
Abstract
Understanding of the molecular mechanisms of protein function requires detailed insight into the conformational landscape accessible to the protein. Conformational changes can be crucial for biological processes, such as ligand binding, protein folding, and catalysis. NMR spectroscopy is exquisitely sensitive to such dynamic changes in protein conformations. In particular, Carr–Purcell–Meiboom–Gill (CPMG) relaxation dispersion experiments are a powerful tool to investigate protein dynamics on a millisecond time scale. CPMG experiments that probe the chemical shift modulation of 15N in-phase magnetization are particularly attractive, due to their high sensitivity. These experiments require high power 1H decoupling during the CPMG period to keep the 15N magnetization in-phase. Recently, an improved version of the in-phase 15N-CPMG experiment was introduced, offering greater ease of use by employing a single 1H decoupling power for all CPMG pulsing rates. In these experiments however, incomplete decoupling of off-resonance amide 1H spins introduces an artefactual dispersion of relaxation rates, the so-called slow-pulsing artifact. Here, we analyze the slow-pulsing artifact in detail and demonstrate that it can be suppressed through the use of composite pulse decoupling (CPD). We report the performances of various CPD schemes and show that CPD decoupling based on the 90x–240y–90x element results in high-quality dispersion curves free of artifacts, even for amides with high 1H offset.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Biological macromolecules such as nucleic acids and proteins are non-rigid entities that can populate a variety of conformers in their energy landscape (Frauenfelder et al. 1991; Wolynes 2005; Henzler-Wildman and Kern 2007). The lowest energy conformation, the ground state, is often able to transiently access higher-energy conformations. Even when their population is low (< 10%) and life times is short (~ ms), these excited states can be crucial for biologically important processes such as enzyme catalysis (Hammes 1964; Eisenmesser et al. 2005; Henzler-Wildman et al. 2007; Palmer 2015; Kim et al. 2017), ligand binding or protein–protein interactions (Sugase et al. 2007; Schneider et al. 2015; Pratihar et al. 2016; Xiao et al. 2016; Zhao et al. 2017; Delaforge et al. 2018), and protein folding (Korzhnev et al. 2010; Neudecker et al. 2012; Kimsey et al. 2015; Libich et al. 2015; Franco et al. 2017; Culik et al. 2018). While these states cannot be detected directly due to their transient and lowly populated nature, NMR experiments (Akke and Palmer 1996; Fawzi et al. 2010; Kovermann et al. 2016) are uniquely able to provide a detailed, atomistic description of the energy landscape. In particular, relaxation dispersion and chemical exchange saturation transfer experiments are particularly powerful herein, as they give access to the population, life times and structures of excited states (Palmer et al. 2001; Vallurupalli et al. 2012, 2017; Sauerwein and Hansen 2015; Xue et al. 2015; Lisi 2016; Massi and Peng 2018; Gopalan et al. 2018).
In Carr–Purcell–Meiboom–Gill (CPMG) relaxation dispersion experiments, the characterization of the minor state is derived from the major state peaks by measurement of their effective transverse relaxation rate R2,eff, as a function of the pulsing rate in the CPMG period. Signals of nuclear spins that experience exchange between states with different chemical shifts are affected by exchange-induced line broadening, an effect that depends on the free precession interval (2τcp) between the refocusing pulses in the CPMG element (Palmer et al. 2001; Sauerwein and Hansen 2015). Analysis of the resulting relaxation dispersion curve, a plot of the R2,eff versus CPMG frequency (1/4τcp), allows determination of the rate of exchange (kex), population of minor state (pb) and the absolute chemical shift difference (|Δϖ|) between the exchanging states. Importantly, since the shape of the dispersion profile depends on Δϖ, data is typically acquired at two fields to accurately determine the exchange parameters (Sauerwein and Hansen 2015).
The 15N backbone amide spin is the most popular nucleus for CPMG RD experiments, due to the simplicity of isotope-labeling, the straightforwardness of the two-spin 1H–15N spin system, and the high sensitivity and resolution afforded by these experiments. A critical aspect of these experiments is appropriate handling of differences in the intrinsic R2 of the in-phase (Nx,y) and anti-phase (2Nx,yHz) 15N magnetization which are generated in the free evolution periods. Anti-phase terms have higher intrinsic relaxation rates due to a contribution of 1H spin flips to their decay. The original implementation of the 15N CPMG RD experiment uses a relaxation-compensation scheme to average the Nx,y and 2Nx,yHz relaxation rates (Loria et al. 1999b). The 15N CPMG sequence of Hansen et al. (2008b) (CW–CPMG) measures the dispersion profile of pure in-phase Nx,y by applying high-power continuous wave (CW) 1H decoupling during the CPMG train, offering enhanced sensitivity for non-deuterated proteins. Recently, Jiang et al. (2015) modified this sequence (ST–CW–CPMG) to use a single CPMG train with the Yip and Zuiderweg phase cycle (2004) and a single CW decoupling power, yielding dispersion curves free of off-resonance artifacts for a wider range of 15N offset frequencies.
Both CW–CPMG sequences are nevertheless sensitive to artifacts from 1H off-resonance effects (Hansen et al. 2008b; Yip and Zuiderweg 2004). Amide 1H spins that are far off-resonance from the CW decoupling field are not fully decoupled from the 15N spin, resulting in generation of 2Nx,yHz magnetization through the residual J-coupling. Consequently, higher R2,eff values will be measured for low νCPMG values, for which free precession periods are long and more of the antiphase terms will be generated. This so-called slow-pulsing artifact shows up as an artefactual dispersion curve, interfering with accurate extraction of minor-state parameters.
Here, we analyze the slow-pulsing artifact in 15N CW–CPMG sequences in detail and demonstrate a simple method for its removal. In that, we took inspiration from the work of Chakrabarti et al. (2016), where composite pulse decoupling (CPD) was used to suppress 1H off-resonance effects in exchange mediated saturation transfer experiments. We investigated the performance of various CPD schemes in CW–CPMG sequences and demonstrate here that high power CPD based on the 90x–240y–90x element achieves artifact-free dispersion curves over a wide range of 1H offsets.
Materials and methods
NMR samples
NMR experiments were recorded on a sample of 2.5 mM uniformly 15N/13C-labelled Cu(II) azurin in 25 mM potassium phosphate buffer at pH 5.49 with 5% D2O. Labelled azurin was produced and purified according to a previously published protocol with modifications for incorporating 13C-glucose and 15N-ammoniumchloride (Karlsson et al. 1989).
NMR experiments
Relaxation dispersion experiments, using the ST–CW–CPMG sequence, were recorded at 298 K on Bruker Avance III HD spectrometers operating at 850 and 950 MHz 1H Larmor frequency and equipped with TCI cryoprobes. The constant-time CPMG relaxation delay (Trelax) was set to 40 ms with νCPMG set to 25, 50, 75, 100, 125, 175, 225 (2×), 275, 300, 350, 400 (2×), 500, 550, 600, 650, 700 (2×), 750, 800 (2×), 850, 900, 950 and 1000 Hz respectively, run in an interleaved manner. Duplicates were used to estimate the error in R2,eff. The errors were set to 0.2 s−1 at minimum. The pulse length of the 15N refocusing pulses in the CPMG train was 90 µs. For 1H decoupling, either CW decoupling or a CPD-scheme (GARP, DIPSI, MLEV16, WALTZ16, 90x–240y–90x) was used. This was implemented by changing the “cw:f1” statement in the pulse program to read “cpds1:f1” (pulse program available upon request). In either case, the decoupling field strength was 14.7 kHz (17 µs 1H 90° pulse), applied at 8.2 ppm 1H offset. A total of 3072/120 points were acquired in the 1H/15N dimension with an acquisition time of 90/27.85 ms and a relaxation delay of 2 s and 4 scans per FID. A reference spectrum, without the relaxation delay, was also recorded. NMR data were processed with NMRPipe (Delaglio et al. 1995), using linear prediction in the 15N dimension and Lorentz-to-Gauss window functions. Peak volumes were obtained by peak fitting using FuDa (Hansen, http://www.biochem.ucl.ac.uk/hansen/fuda/), and subsequently converted into effective relaxation rates via R2,eff(νCPMG) = − 1/Trelax·ln(I (νCPMG)/I0), where I0 is the peak intensity in a reference spectrum recorded without the relaxation delay Trelax. The R2,eff values measured using the ST–CW–CPMG sequence were corrected for R1-contribution according to the formula described by Jiang et al. (2015) using an estimate of 0.95 s−1 R1- and 10.5 s−1 for R2-contribution for all residues. Dispersion curves obtained with either CW or CPD decoupling were compared by calculating the RSMD between the curves for all residues:
where i is the index of a particular νCPMG value and the summation runs over the N recorded points, equal to the number of points per dispersion curve (M) times the number of residues. The systematic difference between the CW or CPD-based dispersion curves was calculated from the average point-by-point difference per residue and is tabulated in Table S1. To compensate for these systematic differences, an “R2-offset compensated” RMSD was calculated by replacing the CPD-based R2,eff values with the offset compensated values:
Simulation of 15N CW–CPMG dispersion profiles
To evaluate the magnitude of the slow-pulsing artifact in relaxation dispersion profiles, numerical simulations of a two-spin 1H–15N system were carried out, assuming a non-deuterated protein. The evolution of magnetization in this spin system was calculated for the CPMG part of the CPMG–CW and CPMG–ST–CW sequence, including the flanking 15N 90° pulses. Simulations in the absence of exchange and neglecting pulse imperfections were performed in operator space by solving the complete homogeneous master equation as described by Allard et al. (1998) and Helgstrand et al. (2000) using the open source computing language GNU Octave (http://www.gnu.org/software/octave/) (Eaton et al. 2008). All simulation used the parameters detailed below unless noted otherwise. The 15N spin was assumed to be on-resonance. The magnetic field strength was set to 19.9 T, corresponding to 1H Larmor frequency of 850 MHz. Relaxation rates were calculated using overall rotational correlation time τc of 9 ns, a value of 0.85 for the squared generalized order parameter, 100 ps for the correlation time for internal motions, and − 172/+10 ppm for the 15N/1H chemical shift anisotropy. Relaxation due to neighboring protons was included as described in ref. (Allard et al. 1998) by including a virtual proton at 1.85 Å, resulting in R2 values of in-phase and anti-phase 15N magnetizations of 13.6 and 26.7 s−1 respectively. Dispersion experiments were simulated with Trelax set to 40 ms, and νCPMG values ranging from 25 to 1000 Hz, the 15N 180° refocusing pulse was set to 90 µs, 1H CW decoupling field strength was set to 14.7 kHz (17 µs 1H 90°).
Results
Measurement of in-phase 15N CPMG relaxation dispersion profiles critically relies on 1H decoupling to measure the pure in-phase Nx,y relaxation rate without contamination by the anti-phase relaxation rate. As pointed out in the work of Jiang et al. (2015), the decoupling field strength has a practical limit of roughly 14 kHz, resulting in a residual J coupling interaction for amide protons at non-zero offset to the decoupling field. This interaction causes slow interconversion of in-phase and anti-phase magnetization during the CPMG period, which will lead to undesired averaging of the in-phase and anti-phase relaxation rates (Loria et al. 1999b; Hansen et al. 2008b). To first approximation, this averaging can be described by the equation derived by Palmer et al. (1992) for calculating the effective relaxation rate in spin echo sequences. Here, it is adapted and reformulated to express to the size of the slow-pulsing artifact A:
where R2in and R2anti are the 15N in-phase and anti-phase transverse relaxation rates, Jr is the residual J-coupling, and 2τcp is the inter-pulse delay in the CPMG pulse train. In the limit of perfect decoupling Jr ≈ 0, the sinc factor approaches 1 and A ≈ 0 for all τcp values. For non-zero Jr, A approaches zero in the limit of fast pulsing where τcp is very small. For slow pulsing, however, there is a non-zero artifact, with a theoretical limit of 0.5 (R2,anti − R2,in) for infinitely slow pulsing. In practice, Jr can be as much 16 Hz (for 3 ppm 1H offset at 850 MHz) and 2τcp is typically at most 20 ms, which would generate a maximum artifact of roughly 10% of the difference between the anti-phase and in-phase relaxation rate.
To assess more precisely how the slow pulsing artifact is manifested in 15N CPMG–CW and ST–CW experiments, numerical simulations of these sequences were performed in Liouville space for a non-exchanging two spin N–H system. Figure 1a compares the obtained dispersion profiles for the two experiments with the predicted curve based on Eq. 1, for a N–H system with 3 ppm 1H offset at an 850 MHz spectrometer. Whereas a flat curve is expected for a non-exchanging system, systematically increased R2,eff values are measured in the slow pulsing regime for both pulse sequences. While Eq. 1 is derived for periods of free evolution in absence of a decoupling field, the curvature of the slow-pulsing artifact matches the predicted sinc dependence on the pulsing rate. The size of the artifact is somewhat underestimated by Eq. 1. The original CW sequence shows slightly lower sensitivity to the artifact than the ST–CW experiment. This difference can be traced back to presence of the 15N refocussing pulse in between the two halves of the total CPMG period in the CW experiment. Importantly, since the shape of the artifact is virtually indistinguishable from a bona-fide dispersion profile, the artefactual R2,eff values can be fitted to an actual dispersion curve (see solid lines in Fig. 1a), illustrating the potential impact on the extracted exchange parameters.

Impact of the slow-pulsing artifact on simulated relaxation dispersion profiles. a Simulated slow-pulsing artifact caused by incomplete JNH decoupling in the CW–CPMG and the ST–CW–CPMG implementation of the in-phase 15N CPMG experiment. Solid lines are fits obtained using the program CATIA (Hansen, http://www.biochem.ucl.ac.uk/hansen/catia/) assuming two-site exchange. The artifact expected based on Eq. 1 is shown for comparison. The boxed region is expanded in the inset. The 1H offset from the decoupling field was set to 3 ppm, assuming an 850 MHz spectrometer. b, c Maximum size of the artifact (ΔR2,eff) as a function of (b) 1H offset for proteins of 4, 6.5 and 9 ns correlation times at 850 MHz; c magnetic field strength for 1 and 2 ppm 1H offset for proteins of 4 and 9 ns tumbling times. The gray area indicates the typical experimental error in range of 0.1–0.3 s−1. d The typical accessible 1H offset ranges, color coded into a 15N–1H HSQC spectrum. Assuming the 1H CW field is centered at 8 ppm, the blue region is accessible up to the highest magnetic fields, orange is accessible up to 600 MHz, and the red region is inaccessible. In a–c simulated profiles are shown for both CW–CPMG (open triangle) and ST–CPMG (asterisk) pulse sequences; color coding indicated in the figure. All simulations are based on a non-exchanging N–H spin system. Simulation parameters are given in “Materials and Methods” section, unless noted otherwise
Since the size of the slow pulsing artifact is governed by the relaxation difference between in-phase and anti-phase magnetization, it is dependent on protein size. Large proteins have more efficient 1H–1H spin flips which increase the anti-phase relaxation rate. Figure 1b compares the magnitude of the artifact for three different protein sizes as function of 1H offset from the decoupling field. For larger proteins, where the chance of finding amide protons at high offset is also higher, the artifact can be well above 1 s−1. At offsets larger than ~ 1000 Hz the slow pulsing artifact will be higher than the typical experimental error (on the order of 0.1–0.3 s−1) (Hansen et al. 2008a), as also noted by Jiang et al. (2015).
Since relaxation dispersion data need to be acquired at two magnetic fields in order to extract accurate protein dynamics parameters, we compared the size of the slow pulsing artifact for amide groups at 1 and 2 ppm 1H offset as function of magnetic field strength in Fig. 1c. High field strengths are not only attractive because of the sensitivity and resolution they afford, but also because they are more sensitive to exchange processes as they increase the frequency difference between states, Δϖ. However, for a given resonance, the offset from the decoupling field, and thus the slow pulsing artifact, will increase with increasing magnetic field strength. Strikingly, the artifact will already be significant at 1 ppm offsets for medium-sized proteins in a future 1.2 GHz spectrometer. To illustrate the impact of the slow pulsing artifact, generated by the inability of CW irradiation to decouple the full width of the amide spectrum, the HSQC can be divided in three areas: a narrow region ± ~1 ppm around the carrier frequency of the decoupling field that will be free of significant artifacts, the region beyond ± ~2 ppm in which significant artifacts will already occur at the lowest typical field strength, and the intermediate region (Fig. 1d).
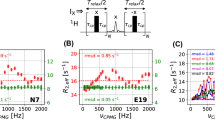
To confirm the results obtained from simulations, we experimentally demonstrated the problem using the ST–CW–CPMG pulse sequence on a sample containing azurin, a 16 kDa electron transfer metalloprotein (Adman 1991). A small subset of residues in azurin have been reported to undergo conformational exchange on the millisecond timescale (Korzhnev et al. 2003). To emphasize the slow pulsing artifact, we purposely centered the 1H decoupling field at 16 ppm such that the dispersion profiles are dominated by the artifact (Fig. 2a). Using this setup, we next screened several broadband decoupling sequences for their ability to suppress the artifact. These sequences rely on composite pulses to offer good population inversion even in the presence of off-resonance effects (Shaka and Keeler 1987), and thus should be able to suppress the artifact in theory. As can be seen in Fig. 2b, a wide range of CPD schemes indeed suppressed the artifact. Notably, the use of GARP (Shaka et al. 1985) and DIPSI2 (Shaka et al. 1988) results in spurious elevated R2,eff values at high pulsing rates, rendering the dispersion curves unusable. These spikes originate from the timing mismatch between the continuous train of (composite) 180° pulses on the 1H channel on the one hand and the repetition of free-evolution and 180° refocusing pulses on the 15N channel on the other hand. This mismatch results in incomplete decoupling at the end of each τcp period and thus elevated R2,eff values (Fig. 2c). As noted by Jiang et al. (2015), the duration of the mismatch is short when using adequately high power CW 1H decoupling, and thus the effect is small. Both DIPSI2 and GARP use particularly long composite pulses (corresponding to the length of 2590° and 1054° rotation, respectively), which aggravates the impact of the timing mismatch, in particular at high pulsing rates, where the effects from each τcp period are compounded. Indeed, use of WALTZ (540° duration) (Shaka et al. 1983) and MLEV (360° duration) (Levitt and Freeman 1981; Levitt et al. 1982a, b) with shorter duration of the composite pulse did not cause such high spikes. We next applied the 90x–240y–90x CPD scheme, which was recently used to suppress artifacts from incomplete 1H decoupling in exchange mediated saturation transfer experiments (Chakrabarti et al. 2016). The 90x–240y–90x CPD sequence has a short overall duration (420° rotation) and offers relatively broadband inversion, free from off-resonance effects without relying on supercycles (Levitt et al. 1982a; Levitt 1982). Gratifyingly, the 90x–240y–90x sequence effectively eliminated the artifacts without causing appreciable spikes or scatter in R2,eff values (Fig. 2d). The requirement for a short duration of the CPD element also means that the broadband performance of CPD decoupling cannot be used to reduce the decoupling power. Tests showed that reducing the decupling power to 7 kHz (34 µs decoupling pulse) resulted in spurious artifacts dominating the dispersion curves at high CPMG pulsing rates (data not shown).

Suppression of slow pulsing artifacts by composite pulse based broadband 1H decoupling. a Experimental relaxation dispersion curves for three residues measured using the ST–CW–CPMG sequence with the 1H decoupling field centered at 16 ppm. Dotted lines are fits to Eq. 1 to guide the eye. b Experimental dispersion curves for residue D62 using the indicated 1H decoupling schemes, all centered at 16 ppm. c When the 1H decoupling power is fixed, there is an inevitable timing mismatch between complete 1H inversion at the end of each composite pulse block (CPD) and the point (dotted line) of complete 15N inversion. As a result, decoupling is incomplete and resulting in elevated R2,eff values. d Experimental dispersion curves for residue D62 using CW and 90x–240y–90x 1H decoupling schemes, both centered at 16 ppm. In b, d between brackets are the average root-mean-square deviation (RMSD) to a straight line over all 114 residues in azurin. Note that the RMSD obtained with CW decoupling centered at 8.2 ppm was 0.7 s−1
While successful in suppressing the slow-pulsing artifacts, the use of composite pulse sequences for decoupling results in systematic differences in R2,eff values compared to those obtained using CW decoupling. This is most apparent from the WALTZ data in Fig. 2b, showing systematically reduced R2,eff values compared to the CW reference data. Such offsets between the CPD-derived and CW-derived dispersion curve are also found for MLEV and 90x–240y–90x decoupling, although typically much smaller. When using the 90x–240y–90x sequence, the average offset over all residues was found to be ~ 0.3 s−1 with 90% of the profiles having offsets below 0.6 s−1 (see Supplemental Table S1). Since this offset is small and the absolute value of R2,eff is not of importance when fitting dispersion curves, it will have negligible impact on the usefulness of the data obtained with CPD decoupling schemes.
Having established that WALTZ, MLEV and 90x–240y–90x decoupling sequences are able to suppress the slow pulsing artifact, we further tested their efficacy in a regular experimental setup with the decoupling field centered at 8.2 ppm. The obtained R2,eff values were compared point-by-point between the CPD and the CW data-set, and the root-mean-square deviation (RMSD) between data sets was calculated with and without compensating for the systematic offset in R2,eff values between the two datasets (Fig. 3a). Clearly, the 90x–240y–90x sequence performs best with an average RMSD to the reference CW data set of 0.17 s−1, which is on the order of the experimental error. The high quality of the data is visible from comparison of profiles obtained for residues with negligible 1H offset, such as shown in Fig. 3b. At high 1H offset from the decoupling field, the CW data suffers from the slow pulsing artifact, which is absent when using the 90x–240y–90x CPD sequence, as exemplified for T52 in Fig. 3c. Notably, this residue shows the slow pulsing artifact superimposed on a genuine dispersion of R2,eff values. From the comparison to the CPD-based experiment, it becomes clear that the data point at 25 Hz νCPMG pulsing rate is strongly affected by the slow pulsing artifact with R2,eff value spuriously elevated by ~ 1 s−1. As a final experiment, we recorded both CW and CPD-based dispersion profiles at the national ultra-high field NMR Facility at 950 MHz. At this field, the resonance with the highest 1H offset shows a slow-pulsing artifact of ~ 1.5 s−1 in the CW experiment, which is effectively suppressed when using the 90x–240y–90x decoupling sequence (Fig. 3d).

The 90x–240y–90x decoupling scheme offers high-quality dispersion curves free of slow-pulsing artifact. a Average RMSD between CPD- and CW-based dispersion curves over all analyzed residues in azurin. Open/closed bars refer to the RMSD without/with compensating for the offset between the curves. b–d Experimental dispersion curves for both CW and 90x–240y–90x based experiments for amide resonances of A92 (no 1H offset, panel b), and T52, and an unassigned Arg sidechain resonance (Rsc) both with significant 1H offset. Data for panels b, c recorded at 850 MHz. Data for panel d recorded at 950 MHz. Dotted lines are best-fit dispersion curves obtained using CATIA (Hansen, http://www.biochem.ucl.ac.uk/hansen/catia/). The CPD data were corrected for the systematic offset to the CW data before plotting
Discussion
We have investigated the impact of the slow-pulsing artifact in in-phase 15N relaxation dispersion experiments by theoretical considerations, numerical simulations and experiments. We show that the artifact can be removed by using CPD-based 1H decoupling during the CPMG period. Out of the tested CPD sequences, the 90x–240y–90x sequence offers the best performance: the artifact is fully suppressed, while retaining shape of the dispersion curve obtained using CW decoupling within experimental error. Notably, this is done without introducing spurious spikes in R2,eff values at high pulsing rates, and with minimal offset to the CW-based dispersion profiles. Critical to its performance seems to be short duration of the composite pulse combined with relatively high quality of off-resonance performance.
The cause of the slight offset between the CPD and CW-based dispersion profiles is unclear. Closer inspection shows that the magnitude of the offset shows no correlation to either the N, HN, or Hα chemical shift and that both reference (no CPMG delay) and dispersion experiment (with CPMG delay) have slightly altered intensities (~ 2–5%) in the CPD experiment compared to the CW experiment. The effect on the reference experiment, where the decoupling block is carried out before the recycle delay, signifies that the both types of decoupling result in a different steady-state magnetization, presumably both for water and protein protons.
As for the water magnetization, a disadvantage of using CPD over CW decoupling is the loss of control over its state. Whereas in the CW case the water magnetization is spin-locked and returned to + z after the CPMG period, continuous alteration between x and y-pulse phase during the 90x–240y–90x CPD element causes dephasing and loss of water polarization. Experimental tests (Hiller et al. 2005) demonstrate this effect and show that after a 2 s delay, corresponding to the recycle delay to the next proton excitation pulse, there is minimal difference between the water polarization in the CPD and CW case (see Supplemental Fig. S1). Here, radiation damping caused by the high Q of the cryogenic probe likely aids the recovery of the water magnetization in the CPD case. Additionally, the low pH of the sample (5.5) will slow down amide-water exchange and thus additionally dampen the effect of (residual) water saturation.
In the original implementation of the in-phase dispersion experiment described by Hansen et al. (2008b), the strength of the decoupling field is matched to the CPMG pulsing rate to avoid the timing mismatch as indicated in Fig. 2c. In principle, such matching could also be done when using CPD decoupling schemes, which should result in decreased scatter in the dispersion curves. While simulations indeed show such improvement in performance, an experimental test showed a severe increase in scatter, presumably due to a point-to-point variation in the steady state of the water and aliphatic proton magnetization.
As noted in Fig. 1, the slow-pulsing artifact will be particularly problematic at high magnetic field strengths. At such high fields, it may be better to use TROSY–CPMG sequences (Loria et al. 1999a), which do not suffer from the slow-pulsing artifact, even for non-deuterated moderately sized proteins. The relative sensitivity of TROSY and in-phase CPMG experiments is best assessed experimentally as it not only depends on magnetic field strength but also on protein size, labeling pattern, and temperature. Next to the absolute sensitivity, one may also consider that lower 15N relaxation rates during the CPMG period allow the use of longer CPMG delays, increasing the sensitivity to slow motions (Loria et al. 1999a), as well as spectral quality of TROSY spectra (reduced overlap vs. presence of anti-TROSY lines). Additionally, in case data at lower field strength have been recorded using the in-phase CPMG experiment it may be necessary to record these at high fields too.
In conclusion, we show here that the use of broadband 1H decoupling, in particular using the 90x–240y–90x sequence, is a viable and attractive option for recording in-phase 15N relaxation dispersion data. This option is particularly relevant when the protein spectrum contains resonances far from center. It offers artifact-free dispersion profiles without the need for recording data in multiple sets or the need for eliminating of data points, all without compromising data quality.
References
Adman ET (1991) Copper protein structures. Adv Protein Chem 42:145–197
Akke M, Palmer AG (1996) Monitoring macromolecular motions on microsecond to millisecond time scales by R1ρ–R1 constant relaxation time NMR spectroscopy. J Am Chem Soc 118:911–912
Allard P, Helgstrand M, Hard T (1998) The complete homogeneous master equation for a heteronuclear two-spin system in the basis of cartesian product operators. J Magn Reson 134:7–16
Chakrabarti KS et al (2016) High-power 1H composite pulse decoupling provides artifact free exchange-mediated saturation transfer (EST) experiments. J Magn Reson 269:65–69
Culik RM et al (2018) Effects of maturation on the conformational free-energy landscape of SOD1. Proc Natl Acad Sci 115:E2546
Delaforge E et al (2018) Deciphering the dynamic interaction profile of an intrinsically disordered protein by NMR exchange spectroscopy. J Am Chem Soc 140:1148–1158
Delaglio F et al (1995) NMRPipe: a multidimensional spectral processing system based on UNIX pipes. J Biomol NMR 6:277–293
Eaton JW, Bateman D, Hauberg S (2008) GNU Octave manual version 3. Network Theory Ltd., Bristol, p 568
Eisenmesser EZ et al (2005) Intrinsic dynamics of an enzyme underlies catalysis. Nature 438:117
Fawzi NL, Ying J, Torchia DA, Clore GM (2010) Kinetics of amyloid β monomer to oligomer exchange by NMR relaxation. J Am Chem Soc 132:9948–9951
Franco R, Gil-Caballero S, Ayala I, Favier A, Brutscher B (2017) Probing conformational exchange dynamics in a short-lived protein folding intermediate by real-time relaxation–dispersion NMR. J Am Chem Soc 139:1065–1068
Frauenfelder H, Sligar SG, Wolynes PG (1991) The energy landscapes and motions of proteins. Science 254:1598–1603
Gopalan AB, Hansen DF, Vallurupalli P (2018) CPMG experiments for protein minor conformer structure determination. In: Ghose R (ed) Protein NMR: methods and protocols. Springer, New York, pp 223–242
Hammes GG (1964) Mechanism of enzyme catalysis. Nature 204:342
Hansen F. http://www.biochem.ucl.ac.uk/hansen/catia/. Accessed 1 June 2018
Hansen F. http://www.biochem.ucl.ac.uk/hansen/fuda/. Accessed 1 June 2018
Hansen DF, Vallurupalli P, Lundström P, Neudecker P, Kay LE (2008a) Probing chemical shifts of invisible states of proteins with relaxation dispersion NMR spectroscopy: how well can we do? J Am Chem Soc 130:2667–2675
Hansen DF, Vallurupalli P, Kay LE (2008b) An improved 15N relaxation dispersion experiment for the measurement of millisecond time-scale dynamics in proteins. J Phys Chem B 112:5898–1904
Helgstrand M, Härd T, Allard P (2000) Simulations of NMR pulse sequences during equilibrium and non-equilibrium chemical exchange. J Biomol NMR 18:49–63
Henzler-Wildman K, Kern D (2007) Dynamic personalities of proteins. Nature 450:964–972
Henzler-Wildman KA et al (2007) Intrinsic motions along an enzymatic reaction trajectory. Nature 450:838
Hiller S, Wider G, Etezady-Esfarjani T, Horst R, Wüthrich K (2005) Managing the solvent water polarization to obtain improved NMR spectra of large molecular structures. J Biomol NMR 32:61–70
Jiang B, Yu B, Zhang X, Liu M, Yang D (2015) A (15)N CPMG relaxation dispersion experiment more resistant to resonance offset and pulse imperfection. J Magn Reson 257:1–7
Karlsson BG, Pascher T, Nordling M, Arvidsson RHA, Lundberg LG (1989) Expression of the blue copper protein azurin from Pseudomonas aeruginosa in Escherichia coli. FEBS Lett 246:211–217
Kim TH et al (2017) The role of dimer asymmetry and protomer dynamics in enzyme catalysis. Science. https://doi.org/10.1126/science.aag2355
Kimsey IJ, Petzold K, Sathyamoorthy B, Stein ZW, Al-Hashimi HM (2015) Visualizing transient Watson-Crick like mispairs in DNA and RNA duplexes. Nature 519:315–320
Korzhnev DM, Karlsson BG, Orekhov VY, Billeter M (2003) NMR detection of multiple transitions to low-populated states in azurin. Protein Sci 12:56–65
Korzhnev DM, Religa TL, Banachewicz W, Fersht AR, Kay LE (2010) A transient and low-populated protein-folding intermediate at atomic resolution. Science 329:1312–1316
Kovermann M, Rogne P, Wolf-Watz M (2016) Protein dynamics and function from solution state NMR spectroscopy. Q Rev Biophys 49:e6
Levitt MH (1982) Symmetrical composite pulse sequences for NMR population inversion. II. Compensation of resonance offset. J Magn Reson (1969) 50:95–110
Levitt MH, Freeman R (1981) Composite pulse decoupling. J Magn Reson 43:502–507
Levitt MH, Freeman R, Frenkiel T (1982a) Broadband heteronuclear decoupling. J Magn Reson (1969) 47:328–330
Levitt MH, Freeman R, Frenkiel T (1982b) Supercycles for broadband heteronuclear decoupling. J Magn Reson 50:157–160
Libich DS, Tugarinov V, Clore GM (2015) Intrinsic unfoldase/foldase activity of the chaperonin GroEL directly demonstrated using multinuclear relaxation-based NMR. Proc Natl Acad Sci USA 112:8817–8823
Lisi GP (2016) & Patrick Loria, J. Using NMR spectroscopy to elucidate the role of molecular motions in enzyme function. Prog Nucl Magn Reson Spectrosc 92–93:1–17
Loria JP, Rance M, Palmer AG (1999a) A TROSY CPMG sequence for characterizing chemical exchange in large proteins. J Biomol NMR 15:151–155
Loria JP, Rance M, Palmer AG (1999b) A relaxation-compensated Carr–Purcell–Meiboom–Gill sequence for characterizing chemical exchange by NMR spectroscopy. J Am Chem Soc 121:2331–2332
Massi F, Peng JW (2018) Characterizing protein dynamics with NMR R 1ρ relaxation experiments. In: Ghose R (ed) Protein NMR: methods and protocols. Springer, New York, pp 205–221
Neudecker P et al (2012) Structure of an intermediate state in protein folding and aggregation. Science 336:362–366
Palmer AG 3rd, Kroenke CD, Loria JP (2001) Nuclear magnetic resonance methods for quantifying microsecond-to-millisecond motions in biological macromolecules. Methods Enzymol 339:204–238
Palmer AG (2015) Enzyme dynamics from NMR spectroscopy. Acc Chem Res 48:457–465
Palmer AG, Skelton NJ, Chazin WJ, Wright PE, Rance M (1992) Suppression of the effects of cross-correlation between dipolar and anisotropic chemical shift relaxation mechanisms in the measurement of spin-spin relaxation rates. Mol Phys 75:699–711
Pratihar S et al (2016) Kinetics of the antibody recognition site in the third IgG-binding domain of protein G. Angew Chem Int Ed 55:9567–9570
Sauerwein A, Hansen D (2015) Relaxation dispersion NMR spectroscopy. In: Berliner L (ed) Protein NMR. Biological magnetic resonance. Springer, Boston
Schneider R et al (2015) Visualizing the molecular recognition trajectory of an intrinsically disordered protein using multinuclear relaxation dispersion NMR. J Am Chem Soc 137:1220–1229
Shaka AJ, Keeler J (1987) Broadband spin decoupling in isotropic-liquids. Prog Nucl Magn Reson Spectrosc 19:47–129
Shaka AJ, Keeler J, Frenkiel T, Freeman R (1983) An improved sequence for broadband decoupling: WALTZ-16. J Magn Reson (1969) 52:335–338
Shaka AJ, Barker PB, Freeman R (1985) Computer-optimized decoupling scheme for wideband applications and low-level operation. J Magn Reson (1969) 64:547–552
Shaka AJ, Lee CJ, Pines A (1988) Iterative schemes for bilinear operators; application to spin decoupling. J Magn Reson (1969) 77:274–293
Sugase K, Dyson HJ, Wright PE (2007) Mechanism of coupled folding and binding of an intrinsically disordered protein. Nature 447:1021
Vallurupalli P, Bouvignies G, Kay LE (2012) Studying “invisible” excited protein states in slow exchange with a major state conformation. J Am Chem Soc 134:8148–8161
Vallurupalli P, Sekhar A, Yuwen T, Kay LE (2017) Probing conformational dynamics in biomolecules via chemical exchange saturation transfer: a primer. J Biomol NMR 67:243–271
Wolynes PG (2005) Recent successes of the energy landscape theory of protein folding and function. Q Rev Biophys 38:405–410
Xiao T, Fan Js, Zhou H, Lin Q, Yang D (2016) Local unfolding of fatty acid binding protein to allow ligand entry for binding. Angew Chem Int Ed 55:6869–6872
Xue Y et al (2015) Characterizing RNA excited states using NMR relaxation dispersion. Methods Enzymol 558:39–73
Yip G, Zuiderweg E (2004) A phase cycle scheme that significantly suppresses offset-dependent artifacts in the R 2-CPMG 15 N relaxation experiment. J Magn Reson 171:25–36
Zhao B, Guffy SL, Williams B, Zhang Q (2017) An excited state underlies gene regulation of a transcriptional riboswitch. Nat Chem Biol 13:968–974
Acknowledgements
We thank prof. Daiwen Yang (National University of Singapore) for sharing the pulse sequence code for the ST-CW-CPMG experiment. This work was supported by financial support from the Dutch Science Foundation NWO by a VIDI grant (723.013.010) to HvI and grant 184.032.207 to the uNMR-NL National Roadmap Large-Scale Facility of the Netherlands.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Chatterjee, S.D., Ubbink, M. & van Ingen, H. Removal of slow-pulsing artifacts in in-phase 15N relaxation dispersion experiments using broadband 1H decoupling. J Biomol NMR 71, 69–77 (2018). https://doi.org/10.1007/s10858-018-0193-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10858-018-0193-2