
Abstract
Fermentation is an ancient technique for preserving food and feed, and for moderating taste and texture of foods. Fermentation of seaweeds for generating novel food products has yet only been described for few red algae. Here, sugar kelp (Saccharina latissima) was heat-treated and fermented using lactic acid bacteria (LAB). Taste, smell and texture of the fermented product was compared to fresh sugar kelp and two commercial seaweed products (nori and wakame). Tissue contents of dry matter, nitrogen, mannitol, and selected minerals and trace metals of the fresh and fermented sugar kelp were quantified and compared. In the fermentation process, the pH was reduced to 4.5 within 40 h, with LAB counts increasing 100-fold and no Bacillus cereus present. Heat-treatment and fermentation caused a reduced saltiness and umami flavour of the sugar kelp, a less slimy visual appearance and a reduced smell of sea, whereas the texture and protein content was unchanged compared to the fresh sugar kelp. The fermented sugar kelp had a stronger bite than nori and wakame, a stronger smell of sea and a more salty, irony and umami rich taste than nori, but less umami and salt taste than wakame. The fermentation process reduced the contents of sodium (− 15%), cadmium (− 35%) and mercury (− 37%) in the sugar kelp. LAB fermentation of sugar kelp showed promising for broadening the food market for seaweeds as the fermented product had a milder taste, improved visual impression and smell, and a reduced content of harmful trace metals.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Fermentation is an ancient and widely used method for preservation of perishable foods, i.e. milk, cereals, meat and vegetables, hereby producing new foods or beverages. The process makes use of the microbial conversion of sugars to acids such as lactic acid, acetic acid and propionic acid or to ethanol. For fermentation of foods, lactic acid bacteria (LAB) are the most commonly used and characterised microorganisms (Caplice and Fitzgerald 1999). Lactic acid fermentation not only improves the shelf life and food safety of the end-product but also alters the nutritional and sensory properties of the product (Caplice and Fitzgerald 1999; Karovicova and Kohajdova 2005) and promotes health benefits through increasing the content of probiotic bacteria (Gupta and Abu-Ghannam 2012).
Seaweeds are highly nutritious seafood products that are commonly used in a broad range of Asian foods, and having the potential for becoming a more central element in human diets also in the Western world (Chapman et al. 2015). On a global scale, 30.1 million wet tonnes of seaweeds are produced annually (2016), representing a value of 11.7 billion US $ (FAO 2018). More than 99% of this is produced in Asia (FAO 2016); however, the cultivation of seaweeds for food, energy and biomolecules in Europe is increasing (Kerrison et al. 2015; Kim et al. 2017). Fermentation of seaweeds offers several potential benefits: (1) a resource efficient method for stabilising a wet biomass that rapidly degrades upon harvest (Enriquez et al. 1993); (2) improvement of food safety and shelf life of non-dried products; (3) refining of sensory properties, with options for innovation in seaweed-based food products; and ultimately (4) providing a source of non-dairy probiotic food of which the demand is increasing (Gupta and Abu-Ghannam 2012). Ensiling of seaweeds as a means for stabilising of the biomass and as a pre-treatment prior to production of biogas has been investigated using spontaneous fermentation as well as fermentation induced by addition of LAB (Herrmann et al. 2015; Redden et al. 2017). Ensiling proved a less resource-demanding alternative to the drying of seaweeds (Black 1955), as well as a means for combined stabilisation and pre-treatment preparing and optimising the biomass for energy conversion or extraction of high value compounds (Herrmann et al. 2015; Milledge et al. 2016; Milledge and Harvey 2016a, b). LAB fermentation of seaweeds has also been applied for production of Marine Single Cell Detritus (MSCD) feed for use in aquaculture of i.e. sea urchins and larval stages of shrimps, fish and pearl oysters, representing a cheap and health promoting supplement to microalgae (Uchida and Murata 2002; Uchida et al. 2004; Felix and Pradeepa 2011). Despite the long history of fermentation technology in food production, only few seaweed-based fermented food products have yet been developed (Uchida et al. 2017). The protein-rich red seaweed Pyropia yezoensis (nori) formed the basis of two high-salt sauces produced by fermentation using halophilic LAB (Uchida et al. 2014, 2017). The nori sauces had a unique taste due to the composition of the free amino acids and contained more vitamin B1, B12 and folic acid as compared to soy sauce. Food safety wise, no allergy-causing agents were detected in the fermented seaweed sauces and their contents of harmful metals were below limit values (Uchida et al. 2017). In addition, LAB fermented nori can contain compounds that have beneficial effects on the cardiovascular system (Tsuchiya et al. 2007; Uchida et al. 2017). Regarding brown algae, LAB fermentation by Lactobacillus plantarum proved feasible with two common North Atlantic kelps: Saccharina latissima and Laminaria digitata following heat-treatment, whereas the fucoid species Ascophyllum nodosum did not support growth of L. plantarum (Gupta et al. 2011a). Heat-treatment of the two kelp species followed by fermentation with Lactobacillus rhamnosus resulted in leaching of antioxidants to the fermentation broth, as well as to an increase in the content of phenolic compounds in the seaweed biomass. This emphasises the potential for developing seaweed-based functional foods (Gupta et al. 2012).
Sugar kelp, S. latissima, is the most commonly cultivated seaweed species in Europe (Fernand et al. 2017; Kerrison et al. 2017). Being rich in minerals, and having a salty, umami-rich flavour, this large brown algae species is attractive for use in food applications, either on its own as a sea vegetable, or as a food ingredient or flavour enhancer (Mouritsen et al. 2012; Chapman et al. 2015). So far, no studies have characterised the effect of LAB fermentation on the sensory properties and food safety of S. latissima or other commonly cultivated kelps.
With the perspective of developing novel food products, the aim of this study was to analyse the effect of LAB fermentation on the taste, smell, visual appearance, and content of protein and specific minerals and metals of sugar kelp (Saccharina latissima).
Materials and methods
Sugar kelp raw material
Freshly harvested, drained and frozen sugar kelp was obtained in 12 kg blocks from Ocean Rainforest Sp/F, Faroe Islands (product number 101120; Batch number A71-1-1-0605) harvested in June. Only biomass from June was studied since early June is a typical time of harvest of S. latissima in Denmark). Upon arrival at the research kitchen at Erhvervsakademi MidtVest (EAMV), the frozen sugar kelp was size-reduced to fragments of approximately 0.5 × 2 cm using an industrial flash cutter (Mado MTK560 D) and frozen at − 20 °C in smaller portions of 200 g. Minor amounts of thawing water were included evenly in the frozen portions of seaweed.
Heat-treatment and fermentation
The size-reduced sugar kelp was defrosted and mixed with cooled, boiled water (batches of 200 g of sugar kelp with 750 mL water). Following it was heated to 95 °C for 15 min and then cooled to 37 °C.
The heat-treated sugar kelp was poured into a sealed fermentor with a total volume of 1.0 L (Biostat Aplus MO, Aage Christensen A/S, Denmark). To each batch, 3 g of starter culture of Lactobacillus plantarum (HOLDBAC™ Listeria 10 IP, DuPont) was added according to the manufacturer’s specifications, in order to achieve an initial concentration of 1 × 106 colony forming units (CFU) per gram fresh weight (FW) of seaweed. According to the product description, the starter culture contained dextrose as a carrier for the LAB. The concentration of dextrose in the starter culture was not clear from the product information, but constituted maximally 3.0 g L−1of each batch of fermentation mix, as 3 g of starter culture was added to 1 L of fermentation mix (seaweed + water). The fermentation process ran at 37 °C for approximately 48 h until reaching a pH of 4.5. The fermentation was repeated three times with three individual batches of sugar kelp. During the fermentation process, the pH was measured every 5 min using the pH monitor of the fermentor. Samples for counting of LAB were taken every 24 h.
Taste
Training and sensory analyses were planned and carried out according to the international standard recommendations (ISO 2005).
Basis training and selection of tasting panel
The members of the tasting panel were selected from among the students at Process Technology at EAMV on the basis of five training sessions with focus on assessing the five basic tastes using the geometric dilution series. The following methods were applied: recognition, dilution and triangular estimation around the lower threshold values (ISO 2005).
A final tasting panel of 13 assessors was selected from the original 50 students tested. All selected assessors had threshold values of all basic tastes of G3 or lower, and further, the selected assessors had the highest frequency of correct evaluation in the tests applied for selection.
Descriptive test
A Sensory Profile analysis was carried out as a Quantitative Descriptive profile as according to international standards (ISO 2016).
The tasting panel was involved in developing the terminology of the flavour descriptors (method 2 (ISO 2016)). Based on tasting of a number of different seaweed samples, a set of 100 attributes was produced to describe the taste, smell, texture and visual appearance of seaweed (Table 1). Following, the 100 attributes were reduced to 11 key attributes (Table 1).
Simultaneous to the development of the flavour descriptors, a common list of attributes was developed. Hereafter, the panel took part in a session where the evaluation scale for each attribute was determined and described. The list of key attributes, the evaluation guidelines and the scales were adjusted during three following training sessions, and hereafter, the panel performed a test evaluation, comparing the tastes of fresh and fermented sugar kelp. Based on an analysis of the variance of these results, final adjustments were made to the list of key attributes, the evaluation guidelines and the scale.
Taste assessments
Thirteen selected members of the taste panel participated in the final taste assessments (ISO 2005). All samples were assessed three times. The samples were served one at a time in a randomised fashion in small neutral plastic trays, coded with random three-digit numbers. Samples were all at room temperature when served. The samples were evaluated using a 15-cm unstructured linear scale, with endpoints at 0.5 cm from each end. The taste assessment included the three experimental treatments of sugar kelp (fresh, heat-treated and fermented), and dried samples of three other seaweed species: wakame (Undaria pinnitifida), bladderwrack (Fucus vesiculosus) and nori (Pyropia sp.) (Table 2).
Analyses of chemical composition
For all analyses, the seaweed samples from the different treatments were freeze-dried at − 40 °C and following homogenised by dry milling to a particle size of < 500 μm. In general and unless specifically mentioned, triplicate independent samples were analysed and results are given as average ± standard error (SE), n = 3.
Dry matter (DM)
The seaweed material from each of the three treatments (fresh, heat-treated and fermented) was drained for 15 min and then 3 g of the drained seaweed was freeze-dried as described above and used for analysis of DM content. Triplicate samples were analysed for each treatment, and the dry matter content was calculated as percentage of FW.
Carbon (C), nitrogen (N) and protein
Concentrations of C and N in the freeze-dried algae tissue were determined by Pregl-Dumas ignition in pure oxygen atmosphere, followed by chromatographic separation of C and N with detection of the individual elements by thermal conductivity (Marco et al. 2002).
Protein content was following estimated from the N content using an N to protein conversion factor of 5 (Angell et al. 2016).
Mannitol
Triplicate samples (500 mg) of the dried material from each treatment were each added to 5 mL of ethanol (50%) and heated at 70–80 °C for 1 h in a water bath. The material was then centrifuged at 956×g for 10 min. In the following derivation process, the extract supernatant and a reference standard of D-mannitol (Sigma-Aldrich, Denmark) were treated in parallel. All reagents were of analytical grade. First, 500 μL of the supernatant/standard was transferred to a vial, cooled to − 80 °C and freeze-dried (CHRIST LOC-1, VWR International). Then, 40 μL dimethylsulfoxide (DMSO) was added and the samples were heated in a water bath to 75 °C and repeatedly shaken until sugars were fully dissolved. Following, 500 μL pyridin reagent including internal standards of phenyl-β-D-gluco-pyranoside (1 mg mL−1) and hydroxylamin hydrochloride (25 mg mL−1) was added and samples were heated to 75 °C for 30 min while vigorously shaken. Hereafter, 500 μL of hexamethyldisiazane (HMDS) and three drops of trifluoroacetic acid were added and the samples were again heated to 75 °C for 30 min. The mannitol content of the samples was following analysed using a Gas Chromatograph (GC) (Agilent Technologies, 6890N, Network GC System), equipped with a flame ionisation detector. The column was a 10 m Chrompack CP-SIL 5 CB, 100% dimethylpolysiloxane, with ID 0.25 mm and 0.12-μm film thickness. Operation conditions were as follows: injection port 360 °C, splitless manual injection, detector 360 °C, column gradient program: 0 min 145 °C, 0–7.5 min 2 °C min−1 to 160 °C, 7.5–12 min 20 °C min−1 to 250 °C, 12–15 min 12 °C min−1 to 285 °C. The N carrier flow rate was 4 mL min−1. The sample volume was 1 μL. Data were acquired with Agilent Technologies, ChemStation, and the mannitol content of each sample was calculated using the following equation:
where Amannitol and Ais are peak areas, Rmannitol and Ris are response factors, and Wsugar kelp and Wis are the DM content of the sugar kelp and the internal standard, respectively.
Minerals and metals
Concentrations of the following minerals and metals were determined by Inductively Coupled Plasma Mass Spectrometry (ICP-MS): iron (Fe), iodine (I), potassium (K), magnesium (Mg), sodium (Na), selenium (Se), zinc (Zn), total arsenic (As), cadmium (Cd), mercury (Hg) and lead (Pb). In brief, a 0.2g dry sub-sample was digested in a closed vessel microwave oven using 5 mL of nitric acid (7 M) and 1 mL of hydrogen peroxide, then diluted to 50 mL with milliQ water, followed by ICP-MS determination using internal standards of Rh, Ir and Ge to correct for drift as in Nielsen et al. (2012). Certified reference material of seaweed from IAEA-140 (Coquery et al. 2000) was used for quality assurance. Triplicate samples were analysed for each treatment. The molar Na:K ratio of the samples was calculated.
Microbiology
All microbiological analyses were performed using the liquid phase of the experimental treatments extracted following a thorough mixing, assuming that the mixing caused the sampling to be representative of the microflora on the seaweed. From each experimental series, duplicate samples were analysed for all treatments (fresh, heat-treated and fermented sugar kelp).
Counts of lactic acid bacteria were performed according to ISO 15214 (1998) with the following modifications: 1 mL of sample was inoculated in melted and cooled De Man, Rogosa and Sharpe (MRS) agar (Oxoid. CM 1153. Thermo Fisher Scientific) and following incubated in an aerobe environment at 37 °C for 48 h.
Counts of Bacillus spp. were performed according to ISO 15214 (1998), with the following modifications: A sample of 0.1 mL was inoculated on to Bacillus cereus Selective agar (Oxoid CM0617, Thermo Fisher Scientific), added Polymyxin (Oxoid, Thermo Fisher Scientific), and following incubated in an aerobe environment at 37 °C for 24 h. Following, counting of all colonies, regardless of morphology, was attempted and identification of four representative colonies from each sample was made using API CHB medium—Identification of Bacillus (bioMérieux).
Statistics
Normal distribution and equality of variance of data regarding sensory properties was confirmed, and following the results were analysed using a two-sided analysis of variance according to ISO (2016).
Data for the chemical composition were analysed using t test in JMP, version 12.1.0. (SAS Institute Inc., USA). Unless specifically mentioned, the level of significance applied was 0.05.
Results
Fermentation process
Generally, 36 h after addition of the starter culture to the sugar kelp, the pH was in the range of 4.5 to 5, stabilising at a value of 4.5 after 40 h (Fig. 1a). During the first 48 h of fermentation the total count of LAB increased by a factor of 100 from 106 to 108 CFU mL−1 (Fig. 1b). The concentration of mannitol decreased by a factor of 9 from 161.4 ± 23.4 mg L−1 in the heat-treated seaweed to 18.8 ± 3.4 mg L−1 in the fermented seaweed.

The fermentation process. a pH during the process. b Total counts of lactic acid bacteria in the liquid phase of the fermented kelp at the beginning of fermentation, and at 24 and 48 h later (note that the y-axis is a log scale). Data are given as average ± standard error (n = 3)
Taste, smell and visual impression
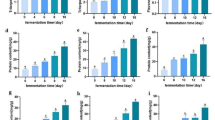
The 11 key attributes of sensory characteristics of the sugar kelp were: “Slimy” and “yellow green” for the visual appearance; “smell of sea” for the smell; and “iron” and “boiled green vegetables” in addition to the 5 fundamental tastes; “sour”, “sweet”, “salty”, “bitter” and “umami”. In addition, the “bite” of the seaweed was selected to evaluate the texture. Comparing the taste, smell and visual appearance of the fresh sugar kelp to the treated sugar kelp, the heat-treatment caused a significant decrease in the taste components of salt and umami, and resulted in a significantly less slimy visual appearance of the sugar kelp (Fig. 2. Table 3). Adding to this, the fermentation process caused a significant reduction of the sea smell of the sugar kelp (Fig. 2, Table 3). None of the two treatments affected the taste components of sour, iron, sweet, bitter or boiled green vegetables. The texture also remained unchanged through the treatments (Fig. 2). Comparing the fermented sugar kelp to commercial seaweed products (wakame and nori) as well as to fresh bladderwrack, the fermented sugar kelp had a stronger bite than the commercial seaweeds, a stronger smell of sea than nori, a more salty, irony and umami rich taste than nori and bladderwrack, but less umami and salt taste compared to wakame (Fig. 3, Table 3).
Chemical composition
Dry matter
The DM content of the fresh sugar kelp was 6.2 ± 0.2% of FW (Fig. 4a). Heat-treatment significantly decreased the DM content by 24% to 4.7 ± 0.2% of FW (p = 0.04, t test, JMP). However, the fermentation increased the DM content of the heat-treated sugar kelp product by 35% (p = 0.02) to 6.3 ± 0.2% of FW, resulting in no significant difference in DM content between the fresh and the fermented sugar kelp.
Nitrogen, protein and carbon content
The N content did not change significantly with neither the heat-treatment nor the fermentation. The N content in sugar kelp was 3.1 ± 0.1% of DM, 3.3 ± 0.2% of DM and 3.2 ± 0.1% of DM in the fresh, heat-treated and fermented sugar kelp, respectively. Converting the N content to protein using a universal conversion factor of 5 (Angell et al. 2016), the N content reflected a protein content of 15.6 ± 0.3% of DM, 16.3 ± 1.1% of DM and 15.9 ± 0.4% of DM in the fresh, heat-treated and fermented sugar kelp, respectively (Fig. 4b). The C content also did not change significantly with heat-treatment or fermentation: The C content was 31.6 ± 0.2%, 32.8 ± 1.8% and 33.4 ± 0.6% of DM, in the fresh, heat-treated and fermented sugar kelp, respectively (Table 4).
Mannitol
The content of mannitol was unchanged from the fresh to the heat-treated sugar kelp (1.43 ± 0.24 and 1.30 ± 0.15% of DM, respectively), but decreased significantly during the fermentation process to 0.12 ± 0.06% of DM in the fermented sugar kelp (p = 0.03) (Fig. 4c).
Minerals and metals
The heat-treatment and fermentation processes had different effects on the content of different minerals and metals (Table 4): The heat-treatment reduced the content of Na in the sugar kelp biomass, and following the fermentation further reduced the Na content to 15% less than in the fresh sugar kelp. The content of Mg also was significantly reduced in the fermented, as compared to the fresh sugar kelp (by 21%). The contents of the two harmful metals, Hg and Cd, were not affected by the heat-treatment, but were both reduced in the fermentation: Hg by 37% and Cd by 35%, as compared to the fresh sugar kelp. The content of iodine (I) also was reduced in both the heat-treatment and further in the fermentation; however, the reduction was not significant due to large variation. The concentrations of the remaining minerals analysed for did not change significantly between the fresh and the fermented products (As, Fe, K, Pb, Se and Zn).
Microbiology
No colonies with morphology of B. cereus were observed in any of the treatments.

Development in taste, smell, texture and visual appearance of sugar kelp during processing steps from fresh sugar kelp over heat-treatment to fermented sugar kelp. Heat-treatment significantly reduced the taste components of umami and salt, as well as the slimy visual appearance, whereas fermentation significantly reduced the sea smell

Evaluation of taste, smell, texture and visual appearance, comparing fermented sugar kelp and other seaweed products: wakame, nori and fresh bladderwrack

Changes in nutritional quality with the processing steps of heat-treatment and fermentation of sugar kelp. Content of a DM (% of FW), b protein (estimated from total nitrogen content (% of DM), and c mannitol in the fresh, heat-treated and fermented sugar kelp. Data are given as average ± standard error (n = 3)
Discussion
The processes of heat-treatment and LAB fermentation of the sugar kelp had an overall positive impact on the stabilisation of the sugar kelp biomass, the sensory characteristics and the nutritional value as well as the food safety of the final seaweed product.
Stabilising the biomass
LAB fermentation proved an efficient method for stabilising the sugar kelp biomass. By using a defined and competitive LAB strain as starter culture, the pH was reduced from 6.5 to 4.5 within 40 h. The process of adding LAB, together with water soluble carbohydrates (dextrose) as a carrier, to the heat-treated biomass generated a considerably faster decrease in pH, comparing to ensilation of S. latissima relying on natural LAB and carbohydrates present in the seaweed biomass, where a pH of 4.5 was not achieved until after approximately 10 days (Herrmann et al. 2015). Fresh seaweeds naturally contain relatively low numbers of LAB (102 CFU g−1) (Uchida et al. 2004; Herrmann et al. 2015). Thus, it is necessary, as done in this study, to add LAB to an initial concentration of 106 CFU mL−1 in order to control the fermentation process and suppress growth of yeasts or potentially harmful bacteria, such as Clostridia (Pahlow et al. 2003). The fermentation coefficient (FC) suggested by Weissbach (1996) describes the ensilability of a biomass by the equation: FC = TS + 8 WSC/BC, taking into account the total solid content (TS (g 100 g−1), same as DM), the concentration of water soluble carbohydrates (WSC (g kg−1 TS)) and the buffering capacity (BC (g lactic acid kg−1 TS)) of the biomass. Values of FC above 45 have been suggested to indicate good ensilability of a biomass. Saccharina latissima has been described to be the brown algal species best suitable for fermentation, in having FCs above the critical 45, due to a low buffering capacity, an adequate level of total solids concentration and a sufficient content of water soluble carbohydrates as fermentable substrate (Herrmann et al. 2015). However, these parameters all strongly depend on the time of harvest of the seaweed biomass, with the optimal FC most likely obtained for biomass harvested in (late) summer as in Herrmann et al. (2015), where the content of water soluble carbohydrates is maximal and the ash fraction of the total solids is minimal (Manns et al. 2017). In this study, the biomass was harvested in early summer (June), where the content of the water soluble fermentable carbohydrate mannitol is high but not yet expectedly maximal, as it typically peaks in late June (Schiener et al. 2015; Manns et al. 2017). Early June however represents a realistic time of harvest of S. latissima for food purposes in North Atlantic waters since biofouling of the seaweed fronds increasingly sets in from June and onwards, with a consequent loss of biomass quality (Handå et al. 2013; Bruhn et al. 2016). The concentrations of mannitol in the sugar kelp in this study were generally a factor of 10 lower than observed for this species in general (5–22% of DM) (Adams et al. 2011; Holdt and Kraan 2011; Nielsen et al. 2016; Manns et al. 2017). The relatively low mannitol concentrations observed were likely due to the analysis method applied in this study, as neither acid nor enzymatic hydrolysis pretreatments were involved in the extraction process (Manns et al. 2014). Despite the low mannitol concentration, the fermentation process advanced successfully, presumably on the basis of the dextrose used as carrier in the LAB starter culture, the concentration of which was up to 18 times as high as the natural concentration of mannitol from the seaweed. Regardless of the low absolute level of mannitol detected, the almost total absence of mannitol in the fermented biomass, indicated that either the mannitol was drained to the liquid fraction of the fermentation (Herrmann et al. 2015) or consumed by the LAB in the fermentation process, as previously described (Black 1955; McDonald et al. 1991; Hwang et al. 2011). The content of WSC (dextrose + mannitol) in this study was in the same range as in Herrmann et al. (2015): 256 g kg−1 TS, as compared to 217 g kg−1 TS.
A TS content of 30% of FW is considered optimal for ensiling of biomass (McDonald et al. 1991). Notably, the TS content of the S. latissima biomass in this study was very low (up to 6% of FW) compared to S. latissima biomass described in other studies, where the content of DM is in the rage of 11.3–22.5% of FW (Marinho et al. 2015; Schiener et al. 2015). One negative effect of a high moisture content is an increased drainage of effluent from the biomass during the fermentation process, with an implicit loss of minerals and biomolecules such as small organic acids, alcohols and mannitol to the effluent (Herrmann et al. 2015). The heat-treatment prior to the fermentation reduced the DM contents of the biomass. During the fermentation process, however, the DM content of the sugar kelp biomass increased to reach the initial concentrations of the fresh sugar kelp. This may be explained by a dewatering of the biomass caused by the fermentation, leading to an increase of the DM content (Herrmann et al. 2015), and by the growing population of LAB bacteria assimilating dissolved compounds and by their cell constituents increasing the DM content of the total biomass (Black 1955).
Nutritional value and food safety
The N concentrations observed in the S. latissima biomass were in agreement with values reported in the literature for this species at this time of the year (Nielsen et al. 2014; Bruhn et al. 2016; Mols-Mortensen et al. 2017), and the estimated protein content of the sugar kelp product did not significantly change in the processes of heat-treatment and fermentation. A specific analysis of the amino acid composition of the biomass would be needed in order to fully evaluate the effects of the heat-treatment and fermentation processes on the nutritional value of the final product (Mols-Mortensen et al. 2017).
During the processing, the content of Na was reduced by 15%. Human intake of Na increases the risk of cardiovascular diseases, and hence the reduction, albeit limited, in Na content with the fermentation is positive (Ortega-Calvo et al. 1993). The molar Na:K ratio of the seaweed products was not altered by the processing, but remained below one.
Regarding pathogens, no presence of B. cereus was detected in the fermented seaweed. In this study, only one specific analysis was carried out in order to detect the presence of one specific pathogenic bacterium (B. cereus) in the seaweed material. Thus, it is beyond the scope of this paper to fully evaluate the microbial food safety aspects and shelf life of this particular product. However, the relatively rapid pH decrease to below pH 5 within 40 h indicated a fast preservation of the seaweed material (Caplice and Fitzgerald 1999). Also, the starter culture of L. plantarum applied in this study is a highly competitive culture, even able to produce bacteriocins specifically limiting the growth of Listeria, and marketed for this ability (Tolvanen et al. 2008).
Regarding harmful metals, fermentation improved the food safety of the sugar kelp in reducing the content of two out of four specific harmful trace metals (Cd and Hg) of which the EU and European Food Safety Authority have set limit values in food for human consumption and feeds for animals: arsenic (As), cadmium (Cd), lead (Pb) and mercury (Hg). Through fermentation, the Cd concentrations of the sugar kelp was brought below the limit values of 3 μg g−1 DM set for dietary supplements based on algae (EU 2013). Regarding Hg, the concentrations in any treatment of the sugar kelp were not problematic with respect to food and feed legislation, as they were a factor of 4–7 below limit values regarding food, and a factor of 22–33 below limit values for feed (EU 2013).
While Hg adsorption to seaweed biomass is related to carboxylate groups in the biomass, chelation is the primary mechanism behind Cd sequestering in algae (Roleda et al. 2019). Albeit the mechanisms behind the metal accumulating ability of seaweed biomass differ between Cd and Hg, studies made on another brown alga, Sargassum muticum, demonstrate that the binding capacity was reduced at pH below 4.5 (Lodeiro et al. 2004; Carro et al. 2011). Further studies are needed to document the mechanisms behind the reduction in Cd and Hg during the fermentation process.
The fermentation and heat-treatment did not alter the concentrations of the other two harmful metals, Pb and As. Regarding Pb, the concentrations found in the sugar kelp products were a factor of 3–5 lower than the European food maximum limit (ML), and a further factor 5–10 below the ML for feed (EU 2013), and thus not problematic.
Regarding As, the legislation at national levels may differ between limit values set for total As and/or for the inorganic species of As. In Europe in general, no limit values are set for total As in foods, whereas for inorganic As, the ML is set at 3.0 μg g−1 DM in France and the USA, since only this species of arsenic is considered toxic to the consumer (EFSA Panel on Contaminants in the Food Chain (CONTAM) 2009; Holdt and Kraan 2011). An interesting subject for future studies would be to investigate the specific effect of fermentation on the content of inorganic As.
The tissue concentrations of the four harmful metals were all generally within range of concentrations reported for the same species (Bruhn et al. 2016; Nielsen et al. 2016; Roleda et al. 2019), emphasising that concentrations of As, and to some extent Cd, of kelps may pose a barrier for products entering the European and global food and feed markets.
Refining sensory properties
The majority of the flavour and odour descriptors generated through the taste tests were similar to the key descriptors selected by others to describe the taste and smell of seaweeds, including sugar kelp: salty, sea/marine, sweet, sour, umami, bitter and with a slimy appearance (Mouritsen et al. 2012; Chapman et al. 2015; López-Pérez et al. 2017). Sugar kelp has been described as having a strong salty taste with components of sea and grass, but a weak umami component as compared to other kelp species, L. digitata (oarweed) and Alaria esculenta (winged kelp), as well as the red algae Palmaria palmata (dulse) (Chapman et al. 2015). The dominant descriptors of the smell of sugar kelp were seaweed, marine, hay and spices (López-Pérez et al. 2017). The relatively mild umami taste component of sugar kelp was described by Mouritsen et al. (2012), who also mentioned the strong sea smell of the sugar kelp. The viscosity or slimy appearance of sugar kelp was similarly described by Mouritsen et al. (2012) and Chapman et al. (2015). The loss of saltiness in the heat-treatment and fermentation process may be explained by the simultaneous loss of minerals responsible for a salty taste – predominantly Na but also Mg, the concentrations of which decreased by 15% and 21%, respectively, in the heat-treatment and fermentation process. The decrease in the umami taste component in the heat-treated seaweed went in parallel with a decrease in the N content of the product. Glutamic acid is the amino acid responsible for the umami taste, as the essence of umami taste, the taste enhancer monosodium glutamate (MSG), is the sodium salt of glutamic acid (Fuke and Shimizu 1993; Ikeda 2002). When cooking meat, a reduction of approximately 50% in the content of glutamic acid was observed during the first 15 min of cooking time, with a corresponding increase of glutamic acid in the cooking broth (Sasaki et al. 2007), reflecting how umami taste was extracted from meat to the soup during cooking. In this study, the seaweed was heat-treated at 95 °C for 15 min and a similar extraction of umami taste was expected to have taken place from the seaweed biomass to the boiling water. During the fermentation process, the seaweed biomass stayed in the broth; however, the loss of umami taste did not recover during fermentation. A more thorough analysis identifying changes in the amino acid and nucleotide composition of the seaweed biomass through the different treatments would be needed for quantifying the specific effect of the fermentation process on key amino acids such as glutamic acid, but also aspartic acid, and the nucleotide inosinate, that both also contribute to the umami taste (Mouritsen et al. 2012; Uchida et al. 2017).
The composition of volatiles contributing to the distinct smell of kelp is complex (Seo et al. 2012; López-Pérez et al., 2017). Dimethylsulphide (DMS) is one of the key compounds responsible for the sea smell (López-Pérez et al., 2017). Fermentation of kelp extract using Aspergillus oryzae was shown to reduce the expected odour intensity of off odours of the seaweed extract by a factor of four over four days, including a full elimination of the concentration of DMS after 2 days (Seo et al. 2012). The ability of LAB fermentation to eliminate smell deriving from DMS and other sulphides in fermented foods has also been demonstrated (Zhao et al. 2016).
The less slimy appearance of the heat-treated product may be explained by dilution or removal by washing/boiling of exuded water soluble polysaccharides, such as laminarin or fucoidan (Percival 1979). Laminarin is a storage carbohydrate, the content of which peaks in late summer, and in June attains concentrations around 5–20% of DM depending on environmental conditions and age of seaweed fronds (Black 1950; Schiener et al. 2015; Manns et al. 2017). Laminarin may be utilised in an anaerobic LAB fermentation process (Black 1955).
Market perspectives
Fermentation of kelps using LAB may have an impact on the economic feasibility of kelp products in several links of the process chain. Regarding storage stabilisation of seaweed biomass, drying is an efficient method of stabilisation; however, in temperate climates where sun drying is not an option, drying demands large amounts of energy, hereby lowering the environmental as well as economical sustainability of the process chain (Stévant et al. 2017). Further, sun drying and drying at lower temperatures negatively affect the nutritional value of the seaweed biomass, reducing the content of vitamin C, amino acids and polyunsaturated fatty acids, as well as antioxidants (Chan et al. 1997; Gupta et al. 2011b). Thus, LAB fermentation could offer a more gentle and sustainable method of storage stabilising seaweed biomass for non-dry food products, but at the same time introduce a challenge to the food industry that is currently based on dried products. Regarding food safety and improving shelf life of non-dried products, this study has on the microbiological side only documented absence of B. cereus in the fermented products, and the microbiological aspects needs further investigations. However, the finding that LAB fermentation significantly reduced the concentrations of two harmful metals (Cd and Hg) is new and important for future food productions and biorefinery concepts, since content of harmful metals is one of the challenges that seaweed products face in the food and feed industries (Holdt and Kraan 2011). Also the contents of Na, the reputation of the slimy looks and strong sea smell are some of the challenges that seaweed products are facing among consumers. Thus, the findings of this study that the LAB fermented products had an unchanged protein content, but were less slimy, less smelly, and contained a healthier Na-reduced salt, may help improving the consumer perception of kelp products. Overall, the positive modifications in sensory properties and composition could open for innovation towards new kelp food products based on fermentation, providing also a source of non-dairy probiotic food of which the demand is increasing.
Conclusion
Fermentation with Lactobacillus plantarum changed fresh sugar kelp harvested in June into a product with a milder, less salty taste, a reduced sea smell and a less slimy visual appearance. The fermentation did not alter the protein content of the seaweed biomass, but improved the mineral composition of the product, in reducing the concentrations of two harmful metals (Cd and Hg), as well as the Na content. The fermentation reduced pH to 4.5 within 40 h, with LAB counts increasing by a factor of 100 and with no Bacillus cereus present.
Fermentation as a sustainable method for stabilising of a wet biomass could lower the total cost price and drive innovation towards new food and feed products based on fermentation.
References
Adams JMM, Ross AB, Anastasakis K, Hodgson EM, Gallagher JA, Jones JM, Donnison IS (2011) Seasonal variation in the chemical composition of the bioenergy feedstock Laminaria digitata for thermochemical conversion. Bioresour Technol 102:226–234
Angell AR, Mata L, de Nys R, Paul NA (2016) The protein content of seaweeds: a universal nitrogen-to-protein conversion factor of five. J Appl Phycol 28:511–524
Black WAP (1950) The seasonal variation in weight and chemical composition of the common British Laminariaceae. J Mar Biol Assoc UK 29:45–72
Black WAP (1955) The preservation of seaweed by ensiling and bactericides. J Sci Food Agric 6:14–23
Bruhn A, Tørring DB, Thomsen M, Canal-Vergés P, Nielsen MM, Rasmussen MB, Eybye KL, Larsen MM, Balsby TJS, Petersen JK (2016) Impact of environmental conditions on biomass yield, quality, and bio-mitigation capacity of Saccharina latissima. Aquac Environ Interact 8:619–363
Caplice E, Fitzgerald GF (1999) Food fermentations: role of microorganisms in food production and preservation. Int J Food Microbiol 50:131–149
Carro L, Barriada JL, Herrero R, Sastre de Vicente ME (2011) Adsorptive behaviour of mercury on algal biomass: competition with divalent cations and organic compounds. J Hazard Mater 192:284–291
Chan JCC, Cheung PCK, Ang PO (1997) Comparative studies on the effect of three drying methods on the nutritional composition of seaweed Sargassum hemiphyllum (Turn) C Ag. J Agric Food Chem 45:3056–3059
Chapman AS, Stévant P, Larssen Wenche E (2015) Food or fad? Challenges and opportunities for including seaweeds in a Nordic diet. Bot Mar 58:423–433
Coquery M, Carvalho FP, Azemard S, Bachelez M, Horvat M (2000) Certification of trace and major elements and methylmercury concentrations in a macroalgae (Fucus sp.) reference material, IAEA-140. Fresenius J Anal Chem 366:792–801
EFSA Panel on Contaminants in the Food Chain (CONTAM) (2009) Scientific opinion on arsenic in food. EFSA J 7(10):1351
Enriquez S, Duarte CM, Sandjensen K (1993) Patterns in decomposition rates among photosynthetic organisms - the importance of detritus C-N-P content. Oecologia 94:457–471
EU (2013) Commission regulation (EU) No 1275/2013 of 6 December 2013 - amending annex I to directive 2002/32/EC of the European Parliament and of the council as regards maximum levels for arsenic, cadmium, lead, nitrites, volatile mustard oil and harmful botanical impurities. The European Commission
FAO (2016) The state of world fisheries and aquaculture 2016. Contributing to food security and nutrition for all. FAO, Rome, p 200
FAO (2018) The State of World Fisheries and Aquaculture 2018 - Meeting the sustainable development goals. Rome
Felix S, Pradeepa P (2011) Seaweed (Ulva reticulata) based fermented marine silage feed preparation under controlled conditions for Penaeus monodon larval development. Mar Sci 1:1–3
Fernand F, Israel A, Skjermo J, Wichard T, Timmermans KR, Golberg A (2017) Offshore macroalgae biomass for bioenergy production: environmental aspects, technological achievements and challenges. Renew Sust Energ Rev 75:35–45
Fuke S, Shimizu T (1993) Sensory and preference aspects of umami. Trends Food Sci Technol 4:246–251
Gupta S, Abu-Ghannam N (2012) Probiotic fermentation of plant based products: possibilities and opportunities. Crit Rev Food Sci Nutr 52:183–199
Gupta S, Abu-Ghannam N, Rajauria G (2012) Effect of heating and probiotic fermentation on the phytochemical content and antioxidant potential of edible Irish brown seaweeds. Bot Mar 55:527–537
Gupta S, Abu-Ghannam N, Scannell AGM (2011a) Growth and kinetics of Lactobacillus plantarum in the fermentation of edible Irish brown seaweeds. Food Bioprod Process 89:346–355
Gupta S, Cox S, Abu-Ghannam N (2011b) Effect of different drying temperatures on the moisture and phytochemical constituents of edible Irish brown seaweed. LWT-Food Sci Technol 44:1266–1272
Herrmann C, FitzGerald J, O’Shea R, Xia A, O’Kiely P, Murphy JD (2015) Ensiling of seaweed for a seaweed biofuel industry. Bioresour Technol 196:301–313
Holdt SL, Kraan S (2011) Bioactive compounds in seaweed: functional food applications and legislation. J Appl Phycol 23:543–597
Hwang HJ, Lee SY, Kim SM, Lee SB (2011) Fermentation of seaweed sugars by Lactobacillus species and the potential of seaweed as a biomass feedstock. Biotechnol Bioprocess Eng 16:1231–1239
Handå A, Forbord S, Wang XX, Broch OJ, Dahle SW, Storseth TR, Reitan KI, Olsen Y, Skjermo J (2013) Seasonal- and depth-dependent growth of cultivated kelp (Saccharina latissima) in close proximity to salmon (Salmo salar) aquaculture in Norway. Aquaculture 414:191–201
Ikeda K (2002) New seasonings. Chem Senses 27:847–849
ISO 15214 (1998) Microbiology of food and animal feeding stuffs. Horizontal method for the enumeration of mesophilic lactic acid bacteria. Colony-count technique at 30 degrees C. Technical Committee ISO/TC 34, Agricultural food products, Subcommittee SC 9, Microbiology
ISO (2005) ISO 6658. Sensory analysis. Methodology. General guidance. The International Organisation for Standardization Technical Committee ISO/TC 34, Food products, Subcommittee SC 12, Sensory analysis:1–20
ISO (2016) ISO 13299:2016. Sensory analysis. Methodology. General guidance for establishing a sensory profile. The International Organisation for Standardization ISO/TC 34, Food products, Subcommittee SC 12, Sensory analysis.:1–41
Karovicova J, Kohajdova Z (2005) Lactic acid-fermented vegetable juices - palatable and wholesome foods. Chem Pap 59:143–148
Kerrison PD, Stanley MS, Black KD, Hughes AD (2017) Assessing the suitability of twelve polymer substrates for the cultivation of macroalgae Laminaria digitata and Saccharina latissima (Laminariales). Algal Res 22:127–134
Kerrison PD, Stanley MS, Edwards MD, Black KD, Hughes AD (2015) The cultivation of European kelp for bioenergy: site and species selection. Biomass Bioenergy 80:229–242
Kim JK, Yarish C, Hwang EK, Park M, Kim Y (2017) Seaweed aquaculture: cultivation technologies, challenges and its ecosystem services. Algae 32:1–13
Lodeiro P, Cordero B, Grille Z, Herrero R, Sastre de Vicente ME (2004) Physicochemical studies of cadmium(II) biosorption by the invasive alga in Europe, Sargassum muticum. Biotechnol Bioeng 88:237–247
López-Pérez O, Picon A, Nuñez M (2017) Volatile compounds and odour characteristics of seven species of dehydrated edible seaweeds. Food Res Int 99:1002–1010
Manns D, Deutschle AL, Saake B, Meyer AS (2014) Methodology for quantitative determination of the carbohydrate composition of brown seaweeds (Laminariaceae). RSC Adv 4:25736–25746
Manns D, Nielsen MM, Bruhn A, Saake B, Meyer AS (2017) Compositional variations of brown seaweeds Laminaria digitata and Saccharina latissima in Danish waters. J Appl Phycol 29:1493–1506
Marinho G, Holdt S, Birkeland M, Angelidaki I (2015) Commercial cultivation and bioremediation potential of sugar kelp, Saccharina latissima, in Danish waters. J Appl Phycol 27:1963–1973
Marco A, Rubio R, Compano R, Casals I (2002) Comparison of the Kjeldahl method and a combustion method for total nitrogen determination in animal feed. Talanta 57:1019–1026
McDonald P, Henderson AR, Heron SJE (1991) The Biochemistry of Silage (2nd edn), Chalcombe Publications, Kingston
Milledge JJ, Harvey PJ (2016a) Ensilage and anaerobic digestion of Sargassum muticum. J Appl Phycol 28:3021–3030
Milledge JJ, Harvey PJ (2016b) Potential process “hurdles” in the use of macroalgae as feedstock for biofuel production in the British Isles. J Chem Technol Biotechnol 91:2221–2234
Milledge JJ, Nielsen BV, Bailey D (2016) High-value products from macroalgae: the potential uses of the invasive brown seaweed, Sargassum muticum. Rev Environ Sci Biotechnol 15:67–88
Mols-Mortensen A, Ortind EG, Jacobsen C, Holdt SL (2017) Variation in growth, yield and protein concentration in Saccharina latissima (Laminariales, Phaeophyceae) cultivated with different wave and current exposures in the Faroe Islands. J Appl Phycol 29:2277–2286
Mouritsen OG, Williams L, Bjerregaard R, Duelund L (2012) Seaweeds for umami flavour in the New Nordic Cuisine. Flavour 1:1–12
Nielsen M, Krause-Jensen D, Olesen B, Thinggaard R, Christensen P, Bruhn A (2014) Growth dynamics of Saccharina latissima (Laminariales, Phaeophyceae) in Aarhus Bay, Denmark, and along the species’ distribution range. Mar Biol 161:2011–2022
Nielsen MM, Bruhn A, Rasmussen MB, Olesen B, Larsen MM, Møller HB (2012) Cultivation of Ulva lactuca with manure for simultaneous bioremediation and biomass production. J Appl Phycol 24:449–458
Nielsen MM, Manns D, D'Este M, Krause-Jensen D, Rasmussen MB, Larsen MM, Alvarado-Morales M, Angelidaki I, Bruhn A (2016) Variation in biochemical composition of Saccharina latissima and Laminaria digitata along an estuarine salinity gradient in inner Danish waters. Algal Res 13:235–245
Ortega-Calvo JJ, Mazuelos C, Hermosin B, Saizjimenez C (1993) Chemical composition of Spirulina and eukaryotic algae food products marketed in Spain. J Appl Phycol 5:425–435
Pahlow G, Muck RE, Driehuis F, Oude Elferink SJWH, Spoelstra SF (2003) Microbiology of ensiling. In: Buxton D, Muck, RE, Harrison, JH, (eds) Silage science and technology, vol 42. American Society of Agronomy, pp 31–93
Percival E (1979) Polysaccharides of green, red and brown seaweeds - their basic structure, biosynthesis and function. Brit Phycol J 14:103–117
Redden H, Milledge JJ, Greenwell HC, Dyer PW, Harvey PJ (2017) Changes in higher heating value and ash content of seaweed during ensiling. J Appl Phycol 29:1037–1046
Roleda MY, Marfaing H, Desnica N, Jónsdóttir R, Skjermo J, Rebours C, Nitschke U (2019) Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass: health risk assessment and implication for food applications. Food Control 95:121–134
Sasaki K, Motoyama M, Mitsumoto M (2007) Changes in the amounts of water-soluble umami-related substances in porcine longissimus and biceps femoris muscles during moist heat cooking. Meat Sci 77:167–172
Schiener P, Black K, Stanley M, Green D (2015) The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J Appl Phycol 27:363–373
Seo YS, Bae HN, Eom SH, Lim KS, Yun IH, Chung YH, Jeon JM, Kim HW, Lee MS, Lee YB, Kim YM (2012) Removal of off-flavors from sea tangle (Laminaria japonica) extract by fermentation with Aspergillus oryzae. Bioresour Technol 121:475–479
Stévant P, Marfaing H, Rustad T, Sandbakken I, Fleurence J, Chapman A (2017) Nutritional value of the kelps Alaria esculenta and Saccharina latissima and effects of short-term storage on biomass quality. J Appl Phycol 29:2417–2426
Tolvanen R, Hellstrom S, Elsser D, Morgenstern H, Bjorkroth J, Korkeala H (2008) Survival of Listeria monocytogenes strains in a dry sausage model. J Food Protect 71:1550–1555
Tsuchiya K, Matsuda S, Hirakawa G, Shimada O, Horio R, Taniguchi C, Fujii T, Ishida A, Iwahara M (2007) GABA production from discolored laver by lactic acid fermentation and physiological function of fermented laver. Food Preserv Sci 33:121–125
Uchida M, Kurushima H, Ishihara K, Murata Y, Touhata K, Ishida N, Niwa K, Araki T (2017) Characterization of fermented seaweed sauce prepared from nori (Pyropia yezoensis). J Biosci Bioeng 123:327–332
Uchida M, Miyoshi T, Yoshida G, Niwa K, Mori M, Wakabayashi H (2014) Isolation and characterization of halophilic lactic acid bacteria acting as a starter culture for sauce fermentation of the red alga Nori (Porphyra yezoensis). J Appl Microbiol 116:1506–1520
Uchida M, Murata M (2002) Fermentative preparation of single cell detritus from seaweed, Undaria pinnatifida, suitable as a replacement hatchery diet for unicellular algae. Aquaculture 207:345–357
Uchida M, Numaguchi K, Murata M (2004) Mass preparation of marine silage from Undaria pinnatifida and its dietary effect for young pearl oysters. Fish Sci 70:456–462
Weissbach F (1996) New developments in crop conservation. In: Jones, DIH, Dewhurst, R, Merry, R, Haigh, PM (Eds) 11th international silage conference Aberystwyth, Wales PP 11-25
Zhao N, Zhang CC, Yang Q, Yang B, Lu WW, Li DY, Liu XM, Tian FW, Zhang H, Chen W (2016) Multiple roles of lactic acid bacteria microflora in the formation of marker flavour compounds in traditional Chinese paocai. RSC Adv 6:89671–89678
Acknowledgements
This work was carried out as part of the research and innovation project Fermatang supported by funding allocated to EAMV (FFL-13 funds for research association and development and evidence foundation (the Frascati-manual)), as well as the MacroAlgae Biorefinery 4 (MAB4) and the Macrofuels projects, funded by the Innovation Fund Denmark and the European Union’s Horizon 2020 research and innovation programme under grant agreement no. 654010, respectively. The authors thank Anne Marie Plejdrup, Kitte Linding Gerlich and Kristina Høeg for skilful lab work, Tinna Majlund Christensen for assistance with graphics, the sensory panel for hours of professional seaweed tasting, the food technology students for innovation of new foods based on the fermented sugar kelp as well as numerous curious citizens for tastings and comments during public seaweed taste events.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Bruhn, A., Brynning, G., Johansen, A. et al. Fermentation of sugar kelp (Saccharina latissima)—effects on sensory properties, and content of minerals and metals. J Appl Phycol 31, 3175–3187 (2019). https://doi.org/10.1007/s10811-019-01827-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-019-01827-4