
Abstract
Context
Mediterranean landscapes are undergoing severe transformations (land-use change and homogenization) that threatens biodiversity and the services these ecosystems provide. These effects can differ amongst biodiversity facets (plants vs. birds, taxonomic vs. functional) and ecosystem functions (e.g., seed dispersal, forest regeneration, pest control), although these are still poorly understood in Mediterranean landscapes.
Objectives
Our study aims at understanding how Mediterranean landscape characteristics and land use (tree croplands, shrublands and pine, oak and mixed forest) affect the diversity of birds and plants, and how these changes in biodiversity could affect ecosystem functioning.
Methods
In 2020–2021, we sampled 49 (5 × 5 m) plots in South-eastern Spain and from five different land uses: tree croplands, shrublands, and pine, oak and mixed forests. We measured land use, habitat diversity and size, together with birds and plant diversity. We assessed both direct and indirect (via bird and plant diversity) effects of landscape characteristics on 6 different ecosystem functions related to forest regeneration, predation, herbivory or aboveground biomass, which imply interactions between birds, plants and their environment.
Results
Plant communities responded mainly to local features (land use), whereas birds did so to habitat size and landscape diversity (within 500 m radius). Seed and caterpillar predation were more affected by bird’s diversity, whereas regeneration capacity of woody species was driven by both plants and birds. Land use was amongst the most important drivers of all measured functions, with generally higher functioning levels in oak and mixed forests than in the rest of land uses.
Conclusions
Our results emphasize the complementary information obtained when considering multiple biodiversity facets in studying the consequences of landscape transformation. We also show that these multiple biodiversity changes can be as important as those of land-use or landscape characteristics for ecosystem functioning.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Mediterranean ecosystems have been managed for centuries, generating the diverse landscapes intermingling near natural and anthropogenic land uses, such as forests, croplands, open woodlands and shrublands that foster their characteristic large biodiversity (Blondel and Aronson 1999). These heterogeneous Mediterranean landscapes are home to more than 10% of the world’s vascular plant and 75% of the European’s bird biodiversity (Médial and Quézel, 1997; Covas and Blondel 2008). However, over the last decades to centuries many natural forests in the Mediterranean Basin have been decimated for wood and charcoal extraction, by urbanization, or via frequent and intensive fires (Blondel 2006; Padilla et al. 2010). Therefore, their regeneration is compromised by limited seedling recruitment (Gil-Tena et al. 2019). In parallel, abandoned agricultural lands are being colonized (or have been afforested) by dense and mono-specific pine stands since 1950s, rendering more homogeneous landscapes with limited potential for biodiversity protection and high risks of wildfires (Maestre and Cortina 2004; Ameztegui et al. 2018; Moghli et al. 2022). In addition, most remaining croplands are under intensive management, performed on large plots without crop margins or understorey vegetation, and with commonly monospecific young and dense orchards that often receive large inputs of fertilizers and pesticides (García-Navas et al. 2022).
All these changes (loss of native forests, abandonment of small crops, mono-specific pine stands, and intensive agriculture) are homogenizing Mediterranean landscapes, leading to biodiversity loss (Bascompte and Rodríguez 2001; Fahrig 2003; Gardiner et al. 2018). However, the effect of this homogenization, and that of different features of the changing landscape configuration (e.g., land use, habitat size, diversity or configuration, or connectivity changes), may have contrasting effects on different biodiversity facets (Hertzog et al. 2019), notably across organisms with contrasting mobility. For example, birds seem to respond more strongly to habitat size than to other changes in landscape configuration (Fahrig 2003; De Camargo et al. 2018) whereas other organisms such as rodents or insects may show stronger responses to connectivity, spatial configuration or habitat diversity changes (Fahrig 2003; Brudvig et al. 2015). Plants, in turn, are more responsive to changes in local land use or land-use intensification (Allan et al. 2015; García-Vega and Newbold 2020). Due to their contrasting responses, focusing on multiple organisms, therefore, can shed light on the overall consequences of ongoing changes in Mediterranean landscape configuration on biodiversity. Biodiversity changes in birds and plants are particularly well-understood (Sullivan et al. 2009; Troudet et al. 2017), are generally good indicators of overall biodiversity change (Padoa-Schioppa et al. 2006; Scherber et al. 2010; Morelli et al. 2021), have high conservation value in the Mediterranean (Médial and Quézel, 1997; Covas and Blondel 2008), and show seemingly contrasting responses to land-use and landscape changes (Fahrig 2003; Allan et al. 2014; Brudvig et al. 2015). Hence, they are a good study system to evaluate the effects of the homogenization of Mediterranean landscapes on biodiversity.
In addition to biodiversity loss, landscape changes in the Mediterranean Basin could also likely have profound implications on ecosystem functioning and nutrient stocks, as has been observed elsewhere. For example, large amounts of carbon are stored into the soil when croplands are reforested (Guo and Gifford 2002; Bárcena et al. 2014), a process that also influences temperature and water balance (Bellot et al. 2004; Peng et al. 2014). Changes in landscape configuration and connectivity are also likely to affect ecological processes involving animal movement, including seed dispersal, pollination, predation or herbivory (Levey et al. 2005; Brudvig et al. 2015; Liu et al. 2018). Yet, the responses of ecosystem functioning to changes in Mediterranean landscapes are still poorly understood.
In addition to the direct effects of landscape configuration, biodiversity changes associated to such configuration could strongly influence ecosystem functioning, particularly when affecting multiple groups of organisms (Soliveres et al. 2016; Barnes et al. 2018; Schudt et al. 2018) or multiple facets of biodiversity (such as taxonomic, functional and/or phylogenetic; Le Bagousse-Pinguet et al. 2019), as the latter complement each other in driving ecosystem functioning. Indeed, both landscape and biodiversity changes are likely to interact in determining ecosystem functioning (Liu et al. 2018; Hertzog et al. 2019; Zirbel et al. 2019). In addition to be good indicators of biodiversity change in response to landscape modifications, birds and plants may complement each other in supplying ecosystem services (reviewed in Cardinale et al. 2012; Wenny et al. 2011). Firstly, seed dispersal by birds plays a key role on the recovery of vegetation in human-modified habitats (García et al. 2010). Secondly, resprouter plant species (those able to regenerate and regrow from underground structures after being damaged, Pausas et al. 2016), many of them dispersed by birds in the Mediterranean, have positive impacts on carbon storage, soil fertility, and nutrient cycling, improving the resilience of the ecosystem to drought and wildfires (Maestre et al. 2009; Cavard et al. 2011; Moghli et al. 2022). Finally, bird and plant diversity co-determine the biological control of pests in forests (Tanhuanpää et al. 2001; Jactel et al. 2021) and farmlands (Whelan et al. 2008; García et al. 2018). Therefore, the combined study of both birds and plants offers a good opportunity to study landscape indirect (mediated by biodiversity changes) effects on functioning.
Our main aim was to understand how landscape characteristics and land use affect the diversity of birds and plants, and how these changes in biodiversity affect ecosystem functioning. To do so, we sampled bird and plant communities (organisms with contrasting mobility and therefore likely differently sensitive to landscape characteristics) in 49 plots within five different land uses typical from Mediterranean landscapes: pine forest, oak forest, mixed deciduous/evergreen forest, shrubland and tree cropland. We sampled habitat characteristics (at local and landscape levels) to assess their effects on bird and plant taxonomic and functional compositions. Finally, we considered how changes in landscape characteristics and biodiversity affect ecosystem functions co-determined by both plants and birds (seedling recruitment, biomass production, seed predation, herbivory, pest [rodent, insect] control). We hypothesized that: (i) birds will respond more strongly to landscape characteristics and plants will do so to local land use, (ii) changes in plants´ and birds’ communities will alter the ecosystem functioning, and (iii) indirect effects of landscape characteristics (mediated by biodiversity changes) on ecosystem functioning will be as important as their direct effects.
Materials and methods
Study area and plot selection
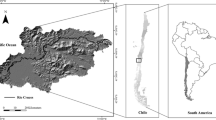
The study took place in two Natural Parks covering similar land uses, landscape configurations, and environmental context: Carrascal de la Font Roja and Serra de Mariola, Alicante, in southeast Spain, (38°39′25″N 0°33′12″W, 2298 ha, 660–1340 m.a.s.l.; 38°43′59″N 0°33′17″W, 17,257 ha, 640–1390 m.a.s.l., respectively, Figure S1). This area is characterized by a dry Mesomediterranean climate, with an average annual temperature of 11° C and mean annual precipitation of 400 mm (Pérez-Cueva 1994), and dominated by calcareous soils. Within this region, we selected 9–10 (5 m × 5 m) plots within five types of land use: Pinus halepensis (pine) forest (N = 10), Quercus rotundifolia (oak) forest (N = 10), shrublands dominated by Cistus albidus, Thymus vulgaris, Stipa offneri and Brachypodium retusum (N = 10), mixed (Q. rotundifolia and deciduous trees such as Fraxinus ornus and Acer opalus) forests (N = 9) and tree cropland, mostly olive (Olea europea) and almond (Prunus dulcis) crops (N = 10). We selected these plots to maximize variation in landscape characteristics (heterogeneity and patch size) and local diversity (based upon local plant diversity).
For each of our 49 plots, we evaluated the area surrounding it (500 m-radius circles with each plot as the centre) to obtain habitat diversity (Shannon index with the proportion of habitats in the 500 m circle), habitat richness (number of different land use types around), and habitat size (proportion of the same land use type within the 500-m circle). The median distance between plots was around 10 km (range 0.05–20 km). Habitat categories considered were mix forest, oak forest, pine forest, shrubland, croplands and urban areas. Habitat richness and diversity were highly correlated to each other (ρ = 0.80) and thus we only considered the latter in our analyses (see below). To obtain these estimates of landscape structure, we used a detailed vegetation map available for the study area (Terrones et al. 2006) and QGIS (QGIS Madeira 3.4) (QGIS.org 2023). Additionally, we visually estimated plant vertical structure as another important plot-level feature, using the cover of different vegetation strata [density of tall and short shrub (plants ha−1), tree cover (%), groundstorey plant cover (%)], litter (%), and rocks, according to Val and others (2018). This index is related to habitat provision for diverse organisms, including vertebrate and invertebrate animals, lichens or fungi.
Plant and bird diversity and composition
At each plot, we estimated the richness of bird species through point counts, where we recorded each bird species heard or seen. The counting time was 15 min, registering each bird detected around the plot (Bibby et al. 1992). This took place between 7:00–11:00 in the morning, avoiding adverse weather conditions such as wind or rain. We distributed the sampling days randomly amongst plots and land uses, to avoid biasing our results. Due to logistical reasons, we only performed bird surveys once per plot (25 plots in 2020, 24 plots in 2021), always with the same observer and during spring (10th–24th June 2020 and 23rd April–21st May 2021). This surely underestimates bird richness, as winter visitors were left out. However, our sampling should still provide a comparative measurement of differences in bird communities across land use types and landscape configurations, and species-accumulation curves suggest that our sampling provided a reasonably complete survey within each land use type (Figure S2). For plants, we estimated plant richness and cover annotating every plant species found in each of our 5 × 5 m plots (see our full database in https://figshare.com/s/466d31af9cdf11991f57).
We used the public databases available of functional traits for plants (TRY; Kattge et al. 2011) and birds (AVONET; Tobias et al. 2022), to obtain the functional traits of the species we found in our plots. From these databases, we obtained plant height, specific leaf area, and seed mass to represent variation in plant resource-use strategy and function (e.g., Wright et al. 2004; Díaz et al. 2016). For birds, we obtained six ecological variables related to trophic niche, range size, habitat, and 11 continuous morphological traits related to size or mobility and flight. From these traits, we estimated functional diversity using Rao’s quadratic entropy (Q), as implemented in the “FD” package in R (Laliberté and Legendre 2010, CRAN). This index is well suited for presence/absence data and mixes of categorical and continuous traits.
Ecosystem processes related to potential services provided by birds and plants
We focused our study on energy and matter transference rates involving both birds and plants, including predation (seeds, leaves, insects and vertebrate) rates, biomass production, and tree regeneration. Specifically, we measured at each plot:
Mouse predation
Some bird species (e.g., birds of prey, Lanius spp.) can render substantial top-down control of potential rodent plagues in croplands, which may also alter, if reaching very high densities, plant composition and biomass, soil fertility, or water infiltration in natural environments (Eldridge and Soliveres 2023). To assess mouse predation rates, we used a modified version of the methodology in Vignieri et al. (2010), see also Bocz et al. (2021). At each plot, we randomly placed six fake mice of different characteristics (contrasting colours (dark grey, white, and grey), material (with hair and without hair), sizes and with/without sound if bitten). Each mouse was separated 50 cm from each other, and loosely fixed to the ground with a 15 cm metal peg and 10 cm wire and left in the field during about 14 days (see further details in Figure S3). We classified between predated (those with missing pieces or pulled out from the wire) or intact mice, estimated the proportion of predated mice, and corrected it by the days spent on the field, as in mouse predation rate = number of predated mice/[total mice (6) * days left in the field (ca. 14)]. Mouse predation could not be assigned to particular predators and thus should be understood as an overall estimation of predation by any animal.
Caterpillar predation
Insectivorous and omnivorous birds help controlling insect pests that may reduce plant productivity (García et al. 2021). Insect damage is also controlled by plant composition and diversity, with rich plant assemblages less likely to be attacked and more likely to recover afterwards (reviewed in Jactel et al. 2021). To assess rates of predation of potential insect pests, we used plasticine caterpillars (Castagneyrol et al. 2019). We placed 20 green plasticine caterpillars in the branches of the most representative plant species at each plot and left them in the field for 14 days approximately. Afterwards, we identified the proportion of caterpillars that were attacked by birds (identified from the marks left in the plasticine). Other caterpillar predators (reptile, mammal, or invertebrates) can also be identified from these marks (Fig. S3), this allowed us to discard these cases to focus on the role of birds as potential pest control agents. In any case, bird attacks accounted for about 20% of the total attacks to caterpillars (the other 80% were attacked by either mammals, arthropods or lizards), and the proportion of caterpillar attacked by birds and those attacked by any predator, results were qualitatively the same as those using only caterpillars damaged by birds. As with the mice, we estimated caterpillar predation rates as the proportion of the 20 caterpillars attacked divided by the number of days in the field.
Herbivory
To complement the measurements of caterpillar predation, we also measured herbivory damage by insects. To do so, we sampled 10 leaves from the main species (i.e., those representing ≥ 50% cover, from 1 to 5 species, 1.86 species on average) of the plot to evaluate presence of signs of herbivory, following the protocol of the Bug-Network (https://bug-net.org/). The method is based upon the estimation of the cover, within each leaf, of each type of damage (four types of invertebrate damage, and four types of fungal damage). Since our study focused on how bird assemblages affect functioning, we focus here only on those leaves damaged by invertebrates (the ones most likely to be influenced by birds). We evaluated the percentage cover of each leaf affected by different insect herbivores (gall-forming, chewers, sap-sucking or rasping, and leaf-miners) and averaged this amount across leaves and dominant plant species to obtain a single value per plot.
Aboveground biomass
As an integrative measure of pest control and aboveground productivity, we measured shrub and herbaceous biomass, clipping and harvesting all plants at 1 cm aboveground in five 0.5 × 0.5 m quadrats at each plot. Samples were then oven-dried (24 h minimum, 60 °C) and weighed.
Seed predation
We followed the methodology of Castro et al. (1999). In each plot, we placed one Petri dish (9 cm in diameter), each with 50 pine nuts, during around 14 days. The Petri dish was hanged from a branch with ropes, to let just birds to access it.
Regeneration capacity
The regeneration of Mediterranean forests is severely limited by the lack of new seedlings (Gil-Tena et al. 2019). This limitation can partly be buffered by seed-dispersal by birds, as these are major dispersers of acorn and fleshy-fruited woody species (García et al. 2010; Bueno et al. 2021). To account for differences in forest regeneration capacity across different land use types and landscape configurations, we estimated the quantity and richness of all young woody plant seedlings at each plot. Not all, but the vast majority, of woody seedlings found, were generally dispersed by birds. Since quantity and richness were highly correlated to each other (ρ = 0.88), we use here quantity as a measurement of tree regeneration capacity.
Statistical analyses
We used linear models to analyse the response of biodiversity to land-use and landscape characteristics. Specifically, our response variables were bird richness, plants richness, functional diversity of birds, and functional diversity of plants. It was necessary to transform RaoQ of birds and plants to obtain a normal distribution of data, with the inverse of RaoQ for birds and the logarithm of RaoQ for plants, the rest of response variables fulfilled analyses´ assumptions. On the other hand, the predictors were the type of land use classified in situ (pine forest, mixed forest, oak forest, shrubland or cropland), vertical structure of vegetation, habitat diversity (evenness of the habitat in the 500 m radius surrounding area) and the percentage of habitat similar to the one established in situ in 500 m radius surrounding area (habitat size). Since we were interested on dissecting the effects of landscape characteristics on diversity, we considered all the possible interactions between habitat and landscape predictors (land-use type, habitat diversity and habitat size), we also included the coordinates of each plot and the year of sampling to control for spatial and temporal autocorrelation in our data. We did not find any strong correlation between predictors, which allowed us to consider all of them simultaneously without inducing multicollinearity issues in our analyses. In sum, our models have the following structure:
From this initial structure, we obtained the most parsimonious models for each of our response variables selecting those by applying multimodel inference using the “dredge” function from MuMIn package in R (Bartoń, 2009). This method tries all possible combinations of predictors departing from the full model and selects those models with a delta AIC < 2 (models within 2 AIC units of that with the lowest AIC were considered equally plausible). Then, we selected the predictors presented in this model’s list.
For the case of birds, we considered plant characteristics (species richness, functional diversity and vertical structure) as additional predictors for bird’s diversity and composition. Hence, we followed the same process mentioned before, but the structure of the full model was:
This structure allowed us to test interactions between landscape and plant features. Those interactions are tested because it allows us to elucidate whether the effect of diversity of plants and birds is context dependent (on landscape characteristics), an important but usually overlooked possibility.
To analyse the direct and indirect (mediated through changes in plant and bird diversity and composition) effects of land-use and landscape configuration on ecosystem functioning, we used structural equation modelling (SEMs; Grace 2006). Our a priori model structure included the direct effect of land use, habitat size and habitat diversity on plant diversity and composition (richness, vertical structure and functional diversity), bird diversity and composition (richness, functional diversity, and proportion of bird species that were granivores, invertivores or omnivores) and ecosystem functioning (listed below). We also considered the effects of plant and bird diversity and composition on ecosystem functioning. We used six separate SEMs to analyse our data: one per each function (regeneration capacity, seed predation, mouse predation, caterpillar predation, biomass and herbivory). To fit our categorical “land use” predictor, we used binomial dummy variables (e.g., pine forest yes/no, oak forest yes/no, etc.) coupled to a composite variable to obtain their overall effect (Grace and Bollen 2008).
We estimated how well our a priori structure fitted our data using χ2, RMSEA and Bollen-Stine overall goodness-of-fit estimators. Since all these estimators rendered very similar results, we only present here results on χ2, the P-values of which (much larger than 0.05) suggested a good fit between our data and our a priori structure in all cases. Although we included originally habitat size, this predictor contributed little to the amount of explained variation in the diversity and composition of plants and birds, and it did not enhance the R2 in any of the functions studied. Thus, we decided to remove this predictor, which enhanced the simplicity and overall goodness-of-fit of our models. We also estimated path coefficients, and their associated P-values, using bootstrapping (900 iterations) instead of maximum likelihood because our relatively limited sample size. We used R version 4.2.1 (R Core Team 2022) and RStudio 2022.12.0.353 (Posit Team 2022) to perform all the analyses, excepting the SEMs, which were implemented in AMOS AMOS for windows (SPSS Inc., Chicago, IL, USA).
Results
We found 193 plant and 60 bird species, including some of special conservation interest such as the endemism Centaurea mariolensis (Serra 2011) or the locally rare blue tit (Cyanistes caeruleus; López-Iborra et al. 2015). The most common plant species were Rubia peregrina subsp. peregrina and Quercus rotundifolia appearing in more than 50% of the plots. On the other hand, the most common bird species were Serinus serinus, Cyanistes caeruleus and Fringilia coelebs appearing in around 90% of the plots. Tree croplands were the most species-rich land use, birds usually feed on this type of habitat and annual plants were abundant on croplands (Table 1). See supplementary material for the full species lists (https://figshare.com/s/466d31af9cdf11991f57).
Plant and bird richness showed contrasting responses to land-use and landscape characteristics. Plants richness responded more strongly to local land use whereas that of birds was more responsive to landscape characteristics (habitat diversity and size; Fig. 1). Functional diversity responded different from species richness on both organisms. In general, functional diversity was more responsive to local factors in both organisms, and the amount of variation explained for both plants and birds was much higher than for species richness (0.18 vs. 0.27 in plants; 0.20 vs. 0.49 in birds) (Fig. 1).

Results from linear models, only those predictors selected in the most parsimonious models (based on AIC) are shown. Bars reflect the effect size (+/− standard error) of each predictor. Significance codes mean: P-values are: ***P < 0.001; **P < 0.01; *P < 0.05. ° 0.1 > P > 0.05; “x” means interaction between predictors
Overall, land use (tree cropland, mix, oak and pine forest and shrubland) was perhaps our most important predictor. It affected significantly plant richness and functional diversity and marginally affected bird functional diversity, and had substantial direct and indirect effects on most of the functions considered (Fig. 2). Tree croplands hosted more plants species than mixed forests, although most of them were annuals and functional diversity showed the opposite trend (peaking in oak and mixed forests (Fig. S4). On the other hand, bird richness and functional diversity showed similar values, with both showing slightly higher values in tree croplands and pine forests than on the others land uses (Fig S4). When we added plant attributes on bird’s diversity models, the amount of variance explained dramatically increased (from R2 = 0.18 to 0.35 in species richness, and from 0.27 to 0.58 in functional diversity). We found a positive interaction between habitat size and plant richness: the positive effect of plant richness on bird richness increased with habitat size. In addition, vertical structure and habitat size also interacted to determine bird richness; a complex vertical structure only benefitted bird diversity in large habitats (Fig. S5), although it benefitted functional diversity in any case (Fig. 1).

General structural equation model (A), showing the fixed relationships between land-use and landscape characteristics and our measurements of plant (green dashed box) and bird (orange dashed box) diversity and composition. Overall goodness-of-fit (χ2; degrees of freedom [df] and P-value) and proportion of variance explained for each endogenous variable (R2) are shown. Grey lines show the paths whose values change in the models fitted individually for each one of our ecosystem functions (from B–G). Acronyms: FD: Functional Diversity, TG: Trophic group, SR: Species richness, VS: Vertical structure. Composite variables are shown with hexagons. Standardized path coefficients are shown for each significant path. The width of arrows is proportional to the path coefficient, with black and red for positive and negative relationships, respectively. P-values are: ***P < 0.001; **P < 0.01; *P < 0.05
The percentage of variance explained by our SEMs ranged between 54% (regeneration capacity) and 31% (caterpillar predation), suggesting that our predictors did reasonably well in explaining variation in our ecosystem functions. For all functions, we found strong direct effects of land use (Figs. 2 and 3), which were not along the same lines that those found for richness. Contrary to plant and bird richness, and bird functional diversity, we found generally higher functioning rates in oak or mixed forests than in the rest of land uses, excepting for caterpillar and mouse predation (Fig. 3). These opposite effects on biodiversity and function rendered substantially smaller total than direct effects of land use on functioning (Fig. 3), which was particularly evident for regeneration capacity and seed predation. For these functions, a large proportion of the direct effects of land use was dampened by their indirect effects mediated by biodiversity change, which worked on opposite direction. Changes in plant and bird diversity and composition showed contrasting influences on ecosystem functioning, although in general these were of similar size than land-use direct effects. Bird richness had generally stronger effects than plant richness, enhancing seed and caterpillar predation rates, but declining mice predation. Relationships between functional diversity and ecosystem functioning were generally strong but varied depending on the function and organism considered. For plants, vertical structure and richness were the most important drivers of functioning, with contrasting effects on mouse (positive) and caterpillar (negative) predation. In birds, functional diversity reduced regeneration capacity and increased seed predation rates, which overall suggests that more functionally diverse bird assemblages may hinder plant recruitment dynamics in the studied landscapes. The dominance of particular bird feeding groups was a stronger predictor for functioning than bird functional diversity (four vs. two significant paths), generally enhancing seed, insect and mouse predation rates, and consequently decreasing leaf damage by insect herbivory.

Representation of standardized total effects from SEMs for each ecosystem function. On X axis we specified every predictor, but we grouped by colours categories; red: predictors related to the landscape, orange: type of land use, mauve: predictors related with plants (plant richness, vertical structure, functional diversity of plants (FD plants), and purple: predictors related with birds (birds’ richness, functional diversity of birds (FD birds), trophic group)
Discussion
We evaluated landscape characteristics in a typical Mediterranean area, mixing oak and pine forest, shrublands and croplands, and how they affected biodiversity and ecosystem functioning. As expected, plant richness responded more strongly to local than landscape-level characteristics, like the type of land use (see also Allan. et al (2015) for a report on the effects of land-use intensification). We also found significant positive effects of landscape diversity on both plant and bird richness, similar to those found by Bascompte and Rodríguez (2001), which clearly suggests that ongoing landscape homogenization in Mediterranean landscapes threatens with further biodiversity loss. In addition, we found that more homogeneous habitats (larger size of the same habitat) have a negative impact on plant richness (Yang et al. 2015), contrary to what was found for bird richness. In the case of bird’ richness, they responded more strongly to landscape- than local-level characteristics, perhaps due to their mobility, preferring larger and more diverse habitats, according with De Camargo et al. (2018) where habitat amount was more important than other landscape features. Functional diversity of both birds and plants also showed contrasting responses compared to species richness patterns, especially in response to land use (Fig. 1, S4). Tree croplands showed higher plant species richness than other land uses, but lower functional diversity, suggesting that most species within tree croplands are functionally homogeneous. However, other land uses, such as oak forests, presented less average plant richness, but larger functional diversity, which could partly explain their higher levels of ecosystem functioning (see also Arroyo-Rodríguez et al. 2020).
In birds, functional diversity and species richness showed similar values across the different land uses studied. Perhaps birds could maintain functional diversity in tree croplands relying on the semi-natural area existing in the surroundings (Barbaro et al. 2021). Additionally, some functional groups could benefit from agroecosystems, due to high productivity and greater food resources (Tscharntke et al. 2008). Also, a proper management of agroecosystems can promote functional diversity and species richness (García-Navas et al. 2022). The latter is supported by the interactions between plant and landscape characteristics we found. This could suggest an important modulation of landscape and plant diversity factors to explain bird richness. In particular, plant richness and plant vertical structure were particularly beneficial for birds in large habitats (habitat size x plant richness, and habitat size x vertical structure interactions; Fig. 1, S5). The effect of these different drivers of bird diversity have been studied separately; either habitat size (Farhig, 2003, De Camargo et al. 2018), or vertical structure of vegetation (Val et al. 2018; Barbaro et al. 2021; García-Navas et al. 2022) or plant richness (Cavard et al. 2011; Paker et al. 2014), but interactions between plant and landscape characteristics have been seldom explored. We found that these different drivers can interact strongly, and that benefits of sustainable local managements can be further exacerbated if applied in larger habitat remnants or in more heterogeneous landscapes.
Our initial hypothesis regarding the functional responses to landscape configuration in plant and bird biodiversity was also supported. In general, birds had more relevance than plants in most of the functions considered. This highlights the important role of birds on pest regulation (Mols and Visser 2002; García et al. 2018), reducing herbivory damage on plants (Koh 2008). Bird diversity also enhanced seed predation rates, which could either be interpreted in relation to a “seed dispersal” service (García et al. 2010). Indeed, 65% of seed dispersal is driven by animals in the Mediterranean, and birds play an important role in this process (Bueno et al. 2021). However, seed predation also hinders plant recruitment, as we observed, by damaging the seed bank. Importantly, the prevalence of the relevant bird trophic group (either granivores, omnivores or invertivores, depending on the function of interest) was a major driver of ecosystem functioning (see also Stewart et al. 2022) and did not respond to landscape characteristics. In addition to modulate bird diversity, plant attributes -mainly their functional diversity- also showed direct relationships with ecosystem functioning. Higher plant functional diversity dampened and benefitted caterpillar and mouse predation, respectively, supporting the notion that habitat characteristics can influence predator-prey interactions and that plants can act as refuge for some prey species (Bruno and Cardinale 2008; Grof-Tisza et al. 2017).
As for plants, landscape characteristics (habitat diversity) were less important than land use to determine ecosystem functioning (Fig. 2). We found both direct and indirect (mediated by biodiversity) effects of land use on functioning, as initially hypothesized. However, the opposite trends in the effects of land-use had opposite effects depending on the biodiversity measure (positive for plants and negative for birds), or the function considered (negative for predation-related functions, positive for biomass). These opposite effects led to weaker total than direct effects of land-use on ecosystem functioning. The relative importance of direct vs. indirect effect also depended on the function we considered, with a larger importance of indirect effects for seed or caterpillar predation, or regeneration capacity, than for others. For example, aboveground biomass was directly affected by land use (peaking in shrublands) with little influence of bird or plant attributes. This suggests that biomass could vary more depending on the type of habitat than on local diversity (Anderson et al. 2010), or that the effect of land-use type is already accounting for higher/lower diversity in certain land-use classes. In contrast to aboveground biomass, however, the effect of abiotic attributes (landscape and land use) and biotic attributes (birds and plants) were equally important in general. The only exception to this general rule was regeneration capacity, which was mostly driven by bird and plant attributes. Regeneration capacity was limited by functional diversity of birds, maybe due to the high predation of the seeds of forest species like Pinus spp. (Castro et al. 1999). Also, both vertical structure and some land uses (tree cropland, mixed forest and shrubland) hindered the regeneration of woody species. This could be due to the limitations of dispersal vectors, which could change their behaviour and efficiency of dispersion modulated by the habitat type and environmental heterogeneity (Escribano-Avila et al. 2012).
Our study presents a novel approach collectively considering changes in Mediterranean local and landscape-characteristics, multiple facets of bird and plant diversity and an array of important ecosystem functions. Our results show contrasting responses of different biodiversity facets (plants and birds’ richness, taxonomic vs. functional diversity) and ecosystem functions to landscape characteristics, responding plants and functions more to local characteristics, and birds to habitat size and landscape diversity. Changes in plant and bird biodiversity were as important as those of land-use or landscape characteristics to determine ecosystem functioning. Indeed, seed and caterpillar predation (related to dispersal and pest control services) were more controlled by bird features, whereas regeneration capacity of woody species was mostly driven by plants and birds more than by land use. Our study provides a comprehensive assessment of how ecosystem functions respond to changes in biodiversity of birds and plants, including the modulating role of landscape in this process. By doing so, our study provides a framework that may be of use for ecosystem managing at the landscape level.
Data availability
Data will be made available on Figshare (“https://figshare.com/s/466d31af9cdf11991f57”) after acceptance.
References
Allan E, Bossdorf O, Dormann CF, Prati D, Gossner MM, Tscharntke T, Blüthgen N, Bellach M, Birkhofer K, Boch S, Böhm S, Börschig C, Chatzinotas A, Christ S, Daniel R, Diekötter T, Fischer C, Friedl T, Glaser K, …, Fischer M (2014) Interannual variation in land-use intensity enhances grassland multidiversity. Proc Natl Acad Sci USA 111(1):308–313.
Allan E, Manning P, Alt F, Binkenstein J, Blaser S, Blüthgen N, Böhm S, Grassein F, Hölzel N, Klaus VH, Kleinebecker T, Morris EK, Oelmann Y, Prati D, Renner SC, Rillig MC, Schaefer M, Schloter M, Schmitt B, …, Fischer M (2015) Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecol Lett 18(8):834–843.
Ameztegui A, Gil-Tena A, Faus J, Piqué M, Brotons L, Camprodon J (2018) Bird community response in mountain pine forests of the Pyrenees managed under a shelterwood system. For Ecol Manag 407:95–105.
Anderson PML, Hoffman MT, O’Farrell PJ (2010) Above ground perennial plant biomass across an altitudinal and land-use gradient in Namaqualand, South Africa. South Afr J Bot 76(3):471–481.
Arroyo-Rodríguez V, Fahrig L, Tabarelli M, Watling JI, Tischendorf L, Benchimol M, …, Tscharntke T (2020) Designing optimal human‐modified landscapes for forest biodiversity conservation. Ecol Lett 23(9):1404–1420
Barbaro L, Assandri G, Brambilla M, Castagneyrol B, Froidevaux J, Giffard B, Pithon J, Puig-Montserrat X, Torre I, Calatayud F, Gaüzère P, Guenser J, Macià‐Valverde F, Mary S, Raison L, Sirami C, Rusch A (2021) Organic management and landscape heterogeneity combine to sustain multifunctional bird communities in european vineyards. J Appl Ecol 58(6):1261–1271.
Bárcena TG, Kiær LP, Vesterdal L, Stefánsdóttir HM, Gundersen P, Sigurdsson BD (2014) Soil carbon stock change following afforestation in Northern Europe: a meta-analysis. Glob Change Biol 20(8):2393–2405.
Barnes AD, Jochum M, Lefcheck JS, Eisenhauer N, Scherber C, O’Connor MI, de Ruiter P, Brose U (2018) Energy Flux: the link between Multitrophic Biodiversity and Ecosystem Functioning. Trends Ecol Evol 33(3):186–197.
Barton K (2009) Mu-MIn: Multi-model inference. R Package Version 0.12.2/r18. http://R-Forge.R-project.org/projects/mumin/
Bascompte J, Rodríguez M (2001) Habitat patchiness and plant species richness. Ecol Lett 4(5):417–420.
Bellot J, Maestre FT, Chirino E, Hernández N, de Urbina JO (2004) Afforestation with Pinus halepensis reduces native shrub performance in a Mediterranean semiarid area. Acta Oecol 25(1–2):7–15.
Bibby CJ, Burgess ND, Hill DA (1992) Point Counts. Bird Census Techniques. Academic press, Cambridge, pp 85–104.
Blondel J (2006) The ‘Design’ of Mediterranean Landscapes: a millennial story of humans and Ecological Systems during the historic period. Hum Ecol 34(5):713–729.
Blondel J, Aronson J (1999) Biology and wildlife of the Mediterranean region. Oxford University Press, Oxford
Bocz R, Batáry P, Purger JJ (2021) Scent, rather than fur pattern, determines predation of mice: an in-the-wild experiment with plasticine mouse models. J Zool 316:223–228.
Brudvig LA, Damschen EI, Haddad NM, Levey DJ, Tewksbury JJ (2015) The influence of habitat fragmentation on multiple plant–animal interactions and plant reproduction. Ecology 96(10):2669–2678.
Bruno JF, Cardinale BJ (2008) Cascading effects of predator richness. Front Ecol Environ 6(10):539–546.
Bueno RdaS, García D, Galetti M, La Mantia T (2021) Trophic and spatial complementarity on seed dispersal services by birds, wild mammals, and cattle in a Mediterranean woodland pasture. Global Ecol Conserv 31:e01880.
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, MacE GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature 486(7401):59–67.
Castagneyrol B, Valdés-Correcher E, Bourdin A, Barbaro L, Bouriaud O, Branco M, Csóka G, Duduman M-L, Dulaurent A-M, Eötvös CB, Ferrante M, Fürjes-Mikó Á, Galman A, Gossner MM, Harvey D, Howe AG, Kaennel-Dobbertin M, Koricheva J, Löveï GL, Lupaștean D (2019) Can school children support ecological research? Lessons from the “Oak bodyguard” citizen science project. Juha-Matti Pitkänen 14:22.
Castro J, Gómez JM, García D, Zamora R, Hódar JA (1999) Seed predation and dispersal in relict Scots pine forests in southern Spain. Plant Ecology 145:115–123
Cavard X, Macdonald SE, Bergeron Y, Chen HYH (2011) Importance of mixedwoods for biodiversity conservation: evidence for understory plants, songbirds, soil fauna, and ectomycorrhizae in northern forests. Environ Rev 19:142–161.
R Core Team (2022) R: A language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/
Covas R, Blondel J (2008) Biogeography and history of the Mediterranean bird fauna. Ibis 140(3):395–407.
De Camargo RX, Boucher-Lalonde V, Currie DJ (2018) At the landscape level, birds respond strongly to habitat amount but weakly to fragmentation. Divers Distrib 24(5):629–639.
Díaz S, Kattge J, Cornelissen JHC, Wright IJ, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Prentice C, Garnier I, Bönisch E, Westoby G, Poorter M, Reich H, Moles PB, Dickie AT, Gillison J, Zanne AN, Gorné AE, L. D (2016) The global spectrum of plant form and function. Nature 529(7585):167–171.
Eldridge DJ, Soliveres S (2023) Rewilding soil-disturbing vertebrates to rehabilitate degraded landscapes: benefits and risks. Biol Lett 19:20220544. https://doi.org/10.1098/rsbl.2022.0544
Escribano-Avila G, Sanz-Pérez V, Pías B, Virgós E, Escudero A, Valladares F (2012) Colonization of Abandoned Land by Juniperus thurifera is mediated by the Interaction of a diverse dispersal assemblage and environmental heterogeneity. PLoS ONE 7(10):1–10.
Fahrig L (2003) Effects of Habitat Fragmentation on Biodiversity. Ann Rev Ecol Evol Syst 34:487–515.
García D, Zamora R, Amico GC (2010) Birds as suppliers of seed dispersal in temperate ecosystems: conservation guidelines from real-world landscapes: seed-dispersal by frugivorous birds. Conserv Biol 24(4):1070–1079.
García D, Miñarro M, Martínez-Sastre R (2018) Birds as suppliers of pest control in cider apple orchards: avian biodiversity drivers and insectivory effect. Agric Ecosyst Environ 254:233–243.
García D, Miñarro M, Martínez-Sastre R (2021) Enhancing ecosystem services in apple orchards: nest boxes increase pest control by insectivorous birds. J Appl Ecol 58(3):465–475.
García-Navas V, Martínez-Núñez C, Tarifa R, Manzaneda AJ, Valera F, Salido T, Camacho FM, Isla J, Rey PJ (2022) Agricultural extensification enhances functional diversity but not phylogenetic diversity in Mediterranean olive groves: a case study with ant and bird communities. Agric Ecosyst Environ 324:107708.
García-Vega D, Newbold T (2020) Assessing the effects of land use on biodiversity in the world’s drylands and Mediterranean environments. Biodivers Conserv 29(2):393–408.
Gardiner R, Bain G, Hamer R, Jones ME, Johnson CN (2018) Habitat amount and quality, not patch size, determine persistence of a woodland-dependent mammal in an agricultural landscape. Landscape Ecol 33(11):1837–1849.
Gil-Tena A, Morán-Ordóñez A, Comas L, Retana J, Vayreda J, Brotons L (2019) A quantitative assessment of mid-term risks of global change on forests in western Mediterranean Europe. Reg Envriron Chang 19(3):819–831.
Grace J (2006) Structural equation modeling and Natural Systems. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511617799
Grace JB, Bollen KA (2008) Representing general theoretical concepts in structural equation models: the role of composite variables. Environ Ecol Stat 15(2):191–213.
Grof-Tisza P, LoPresti E, Heath SK, Karban R (2017) Plant structural complexity and mechanical defenses mediate predator–prey interactions in an odonate–bird system. Ecol Evol 7:1650–1659.
Guo LB, Gifford RM (2002) Soil carbon stocks and land use change: a meta-analysis: SOIL CARBON STOCKS and LAND USE CHANGE. Glob Change Biol 8(4):345–360.
Hertzog LR, Boonyarittichaikij R, Dekeukeleire D, de Groote SRE, van Schrojenstein Lantman IM, Sercu BK, Smith HK, de la Peña E, Vandegehuchte ML, Bonte D, Martel A, Verheyen K, Lens L, Baeten L (2019) Forest fragmentation modulates effects of tree species richness and composition on ecosystem multifunctionality. Ecology. https://doi.org/10.1002/ecy.2653
Jactel H, Moreira X, Castagneyrol B (2021) Tree diversity and forest resistance to insect pests: patterns, mechanisms, and prospects. Ann Rev Entomol 66(1):277–296.
Kattge J, Díaz S, Lavorel S, Prentice IC, Leadley P, Bönisch G, Garnier E, Westoby M, Reich PB, Wright IJ, Cornelissen JHC, Violle C, Harrison SP, Van Bodegom PM, Reichstein M, Enquist BJ, Soudzilovskaia NA, Ackerly DD, Anand M, …, Wirth C (2011) TRY - a global database of plant traits. Glob Change Biol 17(9):2905–2935.
Koh LP (2008) Birds defend oil palms from herbivorous insects. Ecol Appl 18(4):821–825.
Laliberté E, Legendre P (2010) A distance-based framework for measuring functional diversity from multiple traits. Ecology 91(1): 299–305. http://www.ecolag.univ-montp2.fr/softwarei
Le Bagousse-Pinguet Y, Soliveres S, Gross N, Torices R, Berdugo M, Maestre FT (2019) Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality. Proc Natl Acad Sci 116(17):8419–8424.
Levey DJ, Bolker BM, Tewksbury JJ, Sargent S, Haddad NM (2005) Effects of Landscape Corridors on seed dispersal by birds. Science 309(5731):146–148.
Liu J, Wilson M, Hu G, Liu J, Wu J, Yu M (2018) How does habitat fragmentation affect the biodiversity and ecosystem functioning relationship? Landscape Ecol 33(3):341–352.
López Iborra GM, Bañuls Patiño A, Zaragozí Llenes A, Sala Bernabeu J, Izquierdo Rosique A, Martínez Pérez JE, Ramos Sánchez J, Bañuls Patiño D, Morcillo A, Sánchez S, Zapata JA, Roig C, Reig B, Ferrer A (2015) Atlas de las aves nidificantes en la provincia de Alicante. Publicacions de la Universitat d’Alacant - SEO/Alicante, Alicante
Maestre FT, Cortina J (2004) Are Pinus halepensis plantations useful as a restoration tool in semiarid Mediterranean areas? For Ecol Manag 198(1–3):303–317.
Maestre FT, Callaway RM, Valladares F, Lortie CJ (2009) Refining the stress-gradient hypothesis for competition and facilitation in plant communities. J Ecol 97:199–205.
Médail F, Quézel P (1997) Hot-spots analysis for conservation of plant biodiversity in the Mediterranean basin. Ann Mo Bot Gard 84(1):112.
Moghli A, Santana VM, Soliveres S, Baeza MJ (2022) Thinning and plantation of resprouting species redirect overstocked pine stands towards more functional communities in the Mediterranean basin. Sci Total Environ 806:150715.
Mols CMM, Visser ME (2002) Great tits can reduce caterpillar damage in apple orchards: great tits reduce caterpillar damage. J Appl Ecol 39(6):888–899.
Morelli F, Reif J, Díaz M, Tryjanowski P, Ibáñez-Álamo JD, Suhonen J, Jokimäki J, Kaisanlahti-Jokimäki M-L, Møller P, Bussière A, Mägi R, Kominos M, Galanaki T, Bukas A, Markó N, Pruscini G, Jerzak F, Ciebiera L, O., Benedetti Y (2021) Top ten birds indicators of high environmental quality in european cities. Ecol Ind 133:108397.
Padilla FM, Vidal B, Sánchez J, Pugnaire FI (2010) Land-use changes and carbon sequestration through the twentieth century in a Mediterranean mountain ecosystem: implications for land management. J Environ Manage 91(12):2688–2695.
Padoa-Schioppa E, Baietto M, Massa R, Bottoni L (2006) Bird communities as bioindicators: the focal species concept in agricultural landscapes. Ecol Ind 6(1):83–93.
Paker Y, Yom-Tov Y, Alon-Mozes T, Barnea A (2014) The effect of plant richness and urban garden structure on bird species richness, diversity and community structure. Landsc Urban Plann 122:186–195.
Pausas JG, Pratt RB, Keeley JE, Jacobsen AL, Ramirez AR, Vilagrosa A, Paula S, Kaneakua-Pia IN, Davis SD (2016) Towards understanding resprouting at the global scale. New Phytol 209(3):945–954.
Peng S-S, Piao S, Zeng Z, Ciais P, Zhou L, Li LZX, Myneni RB, Yin Y, Zeng H (2014) Afforestation in China cools local land surface temperature. Proc Natl Acad Sci 111(8):2915–2919.
Pérez-Cueva AJ (1994) Atlas climático de la Comunidad Valenciana. Conselleria de Obras Públicas, Urbanismo y Transportes, Valencia
Posit Team (2022) RStudio: Integrated Development Environment for R. Posit Software, PBC. http://www.posit.co/
QGIS.org. (2023). QGIS Geographic Information System. Open Source Geospatial Foundation Project. http://www.qgis.org
Scherber C, Eisenhauer N, Weisser WW, Schmid B, Voigt W, Fischer M, Schulze ED, Roscher C, Weigelt A, Allan E, Beler H, Bonkowski M, Buchmann N, Buscot F, Clement LW, Ebeling A, Engels C, Halle S, Kertscher I, …, Tscharntke T (2010) Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment. Nature 468(7323):553–556.
Schuldt A, Assmann T, Brezzi M, Buscot F, Eichenberg D, Gutknecht J, Härdtle W, He J-S, Klein A-M, Kühn P, Liu X, Ma K, Niklaus PA, Pietsch KA, Purahong W, Scherer-Lorenzen M, Schmid B, Scholten T, Staab M, …, Bruelheide H (2018) Biodiversity across trophic levels drives multifunctionality in highly diverse forests. Nat Commun 9(1):2989.
Serra L (2011) Flora del parc natural de la Font Roja Luis Serra Laliga, Spain
Soliveres S, Van Der Plas F, Manning P, Prati D, Gossner MM, Renner SC, Alt F, Arndt H, Baumgartner V, Binkenstein J, Birkhofer K, Blaser S, Blüthgen N, Boch S, Böhm S, Börschig C, Buscot F, Diekötter T, Heinze J, …, Allan E (2016) Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality. Nature 536(7617):456–459.
Stewart PS, Voskamp A, Santini L, Biber MF, Devenish AJM, Hof C, Willis SG, Tobias JA (2022) Global impacts of climate change on avian functional diversity. Ecol Lett 25(3):673–685.
Sullivan BL, Wood CL, Iliff MJ, Bonney RE, Fink D, Kelling S (2009) eBird: a citizen-based bird observation network in the biological sciences. Biol Conserv 142(10):2282–2292.
Tanhuanpää M, Ruohomäki K, Uusipaikka E (2001) High larval predation rate in non-outbreaking populations of a geometrid moth. Ecology 82:281–289.
Terrones B, Bonet A, Carchano R, Brotons J, Segura M (2006) Cartografía de la cubierta vegetal del parque natural del carrascal de la font roja. Iberis 4:73–88
Tobias JA, Sheard C, Pigot AL, Devenish AJM, Yang J, Sayol F, Neate-Clegg MHC, Alioravainen N, Weeks TL, Barber RA, Walkden PA, MacGregor HEA, Jones SEI, Vincent C, Phillips AG, Marples NM, Montaño‐Centellas FA, Leandro‐Silva V, Claramunt S, …, Schleuning M (2022) AVONET: morphological, ecological and geographical data for all birds. Ecol Lett 25(3):581–597.
Troudet J, Grandcolas P, Blin A, Vignes-Lebbe R, Legendre F (2017) Taxonomic bias in biodiversity data and societal preferences. Sci Rep 7(1):9132.
Tscharntke T, Sekercioglu CH, Dietsch TV, Sodhi NS, Hoehn P, Tylianakis JM (2008) Landscape constraints on functional diversity of birds and insects in tropical agroecosystems. Ecology 89(4):944–951.
Val J, Eldridge DJ, Travers SK, Oliver I (2018) Livestock grazing reinforces the competitive exclusion of small-bodied birds by large aggressive birds. J Appl Ecol 55(4):1919–1929.
Vignieri SN, Larson JG, Hoekstra HE (2010) The selective advantage of crypsis in mice. Evolution 64(7):2153–2158.
Wenny DG, DeVault TL, Johnson MD, Kelly D, Sekercioglu CH, Tomback DF, Whelan CJ (2011) Perspectives in ornithology the need to quantify ecosystem services provided by birds. Auk 128(1):1–14.
Whelan CJ, Wenny DG, Marquis RJ (2008) Ecosystem services provided by birds. Ann NY Acad Sci 1134(1):25–60.
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Lusk C, …, Villar R (2004) The worldwide leaf economics spectrum. Nature 428(6985):821–827.
Yang Z, Liu X, Zhou M, Ai D, Wang G, Wang Y, Chu C, Lundholm JT (2015) The effect of environmental heterogeneity on species richness depends on community position along the environmental gradient. Sci Rep 5(1):15723.
Zirbel CR, Grman E, Bassett T, Brudvig LA (2019) Landscape context explains ecosystem multifunctionality in restored grasslands better than plant diversity. Ecology. https://doi.org/10.1002/ecy.2634
Acknowledgements
Staff from P.N. del Carrascal de la Font Roja and P. N. Sierra de Mariola helped finding suitable study sites. Dr. Lluís Serra helped identifying plant species. Staff from “Fundación Victoria Laporta” support our study with pictures. This research is funded by the Spanish Ministry of Science, Innovation and Universities, through project “Identifying the functional consequences of changes in biodiversity at various spatial scales” (FOBIASS; RTI2018-098895-A-100). The author(s) declare no conflict of interest.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This work is supported by Ministerio de Ciencia, Innovación y Universidades (Grant no.: RTI2018-098895-A-100).
Author information
Authors and Affiliations
Contributions
SS conceived the idea. PL and SS conducted fieldwork. PL curated and analysed the data, with assistance of MB and SS. PL wrote the first draft of the manuscript, and all authors contributed revisions.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lopezosa, P., Berdugo, M. & Soliveres, S. On the relative importance of land use, landscape characteristics, bird and plant assemblages as drivers of Mediterranean ecosystem functioning. Landsc Ecol 38, 3605–3619 (2023). https://doi.org/10.1007/s10980-023-01739-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-023-01739-2