
Abstract
Insect habitat specialists have declined more than generalists over the last decades. Understanding how habitat requirements and increased climate variability interact to explain persistence of populations is needed to enable effective conservation measures. Habitat specialists with a complex life cycle, such as myrmecophilous butterflies, are likely to be most vulnerable. Here, we explored to what extent the habitat factors explaining population persistence of the Alcon blue (Phengaris alcon), changed over a 25-year period in the Netherlands, contrasting the recent years 2018–2022 with extreme droughts with the preceding period 1998–2017 characterized by occasional climatic extremes. Population persistence of P. alcon over 1998–2017 as explained by historical habitat conditions was significantly higher in large heathland areas with large patches of its host plant Gentiana pneumonanthe. On the basis of current conditions, population persistence until 2017 was best explained by a combination of host plant area and quality. In contrast, persistence over the recent drought years was most closely associated with increasing occupancy of the optimal host ant Myrmica ruginodis and lower occupancy of competitive and predatory Lasius ants.
Implications for insect conservation We conclude that critical habitat conditions change under increasing climatic extremes and that the availability of sufficient optimal host ants has become more critical than that of host plants. The results emphasise that both large-scale hydrological restoration and small-scale management for heterogeneity are necessary to preserve P. alcon and its habitat in the Netherlands. This is also likely to apply to other habitat specialists with complex life cycles from threatened habitats.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Recent years have seen mounting evidence of declines in insect abundance and diversity across many insect groups (Forister et al. 2019; Klink et al. 2020). Direct impacts from habitat loss and fragmentation and indirect impacts from increasing land use intensity (including nitrogen deposition and pesticides) and climate change appear to be the main drivers of these declines (Wagner et al. 2021; Warren et al. 2021; Outhwaite et al. 2022). Also, habitat specialist species appear more severely affected than habitats generalists (Warren et al. 2001; Menéndez et al. 2007; Clavel et al. 2010; Brereton et al. 2011), although this trend is not consistent across insect groups (Engelhardt et al. 2022). Indeed, distinguishing habitat specialists from generalists is fraught with ambiguity (WallisDeVries 2014), because the greater vulnerability of habitat specialists may stem from a variety of reasons: their habitat may simply always have been rare, it may have become rare in recent times, resources such as host plants may be rare or habitat quality may have deteriorated. If we are to successfully preserve endangered habitats specialists, there is an urgent need to elucidate which are the critical threats that need to be alleviated. This task is growing more urgent with the impacts of climate change (Halsch et al. 2021; Hill et al. 2021), which may become especially severe with increased frequencies of extreme weather events (Evans et al. 2022; Mahecha et al. 2022).
The most vulnerable species of all probably are those habitat specialists with complex life cycles. Although these may have evolved as successful adaptations, their complexity can undermine their survival under the recent rapid environmental changes. Myrmecophilous butterflies provide a perfect case to illustrate this point, as they depend on both a specific host plant and a specific host ant to complete their larval development. Their adaptation to escape predation by ants and instead exploit them to provide both a safe space and food ensures a high larval survival, which confers a clear evolutionary advantage (Als et al. 2004; Tartally et al. 2019). In times of change, however, the dependence on both host ants and host plants from low-productive environments (Elmes et al. 1998), which are threatened by land use intensification as well as abandonment (WallisDeVries 2014; Warren et al. 2021), has led to the demise of all four species of Phengaris (Maculinea) butterflies in large parts of Europe (Wynhoff 1998; Thomas et al. 2009; Van Swaay et al. 2011).
This study takes the fate of Alcon blue (Phengaris alcon) populations in the Netherlands as a case to unravel the critical factors of habitat area and quality determining the persistence of an endangered habitat specialist with a complex life cycle. Although P. alcon may use several species of Gentiana and Gentianella as host plants, in this region it currently depends exclusively on the rare Marsh gentian, Gentiana pneumonanthe. Gentiana cruciata has been documented as an alternative host plant in coastal dunes, but the last record from this isolated population dates back to 1979 (Bos et al. 2006). Eggs are laid on the gentian’s young flower buds, where the larvae spend the first 2–3 weeks feeding on the developing seeds. After falling to the ground, the larvae depend on being adopted by Myrmica host ants, such as M. ruginodis, M. scabrinodis and possibly M. rubra, with M. ruginodis as probably the optimal host ant species (Elmes et al. 1998; Vanreusel et al. 2000). The caterpillars develop in the ants’ nest until pupation. They are fed by trophallaxis by the worker ants as if they were part of the ants’ brood and occasionally also feed on the grubs (Tartally et al. 2019). The butterfly generally emerges from the nest between mid-June and early August, with individuals living about three days on average (Bubová et al. 2016).
In the Netherlands, Phengaris alcon has declined by 79% in distribution since 1990 and by even more in population size (CBS and RIVM 2023). Critical habitat factors for population persistence have been studied earlier over the period 1990–1998 (WallisDeVries 2004), highlighting the importance of habitat area, host plant abundance and host ant presence. Here, we test to what extent the habitat factors explaining population persistence of the Alcon blue (Phengaris alcon), changed over a 25-year period in the Netherlands, contrasting the recent years 2018–2022 with extreme droughts with the preceding period 1998–2017 characterized by incidental and less severe climatic extremes. We expected impacts of host plant quality on population persistence through the sensitivity of gentians to desiccation resulting from both local conditions (Oostermeijer et al. 1998; Wamelink et al. 2012) and climate (Cerrato et al. 2016). Beneficial impacts of small-scale management and detrimental effects of large-scale management, already shown by WallisDeVries (2004), were expected to remain important. In addition, we surmised possible impacts of changing moisture conditions on the ant community (Seifert 2018), with potentially detrimental effects on the host ants of P. alcon, but the scant evidence on host ant dynamics precluded more definite expectations of such impacts.
Materials and methods
Study design
The focus of our study was to determine differences in current habitat conditions explaining survival and extinction of P. alcon populations between the period preceding frequent climatic extremes (1998–2017) and recent extreme years (2018–2022), with extreme summer drought in 4 out of 5 years as compared to 4 milder drought extremes out of 20 years in the preceding period (Fig. S1). In the analysis of population persistence, we adopted a resource-based habitat approach (Dennis et al. 2006), in which a habitat is defined as a suite of resources and environmental conditions (Caughley and Sinclair 1994).
Data were collected in 2019–2022 for 62 sites, and in 2018 for 2 additional sites (Table S1). In addition, we tested to what extent population persistence could be explained by historical habitat characteristics. To that effect, we used data from WallisDeVries (2004) on extant populations in 1998 for 62 sites and used habitat data from that year to assess critical factors determining population persistence between 1998 and 2017.
Study areas
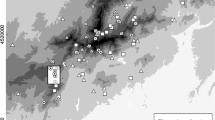
All study sites were located on Pleistocene sandy soils in the Netherlands, as shown in Fig. 1. A site was typically defined as a large patch of host plants (formerly) occupied by P. alcon. In larger areas, multiple sites were sampled at distances of > 500 m from each other (Fig. S2). Data from historical habitat conditions of extant populations in 1998 were available from 62 sites. Current habitat conditions were studied in 64 sites in the recent period 2018–2022, including the sites surveyed in 1998, but excluding already abandoned sites. Recent field data included all sites with potential for P. alcon: extant populations in 2017, satellite sites abandoned since 1998 within 5 km from extant populations, as well as eight sites in semi-natural areas where extensive habitat restoration has taken place after the local extinction of P. alcon. These additional sites included four sites with habitat restoration on former agricultural land by removal of the eutrophic topsoil. Including these restoration sites extended the range of potential habitat conditions encountered by P. alcon.

Location of study sites in the Netherlands where population persistence of P. alcon was studied from (a) 1998–2022 on the basis of recent habitat conditions (white dots, n = 64) and (b) 1998–2017 based on historical habitat conditions (black dots, n = 62)
Most of the recent sites (55) were situated in seminatural wet heathland habitats, while 9 were located in fen meadows or moist acidic grassland. The most common habitat management in these areas consisted of sod-cutting or choppering (i.e. flail-mowing at soil level with removal of the cuttings), followed by low-intensity grazing, removal of tree saplings and hydrological restoration by rewetting; mowing and abandonment were less common. Sod-cutting was typically carried out in small patches of less than 10 m2 (small-scale sod-cutting), and less frequently in patches exceeding 200 m2 (large-scale sod-cutting), although large-scale topsoil removal was applied in four restoration areas on former agricultural land. Only management actions carried out after 2000 were included in the analysis. Grazing impact was assessed indirectly through vegetation structure and indications on hydrological conditions were derived from plant species composition and soil moisture.
Field sampling of habitat conditions and resource availability
For the field sampling in 2018–2022, we collected data on habitat conditions within gentian patches on all sites from vegetation relevés and soil samples and resource availability of host plants and host ants (variables listed in Table S2). Nectar resources are not considered to be a main limiting factor for the short-lived P. alcon butterfly (WallisDeVries 2004). Therefore, they have not been treated with specific emphasis, although they are largely reflected by the abundance of the main nectar source Erica tetralix, which has been included in the analysis. Habitat management by sod-cutting, choppering or topsoil removal carried out after 2000 was recorded as either small-scale or large-scale, i.e. <> 100 m2.
Gentian patch area (in ha) at each site was calculated as the area of the minimum convex polygon encompassing all gentians clustered within a radius of 100 m from each other, based on records of our own survey supplemented with records at a precision of < 100 m from the national database flora and fauna, NDFF (https://www.ndff.nl/english/), over the period 2010–2022.
In the centre of the gentian patch, vegetation structure and plant species composition were assessed in 5 × 5 m quadrats. Vascular plant species composition was assessed using a 9-point Braun-Blanquet scale for abundance/cover. Vegetation structure was described by cover estimates of bare ground, litter, bryophytes, herbaceous vegetation < 25 cm, 25–50 cm and > 50 cm in height, and shrubs or trees (total cover adding up tot 100%). The abundance of the gentian host plants was counted as the number of flowering plants, similar to 1998. Host plant regeneration was recorded as the abundance of seedlings and juvenile vegetative gentians (cf. Oostermeijer et al. 1994) on a 4-point log5-scale.
The availability of high-quality host plants (see Kuer and Fartmann 2004; Wynhoff et al. 2014) was calculated as the proportion of gentians with at least 3 buds on the primary flowering stem from a subsample of 30 flowering gentians per site at mutual distances of > 2 m.
Ant occupancy was sampled at each site with a series of 30 trap tubes (17 mm diameter, 15 ml) baited with fruit wine. Traps were placed at 4 m intervals next to a host plant, with the opening at soil level and marked with a flag for retrieval. Traps were collected again after > 24 h (mostly 2 days). In the lab, samples were rinsed and ants were sorted and stored in 96% ethanol for later species identification, after Boer et al. (2023). Species occupancy was expressed as the percentage of occupied traps. Only the three potential host ants Myrmica ruginodis, M. scabrinodis and M. rubra as well as the potentially competitive and predatory Lasius niger and L. platythorax were included in the analysis.
Soil samples were taken with a 15 mm gauge of the top 10 cm of the soil. For each core, the thickness (in cm) of the dark organic mineral layer was recorded with 0.5 cm accuracy to obtain an average organic layer thickness. Soil samples from cores taken at the first 10 ant traps were pooled for further analysis. The samples were analysed for concentrations (in ppm) of minerals (Ca, K, Mg) and nitrogen available as NH4 or NO3 measured by photometric determination with a Segmented Flow Analysis (SFA) in a Skalar San + + Automated Wet Chemistry Analyser; P was not included as it was known to occur only in very low concentrations in these habitats. Soil pH was measured both in demineralised water and in a KCl solution with a Eutech pH 5 at room temperature. Soil moisture fraction was determined after drying at 105 C for 24 h. For the organic matter faction, samples were weighed again after three hours in a stove at 550 C.
Habitat conditions in 1998 were taken from WallisDeVries (2004). Habitat variables included heathland area (in ha, derived from topographical maps), patch area of the host plant (in m2, field survey), density of flowering gentian plants (in N/m2) and mean host ant nest occupancy of the three potential host ant species (after 15 min searches) over typical three 10 × 10 m plots on each site. Connectivity between occupied sites in 1998 was calculated as the sum of the negative exponentials of the distances d (in km) between a specific site i and all other sites j (i.e. Σ exp− dij where i ≠ j), which is the negative equivalent of I3 in Hanski et al. (1994).
Population-scale habitat conditions
The status of P. alcon populations was determined from our own field study and monitoring data from the Dutch Butterfly Monitoring Scheme (CBS and RIVM 2023), mostly by egg counts in permanent plots and occasionally also by butterfly transect counts.
P. alcon population area (in ha) was also considered as a resource, encompassing host plants and host ants as well as the traveling range between suitable patches for reproduction, following the resource-based habitat concept developed by Vanreusel and Van Dyck (2007). Its area was calculated with a similar approach as used for determining gentian patch area, with the area of the minimum convex polygon encompassing all precise records (accuracy of < 100 m) of eggs and butterflies from the national database flora and fauna NDFF since 1990. As the butterfly is more mobile than its host plant, the radius to cluster records of the same population was set at 250 m, a distance over which butterflies only rarely disperse (Maes et al. 2004). Polygons were only determined for clusters with ≥ 10 records.
As a measure of habitat heterogeneity in relation to groundwater influence, we used the actual height dataset for the Netherlands (AHN3, https://www.ahn.nl/) to calculate the average surface elevation (in m) at soil level in 5 × 5 m squares with the ‘Zonal Statistics As Table’ tool in ArcGIS 10.6. Topographical heterogeneity was then taken as the standard deviation in elevation of all 5 × 5 m squares within each population polygon.
For analyses of population persistence, habitat variables of the 2018–2022 field survey were aggregated to a single value for each population. Variables were averaged over sites after log10(n + 1) transformation. If populations contained multiple sites with different P. alcon status, only sites with the most recent P. alcon occurrence were included. For gentian patch area, we summed the area of polygons within the P. alcon population polygon. Restoration sites where P. alcon had never been recorded were excluded. This resulted in a dataset of 31 populations (see Table S1).
Statistical analysis
We analysed habitat conditions at site level, taking each site as an independent replicate, because the distances to neighbouring sites were considered sufficiently large (> 250 m with median 1.05 km) to assume negligible exchange between host plant and ant populations among sites. At the scale of P. alcon populations, we pooled the information of different sites within a population polygon, as explained above. Again, different populations were considered sufficiently distant from their neighbours (0.8–31.5 km, median 3.75 km) to exclude significant interference. Also, given the similar physico-geographical conditions and the limited geographical extent of the total study region (220 × 70 km), we did not expect significant regional differentiation.
All statistical analyses were carried out in R version 4.2.1 (R Core Team 2022). To arrive at independent factors describing the main variation in habitat conditions at site level, we applied Principal Component Analysis (PCA) to 27 variables of the 2018–2022 dataset describing vegetation structure, botanical composition and soil chemistry. Botanical composition included only the 10 most frequent species (with the main nectar plant Erica tetralix), occurring in ≥ 20% of all relevés. Vegetation cover values were log10(n + 1) transformed beforehand. Additionally, as a general habitat characteristic of the grassland-heathland ecotone, we included the richness of typical fen meadow (Junco-Molinion) species (after Schaminée et al. 2017). PCA was carried out with varimax rotation using “principal” from the package “psych” (Revelle 2020).
Subsequently, generalized linear models were applied, using the function “glm” from the package “stats”, to analyse (a) habitat conditions (including Lasius ants) as explained by management, (b) resource availability for P. alcon as explained by habitat conditions and management and (c) the persistence of P. alcon populations: (i) over the period 1998–2017 by forecasting from 1998 habitat conditions, (ii) over the period 1998–2017 by hind-casting from current conditions and (iii) over the period 2017–2022 from current conditions (full models are listed in Table S3). A stepwise backwards selection procedure was followed to retain only significant variables at p < 0.05, allowing at most one variable to contribute at p < 0.10; only for population persistence over recent years, the small dataset forced us to apply a forward selection (but forwards and backwards procedures typically led to the same outcome with the larger datasets).
Regarding habitat conditions, principal components were analysed as a function of habitat management with family = gaussian. The occupancy of Lasius ants was examined as a function of both principal components and habitat management with family = quasibinomial. For host ant resources, the occupancy of each of the potential Myrmica host ants was analysed likewise, including Lasius occupancy as an additional factor. Gentian resources, i.e. flowering gentian patch area (log10(n + 1) transformed), gentian density (log10(n + 1) transformed), gentian regeneration and gentian quality, were also examined as a function of both principal components and habitat management with family = gaussian for the first three variables and family = quasibinomial for the fourth variable. For gentian density, gentian regeneration was added as an additional explanatory variable.
Population persistence over the period 1998–2017 based on habitat conditions in 1998 was examined in response to connectivity, heathland size, gentian patch area, gentian density (including its squared component) and the occupancy of the three potential host ant species (Myrmica ruginodis, M. scabrinodis and M. rubra), using a glm with family = binomial.
Population persistence based on the 2018–2022 dataset was analysed with the same glm set-up as a function of P. alcon population area, topographical heterogeneity, as well as large-scale and small-scale cutting, occurrence of Lasius ants and resource availability of host plants (gentian area, gentian density, gentian quality) and the occupancies of the three potential host ants.
Results
Habitat conditions
Four principal components explained 46.6% of the variance in the local habitat conditions based on soil chemistry, organic matter and moisture, vegetation structure and botanical composition (Table 1). The first component (15.6% of the variance) was characterised by variables correlated with soil buffering with positive correlations for pH, calcium and potassium as well as plants from fen meadows. Negative correlations were found for litter cover and the wet heathland species Erica tetralix and to a lesser extent Molinia caerulea. Magnesium was not included in the PCA due to some missing values, but it was significantly correlated with PC1 (r = + 0.72, p < 0.0001). First component values were lower under small-scale management, underlining that this measure was largely restricted to heathland (Table 2).
The second component (11% of the variance) reflected soil moisture, with positive correlations for soil moisture, organic matter, and thickness of the dark mineral soil layer. The third component (10.5% of the variance) expressed vegetation succession, with negative correlations for the cover of the early successional plant species Drosera intermedia and Rhynchospora fusca and the cover of bare soil and positive correlations with Molinia caerulea and to a lesser extent soil ammonium concentration. This component had significantly lower values under especially large-scale management, but also under small-scale management (Table 2; Fig. 2). The fourth component (8.8% of the variance) captured vegetation structure and correlated positively with the cover of short vegetation and negatively with that of taller vegetation. The second and fourth components were not significantly affected by management.

Schematic representation of relations between habitat conditions (green boxes), management and topography (brown), competitive and predatory ants (red), potential host ants (orange), host plants (blue) and the area occupied by P. alcon (light blue). Solid lines show significant effects (p < 0.05) while dashed lines indicate marginally significant effects (p < 0.10), with blue lines showing positive effects and red lines showing negative effects
Lasius ants were found in 33.6% of the traps (L. niger 13.8%, L. platythorax 22.8%, with occasional overlap) and occurred most frequently after large-scale management and under the more acidic conditions in heathlands (Table 2). The contribution of L. niger and L. platythorax occupancy when pooled contributed more to explain P. alcon population persistence than when both species were entered as separate variables.
Resource availability
At site level, resource availability for P. alcon was explained most significantly by habitat conditions related to the quality of it host plant and availability of host ants (Table 2). Gentian regeneration was most abundant when management had been carried out, irrespective of scale. Gentian abundance was higher with more abundant gentian regeneration, but not with other habitat conditions. Gentian quality, in terms of the number of flower buds per plant, was significantly lower in areas with pioneer vegetation than in later successional stages and it tended to increase with soil moisture. In contrast, gentian area was not significantly explained by any variable.
The expected optimal host ant, M. ruginodis, was present in 8.2% of the traps on average and occurred more frequently in moist conditions and less in taller vegetation (< 20% cover of the vegetation layer exceeding 50 cm in height) and after small-scale management. The alternative host ant M. scabrinodis was more than twice as abundant as M. ruginodis (18.8% of the traps) and was only marginally affected by habitat conditions, with weak positive effects of soil buffering and moisture. The second alternative host ant, M. rubra, was comparatively rare (5.7% of the traps) and was found significantly more on acid heathland soils in lower vegetation and in the absence of Lasius ants; small-scale management also tended to negatively affect its occurrence.
At the scale of P. alcon populations, habitat area or population extent (as defined by the extent of egg and butterfly records encompassing host plant patches as well as foraging area and travelling range) was strongly related to the area covered by the host plant Gentiana pneumonanthe, as was to be expected, and furthermore by the application of small-scale management. Variation in topography contributed only marginally to explain habitat area.
Population persistence
The persistence of P. alcon in 2017 as predicted by habitat conditions assessed in 1998 showed a survival on only 17 of the 62 sites. Population persistence was explained by two factors: it was higher in larger heathlands and with larger gentian patches (Table 3). There was no significant contribution of local resources, i.e. gentian abundance and host ant presence. Surprisingly, the host ant M. ruginodis was even more frequently present in populations that went extinct (p = 0.0128), but this effect was not retained when including other explanatory variables in the glm (p > 0.05).
The hind-casting of P. alcon persistence in 2017 from current habitat conditions involved 31 populations, with 22 surviving and 9 going extinct before 2017. The area covered by surviving P. alcon populations in 2017 ranged between 0.44 and 514.11 ha (median 38.85 ha). Persistence was solely explained by host plant resources: gentian quality and gentian patch area (Table 3). Gentian quality was significantly higher and gentian patch area was significantly greater in surviving than in extinct populations (Fig. 2a and b; gentian quality t-value 4.046, p = 0.0004; gentian area t-value 2.814, p = 0.0087). In contrast, the fate of the populations after 2017, with 7 extinctions and 15 surviving populations, was explained by ant occupancy. Despite the smaller dataset, significant effects did emerge: population persistence was lower at higher Lasius occupancies whereas the host ant M. ruginodis tended to have a positive effect on P. alcon persistence (Table 3). The percentage of occupied traps of Lasius ants was significantly lower and M. ruginodis host ant occupancy was significantly higher in surviving than in recently extinct populations (Fig. 3c and d; Lasius ants t-value 2.493, p = 0.0215; M. ruginodis t-value 2.119, p = 0.0486).

Differences in habitat conditions of P. alcon populations that survived until 2022 (n = 15) or went extinct, either after or before 2017 (respectively, n = 9 and n = 7): (a) gentian quality (expressed as the percentage of gentians with > 2 flower buds on the primary stem), (b) gentian area, (c) M. ruginodis host ant occupancy and (d) Lasius competitor/predator ant occupancy. The occupancy of ants is expressed as percentage of occupied traps. Boxes show the median and 25–75% quartiles with the mean value as a cross and whiskers extending to the lowest and highest value within the 1.5 interquartile range, and outliers beyond that range
Discussion
Disentangling the various causes determining the decline of habitat specialists is of crucial importance for their conservation (Thomas et al. 2009). For P. alcon, a habitat specialist with a complex life cycle, this study shows that both habitat area and quality contribute significantly to population persistence in the Netherlands. Moreover, it appears that critical habitat factors may change over time. This can have consequences for prioritising conservation actions.
Our study analysed population persistence over two time periods 1998–2017 and 2018–2022, with the latter period characterised by severe and repeated summer droughts. For most sites, the status of P. alcon was known from yearly population monitoring. Ideally, data on habitat conditions should have been collected in multiple years over such a long period. However, this was not practically feasible. Habitat area and gentian patch area are likely to vary much less over time than host plant quality, which is strongly influenced by soil moisture (Oostermeijer et al. 1994, 1998; Wamelink et al. 2012), whilst fluctuations in host ant occupancy remain largely unknown. Here, we relied on recent data from the period 2018–2022 and early data from 1998 without data from intervening years. This should caution against overly confident conclusions on the critical factors determining the persistence of a butterfly with such a complex life cycle. Still, our findings are in line with a growing number of studies showing an increasing impact of climatic extremes on insect populations (Evans et al. 2022; Ubach et al. 2022; Termaat et al. 2023).
Changes in critical factors
The persistence of P. alcon populations in the Netherlands from 1990 till 1998 was explained earlier by both habitat area and quality, based on habitat conditions as determined in 1998 (WallisDeVries 2004). Here, we predicted population persistence from 1998 until 2017 on the basis of the same habitat conditions but now we found that only habitat area (heathland size and host plant patch size) contributed significantly. However, based on recent habitat conditions, population persistence was not only determined by host plant patch area, but also by host plant quality. For the persistence between 2017 and 2022, area factors were no longer significant, but shifted to quality factors, notably those relating to host ant availability.
It is likely that the importance of habitat area in determining population persistence has declined, because populations from the smallest areas have mostly become extinct. A few populations still survive in small host plant patches, but these are either a surviving remnant from a former metapopulation or are still part of one. Effectively, only populations located in larger nature reserves persist. There, the size of suitable host plant patches is mostly determined by landscape factors, such as geology and geomorphology allowing a stable groundwater influence and soil development (Oostermeijer et al. 1994, 1998), potentially influenced by drainage from surrounding land use. Our analysis could not explain gentian patch size from local site conditions (Fig. 4a), but this could well be due to the larger scale needed to properly assess such characteristics.

Factors explaining population persistence of P. alcon in the periods (a) 1998–2017 and (b) 2017–2022. Green boxes reflect habitat conditions with a red box for competitive or predatory Lasius ants, brown boxes reflect habitat management, blue boxes cover host plant variables, orange boxes host ants and grey boxes represent factors that did not contribute to explain population persistence. Solid lines show significant effects (p < 0.05) while dashed lines indicate marginally significant effects (p < 0.10), with blue lines showing positive effects and red lines showing negative effects
Habitat quality factors did not significantly contribute to forecasting population persistence from 1998 till 2017 on the basis of initial habitat conditions, but they became relevant when hindcasting persistence from habitat conditions in recent years. This may be due to our new focus on host plant quality in addition to area and abundance, because of emerging evidence of selective oviposition on larger gentians with more and greater buds (Kuer and Fartmann 2004; Wynhoff et al. 2014). Our analysis indicated that gentian quality is associated with later-successional vegetation and by higher soil moisture. Gentian population structure data suggest that this is because individual gentian plants become older and larger during succession (Oostermeijer et al. 1994). The effect of soil moisture was marginally significant in our case, but its importance for gentians has been well established by detailed measurements (Wamelink et al. 2012). Management actions to set back vegetation succession increase the cover of bare ground and the proportion of young gentians (Oostermeijer et al. 1994) and are thus detrimental to host-plant quality in the short term (Fig. 4a), henceforth undermining population persistence of P. alcon if carried out at a large spatial scale. Habitat desiccation by drainage or during drought periods may equally lower the survival of P. alcon populations.
During the recent drought years, however, the critical factors explaining population persistence shifted from host plants to host ants and their competitors. Although drought years also occurred four times in the period 1998–2017, these droughts were much less severe and they were always separated by several milder years (Fig. S1). This makes it likely that their impact was much less important than in the period 2018–2022, in which four out of five years had severe droughts. Lasius ants, in particular L. niger, appear to benefit from drought as well as from more bare ground after large-scale management (Fig, 4b). Their negative impact on P. alcon may arise from either increased predation of young caterpillars that emerge from their host plant or from increased competition for nest sites or food with host ants. In contrast, M. ruginodis appears to profit from soil moisture and small-scale management, combined with a sufficiently open vegetation structure (Fig. 4b), which should still provide sufficient tussocks for nesting sties (see WallisDeVries 2004). The dependence of M. ruginodis on cool, moist microclimates is well known (Seifert 2018). Indeed, its higher occupancy in 1998 on sites where P. alcon went subsequently extinct may testify of grass encroachment that was detrimental to the butterfly’s host plant (WallisDeVries 2004). Hence, it is likely that M. ruginodis is vulnerable to periods of drought, especially in fairly open vegetation. The alternative host ants did not contribute significantly to P. alcon persistence. For M. rubra, this could simply result from its rare occurrence, but M. scabrinodis was twice as abundant as M. ruginodis, without apparently benefiting P. alcon. This suggests that M. scabrinodis is not as suitable a host as M. ruginodis, which is supported by the analysis of cuticular hydrocarbon profiles by Oostermeijer et al. (in prep.).
Our study suggests negative impacts of drought extremes on the population persistence of P. alcon, but at other times extreme rainfall may also pose a risk through prolonged flooding during active periods of butterfly and larval stages. This was apparent from the earlier study of population persistence in the 1990s (WallisDeVries 2004), but we know of at least 9 sites where P. alcon populations became extinct after such periods of flooding, particularly in 2016. It can even be argued that under drought periods, suitable habitat gradually shifts to refuges in wet depressions in the landscape that are also most vulnerable to flooding incidents, increasing risk of extinction (see WallisDeVries et al. 2021).
Conservation actions
Given the increasing frequency of climatic extremes, the importance of mitigating extremes in soil moisture becomes critical. Drier surface conditions may alter both ant community composition and host plant quality at the detriment of P. alcon. The other extreme of flooding needs to be mitigated as well. Here, a landscape-scale perspective becomes necessary. While it has been shown that small populations of P. alcon can remain stable in isolation (Nowicki et al. 2019), habitat area is still likely to become critical in times of climate change. Not only are fluctuations in population size expected to become more frequent, increasing the extinction risk of small populations (Hill et al. 2021; Evans et al. 2022), but achieving the necessary improvement of habitat quality (see Jeltsch et al. 2011) also requires space. Especially the conservation and restoration of hydrological systems requires a landscape-scale approach (Von Holle et al. 2020) and this appears necessary to maintain suitable habitat for P. alcon. Moreover, larger areas can comprise more habitat patches that encompass a larger hydrological gradient. This should ensure a better resilience of the metapopulation during extreme events through risk spreading and subsequent demographic-genetic rescue (Kanarek et al. 2015; McLaughlin et al. 2017).
For the remaining populations of P. alcon, habitat quality appears to be more important than habitat area for their persistence. In terms of habitat management, the results provide further evidence supporting the recommendations by WallisDeVries (2004) to manage for heterogeneity. Host plants require bare patches to regenerate, but their suitability as host plants for the Alcon blue is highest in more developed vegetation (Oostermeijer et al. 1994). Moreover, the nests of the optimal host ant M. ruginodis are also found in established vegetation (Seifert 2018), little disturbed by small-scale management of shallow sod-cutting or choppering and perhaps benefitting from microclimatic heterogeneity. Together, these results suggest caution against both large-scale interventions and encroachment by tall vegetation that offers less heterogeneity to accommodate microclimatic preferences of both the Alcon blue and its host ants. However, beyond these crude indications we still have insufficient knowledge on the habitat requirements of M. ruginodis. We certainly require further study to elucidate impacts of climate change and nitrogen deposition on the abundance of this apparently crucial link in the ecology of P. alcon. In the meantime, managing for small-scale heterogeneity is the safest bet, to promote both its host plant and its optimal host ant.
We thus conclude that, in the face of climate change, both large-scale hydrological restoration and small-scale management for heterogeneity are key to preserve P. alcon in the Netherlands. Such a broad range of actions is likely to be equally necessary for other habitat specialists with complex life cycles from threatened habitats.
References
Als TD, Vila R, Kandul NP, Nash DR, Yen S-H, Hsu Y-F, Mignault AA, Pierce N (2004) The evolution of alternative parasitic life histories in large blue butterflies. Nature 432:386–390. https://doi.org/10.1038/nature03020
Boer P, Zegers PA, Njegowan J (2023) Mieren van de benelux. 3rd revised edition. Stichting Jeugdbondsuitgeverij, ‘s Graveland
Bos F, Bosveld M, Groenendijk D, Van Swaay CAM, Wynhoff I (2006) De Dagvlinders Van Nederland, verspreiding en bescherming (Lepidoptera: Hesperioidea, Papilionoidea). Nederlandse Fauna 7. Nationaal Natuurhistorisch Museum Naturalis. KNNV Uitgeverij European nvertebrate Survey-Netherlands, Leiden
Brereton T, Roy D, Middlebrook I, Botham M, Warren M (2011) The development of butterfly indicators in the United Kingdom and assessments in 2010. J Insect Conserv 15:139–151. https://doi.org/10.1007/s10841-010-9333-z
Bubová T, Kulma M, Vrabec V, Nowicki P (2016) Adult longevity and its relationship with conservation status in European butterflies. J Insect Conserv 20:1021–1032. https://doi.org/10.1007/s10841-016-9936-0
Caughley G, Sinclair ARE (1994) Wildlife Ecology and Management. Blackwell, Oxford
CBS PBL, RIVM WUR (2023) Trend van dagvlinders, 1992–2022 (indicator 1386, versie 19, 16 mei 2023). www.clo.nl. Centraal Bureau voor de Statistiek (CBS), Den Haag, PBL Planbureau voor de Leefomgeving, Den Haag, RIVM Rijksinstituut voor Volksgezondheid en Milieu, Bilthoven, en Wageningen University and Research, Wageningen. https://www.clo.nl/indicatoren/nl1386-dagvlinders
Cerrato C, Lai V, Balletto E, Bonelli S (2016) Direct and indirect effects of weather variability in a specialist butterfly. Ecol Entomol 41:263–275. https://doi.org/10.1111/een.12296
Clavel J, Julliard R, Devictor V (2010) Worldwide decline of specialist species: toward a global functional homogenization? Front Ecol Env 9(4):222–228. https://doi.org/10.1890/080216
R Core Team (2022) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Dennis RLH, Shreeve TG, Van Dyck H (2006) Habitats and resources: the need for a resource-based definition to conserve butterflies. Biodiv Conserv 15:1943–1966. https://doi.org/10.1007/s10531-005-4314-3
Elmes G, Thomas J, Wardlaw J, Hochberg M, Clarke R, Simcox D (1998) The ecology of Myrmica ants in relation to the conservation of Maculinea butterflies. J Insect Conserv 2:67–78. https://doi.org/10.1023/A:1009696823965
Evans LC, Melero Y, Schmucki R, Boersch-Supan PH, Brotons L, Fontaine C, Jiguet F, Kuussaari M, Massimino D, Robinson RA, Roy DB, Schweiger O, Settele J, Stefanescu C, van Turnhout CAM, Oliver TH (2022) Bioclimatic context of species’ populations determines community stability. Glob Ecol Biogeogr 8:1542–1555. https://doi.org/10.1111/geb.13527
Forister M, Pelton E, Black S (2019) Declines in insect abundance and diversity: we know enough to act now. Conserv Sci Pract 1:e80. https://doi.org/10.1111/csp2.80
Halsch CA, Shapiro AM, Fordyce JA, Nice CC, Thorne JH, Waetjen DP, Forister ML (2021) Insects and recent climate change. PNAS 118(2):e2002543117. https://doi.org/10.1073/pnas.2002543117
Hanski I, Kuussaari M, Nieminen M (1994) Metapopulation structure and migration in the butterfly Melitaea Cinxia. Ecology 75:747–762. https://doi.org/10.2307/1941732
Hill GM, Kawahara AY, Daniels JC, Bateman CC, Scheffers BR (2021) Climate change effects on animal ecology: butterflies and moths as a case study. Biol Rev 96:2113–2126. https://doi.org/10.1111/brv.12746
Jeltsch F, Moloney KA, Schwager M, Körner K, Blaum N (2011) Consequences of correlations between habitat modifications and negative impact of climate change for regional species survival. Agric Ecosyst Env 145:49–58. https://doi.org/10.1016/j.agee.2010.12.019
Kanarek AR, Webb CT, Barfield M, Holt RD (2015) Overcoming Allee effects through evolutionary, genetic, and demographic rescue. J Biol Dyn 9:15–33
Küer A, Fartmann T (2004) Prominent shoots are preferred: microhabitat preferences of Maculinea alcon ([Denis & Schiffermüller], 1775) in Northern Germany (Lycaenidae). Nota Lepid 27(4):309–319
Maes D, Vanreusel W, Talloen W, Van Dyck H (2004) Functional conservation units for the endangered Alcon Blue butterfly Maculinea alcon in Belgium (Lepidoptera: Lycaenidae). Biol Conserv 120:229–241. https://doi.org/10.1016/j.biocon.2004.02.018
Mahecha MD, Bastos A, Bohn FJ, Eisenhauer N, Feilhauer H, Hartmann H, Hickler T, Kalesse-Los H, Migliavacca M, Otto FEL, Peng J, Quaas J, Tegen I, Weigelt A, Wendisch M, Wirth C (2022) Biodiversity loss and climate extremes – study the feedbacks. Nature 612(7938):30–32. https://doi.org/10.1038/d41586-022-04152-y
McLaughlin BC, Ackerly DD, Klos PZ, Natali J, Dawson TE, Thompson SE (2017) Hydrologic refugia, plants, and climate change. Glob Change Biol 23:2941–2961. https://doi.org/10.1111/gcb.13629
Menéndez R, González-Megías A, Collingham Y, Fox R, Roy D, Ohlemüller R, Thomas C (2007) Direct and indirect effects of climate and habitat factors in butterly diversity. Ecology 88(3):605–611. https://doi.org/10.1890/06-0539
Nowicki P, Deoniziak K, Dziekanska I, Kostro-Ambroziak A, Plazio E, Rutkowski R, Sielezniew M (2019) What keeps ‘living dead’ alive: demography of a small and isolated population of Maculinea (= Phengaris) alcon. J Insect Conserv 23:201–210. https://doi.org/10.1007/s10841-018-0078-4
Oostermeijer JGB, Van’t Veer R, Den Nijs JCM (1994) Population structure of the rare, long-lived perennial Gentiana pneumonanthe in relation to vegetation and management in the Netherlands. J Appl Ecol 31:428–438. https://www.jstor.org/stable/2404440
Oostermeijer JGB, Luijten SH, Krenová ZV, den Nijs HCM (1998) Relationships between population and habitat characteristics and reproduction of the rare Gentiana pneumonanthe L. Conserv Biol 12(5):1042–1053. https://www.jstor.org/stable/2387578
Outhwaite CL, McCann P, Newbold T (2022) Agriculture and climate change are reshaping insect biodiversity worldwide. Nature 605:97–102. https://doi.org/10.1038/s41586-022-04644-x
Revelle W (2020) psych: Procedures for Personality and Psychological Research. Northwestern University, Evanston, Illinois, USA. https://CRAN.R-project.org/package=psychVersion = 2.0.12
Schaminée JHJ, Haveman R, Hommel PWFM, Janssen JAM, De Ronde I, Schipper PC, Weeda EJ, van Dort KW, Bal D (2017) Revisie Vegetatie Van Nederland. Westerlaan, Lichtenvoorde
Seifert B (2018) The ants of central and North Europe. Lutra Verlags- und Vertriebsgesellschaft, Tauer
Tartally A, Thomas JA, Anton C, Balletto E, Barbero F et al (2019) Patterns of host use by brood parasitic maculinea butterflies across Europe. Phil Trans R Soc B 374:2018202. https://doi.org/10.1098/rstb.2018.0202
Termaat T, Ketelaar R, van Kleef HH, Verberk WECP, Van Grunsven RHA, WallisDeVries MF (2023) Spearhead blues: how threats to the damselfly Coenagrion Hastulatum changed over time. J Insect Conserv. https://doi.org/10.1007/s10841-023-00537-0
Thomas JA, Simcox DJ, Clarke RT (2009) Successful conservation of a threatened Maculinea butterfly. Science 325(5936):80–83. https://doi.org/10.1126/science.1175726
Ubach A, Páramo F, Prohom M, Stefanescu C (2022) Weather and butterfly responses: a framework for understanding population dynamics in terms of species’ life-cycles and extreme climatic events. Oecologia 199:427–439. https://doi.org/10.1007/s00442-022-05188-7
Van Klink R, Bowler DE, Gongalsky KB, Swengel AB, Gentile A, Chase JM (2020) Meta-analysis reveals declines in terrestrial but increases in freshwater insect abundances. Science 368:417–420
Van Swaay C, Maes D, Collins S, Munguira ML, Šašic M, Settele J, Verovnik R, Warren M, Wiemers M, Wynhoff I, Cuttelod A (2011) Applying IUCN criteria to invertebrates: how red is the red list of European butterflies? Bio Conserv 144(1):470–478. https://doi.org/10.1016/j.biocon.2010.09.034
Vanreusel W, Van Dyck H (2007) When functional habitat does not match vegetation types: a resource-based approach to map butterfly habitat. Biol Conserv 135(2):202–211. https://doi.org/10.1016/j.biocon.2006.10.035
Vanreusel W, Maes D, Van Dyck H (2000) Soortbeschermingsplan gentiaanblauwtje: deel 1 - Hoofdrapport. Universiteit van Antwerpen (UIA-UA), Wilrijk. https://docplayer.nl/10549952-Soortbeschermingsplan.html
Von Holle B, Yelenik S, Gornish ES (2020) Restoration at the landscape scale as a means of mitigation and adaptation to climate change. Curr Landsc Ecol Rep 5:85–97. https://doi.org/10.1007/s40823-020-00056-7
Wagner DL, Grames EM, Forister ML, Berenbaum MR, Stopak D (2021) Insect decline in the Anthropocene: death by a thousand cuts. PNAS 118(2):e2023989118. https://doi.org/10.1073/pnas.2023989118
WallisDeVries MF (2004) A quantitative conservation approach for the endangered butterfly Maculinea alcon. Conser Biol 18(2):489–499. https://doi.org/10.1111/j.1523-1739.2004.00336.x
WallisDeVries MF (2014) Linking species assemblages to environmental change: moving beyond the specialist-generalist dichotomy. Basic Appl Ecol 15. https://doi.org/10.1016/j.baae.2014.05.001. :279 – 287
WallisDeVries MF, Bokelaar J, Smit JT, Versluijs R, Jansen AJM Knelpunten voor terugkeer van bedreigde insecten in het natte zandlandschap: zijn de grote gebieden hersteld? Rapportnummer 2021/OBN246-NZ, Kennisnetwerk OBN, Driebergen (2021) https://www.natuurkennis.nl/Uploaded_files/Publicaties/obn-2019-112-nz-knelpunten-terugkeer-bedreigde-insecten-eindrapport.pdf
Wamelink GWW, van Adrichem MHC, van Dobben HF, Frissel JY, den Held M, Joosten V, Malinowska AH, Slim PA, Wegman RJM (2012) Vegetation relevés and soil measurements in the Netherlands, a database. Biodiv Ecol 4:125–132. https://doi.org/10.7809/b-e.00067
Warren MS, Hill JK, Thomas JA, Asher J, Fox R, Huntley B et al (2001) Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 414:65–69. https://doi.org/10.1038/35102054
Warren MS, Maes D, Van Swaay CAM, Goffart P, Van Dyck H, Bourn NAD, Wynhoff I, Hoare D, Ellis S (2021) The decline of butterflies in Europe: problems, significance, and possible solutions. PNAS 118(2):e2002551117. https://doi.org/10.1073/pnas.2002551117
Wynhoff I (1998) The recent distribution of the European Maculinea species. J Insect Conserv 2:15–27. https://doi.org/10.1023/A:1009636605309
Wynhoff I, Bakker RB, Oteman B, Arnaldo PS, van Langevelde F (2014) Phengaris (Maculinea) alcon butterflies deposit their eggs on tall plants with many large buds in the vicinity of Myrmica ants. Insect Conserv Div 8:177–188. https://doi.org/10.1111/icad.12100
Acknowledgements
This study was funded by the Provinces of Drenthe, Gelderland, Noord-Brabant and Overijssel as well as the Cultuurfonds. We are grateful to all the site owners for granting access to their nature reserves, to the volunteers for their monitoring efforts, to Robin Satter for help with field work, to Isabel Negri and Jinze Noordijk for ant identifications and, finally, to Jan van Walsum and Hennie Halm for their work in analysing soil chemistry.
Author information
Authors and Affiliations
Contributions
MW, IW, SL and GO conceived and designed the study. MW wrote the first draft and JL, GO, IV and IW contributed to the final version of the manuscript. Field data were chiefly collected and processed by JB, AS, RM, CV and AK under supervision of MW and JL. Ant species were identified by JB, AS, RM, CV and IV. Data analysis was carried out by MW and IV.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
WallisDeVries, M.F., Bokelaar, J., Kok, A. et al. Alcon Blues: recent shifts in limiting habitat factors challenge the persistence of an endangered butterfly. J Insect Conserv (2024). https://doi.org/10.1007/s10841-024-00587-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10841-024-00587-y