel M, Ćirović D et al (2014) Population densities and habitat use of the golden jackal (Canis aureus) in farmlands across the Balkan Peninsula. Eur J Wildl Res 60:193–200
Šálek M, Drahníková L, Tkadlec E (2015) Changes in home range sizes and population densities of Carnivore species along the natural to urban habitat gradient. Mamm Rev 45:1–14
Samia DS, Nakagawa S, Nomura F, Rangel TF, Blumstein DT (2015) Increased tolerance to humans among disturbed wildlife. Nat Commun 6:8877
Schmitt CB, Burgess ND, Coad L, Belokurov A, Besançon C et al (2009) Global analysis of the protection status of the world’s forests. Biol Conserv 142:2122–2130
Schmitz OJ, Wilmers CC, Leroux SJ, Doughty CE, Atwood TB et al (2018) Animals and the zoogeochemistry of the carbon cycle. Science 362:eaar3213
Shahabuddin G, Thadani R (2018) Biodiversity in community managed landscapes: a view of the potential and constraints in the Van panchayats of the Kumaon Himalaya. Conservation and development in India. Reimagining Wilderness, Routledge, New York, pp 109–131
Shamoon H, Maor R, Saltz D, Dayan T (2018) Increased mammal nocturnality in agricultural landscapes results in fragmentation due to cascading effects. Biol Conserv 226:32–41
Sharp TR, Swaminathan S, Arun AS, Smith T, Satyanarayan K, Seshamani G (2020) Sloth bear attacks on the Deccan Plateau of Karnataka, India. Ursus (31e8):1–11
Sodhi NS, Koh LP, Clements R, Wanger TC, Hill JK et al (2010) Conserving southeast Asian forest biodiversity in human-modified landscapes. Biol Conserv 143:2375–2384
Sunquist M, Sunquist F (2002) Wild cats of the World. University of Chicago Press
Suraci JP, Gaynor KM, Allen ML, Alexander P, Brashares JS et al (2021) Disturbance type and species life history predict mammal responses to humans. Glob Chang Biol 27:3718–3731
Torre I, Pulido T, Vilella M, Díaz M (2022) Mesocarnivore distribution along gradients of anthropogenic disturbance in Mediterranean landscapes. Diversity 14:133
Van Cleave EK, Bidner LR, Ford AT, Caillaud D, Wilmers CC, Isbell LA (2018) Diel patterns of movement activity and habitat use by leopards (Panthera pardus pardus) living in a human-dominated landscape in central Kenya. Biol Conserv 226:224–237
Vanak AT, Gompper ME (2010) Multi-scale resource selection and spatial ecology of the Indian fox in a human‐dominated dry grassland ecosystem. J Zool 281:140–148
Wan X, Jiang G, Yan C, He F, Wen R et al (2019) Historical records reveal the distinctive associations of human disturbance and extreme climate change with local extinction of mammals. PNAS 116:19001–19008
Wang Y, Allen ML, Wilmers CC (2015) Mesopredator spatial and temporal responses to large predators and human development in the Santa Cruz Mountains of California. Biol Conserv 190:23–33
Wei T, Simko V (2021) R Package corrplot: visualization of a correlation matrix. (Version 0.89). https://github.com/taiyun/corrplot
White D, Minotti PG, Barczak MJ, Sifneos JC, Freemark KE et al (1997) Assessing risks to Biodiversity from Future Landscape Change: Evaluación De Riesgos Para La Biodiversidad Debido a Cambios Futuros en El Paisaje. Conserv Biol 11:349–360
Whitworth A, Beirne C, Pillco Huarcaya R, Whittaker L, Serrano Rojas SJ, Tobler MW, MacLeod R (2019) Human disturbance impacts on rainforest mammals are most notable in the canopy, especially for larger-bodied species. Divers Distrib 25:1166–1178
Yoganand K, Rice CG, Johnsingh AJT (2005) Evaluating Panna National Park with special reference to ecology of sloth bear (Melursus ursinus). Final project report. Wildlife Institute of India, Dehradun, India
Yusefi GH, Safi K, Tarroso P, Brito JC (2021) The impacts of extreme climate change on mammals differ among functional groups at regional scale: the case of Iranian terrestrial mammals. Divers Distrib 27:1634–1647
Zamuda KM, Duguid MC, Schmitz OJ (2022) Human land-use effects on mammalian mesopredator occupancy of a northeastern Connecticut landscape. Ecol Evol 12:e9015
Zimmermann F, Foresti D (2016) Capture-recapture methods for density estimation In Rovero F. & Zimmermann F. (Eds.), Camera trapping for wildlife research, pp. 95–141
Acknowledgements
We thank the Director of SACON for providing permission and necessary logistic support to execute this work. We are grateful to Karnataka Forest Department, Ballari Division for providing camera traps, necessary permits, and logistic support. A special thanks to the staff of Karnataka Forest Department, Ballari Division for helping conduct camera trapping surveys. The study was carried out with support from Ballari Forest Division, Karnataka government and the INSPIRE Fellowship, Department of Science and Technology (DST/INSPIRE Fellowship/2028/IF180359), and the SERB Overseas Visiting Doctoral Fellowship (OVDF) (SB/S9/Z-03/2017-V (2020-21)) to A.K.B. A sincere thanks to Zollner lab, Flaherty lab and Swihart lab of The Department of Forestry and Natural Resources at Purdue University for diligent proofreading and providing suggestions that improved this paper.
Funding
The study was carried out with support from Ballari Forest Division, Karnataka Government, the INSPIRE Fellowship, Department of Science and Technology (DST/INSPIRE Fellowship/2028/IF180359) and the SERB Overseas Visiting Doctoral Fellowship (OVDF) (SB/S9/Z-03/2017-V (2020-21)) to the First Author.
Author information
Authors and Affiliations
Contributions
AKB, PAZ, TR & RK conceived the ideas; AKB, PRK & MMP collected the data; AKB, PAZ and TR analysed the data; AKB led the writing; PRK, MMP, PAZ, TR & RK provided editorial input.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Behera, A.K., Kumar, P.R., Priya, M.M. et al. Coexistence in multi-use landscape: linking human activities with functional traits of wild mammals in southern India. Landsc Ecol 39, 41 (2024). https://doi.org/10.1007/s10980-024-01847-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10980-024-01847-7