
Abstract
Purpose
In this dose-finding Phase II study (NCT00621322), we evaluated the safety and immunogenicity of different formulations of the candidate tuberculosis vaccine containing the M72 antigen (10/20/40 μg doses) and the liposome-based AS01 Adjuvant System. We aimed to select the lowest-dose combination of M72 and AS01 that was clinically well tolerated with immunogenicity comparable to that of the previously tested M72/AS01B (40 μg) candidate vaccine.
Methods
Healthy PPD-positive (induration 3–10 mm) adults (18–45 years) in The Philippines were randomized (4:4:4:4:1:1) to receive 2 injections, 1 month apart, of M72/AS01B (40 μg), M72/AS01E (10 μg), M72/AS01E (20 μg), M72/AS02D (10 μg), M72/Saline (40 μg) or AS01B alone, and were followed up for 6 months. AS01E and AS02D contain half the quantities of the immunostimulants present in AS01B. AS02D is an oil-in-water emulsion. Vaccine selection was based on the CD4+ T-cell responses at 1 month post vaccination.
Results
All formulations had a clinically acceptable safety profile with no vaccine-related serious adverse events reported. Two vaccinations of each adjuvanted M72 vaccine induced M72-specific CD4+ T-cell and humoral responses persisting at 6 months post vaccination. No responses were observed with AS01B alone. One month post second vaccination, CD4+ T-cell responses induced by each of the three M72/AS01 vaccine formulations were of comparable magnitudes, and all were significantly higher than those induced by M72/AS02D (10 μg) and M72/Saline.
Conclusions
The formulation with the lowest antigen and adjuvant dose, M72/AS01E (10 μg), fulfilled our pre-defined selection criteria and has been selected for further clinical development.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Tuberculosis (TB) is the second leading cause of mortality worldwide, resulting in 1.4 million deaths and 8.7 million incident cases in 2011 [1]. The only licensed TB vaccine, Bacille Calmette-Guérin (BCG) confers protection against meningeal and disseminated TB in children [2]. This vaccine has however a poor efficacy in preventing adult pulmonary disease [3], which is the foremost source of Mycobacterium tuberculosis (Mtb) transmission and also the primary cause of death [4]. Moreover, the efficacy of BCG wanes over time [5–7], highlighting the need to develop a highly efficacious and safe TB vaccine.
Several novel prophylactic vaccines are being developed to prevent TB disease [8], including the adjuvanted M72 candidate vaccine [9]. Initial development of the latter vaccine started with the adjuvanted Mtb72F vaccine, which is composed of the Mtb72F antigen formulated with one of two different Adjuvant Systems (the liposome-based AS01 or the oil-in-water emulsion-based AS02 [10, 11]). Mtb72F is a recombinant fusion protein [12], derived from the Mtb proteins Mtb32A (encoded by Rv0125) [13] and Mtb39A (encoded by Rv1196) [14]. Both AS01 and AS02 contain the immunostimulants 3-O-desacyl-4′-monophosphoryl lipid A (MPL) and Quillaja saponaria Molina fraction 21 (QS-21) and are known to induce T-helper cell 1 (Th1)-type responses [10, 11]. Mtb72F combined with AS02A had a favourable safety and immunogenicity profile in Purified Protein Derivative (PPD)-negative and PPD-positive adults [15–17]. The Mtb72F antigen was improved to generate the current vaccine antigen, M72, and a subsequent clinical study showed that 40 μg doses of M72 in either AS01B or AS02A were equally well tolerated in PPD-negative adults [9]. Both candidate vaccines elicited robust M72-specific humoral and CD4+ T-cell responses persisting for up to 3 years, however M72/AS01B was significantly more immunogenic than M72/AS02A in terms of CD4+ T-cell responses. We hypothesize that in a PPD-positive population, a lower-dose formulation of M72/AS01 may be sufficient to induce robust immune responses, thus allowing for a decrease of the antigen and/or adjuvant doses without compromising vaccine immunogenicity.
To test this hypothesis and to select the optimal candidate vaccine formulation, we conducted a Phase II antigen and adjuvant dose-finding study with different formulations of M72/AS01 in healthy adults living in a region with a high TB burden. Two variants of AS01 (AS01B and AS01E) and one variant of AS02 (AS02D) have been tested. Per dose injected, AS01E and AS02D both contain half the quantities of immunostimulants that are present in AS01B, as detailed in the “Methods” section. From the M72/AS01 candidate vaccines tested in this study, we aimed to select the lowest-dose formulation that had an acceptable safety profile and immunogenicity comparable to M72/AS01B (40 μg) candidate vaccine. We therefore evaluated lower quantities of the M72 antigen (10 and 20 μg) in AS01E, using an AS02D-adjuvanted and a non-adjuvanted M72 vaccine as comparators. The tested formulations thus included M72/AS01B (40 μg), M72/AS01E (10 μg), M72/AS01E (20 μg), M72/AS02D (10 μg), M72/Saline (40 μg) and AS01B alone (control).
Methods
Study Design and Ethics
This Phase I/II, observer blind, randomised, single center, controlled trial was conducted between April 2008 and April 2009 at the City Health Office I, Santa Rosa City, Laguna, The Philippines (www.clinicaltrials.gov NCT00621322). The study protocol and informed consent forms were reviewed and approved prior to initiation of the study by the National Ethics Committee, Philippine Council for Health Research and Development. Written informed consent was obtained from each subject prior to enrollment. The study was conducted in accordance with the Declaration of Helsinki and Good Clinical Practice.
Eligible subjects were randomized (4:4:4:4:1:1) to receive M72/AS01B (40 μg), M72/AS01E (10 μg), M72/AS01E (20 μg), M72/AS02D (10 μg), M72/Saline (40 μg) or AS01B alone. A randomization list generated with standard statistical analysis software (SAS Institute Inc., Cary, NC, USA) was used to number the vaccines, with a block randomization scheme to ensure a balanced allocation between groups. All groups received 2 injections of study vaccine at a one-month interval (Days 0 and 30), and were followed for up to 6 months after the second vaccination (Day 210).
Vaccination was staggered in two sets and each set was observer-blind (i.e., the subject and site or sponsor staff involved in the evaluation of subjects were blinded, whilst staff involved in vaccine administration were aware of the treatment). Subjects of the first set were randomized to receive study vaccine containing the lowest antigen quantities used in this study (M72/AS01E (10 μg) and M72/AS02D (10 μg)) or AS01B alone. The second set (M72/AS01E (20 μg), M72/AS01B (40 μg) and M72/Saline (40 μg) groups) was vaccinated if no safety signals were observed post vaccination of Set 1 subjects.
Study Participants
Healthy subjects were eligible if they were PPD-positive (PPD induration ≥ 3 mm but ≤ 10 mm 48–72 hours after administration of the PPD skin test; Tuberculin PPD RT23 SSI, Statens Serum Institute), HIV-negative, aged between 18 and 45 years, had clinically normal screening laboratory values, no history of TB disease and no sign of active TB disease on chest X-ray. The selection of a skin test induration between 3 and 10 mm was made using the assumption that the test is positive maybe due to an infection with a non-tuberculous mycobacterium and/or a previous BCG vaccination, and not to an infection by Mtb. Histories of BCG vaccination were not collected. Subjects were excluded if they had a history of any acute or chronic illness, or of medication with the potential to interfere with the safety and immunogenicity evaluations. Pregnancy and lactation were exclusion criteria.
Study Vaccines
The M72 antigen has been described previously [9]. Each lyophilised cake of M72 antigen was reconstituted with the appropriate liquid adjuvant or saline. AS01 and AS02 both contain the immunostimulants QS-21 (Antigenics Inc., a wholly owned subsidiary of Agenus Inc., Lexington, MA, USA) and MPL, either combined with liposomes (AS01) or in an oil-in-water emulsion (AS02) [10]. AS01B contained 50 μg MPL and 50 μg QS-21 per injection, and AS01E and AS02D both contained 25 μg MPL and 25 μg QS-21 per injection. Injected volumes were 0.5 mL for all study vaccines, except for M72/AS01E (20 μg) which was a half of the injected volume of M72/AS01B (40 μg), i.e., 0.25 mL. The vaccines were administered by intramuscular injection with a 23-gauge needle in the deltoid muscle.
Study Objectives
The primary objective was to evaluate the safety and reactogenicity of the M72 candidate vaccines in the targeted PPD-positive population. Secondary objectives were to evaluate cell-mediated and humoral immune responses.
Safety Assessment
Safety was assessed by daily recording of solicited local adverse events (AEs) (pain, redness and swelling) and general AEs (fatigue, fever [axillary temperature ≥37.5°C], gastrointestinal (GI) symptoms, headache, malaise and myalgia) for 7 days after each dose (Days 0–6). Unsolicited AEs occurring within 30 days after each dose and serious AEs (SAEs) until study end were also recorded. Biochemical and haematological parameters (complete blood count, renal and liver function tests) were monitored on the day of each vaccination (Days 0 and 30), on Day 7 after each dose (Days 7 and 37) and 30 days after Dose 2 (Day 60). All solicited local AEs were considered vaccination-related; the relationship of all other AEs to vaccination was determined by the Investigator. AE intensities were scored, and grade 3 (severe) AEs were defined as those preventing normal activity, redness or swelling >50 mm in diameter, or an axillary temperature >39.5°C.
Within Set 1, all safety and reactogenicity data collected for 6 days following vaccination were reviewed in a blinded manner by the Investigator group and a sponsor Safety Review Team to see if pre-defined holding/suspension criteria were met, prior to vaccination of Set 2. Vaccination would be put on hold pending unblinded safety review by an independent sponsor Vaccine Safety Monitoring Board if more than 8 subjects were withdrawn for severe or unexpected AEs judged to be related to vaccination 1 week after the last subject was vaccinated, in the event of a death or life-threatening SAE judged to be related to vaccination, or if a subject experienced anaphylactic shock following vaccination.
Immunological Assessment Time-Points
Blood samples were collected prior to each vaccination (Days 0 and 30) and 1 and 6 month(s) after the second vaccination (Days 60 and 210, respectively). Laboratory assessments were conducted in a blinded manner.
Humoral Responses
M72-specific IgG antibodies were measured by enzyme-linked immunosorbent assay (ELISA) based on a previously described method [9] with an assay cut-off of 2.8 ELISA units (EU)/ml (i.e., the lower limit of quantitation).
T-Cell Responses Assessed by Intracellular Cytokine Staining (ICS) and Flow Cytometry
M72 or PPD-specific CD4+ and CD8+ T cells expressing the immune markers IFN-γ and/or IL-2 and/or TNF-α and/or CD40L were detected by ICS upon short-term in vitro stimulation, based on a previously described methodology [18]. We used an adaptation of the method described by Maecker et al. [19, 20], in which PBMCs were stimulated ex vivo by incubation with antigen in the presence of costimulatory antibodies to CD28 and CD49d, and of Brefeldin A to inhibit cytokine secretion and allow intracellular accumulation. PBMCs were then stained using fluorochrome-conjugated antibodies before enumeration by flow cytometry.
To measure the M72-specific T-cell responses, PBMC were stimulated with a pool of 15-mer peptides (Eurogentech s.a., Seraing, Belgium; final concentration 1.25 μg/ml of each peptide) overlapping by 11 amino acids and spanning the entire sequence of the M72 antigen. The peptides were shown to have >80 % purity by high-performance liquid chromatography. Lyophilized peptides were reconstituted in phosphate buffered saline (PBS)/dimethylsulfoxide (<0.1 % final concentration). For assessment of PPD-specific T-cell responses, PBMC were stimulated with PPD (10 μg/ml; Staten Serum Institut, Denmark).
For the M72/AS01E (10 μg) vaccine group, an additional ICS assay was done. Th1/Th2 cytokine profiles (at Days 0 and 60 only) were characterized by measuring frequencies of CD4+ T cells expressing IFN-γ and/or IL-2 and/or TNF-α and/or CD40L and/or IL-13, following stimulation of PBMC with pools of overlapping 15-mer peptides of either 10-kDa culture filtrate protein (CFP-10), early secreted antigenic target (ESAT-6) (both 1.25 μg/ml; Eurogentec s.a., Seraing, Belgium), or M72 (1.25 μg/ml), or with PPD (10 μg/ml).
Cell Stimulation and Staining
Purified PBMC were thawed, washed twice in culture medium (RPMI 1640, Cambrex, East Rutherford, NJ, USA), supplemented with 10 % heat-inactivated fetal calf serum (FCS; PAA Laboratories GMbH, Austria), 100 IU/ml penicillin, 100 μg/ml streptomycin sulfate, 2 mM L-glutamine, MEM nonessential amino acids, 100 mM sodium pyruvate, 50 mM 2-mercapto-ethanol (all Life Technologies, Belgium), examined for viability and counted (Trucount, BD Biosciences, San Jose, CA USA), washed again, and resuspended to 2 × 107 cells/ml in culture medium. PBMC (1 × 106 cells per well) were incubated in 96-well microtiter plates with unconjugated and azide-free costimulatory anti-human CD28 and CD49d antibodies (1/250 dilution each) and stimulated for 20 h at 37 °C with the appropriate antigen. Brefeldin A (BD Pharmingen, San Diego, CA, USA; final concentration 1 μg/ml) was added for the last 18 h of culture. Positive (Staphylococcal enterotoxin B (SEB), 1 μg/ml; Sigma-Aldrich, St. Louis, MO, USA) and negative controls (unstimulated; no antigen) were included in each assay. PMA/ionomycin-stimulated CD4+ T cells were used as a positive control for IL-13 staining.
Following incubation, the cells were washed (in PBS containing 1 % FCS) and stained with anti-CD4-PerCP and anti-CD8-APC-Hi7 (all BD Pharmingen). The cells were then washed again, fixed, and permeabilized with the Cytofix/Cytoperm kit (BD Pharmingen) according to instructions and stained with anti CD3-AF700, anti-IFN-γ-FITC, anti-IL-2-APC, anti-TNF-α-PE Cy7 and anti-CD40L-PE (all BD Pharmingen). Following washing (Perm/Wash buffer, BD Pharmingen), the cells were analyzed by flow cytometry.
The same protocol and antibodies were used for the characterization of Th1/Th2-expressing T cells, with the exception that the intracellular staining was done with anti-CD3 PO (Caltag-Medsystems Ltd, UK), anti-IFN-γ-AF 700 and anti-IL-2-FITC (both BD Pharmingen) instead of anti CD3-AF700, anti-IFN-γ-FITC and anti-IL-2-APC, respectively, and that anti-IL-13-APC (Biolegend, San Diego, CA, USA) was used.
Flow Cytometry
Cells were acquired on a FACSCanto II flow cytometer or a LSR II flow cytometer (Becton Dickinson) using seven-color panels. Data were analyzed using FlowJo software (TreeStar, San Carlos, CA). Background (unstimulated control) was subtracted from all values. The remaining positive events were regarded as significant. Samples were only included for analysis if viability was ≥80 %.
Statistical Methods
Safety Evaluation
The evaluation of safety was performed on the Total Vaccinated Cohort, which included all subjects with at least one vaccine administration documented.
Descriptive statistics were performed for the percentage of doses followed by at least one solicited AE (local or general) with exact 95 % confidence interval (CI), and for the proportion of subjects reporting an unsolicited AE, classified by the MedDRA-preferred term level, with exact 95 % CI. Similar analyses were conducted for grade 3 AEs and for the AEs considered to be related to vaccination. Any reported SAEs were described. Any biochemistry and/or haematology values outside of the predefined reference ranges were assessed for clinical significance.
Immunogenicity Evaluation
Immunogenicity analysis was performed on the According to Protocol (ATP) cohort, i.e., all subjects meeting all eligibility criteria, complying with protocol defined procedures, with no elimination criteria and for whom data concerning immunogenicity endpoint measures were available.
Subjects were considered to be seropositive if their anti-M72 IgG antibody concentrations were ≥2.8 EU/mL. Seronegative subjects were given an arbitrary value of half the cut-off. Anti-M72 seropositivity rates and geometric mean concentrations (GMCs) were calculated with a 95 % CI as described previously [9].
Descriptive statistics of the CD4+ and CD8+ T-cell frequencies and serum levels of IgG antibodies were performed at each time-point using SAS version 8.2.
In the ICS assays, results were evaluated as the background-subtracted percentages of antigen-specific CD4+ and CD8+ T cells, identified as expressing any or a combination of IFN-γ and/or IL-2 and/or TNF-α and/or CD40L, or expressing any or a combination of these 4 markers and/or IL-13, upon short term in vitro stimulation. Total immune markers-expressing CD4+ and CD8+ T-cell responses were calculated by summation of the frequencies of the phenotypes expressing one, two, three, four, or five of the above immune markers upon in vitro stimulation over the background level.
A comparison of the magnitudes of vaccine-induced M72-specific responses between groups was made by evaluating for each group the proportions of subjects that responded to vaccination. This was performed descriptively by calculating at each post-vaccination time-point and for each subject the fold increases (at least 2-fold, at least 4-fold, at least 6-fold and at least 8-fold) in the frequency of M72-specific CD4+ T cells expressing at least 2 immune markers (among CD40L, IL-2, IFN-γ and TNF-α) over the frequency of these cells at pre-vaccination.
The selection of the candidate vaccine for further evaluation was based on statistical comparisons of M72-specific CD4+ T-cell responses expressing at least two immune markers (among IL-2, TNF-α, IFN-γ and CD40L) between all groups at Day 60. The expression by CD4+ T cells of at least 2 markers (instead of one) has been used as selection criterion to increase the sensitivity of the assay, allowing for a lower cut-off [21–23].
The frequencies of M72-specific CD4+ T cells were compared pair-wise between vaccine groups using a nonparametric ANOVA on the ranks (Kruskal-Wallis test) without correction for multiplicity. A sample size of 40 subjects per group for the adjuvanted vaccines provided a power of 82 % to detect at least one formulation similar to M72/AS01B (40 μg). The Wilcoxon signed rank test was used to compare ICS results at different time-points within a given vaccine group. Each two treatment groups or time-points compared were considered significantly different if p ≤ 0.05.
Results
Demographic Characteristics
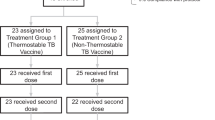
A total of 180 subjects were enrolled and randomly assigned to a study group, of which 169 completed the 6 months follow up (Fig. 1). Of the 11 subjects who withdrew from the study, none withdrew due to an adverse event (AE).

Participant flow. CONSORT diagram of study flow. ATP According to Protocol Cohort, N number of subjects who received the vaccine and for whom data are available. Boxes with dotted-lines represent subjects excluded from the ATP immunogenicity cohort
At Month 0, the demographic profile of subjects across groups was balanced in terms of age and gender. The mean age of subjects was 31.8 years (standard deviation 8.8 years) and the majority of subjects were female (142/180; 78.9 %).
Reactogenicity and Safety
Pain at the injection site was the most frequently reported solicited local AE in all groups (Fig. 2a), occurring with a similar frequency across all adjuvanted M72 vaccine groups and the AS01B alone group. No grade 3 solicited local AEs were reported in the M72/AS01E (10 μg), M72/Saline (40 μg) and AS01B groups; the incidence in all other groups was low (after ≤2.5 % of doses, Fig. 2a). All grade 3 solicited local AEs resolved or reduced in intensity after 2 days. The frequency of solicited local AEs did not increase from the first to the second injection of any of the study vaccines (data not shown).

Percentage of doses followed by solicited local and general adverse events within 7 days after each dose (Total vaccinated cohort). Mean percentage of doses (with 95 % confidence interval) followed by solicited local (a) and general (b) adverse events (AEs) reported within 7 days post-vaccination are shown for all subjects for whom at least one vaccine administration was documented. Gr.3, Grade 3; GI sympt, gastro-intestinal symptom
Headache, malaise and myalgia were the most frequently reported solicited general AEs in all groups (Fig. 2b). Symptoms of grade 3 intensity were infrequent across groups, occurring after ≤6.3 % of vaccine administrations. All grade 3 general AEs resolved or reduced in intensity after 1 day. There was a trend towards a higher incidence of solicited general AEs in the M72/AS01B (40 μg) group.
The percentages of subjects reporting at least one unsolicited AE were comparable across groups (Table 1). Nasopharyngitis, cough and headache were the most commonly reported unsolicited AEs in all groups. The incidence of subjects reporting unsolicited AEs with causal relationship to vaccination was infrequent (≤17.5 %) with no individual vaccine-related unsolicited AE being reported by more than one subject per group. Two unsolicited AEs were graded 3 in intensity: chills (related) in the M72/AS01E (10 μg) group and oropharyngeal pain (not related) in the M72/AS02D (10 μg) group.
Four subjects reported one SAE each during the study and none of these SAEs were considered to be related to vaccination. In the M72/AS01E (10 μg) group, one subject (with a family history of diabetes mellitus) developed type-2 diabetes mellitus, enteric fever and dengue fever 4 months after the second vaccination and was hospitalized, and a second subject developed bronchial asthma 78 days after the second vaccination. In the M72/AS01E (20 μg) group, one subject developed acute pyelonephritis 56 days after the second vaccination and was hospitalized, and a second subject developed food poisoning 68 days after the second vaccination. None of these SAEs were fatal and all resolved except for the diabetes mellitus, for which the subject is under treatment.
No vaccine-related clinically relevant changes in biochemistry or haematology parameters were observed.
Sixty-nine days after the second vaccination, one participant reported having become pregnant between the first and the second vaccination. She delivered a healthy infant after 35 weeks of pregnancy.
Immunogenicity Evaluations
Humoral Immune Responses Elicited by the Candidate Vaccines
All subjects in the four adjuvanted M72 vaccine groups were seronegative prior to vaccination, and after one vaccination (Day 30), seroconversion rates in the M72/AS01B (40 μg), M72/AS01E (10 μg), M72/AS01E (20 μg) and M72/AS02D (10 μg) groups were 100 %, 69.2 %, 90.0 % and 78.9 % of subjects, respectively. All of these subjects were seropositive 1 month after the second vaccination (Day 60). At study end (Day 210), all except one subject (in the M72/AS01E (10 μg) group) remained seropositive. Anti-M72 IgG antibody responses in these four groups were highest at Day 60, with comparable geometric mean concentrations (GMCs) between groups (Fig. 3). The responses persisted until study end at values that remained higher than at 30 days after the first vaccination. In the M72/Saline (40 μg) group, 80.0 % of subjects had seroconverted by Day 60, with a low GMC (31.0 EU/ml). No responses were observed in the AS01B alone group.

Anti-M72 IgG antibody responses. Blood samples were obtained prior to each vaccination (D0 and D30), and at 1 and 6 months post the second vaccination (D60 and D210). GMC geometric mean concentration, CI confidence interval
CD4+ T-Cell Responses Elicited by the Candidate Vaccines
T-cell responses induced by the candidate vaccines were evaluated by ICS and flow cytometry. PBMC were stimulated with either overlapping peptides covering the M72 sequence or PPD, and responding T cells were detected by the expression of CD40L, IFN-γ, TNF-α and/or IL-2. The gating strategy and representative scatterplots for the stimulation with M72 are presented in Fig. 4.

PBMC intracellular cytokine detection by flow cytometry. PBMC intracellular cytokine detection by flow cytometry was performed following overnight stimulation with Staphylococcal enterotoxin B (SEB; positive control), the M72 peptide pool or medium. a Cytokine production by M72-specific CD4+ T cells was determined by discriminating the CD4+ and CD8+ T cells from the “morphological” gating window. b CD4+ T cells were analyzed with respect to the production of CD40L, IL-2, TNF-α and IFN-γ. The unstimulated (medium) samples show background levels of cytokine production. When restimulated with the M72 peptide pool, the production of CD40L, IL-2 and TNF-α by CD4+ T cells and lower production of IFN-γ was detected. The numbers in the quadrant gates of the plots denominate each distinct population based on their cytokine production. Samples from the same subject are shown, with responses at 1 month post second vaccination. Results shown are representative of the range of responses seen with all subjects studied. The lowest, highest and median numbers of acquired viable CD4+ T cells examined were 16975, 75000 and 75000 cells respectively
The vaccine-induced M72-specific CD4+ T-cell responses were evaluated for cells expressing at least two immune markers (Fig. 5). Prior to vaccination (Day 0), M72-specific CD4+ T-cell responses were detected in a minority of subjects. No M72-specific responses were observed in the AS01B alone group throughout the study. M72-specific CD4+ T-cell responses were observed after one administration of each adjuvanted M72 vaccine or M72/Saline (40 μg), although the responses to the latter vaccine were of a lower magnitude. For the M72/AS01E (10 or 20 μg) and M72/AS02D (10 μg) vaccines, these responses were significantly boosted by a second vaccination (p < 0.001; Day 60 compared to Day 30; Wilcoxon signed rank test), which appeared not to be the case for the M72/AS01B (40 μg) and M72/Saline (40 μg) vaccines (p = 0.18 and p = 0.91, respectively).

Frequencies of M72-specific CD4+ T cells expressing at least two immune markers. Blood samples were obtained prior to each vaccination (D0 and D30), and at 1 and 6 months post the second vaccination (D60 and D210). Data are reported as the percentages of M72-specific CD4+ T cells expressing (after in vitro stimulation) at least two immune markers among IFN-γ, IL-2, TNF-α and CD40L of all CD4+ T cells, with first and third quartiles, and the minimum/maximum values measured. Magnitudes of the CD4+ T-cell responses at D210 versus D0 and at D60 versus D30 were compared using the Wilcoxon signed rank test. Values were considered significantly different if p ≤ 0.05, as indicated by * for p ≤ 0.05, ** for p < 0.01 or *** for p < 0.001
The M72-specific CD4+ T-cell responses after vaccination were also evaluated for each subject by calculating the fold increases over the frequency of these cells at pre-vaccination (Table 2). In the adjuvanted vaccine groups, there was a tendency for higher proportions of responders after the second vaccination (Day 60) as compared with after the first vaccination. In addition, the proportions of responders with 6-fold and 8-fold increases were comparable between the M72/AS01E (10 μg) and M72/AS01B (40 μg) groups, both at Day 60 and at Day 210, and the proportions in these two groups tended to be higher relative to those for the other groups.
Six months after the second vaccination, the responses elicited by the adjuvanted M72 vaccines, as well as by M72/Saline (40 μg), persisted at significantly higher magnitudes than at pre-vaccination (p < 0.001 and p = 0.016, respectively; Wilcoxon signed rank test; Fig. 5). A similar persistence in the responses was seen for the adjuvanted M72 vaccine groups when the M72-specific CD4+ T-cell responses were evaluated using the frequencies of cells expressing either at least one cytokine among IFN-γ, TNF-α and IL-2, or any of the four immune markers (p < 0.001 all time-points compared to Day 0; Wilcoxon signed rank test; Fig. 6). In the M72/Saline (40 μg) group such persistence was not consistently detected at Day 210.

Frequencies of total immune-marker expressing M72-specific CD4+ T cells. Blood samples were obtained prior to each vaccination (D0 and D30), and at 1 and 6 months post the second vaccination (D60 and D210). Data are reported as the percentages of M72-specific CD4+ T cells expressing any immune marker among IFN-γ, IL-2, TNF-α and CD40L (‘any marker’) or at least IL-2, TNF-α or IFN-γ among all CD4+ T cells, with first and third quartiles, and the minimum/maximum values measured. Magnitudes of the immune responses post vaccination (D30, D60 and D210) versus D0 were compared using the Wilcoxon signed rank test. Values were considered significantly different if p ≤ 0.05, as indicated by * for p ≤ 0.05, ** for p < 0.01 or *** for p < 0.001
The CD4+ T-cell responses elicited by the adjuvanted M72 vaccines comprised predominantly CD40L+ IL-2+ IFN-γ+ TNF-α+, CD40L+ IL-2+ TNF-α+ and CD40L+ IL-2+ expression profiles, with lower frequencies of single-positive CD40L+ and of CD40L+ IL-2+ IFN-γ+ profiles (Fig. 7a). At Day 60, the majority (83–88 %) of responding CD4+ T cells expressed at least two immune markers simultaneously, and these responses persisted up to Day 210 (Fig. 7b).

Polyfunctional profiles of M72-specific CD4+ T cells expressing any combination of immune markers elicited by the adjuvanted M72 vaccine groups. Blood samples were obtained prior to each vaccination (D0 and D30), and at 1 and 6 months post the second vaccination (D60 and D210). a Phenotypes of M72-specific CD4+ T cells expressing (after in vitro stimulation) single markers and any combination of IFN-γ, IL-2, TNF-α and CD40L. Box-and-whiskers plots represent the percentages of M72-specific CD4+ T cells, with first and third quartiles, and the minimum/maximum values measured. b Pie charts represent the mean proportions of cells expressing (after in vitro stimulation) single markers and any combination of IFN-γ, IL-2, TNF-α and CD40L marker-positive CD4+ T cells out of the total immune-marker expressing CD4+ T-cell response, at Days 60 and 210
Pre-existing PPD-specific CD4+ T cells were observed in all groups, but no vaccine-induced increase in the frequencies of these cells was observed (Table 3).
No vaccine-induced M72-specific CD8+ T-cell responses were detected for any of the formulations at the time-points measured (data not shown).
Selection of the Candidate Vaccine for Further Clinical Evaluation
Vaccine selection was based on the comparison of the frequencies of M72-specific CD4+ T cells expressing at least 2 immune markers at Day 60, with the aim to select the lowest-dose formulation of M72/AS01 with responses comparable to that induced by M72/AS01B (40 μg). There were no significant differences between the responses in the three M72/AS01 vaccine groups (Table 4), and in each of these groups the responses were significantly higher than in the M72/AS02D (10 μg) group. The responses in the M72/Saline (40 μg) and AS01B alone groups were significantly lower relative to all other groups. Based on its acceptable safety profile and on the CD4+ T-cell data described above, M72/AS01E (10 μg) was selected.
Further Characterization of the M72/AS01E (10 μg) Candidate Vaccine
In order to assess the Th1/Th2 bias of the M72-specific responses induced by the M72/AS01E (10 μg) vaccine, CD4+ T cells expressing either any immune marker (amongst CD40L, IFN-γ, TNF-α, IL-2 and IL-13) or at least one cytokine, were evaluated at Day 60. No M72-specific IL-13+ CD4+ T-cell responses were detected (Fig. 8).

Th1/Th2 cytokine expression by M72-specific CD4+ T cells induced by the M72/AS01E (10 μg) vaccine. Blood samples were obtained prior to vaccination (D0) and at 1 month post the second vaccination (D60). Data are presented as frequencies of M72-specific CD4+ T cells expressing any immune marker among CD40L, IFN-γ, IL-2, TNF-α and IL-13 (‘total’) or at least IFN-γ, TNF-α, IL-2 or IL-13 among all CD4+ T cells, with first and third quartiles, and the minimum/maximum values measured
No vaccine-induced PPD, ESAT-6 or CFP-10-specific CD4+ T-cell responses were observed (Table 5).
Discussion
This dose-finding study in PPD-positive adults (induration 3–10 mm) was aimed at evaluating and selecting the lowest-dose formulation of the candidate vaccines tested, that was well tolerated and with immunogenicity comparable to that of the M72/AS01B (40 μg) candidate vaccine. The selection of the study population was driven by the expectation that a major portion of the targeted population for this candidate vaccine would have a PPD-positive status.
All three AS01-adjuvanted vaccines investigated (M72/AS01B (40 μg), M72/AS01E (10 μg) and M72/AS01E (20 μg)) had comparable safety and reactogenicity profiles, which were similar to that seen previously in PPD-negative adults with M72/AS02A and M72/AS01B vaccines (both with 40 μg doses) [9]. For the induced M72-specific CD4+ T-cell responses, we did not observe a dose–response relationship over the three M72 doses and two AS01 formulations tested, and the responses were of similar magnitudes and durable. The lowest-dose vaccine, M72/AS01E (10 μg), was thus selected for further development. It is noted that this Phase I/II study was not designed as a formal equivalence trial for the vaccine formulations tested, as this would have required a larger sample size and an upfront specification of criteria that were unknown at the time of the study design. The selection of M72/AS01E (10 μg) was supported by other evaluations, showing that polyfunctional and persistent CD4+ T-cell responses and strong humoral responses were induced by each of the M72/AS01 vaccines. An additional evaluation of M72/AS01 (10 μg) demonstrated the absence of IL-13+ M72-specific CD4+ T-cell responses. No antigen-specific CD8+ T-cell responses were detected, in line with previous observations with an M72/AS01 vaccine formulation [9].
The data confirmed several outcomes of a previous M72 vaccine study [9]. First, the adjuvant effect of AS01 appeared to be stronger relative to that of AS02 in improving the cell-mediated immunogenicity. This is demonstrated by the comparison of the responses to the two 10 μg-dose vaccines, whereby the AS01E-adjuvanted vaccine induced significantly higher CD4+ T-cell responses than the AS02D-adjuvanted vaccine. This supports previous observations with M72/AS01 and M72/AS02 vaccines [9], as well as other comparative vaccine studies with different antigens (reviewed in [10, 11]). Secondly, the need for vaccine adjuvantation was demonstrated by the significantly higher antigen-specific CD4+ T-cell responses elicited by each of the AS01-adjuvanted M72 vaccines relative to the non-adjuvanted M72 vaccine. Lastly, as also observed in PPD-negative adults, the induced M72-specific CD4+ T-cell and humoral responses were increased by the second vaccination. This is in line with the observed proportions of subjects responding to the adjuvanted vaccines (with regard to CD4+ T-cell and humoral responses) at Days 30 and 60. Taken together, these results suggest that two vaccinations could also be needed in a PPD-positive population.
All study vaccines were clinically well tolerated. Local reactogenicity was reported with a high and similar frequency in recipients of adjuvanted vaccine or adjuvant alone, though symptoms were mainly mild to moderate in intensity. Solicited systemic AEs were reported more frequently after the second dose of an adjuvanted vaccine although the trend was not observed with Grade 3 AEs which were infrequently reported. A similar unsolicited AE profile was observed across vaccine groups. No safety signal was observed for any of the vaccines tested.
Adaptive cellular immunity is considered to be essential for protection against TB and, based on preclinical studies, it has been postulated that protection is mediated by sustained production of Th1 cytokines by polyfunctional CD4+ T cells [24–27]. Indeed, vaccine-induced antigen-specific polyfunctional CD4+ T cells co-expressing IL-2, IFN-γ and TNF-α were associated with protection against Mtb in several preclinical challenge models with other antigens [28–32]. Polyfunctionality (i.e., the expression of at least 2 immune markers) of vaccine-induced CD4+ T cells has also been demonstrated in Mtb72F or M72 vaccine studies in PPD-negative and PPD-positive adults [9, 15–17], and in studies with other TB candidate vaccines in mycobacteria-exposed, BCG-vaccinated and/or Mtb-infected individuals [33–39]. However, reliable biomarkers of protection in humans have been incompletely defined [40], and, adding to the complexity, new clinical data have emerged that may question the potential link between a polyfunctional CD4+ T-cell profile and BCG-induced or natural immunity to TB [41–43]. Nevertheless, the immunogenicity profiles elicited by adjuvanted M72 vaccines in humans [9, 15–17] are encouraging.
In TB-endemic settings, a pre-existing mixed Th1/Th2 response to mycobacterial antigens may exist due to exposure to non-tuberculous Mycobacteria [44–47]. In addition, neonatal BCG vaccination has been shown to be able to initially induce both Th1 and Th2-type responses to mycobacterial antigens [48–51], however the BCG-induced Th2 responses were shown to wane to pre-vaccination levels within 2 years post vaccination [50]. There are preclinical data suggesting that Th2-type responses (IL-4, IL-5 and IL-13) are able to suppress the development of Th1-mediated immunity by down-regulation of IFN-γ and TNF-α [52, 53]. In recent preclinical challenge studies [54, 55], it has been suggested that a stronger Th1 versus Th2 immunity was needed to limit the mycobacterium growth, and in humans Th2-type responses are known to correlate with TB immunopathology [47]. In our study, the vaccine-induced CD4+ T-cell responses were strongly Th1-biased.
The trends observed in the IgG antibody responses did not follow the same pattern as the CMI responses. CD4+ T-cell responses were higher with M72/AS01 as compared with M72/AS02, but all adjuvanted vaccines induced similar humoral responses. Some preclinical studies suggest that antibody responses to Mtb antigens have a protective role, e.g., by impairing the extra-pulmonary dissemination of Mtb [56] or by antibody-mediated enhancement of the uptake of mycobacteria by macrophages (reviewed in Abebe and Bjune (2009) [57]). It is not known if, and to which extent, humoral immunity contributes to protection against TB in humans.
Conclusions
The safety and immunogenicity profiles described for the three formulations of the M72/AS01 candidate vaccine support the selection of M72/AS01E (10 μg) for further clinical development.
References
World Health Organization. Global Tuberculosis Control: WHO report 2012. Geneva, Switzerland: WHO; 2012. Report No.: WHO/HTM/TB/2012.6.
Trunz BB, Fine P, Dye C. Effect of BCG vaccination on childhood tuberculous meningitis and miliary tuberculosis worldwide: a meta-analysis and assessment of cost-effectiveness. Lancet. 2006;367:1173–80.
Fine PE. Variation in protection by BCG: implications of and for heterologous immunity. Lancet. 1995;346:1339–45.
Chan ED, Iseman MD. Current medical treatment for tuberculosis. BMJ. 2002;325:1282–6.
Weir RE, Gorak-Stolinska P, Floyd S, Lalor MK, Stenson S, Branson K, et al. Persistence of the immune response induced by BCG vaccination. BMC Infect Dis. 2008;8:9.
Sterne JA, Rodrigues LC, Guedes IN. Does the efficacy of BCG decline with time since vaccination? Int J Tuberc Lung Dis. 1998;2:200–7.
Anuradha B, Santosh CM, Hari Sai Priya V, Suman Latha G, Murthy KJ, Vijaya Lakshmi V. Age-related waning of in vitro interferon-γ levels against r32kDaBCG in BCG vaccinated children. J Immune Based Ther Vaccines. 2007;5:8.
Ottenhoff TH, Kaufmann SH. Vaccines against tuberculosis: where are we and where do we need to go? PLoS Pathog. 2012;8:e1002607.
Leroux-Roels I, Forgus S, De Boever F, Clement F, Demoitié M-A, Mettens P, et al. the M72 study group. Improved CD4+ T cell responses to Mycobacterium tuberculosis in PPD-negative adults by M72/AS01 as compared to the M72/AS02 and Mbt72F/AS02 tuberculosis candidate vaccine formulations: a randomised trial. Vaccine. 2013;31:2196–206.
Garçon N, Chomez P, Van Mechelen M. GlaxoSmithKline Adjuvant Systems in vaccines: concepts, achievements and perspectives. Expert Rev Vaccines. 2007;6:723–39.
Garçon N, Van Mechelen M. Recent clinical experience with vaccines using MPL- and QS-21-containing Adjuvant Systems. Expert Rev Vaccines. 2011;10:471–86.
Skeiky YA, Alderson MR, Ovendale PJ, Guderian JA, Brandt L, Dillon DC, et al. Differential immune responses and protective efficacy induced by components of a tuberculosis polyprotein vaccine, Mtb72F, delivered as naked DNA or recombinant protein. J Immunol. 2004;172:7618–28.
Skeiky YA, Lodes MJ, Guderian JA, Mohamath R, Bement T, Alderson MR, et al. Cloning, expression, and immunological evaluation of two putative secreted serine protease antigens of Mycobacterium tuberculosis. Infect Immun. 1999;67:3998–4007.
Dillon DC, Alderson MR, Day CH, Lewinsohn DM, Coler R, Bement T, et al. Molecular characterization and human T-cell responses to a member of a novel Mycobacterium tuberculosis mtb39 gene family. Infect Immun. 1999;67:2941–50.
Leroux-Roels I, Leroux-Roels G, Ofori-Anyinam O, Moris P, De Kock E, Clement F, et al. Evaluation of the safety and immunogenicity of two antigen concentrations of the Mtb72F/AS02A candidate tuberculosis vaccine in Purified Protein Derivative-negative adults. Clin Vaccine Immunol. 2010;17:1763–71.
Spertini F, Audran R, Lurati F, Ofori-Anyinam O, Zysset F, Vandepapelière P, et al. The candidate tuberculosis vaccine Mtb72F/AS02 in PPD positive adults: a randomized controlled phase I/II study. Tuberculosis (Edinb). 2013;93:179–88.
Von Eschen K, Morrison R, Braun M, Ofori-Anyinam O, De Kock E, Pavithran P, et al. The candidate tuberculosis vaccine Mtb72F/AS02A: tolerability and immunogenicity in humans. Hum Vaccin. 2009;5:475–82.
Moris P, van der Most R, Leroux-Roels I, Clement F, Dramé M, Hanon E, et al. H5N1 influenza vaccine formulated with AS03A induces strong cross-reactive and polyfunctional CD4 T-cell responses. J Clin Immunol. 2011;31:443–54.
Maecker HT. Multiparameter flow cytometry monitoring of T cell responses. Methods Mol Biol. 2009;485:375–91.
Nomura L, Maino VC, Maecker HT. Standardization and optimization of multiparameter intracellular cytokine staining. Cytometry A. 2008;73:984–91.
Chattopadhyay PK, Yu J, Roederer M. A live-cell assay to detect antigen-specific CD4+ T cells with diverse cytokine profiles. Nat Med. 2005;11:1113–7.
Frentsch M, Arbach O, Kirchhoff D, Moewes B, Worm M, Rothe M, et al. Direct access to CD4+ T cells specific for defined antigens according to CD154 expression. Nat Med. 2005;11:1118–24.
Hammarlund E, Lewis MW, Hansen SG, Strelow LI, Nelson JA, Sexton GJ, et al. Duration of antiviral immunity after smallpox vaccination. Nat Med. 2003;9:1131–7.
Flynn JL. Immunology of tuberculosis and implications in vaccine development. Tuberculosis (Edinb). 2004;84:93–101.
Flynn JL, Chan J. Immunology of tuberculosis. Annu Rev Immunol. 2001;19:93–129.
Mueller H, Detjen AK, Schuck SD, Gutschmidt A, Wahn U, Magdorf K, et al. Mycobacterium tuberculosis-specific CD4+, IFNγ+, and TNFα+ multifunctional memory T cells coexpress GM-CSF. Cytokine. 2008;43:143–8.
Stenger S. Immunological control of tuberculosis: role of tumour necrosis factor and more. Ann Rheum Dis. 2005;64 Suppl 4:iv24–8.
Aagaard C, Hoang TT, Izzo A, Billeskov R, Troudt J, Arnett K, et al. Protection and polyfunctional T cells induced by Ag85B-TB10.4/IC31® against Mycobacterium tuberculosis is highly dependent on the antigen dose. PLoS One. 2009;4:e5930.
Aagaard CS, Hoang TT, Vingsbo-Lundberg C, Dietrich J, Andersen P. Quality and vaccine efficacy of CD4+ T cell responses directed to dominant and subdominant epitopes in ESAT-6 from Mycobacterium tuberculosis. J Immunol. 2009;183:2659–68.
Hoang TT, Nansen A, Roy S, Billeskov R, Aagaard C, Elvang T, et al. Distinct differences in the expansion and phenotype of TB10.4 specific CD8 and CD4 T cells after infection with Mycobacterium tuberculosis. PLoS One. 2009;4:e5928.
Lindenstrom T, Agger EM, Korsholm KS, Darrah PA, Aagaard C, Seder RA, et al. Tuberculosis subunit vaccination provides long-term protective immunity characterized by multifunctional CD4 memory T cells. J Immunol. 2009;182:8047–55.
Aagaard C, Hoang T, Dietrich J, Cardona P-J, Izzo A, Dolganov G, et al. A multistage tuberculosis vaccine that confers efficient protection before and after exposure. Nat Med. 2011;17:189–94.
Abel B, Tameris M, Mansoor N, Gelderbloem S, Hughes J, Abrahams D, et al. The novel tuberculosis vaccine, AERAS-402, induces robust and polyfunctional CD4+ and CD8+ T cells in adults. Am J Respir Crit Care Med. 2010;181:1407–17.
Beveridge NE, Price DA, Casazza JP, Pathan AA, Sander CR, Asher TE, et al. Immunisation with BCG and recombinant MVA85A induces long-lasting, polyfunctional Mycobacterium tuberculosis-specific CD4+ memory T lymphocyte populations. Eur J Immunol. 2007;37:3089–100.
Hawkridge T, Scriba TJ, Gelderbloem S, Smit E, Tameris M, Moyo S, et al. Safety and immunogenicity of a new tuberculosis vaccine, MVA85A, in healthy adults in South Africa. J Infect Dis. 2008;198:544–52.
Sander CR, Pathan AA, Beveridge NE, Poulton I, Minassian A, Alder N, et al. Safety and immunogenicity of a new tuberculosis vaccine, MVA85A, in Mycobacterium tuberculosis-infected individuals. Am J Respir Crit Care Med. 2009;179:724–33.
Scriba TJ, Tameris M, Mansoor N, Smit E, van der Merwe L, Isaacs F, et al. Modified vaccinia Ankara-expressing Ag85A, a novel tuberculosis vaccine, is safe in adolescents and children, and induces polyfunctional CD4+ T cells. Eur J Immunol. 2010;40:279–90.
Scriba TJ, Tameris M, Mansoor N, Smit E, van der Merwe L, Mauff K, et al. Dose-finding study of the novel tuberculosis vaccine, MVA85A, in healthy BCG-vaccinated infants. J Infect Dis. 2011;203:1832–43.
Scriba TJ, Tameris M, Smit E, van der Merwe L, Hughes EJ, Kadira B, et al. A phase IIa trial of the new tuberculosis vaccine, MVA85A, in HIV- and/or Mycobacterium tuberculosis-infected adults. Am J Respir Crit Care Med. 2012;185:769–78.
Parida SK, Kaufmann SH. The quest for biomarkers in tuberculosis. Drug Discov Today. 2010;15:148–57.
Young JM, Adetifa IM, Ota MO, Sutherland JS. Expanded polyfunctional T cell response to mycobacterial antigens in TB disease and contraction post-treatment. PLoS One. 2010;5:e11237.
Caccamo N, Guggino G, Joosten SA, Gelsomino G, Di Carlo P, Titone L, et al. Multifunctional CD4+ T cells correlate with active Mycobacterium tuberculosis infection. Eur J Immunol. 2010;40:2211–20.
Kagina BM, Abel B, Scriba TJ, Hughes EJ, Keyser A, Soares A, et al. Specific T cell frequency and cytokine expression profile do not correlate with protection against tuberculosis after bacillus Calmette-Guérin vaccination of newborns. Am J Respir Crit Care Med. 2010;182:1073–9.
Rook GA, Dheda K, Zumla A. Immune systems in developed and developing countries; implications for the design of vaccines that will work where BCG does not. Tuberculosis (Edinb). 2006;86:152–62.
Rook GA, Dheda K, Zumla A. Immune responses to tuberculosis in developing countries: implications for new vaccines. Nat Rev Immunol. 2005;5:661–7.
Rook GA, Dheda K, Zumla A. Do successful tuberculosis vaccines need to be immunoregulatory rather than merely Th1-boosting? Vaccine. 2005;23:2115–20.
Rook GAW. Th2 cytokines in susceptibility to tuberculosis. Curr Mol Med. 2007;7:327–37.
Lalor MK, Smith SG, Floyd S, Gorak-Stolinska P, Weir RE, Blitz R, et al. Complex cytokine profiles induced by BCG vaccination in UK infants. Vaccine. 2010;28:1635–41.
Finan C, Ota MO, Marchant A, Newport MJ. Natural variation in immune responses to neonatal Mycobacterium bovis Bacillus Calmette-Guerin (BCG) vaccination in a cohort of Gambian infants. PLoS One. 2008;3:e3485.
Djuardi Y, Sartono E, Wibowo H, Supali T, Yazdanbakhsh M. A longitudinal study of BCG vaccination in early childhood: the development of innate and adaptive immune responses. PLoS One. 2010;5:e14066.
Vekemans J, Amedei A, Ota MO, D’Elios MM, Goetghebuer T, Ismaili J, et al. Neonatal bacillus Calmette-Guérin vaccination induces adult-like IFN-γ production by CD4+ T lymphocytes. Eur J Immunol. 2001;31:1531–5.
Ruth JH, Warmington KS, Shang X, Lincoln P, Evanoff H, Kunkel SL, et al. Interleukin 4 and 13 participation in mycobacterial (type-1) and schistosomal (type-2) antigen-elicited pulmonary granuloma formation: multiparameter analysis of cellular recruitment, chemokine expression and cytokine networks. Cytokine. 2000;12:432–44.
Doherty TM, Kastelein R, Menon S, Andrade S, Coffman RL. Modulation of murine macrophage function by IL-13. J Immunol. 1993;151:7151–60.
Keyser A, Troudt JM, Taylor JL, Izzo AA. BCG sub-strains induce variable protection against virulent pulmonary Mycobacterium tuberculosis infection, with the capacity to drive Th2 immunity. Vaccine. 2011;29:9308–15.
Baldwin SL, Bertholet S, Reese VA, Ching LK, Reed SG, Coler RN. The importance of adjuvant formulation in the development of a tuberculosis vaccine. J Immunol. 2012;188:2189–97.
Pethe K, Alonso S, Biet F, Delogu G, Brennan MJ, Locht C, et al. The heparin-binding haemagglutinin of M. tuberculosis is required for extrapulmonary dissemination. Nature. 2001;412:190–4.
Abebe F, Bjune G. The protective role of antibody responses during Mycobacterium tuberculosis infection. Clin Exp Immunol. 2009;157:235–43.
Acknowledgments
The authors thank the participating volunteers, and acknowledge the contributions of the clinicians, nurses and laboratory technicians. In addition, they thank the technicians from the Research Institute for Tropical Medicine (Muntinlupa City, Philippines) for preparation of PBMC, the technicians from the Centre for Vaccinology (Ghent University and Hospital, Belgium), the Human Cellular Immunology group from GSK Vaccines for CMI evaluations, Ellen Oe (XPE Pharma&Science) and Sarah Benns (freelancer) for writing assistance on behalf of GSK Vaccines, and Sofia Dos Santos Mendes (XPE Pharma&Science on behalf of GSK Vaccines) and Ulrike Krause for editorial assistance. Finally, they thank Evi de Ruymaeker and Criselda Villegas for study management activities, Janani Murali and Priya Pavithran for writing of the clinical report and protocol, respectively, Edouard Ledent for statistical input on the protocol, and Didier Lapierre, Ripley W. Ballou, Pierre Vandepapelière and Hans Bock for helpful discussions.
Authors Contribution
All authors participated in the design or implementation or analysis and interpretation of the study; and the development of this manuscript. All authors had full access to the data and gave final approval before submission.
JM, JAS, PhM, MAD, PM, CV, JC and OOA were involved in study conception and design. JM was the coordinating investigator and together with JAS was responsible for the conduct of the trial. JM, JAS, SC, PhM, MJ, LA and SG collected data. JM, AB, PhM, MJ, EJ, MAD, PM, SG, CV, JC and OOA were involved in data analysis and/or interpretation of results. AB was the statistician on the team.
Conflicts of Interest
AB, PhM, MJ, EJ, MAD, PMe, SG, CV, JC and OO are or were employees of GlaxoSmithKline group of companies (GSK) at the time of the study. PhM, MAD, PMe, SG, CV, JC and OO own shares and options to shares in GSK. JC is a named inventor on several patents or patent applications related to the field of vaccine development. JM, JAS and LA received grants or consulting fees for operational expenses in the study conduct. All other authors report no potential conflicts.
GlaxoSmithKline Biologicals S.A. and Aeras Global TB Vaccine Foundation co-funded the study, and GlaxoSmithKline Biologicals S.A., as sponsor of the study, was involved in all stages of the study conduct and analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Montoya, J., Solon, J.A., Cunanan, S.R.C. et al. A Randomized, Controlled Dose-Finding Phase II Study of the M72/AS01 Candidate Tuberculosis Vaccine in Healthy PPD-Positive Adults. J Clin Immunol 33, 1360–1375 (2013). https://doi.org/10.1007/s10875-013-9949-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10875-013-9949-3