
Abstract
Rapid urbanization is among the factors that decrease insect diversity. However, by offering suitable habitats, green roofs could lessen this adverse effect. Certain factors, like a nearby public garden, could be useful predictor variables to analyze to what extent green roofs can support insect communities. The study aimed to measure the insect diversity on intensive green roofs located near public gardens and on more isolated green roofs, within an urban setting. Insect species richness, abundance, and assemblages on the green roofs near public gardens differed from those on isolated green roofs. Results indicate that green roofs near public gardens will host more species, especially pollinators, consequently lessening urbanization’s negative effect. To properly understand how landscape factors impact insect communities on green roofs, future studies on green roofs’ biodiversity should consider public gardens and their influence on urban biodiversity.
Implications for insect conservation
The planning and establishment of a green roof near public gardens would probably enhance biodiversity conservation in an urban setting.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
During the last decade, studies on plants (Köhler 2006), birds (Fernández Cañero and González Redondo 2010), and various arthropod taxa including Araneae, Coleoptera (Kadas 2006), and Hymenoptera (MacIvor et al. 2015; Kratschmer et al. 2018) have supported the idea that green roofs are essential for preserving nature and promoting biodiversity in the urban setting (Hoeben and Posch 2021). For rare and endangered insect species adversely affected by urbanization, well-designed green roofs can offer secondary habitats. This has been shown by research projects examining the enormous potential of green roofs for ecological compensation: for example, in Basel (Brenneisen 2009) and London (Jones 2002; Kadas 2002).
Particularly in urban settings, public gardens generally contain a mixture of native and non-native plants, support insect species, and offer important ecosystem services. Studies by Smith et al. (2006) and Owen (2010) demonstrated that public gardens established with appropriate plant taxa can maintain high insect diversity. However, little is known about the significance of public gardens in supporting insect diversity on green roofs.
The aim of this study was to understand the relation between public gardens and insect diversity on green roofs by documenting some of the insect diversity associated with green roofs near and far from public gardens in Vienna as a first step to understanding insect distribution patterns. This will enable us to optimize the biodiversity potential of green roof designs. It is well known that plant, bird, and pollinator diversity is high in public gardens (Shwartz et al. 2014). Assuming public gardens may act as a source habitat, we expected that insect diversity would be higher on green roofs near public gardens than on more isolated green roofs.
Methodology
Study site
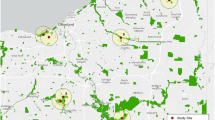
The study was conducted in the capital of Austria, Vienna (48.2082° N, 16.3738° E), which has a total area of 414.9 km2. The capital is characterized by a dry continental climate with an average precipitation of 535 mm per year. It ranges from 30 mm in the driest months (January and February) to 60 mm in the wettest ones (June and July) (Central Institute for Meteorology and Geodynamics 2016). We selected eight intensive green roofs (they have greater substrate depth than the extensive green roofs) having comparable vegetation structures (grasses like Andropogon gerardii, Bouteloua gracilis, Schizachyrium scoparium were commonly found on intensive green roofs; Supplementary Materials S1). The ages of the selected green roofs ranged between 10 and 30 years. Of these, four were located close to public gardens (within a 500-m radius) while the other four were more isolated (distance to the next public garden ranged from 2 to 5 km; Fig. 1). We chose a 500-m radius because roads may act as barriers to the movement of bees and wasps, especially for small species with poor dispersal ability (Andersson et al. 2017), and eventually restrict their activity range. In this study, public gardens are defined as urban green spaces containing ornamental and native plant species (Rakow 2011). The selected public gardens were highly managed for recreation. Thus, they comprised several old and large trees, large lawns that cover at least one-third of the garden area, and several highly managed flowerbeds with native and non-native ornamental flowers. They also contained some children’s recreation areas. The selected public gardens were of similar size (∼1 ha) and spatially independent of each other, typically separated by large buildings and streets. The green roofs studied contained a range of biophysical conditions, including variations in roof area (61–399 m2) and roof height (17–107 m; Supplementary Materials Table A2).

Map of green roofs near public gardens and far from public gardens. The color codes represent the different habitat classes (Land Use of Vienna: modified after Pendle et al. 2022)
Insect sampling
In 2021, insect collection was performed by a semi-quantitative and hand netting method on each green roof (Kratschmer et al. 2018). Insect sampling was conducted in four sessions of 60 min in each month (May to August), when weather conditions were suitable (warm, windless, and dry). Within each session, we actively searched the green roof and documented all observed insect species. Only insects that were observed to interact with the green roof were recorded. The insect taxa ant, bumblebee, heteropteran bug, syrphid, wasp, and wild bee were collected and afterwards prepared and identified to species level using relevant literature (e.g., Wagner 1967; Schmid-Egger and Scheuchl 1997; Gokcezade et al. 2010; Veen 2010). Most bumblebee individuals were identified on site and released afterwards.
Statistical analysis
For analysis, the numbers of species and individuals among all insect taxa were summed from multiple observations per month. The Shapiro test and QQ plots were used to examine the data for normal distribution. The Bartlett test was used to assess homoscedasticity, or homogeneity of variance. To evaluate differences between response variables (richness and abundance) and predictor variables (habitat type: green roof near and far from a public garden), we performed analysis of variance (ANOVA).
To assess differences in species assemblage, we calculated PERMANOVA (Bray-Curtis dissimilarities, 999 permutations) using the function adonis from the R package vegan (as performed in Hussain et al. 2022). The betadisper function was used to check the initial data for equal multivariant dispersion. Later, multilevel pairwise comparison was applied using pairwise.adonis to evaluate the species assemblage differences between green roofs near and far from public gardens. Further, an ordination plot of principal component analysis was made to visualize the species assemblage patterns. For statistical analysis, R software (version 3.5.1) was used (R Core Team 2018).
Results
A total of 124 individuals from the 6 insect taxonomic groups were observed. In total, 106 individuals were observed on green roofs near public gardens and 18 individuals far from public gardens. There were 21 wild bee and eight syrphid species, and two species each of ants, bumblebees, heteropteran bugs, and wasps, totaling 37 species (Supplementary Materials Table A1). The numbers of individuals (ANOVA: F = 8.948, p = 0.0243; Table 1; Fig. 2) and species (ANOVA: F = 6.818, p = 0.0401; Table 1; Fig. 1) were significantly higher on green roofs near public gardens than on the more isolated green roofs. Insect species assemblages differed significantly between green roofs with public gardens and the more isolated green roofs (PERMANOVA: R2 = 0.15, p = 0.0385; Table 2; Fig. 3).

Number of individuals and species on green roofs near (located within 500 m) and far (located more than 500 m) from a public garden. Boxplots show the medians, the 25% and 75% percentiles, and the 10% and 90% percentiles, and notches. Different letters (a, b) indicate significant differences between studied roof gardens

Principal component analysis based on species assemblages of observed insects on green roofs
Discussion
Green roofs near public gardens showed 50% more species and total individual insects compared with isolated green roofs. Insect abundance may increase on green roofs over time (Kadas 2006), although this is not always the case (Kyrö et al. 2018). All insect groups were present on green roofs, with generalists (that can live in diverse environments) being the most abundant and diverse. Our findings imply that green roof location should be considered prior to installation in order to preserve biodiversity, trophic level interactions, and ecosystem services in urban settings (Lundholm 2015).
In general, rapid urbanization is associated with a loss of species diversity and homogenization of insect assemblages (Groffman et al. 2017). The impacts of different local site attributes and habitat quality on insect assemblages are significantly more complex at the scale of a yard, park, or neighborhood (Adams et al. 2020). In general, insect responses to green roofs indicate that the character of the nearby urban setting is likely an important predictor of local insect assemblage structure, especially for wild bees, a group that is highly mobile and might not be hampered by barriers (Haskell 2000; Baxter-Gilbert et al. 2015; Muñoz et al. 2015).
Diverse vegetation in public gardens usually presents more suitable habitat conditions, is higher, and/or offers more resources than green roofs (Holt 2016). This is in line with studies arguing that habitats with greater plant diversity enhance the growth and activity of natural enemies (Letourneau et al. 2011; Hussain et al. 2021). Even though the predictions made by these studies are frequently observed in agricultural systems (Landis et al. 2000; Maas et al. 2021), the influence of plant diversity on insect habitat quality has rarely been considered in regard to green roofs. This study’s results indicate that, for the six observed insect taxa, the resources provided by a diverse plant community in nearby public gardens might be more relevant (mentioned in Schunko et al. 2021), as many generalist insect species can thrive in the small habitats of nearby green roofs.
In the past, researchers thought that only the most migratory insect species could make use of the ecosystems found on green roofs (Dunnett and Kingsbury 2008). We discovered a similar pattern, in which numerous insects, including medium, large, and even flightless insects, spontaneously populated the green roofs. The source habitat of such insect groups could be the nearby public gardens, because the literature shows that these insect groups—especially carabids and heteropteran bugs, which are sensitive to environmental stress—are found predominantly at the soil surface of ground level urban habitats (Niemelä et al. 2000).
We present preliminary results because we acquired just four replicates, leading to a comparably low number of observed individuals. The current study has a few caveats that should be considered. For instance, the sampling of insects was done only on green roofs and according to a specific method, because of the study’s preliminary character and limited funding. Additional sampling methods with increasing sampling duration can help to determine how many insect species are actively using green roofs. Each isolated green roof was in close proximity to some green spaces that could have influenced its biodiversity. Further, factors like urban construction and green roof structure (e.g., vegetation mixture, type of soil, soil depth, construction age, and specific maintenance) may also affect green roof biodiversity (Andersson et al. 2017). Future studies considering these limitations would be valuable in fully understanding how nearby landscape conditions shape insect communities on green roofs. However, our study provides additional information with which to build strategies for biodiversity conservation in the urban setting.
References
Adams BJ, Li E, Bahlai CA, Meineke EK, McGlynn TP, Brown BV (2020) Local-and landscape‐scale variables shape insect diversity in an urban biodiversity hot spot. Ecol Appl 30:e02089
Andersson P, Koffman A, Sjödin NE, Johansson V (2017) Roads may act as barriers to flying insects: species composition of bees and wasps differs on two sides of a large highway. Nat Conserv 18:47–59
Baxter-Gilbert JH, Riley JL, Neufeld CJ, Litzgus JD, Lesbarrères D (2015) Road mortality potentially responsible for billions of pollinating insect deaths annually. J Insect Conserv 19:1029–1035
Brenneisen S (2009) Ökologisches Ausgleichspotenzial von Extensiven Dachbegrünungen: Bedeutung des Ersatz-Ökotops für den arten-und Naturschutz und die Stadtentwicklungsplanung. Selbstverlag der Abteilung Physiogeographie der Universität Basel
Central Institute for Meteorology and Geodynamics (2016) Zentral-anstalt für Meteorologie und Geodynamik. https://www.zamg.ac.at/cms/en/. Accessed 01 September 2022
Dunnett N, Kingsbury N (2008) Planting green roofs and living walls. Timber press, Portland, OR
Fernández Cañero R, González Redondo P (2010) Green roofs as a habitat for birds: a review. J Anim Vet Adv 9:2041–2052
Gokcezade J, Gereben-Krenn B-A, Neumayer J, Krenn H (2010) Feldbestimmungsschlüssel für die Hummeln Österreichs, Deutschlands und der Schweiz (Hymenoptera, Apidae). Linzer Biologischer Beitrag 42:5–42
Groffman PM, Avolio M, Cavender-Bares J, Bettez ND, Grove JM, Hall SJ, Hobbie SE, Larson KL, Lerman SB, Locke DH, Heffernan JB (2017) Ecological homogenization of residential macrosystems. Nat Ecol Evol 1:1–3
Haskell DG (2000) Effects of forest roads on macroinvertebrate soil fauna of the southern Appalachian Mountains. Conserv Biol 14:57–63
Hoeben AD, Posch A (2021) Green roof ecosystem services in various urban development types: a case study in Graz, Austria. Urban for Urban Green 62:127167
Holt RD (2016) Green roofs may cast shadows. Isr J Ecol Evol 62:15–22
Hussain RI, Brandl M, Maas B, Rabl D, Walcher R, Krautzer B, Entling MH, Moser D, Frank T (2021) Re-established grasslands on farmland promote pollinators more than predators. Agric Ecosyst Environ 319:107543
Hussain RI, Brandl M, Maas B, Krautzer B, Frank T, Moser D (2022) Establishing new grasslands on crop fields: short-term development of plant and arthropod communities. Restor Ecol 30:1–5
Jones RA (2002) Tecticolous invertebrates: a preliminary investigation of the invertebrate fauna on green roofs in urban London, 3 edn. English Nature, London
Kadas G (2002) Study of invertebrates on green roofs-How roof design can maximise biodiversity in an urban environment. Unpublished mater’s thesis, University College, London, England
Kadas G (2006) Rare invertebrates colonizing green roofs in London. Urban Habitats 4:66–86
Köhler M (2006) Long-term vegetation research on two extensive green roofs in Berlin. Urban Habitats 4:3–26
Kratschmer S, Kriechbaum M, Pachinger B (2018) Buzzing on top: linking wild bee diversity, abundance and traits with green roof qualities. Urban Ecosyst 21:429–446
Kyrö K, Brenneisen S, Kotze DJ, Szallies A, Gerner M, Lehvävirta S (2018) Local habitat characteristics have a stronger effect than the surrounding urban landscape on beetle communities on green roofs. Urban for Urban Green 29:122–130
Landis DA, Wratten SD, Gurr GM (2000) Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu Rev Entomol 45:175–201
Letourneau DK, Armbrecht I, Rivera BS, Lerma JM, Carmona EJ, Daza MC, Escobar S, Galindo V, Gutiérrez C, López SD, Mejía JL (2011) Does plant diversity benefit agroecosystems? A synthetic review. Ecol Appl 21:9–21
Lundholm JT (2015) Green roof plant species diversity improves ecosystem multifunctionality. J Appl Ecol 52:726–734
Maas B, Brandl M, Hussain RI, Frank T, Zulka KP, Rabl D, Walcher R, Moser D (2021) Functional traits driving pollinator and predator responses to newly established grassland strips in agricultural landscapes. J Appl Ecol 58:1728–1737
MacIvor JS, Ruttan A, Salehi B (2015) Exotics on exotics: Pollen analysis of urban bees visiting Sedum on a green roof. Urban Ecosyst 18:419–430
Muñoz PT, Torres FP, Megías AG (2015) Effects of roads on insects: a review. Biodivers Conserv 24:659–682
Niemelä J, Kotze J, Ashworth A, Brandmayr P, Desender K, New T, Penev L, Samways M, Spence J (2000) The search for common anthropogenic impacts on biodiversity: a global network. J Insect Conserv 4:3–9
Owen J (2010) Wildlife of a garden: a thirty-year study. Royal Horticultural Society
Pendl M, Hussain RI, Moser D, Frank T, Drapela T (2022) Influences of landscape structure on butterfly diversity in urban private gardens using a citizen science approach. Urban Ecosyst 25:477–486
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing. https://www.r-project.org/
Rakow DA (2011) What is a public garden? In Public garden management, edited by D. A. Rakow and S. A. Lee, 3–14. Hoboken, NJ: Wiley
Schmid-Egger C, Scheuchl E (1997) Illustrierte Bestimmungstabellen der Wildbienen Deutschlands und Österreichs, vol 3. Andrenidae. Eigenverlag, Velden
Schunko C, Wild AS, Brandner A (2021) Exploring and limiting the ecological impacts of urban wild food foraging in Vienna, Austria. Urban for Urban Green 62:127164
Shwartz A, Turbé A, Simon L, Julliard R (2014) Enhancing urban biodiversity and its influence on city-dwellers: an experiment. Biol Conserv 171:82–90
Smith RM, Warren PH, Thompson K, Gaston KJ (2006) Urban domestic gardens (VI): environmental correlates of invertebrate species richness. Biodivers Conserv 15:2415–2438
Veen MPV (2010) Hoverflies of Northwest Europe: identi fica-tion Keys to the Syrphidae. KNNV, Utrecht, p 256
Wagner E (1967) Die Tierwelt Deutschlands und der angrenzenden Meeresteile nachihren Merkmalen und nach ihrer Lebensweise, Wanzen oder Heteroptera II,Cimicomorpha. – Fischer, Jena
Acknowledgements
We would like to thank the Department of Culture of the City of Vienna (MA 7) for providing financial support for the study. We acknowledge the caretakers of the buildings for providing flexible access to the study roofs, and thank Elisabeth Gruchmann for helpfully suggesting specific green roofs during study site selection.
Funding
Open access funding provided by University of Natural Resources and Life Sciences Vienna (BOKU).
Author information
Authors and Affiliations
Contributions
Raja Imran Hussain led the data collection and analysis as well as the writing of the manuscript, Sophie Kratschmer supported bee’s identification, and Thomas Frank helped revise the manuscript. All authors contributed critically to the manuscript and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Consent to participate
The authors have provided their consent to participate to the work.
Consent for publication
The authors have provided their consent for publication.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hussain, R.I., Frank, T. & Kratschmer, S. More insect species are supported by green roofs near public gardens. J Insect Conserv 27, 941–946 (2023). https://doi.org/10.1007/s10841-023-00510-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-023-00510-x