
Abstract
The Canary Islands were initially colonized around 200 CE by North African Berber populations who brought with them domestic plants and animals. These communities remained isolated until the arrival of Europeans in the Late Middle Ages which triggered the conquest of the archipelago. Its geostrategic location in the framework of Atlantic expansion facilitated the arrival of people, knowledge, goods, plants, and animals which served to shape the new Canarian society. This study explores the dietary practices of this period through the zooarchaeological analysis of the faunal remains recovered at the Hospital de San Martín (fifteenth to the eighteenth centuries), Las Palmas (Gran Canaria, Spain). Our results suggest that caprines were the main taxa consumed. This follows both the pre-colonial Indigenous tradition and, according to historical sources, the subsequent European period. The faunal assemblage recovered also reveals the first directly radiocarbon-dated cases in the Canary Islands of three hitherto unknown species: cattle, chicken, and rabbit. Moreover, the results evidence a progressive increase throughout the Modern Era of chicken and rabbit, and that translocating these new species to the archipelago during the European expansion led to a diversification of meat resources.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The European colonization of the Canary Islands represents one of the first phases of the expansion into the Atlantic Ocean in the Modern Era (Aznar Vallejo 2008). This colonization entailed major ramifications in all areas of society as it led to the development of international mercantilism and transatlantic trade, movements of vast populations from Europe and Africa to the Caribbean and America, and the emergence of new colonies and territories.
Unlike other Atlantic archipelagos such as the Azores, Madeira or Cape Verde, the Canarian archipelago was occupied prior to the arrival of Europeans in the fourteenth century (Aznar Vallejo et al. 2006). Berber farming populations from North Africa settled sometime between the first and third centuries CE (Velasco-Vázquez et al. 2021). The European and Indigenous inhabitants from the outset established relations that coincided with an evangelizing process and slaving raids that decimated the original inhabitants (Aznar Vallejo 2008). The archipelago was eventually conquered in 1496 when it became a province of the Spanish Kingdom (Aznar Vallejo 2008; Tejera Gaspar and Aznar Vallejo 1991). The islands emerged as a key location for the European Atlantic expansion that led to the control of trade routes toward both the African and American continents (Aznar Vallejo et al. 2000; Onrubia Pintado and González Marrero 2016). During the Modern Era, the Canary Islands became inhabited by Europeans from various geographical backgrounds, including Castilians, Portuguese (Fajardo Spínola 2004), Genovese (Bruquetas de Castro 2015), and Flemish (Viña Brito 2012). Additionally, North African and Sub-Saharan Western African individuals were brought to work in the sugar cane industry of the archipelago (Lobo Cabrera 2016; Santana-Pérez 2018; Viña Brito 2006). The integration of the Indigenous into the new society is also supported by genetic and historical records (Arnay-de-la-Rosa et al. 2015; Lobo Cabrera 2016; Santana et al. 2016). The Canarian society resulting from this process thus consisted of Indigenous, European, North African, and Sub-Saharan populations, each bearing their own unique cultural backgrounds (Fregel et al. 2021; Santana et al. 2016; Velasco-Vázquez et al. 2003).
The Berber populations that initially colonized the islands brought with them domestic plants and animals allowing them to perpetuate their traditional North African strategies of subsistence. It was then that a variety of crops were introduced: barley (Hordeum vulgare), wheat (Triticum), lentil (Lens culinaris), field bean (Vicia faba), peas (Pisum sativum), and fig (Ficus carica). Barley became in fact the main crop followed by wheat, pulses, and in the case of the island of Gran Canaria, on which this paper is focused, fig (Morales et al. 2017). Evidence from Gran Canaria reveals that agriculture was the island’s main source of food (Morales et al. 2023). Yet the North African settlers also translocated domestic animals such as goats (Capra hircus), hairless sheep (Ovis aries), pigs (Sus domesticus), and dogs (Canis familiaris), which likewise played a substantial role in their subsistence (Castellano Alonso et al. 2016). Evidence also suggest that they resorted to the wild resources offered by the islands which included marine species (fish and mollusks) and, to a lesser extent, native terrestrial flora and fauna (Alberto-Barroso 1998; Henríquez-Valido et al. 2020; Mesa Hernández 2016; Morales et al. 2021; Rodríguez-Rodríguez et al. 2021; Rodríguez Santana 1994).
Europeans, mainly Castilians, began to settle permanently in Gran Canaria from the end of the fifteenth century, especially after the end of the conquest in 1483 (Lobo Cabrera 2012). They raised settlements in new areas as well as at the locations of traditional Indigenous rural sites (Onrubia Pintado and González Marrero 2016). This colonization led to the introduction of new plant and animal species from Europe, Africa, Asia, and America, a process bolstered by the archipelago’s geostrategic position along the Atlantic trade network (Armenteros Martínez 2018; Hernández Suárez 2021; Santana Pérez et al. 2004). The Europeans also introduced new farming technologies, food processing techniques, dietary patterns and habits such as the consumption of tobacco (Arnay-de-la-Rosa 2009; Díaz-Sierra 2022). Territorial colonization was put in place by means of a system of land and water distribution appropriated by the Crown according to the right of conquest as well as by granting economic privileges. The land was destined for cash crops (sugar cane and woad) and subsistence agriculture (cereals, vineyards, fruit trees, vegetables) (Díaz-Sierra 2022; Ronquillo Rubio and Aznar Vallejo 1998).
All these developments took place along with the introduction of a new economic model based on the sugar cane industry (Rivero Suárez 1991; Ronquillo Rubio 2008; Viña Brito 2014). The new plantations and mills at times took advantage of lands originally labored for subsistence agriculture by the Indigenous Gran Canarians (Díaz-Sierra 2022). The Europeans likewise resorted to the experience of the Indigenous populations to manage livestock during the initial moments of the colonization (Betancor Quintana 2003). Genetic data point to an isolation of native pigs and goats breeds that most likely endured until the introduction of industrial varieties (Ferrando et al. 2015; Olalde et al. 2015). That was not the case of dogs which appear to have bred with the new breeds introduced after colonization (Suárez et al. 2013). However, the system of ownership and management of farm animals brought by the Europeans differed greatly from that of the Native populations (Hernández Marrero 2001; Lobo Cabrera 2019). Written sources from the time state that livestock management prioritized exploiting dairy products and animal skins. This production not only served the needs of the islanders but also to export goods (cheeses and hides) to the Iberian Peninsula and continental America (Betancor Quintana 2003; Lobo Cabrera 1998; Quintana Andrés 2003).
European colonization led to the introduction of new species into Gran Canaria, too. Old documentary sources mention cattle, horses, donkeys, chickens, and rabbits (Cadamosto 2017 [1456]; Tejera Gaspar 2020). All of them were fundamental to both the subsistence of Canarian society as well as to transform the territory, maintain agricultural activity, and mobility. This also meant the introduction of innovative agricultural practices, including the utilization of animal-driven plows and the adoption of advanced food processing methods such as watermills, windmills, and animal mills (Cullen del Castillo 1947; Ruíz Melgarejo 1974 [1531]). However, historical sources are not clear as to the repercussion or the proportion of these new species with respect to goats, sheep, and pigs present since the Indigenous Period (Capote et al. 2002; Tejera Gaspar 2020; Torres Krupij et al. 2018). The Canary Islands were also key in translocating animals across the Atlantic. Both historical records and genetic analyses reveal that Canarian goats and pigs contributed to the implantation of these species in the Caribbean islands and America (Capote et al. 2002; Delgado et al. 2000; Tejera Gaspar 2020; Torres Krupij et al. 2018). Moreover, it is this framework that saw the only domestication of a native Canarian species: the canary bird (Serinus canaria) (Arnaiz-Villena et al. 1999; Birkhead et al. 2004).
To date archaeological investigations in the archipelago have mainly focused on the earlier Indigenous societies, neglecting the subsequent periods (Arnay-de-la-Rosa 2009; González Marrero and Tejera Gaspar 2011; Mitchell 2023). Despite the key role of these islands in the development of animal translocation between continents (Delsol et al. 2022; López et al. 2013; McTavish et al. 2013; Olalde et al. 2015; Reitz 1992; Speller et al. 2013), local zooarchaeological research is basically limited to either Indigenous sites or sites from the fifteenth century corresponding to the period of contact between the Europeans and the Native populations (Castellano Alonso et al. 2016, 2018). Additionally, while historical written sources of the Modern Era of the Canary Islands provide valuable insights into human-animal interactions, they are often biased, limited in scope, or subject to interpretation. Zooarchaeology analysis offers a complementary perspective by focusing on the actual material evidence left behind. The integration of these two approaches enables a more robust interpretation of the relationship between past peoples and the animals with which they shared the world (O’Connor 2003). The current study is thus the first to directly make use of faunal data from Modern Era in the Canary Islands.
This study explores the dietary practices between the late fifteenth and the late eighteenth centuries at the old Hospital de San Martín (Gran Canaria) (HSM henceforth) by means of zooarchaeological analyses. Apart from the finds themselves, this work resorts to data garnered from historical written sources of the time and by comparison with the results of investigations of other faunal assemblages. The goal is to attempt to grasp, on the one hand, the processes of change linked to the European colonization with respect to the Indigenous Period and, on the other hand, to determine to what extent the patterns of consumption at the HSM resembled those of other contemporary Modern Era contexts in the Atlantic world. The main hypothesis is that caprines (sheep and goats) served as the nutritional base of the Canarians during this period probably due to their availability since the Indigenous Period and their adaptative capacity to the island environment.
Site Description
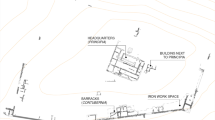
The archaeological site of HSM is located in the historic center of Las Palmas de Gran Canaria (Fig. 1C), a port city along the northeastern coast of the island founded in 1478 during the conquest by the Crown of Castile. Las Palmas became the nucleus of the civil, military, and ecclesiastical power in the archipelago during the first century of Castilian rule (Aznar Vallejo 1983). The HSM was founded in 1481 to care for the sick, pobres de solemnidad (the very poor), and orphans (Quintana Andrés 2003; Santana-Pérez 2005). It maintained its activities until 1780 when it was razed and buried during work related to the expansion of the Cathedral of Santa Ana and the construction of the San Marcial School (Bosch Millares 1940). Due to a new construction project in 2005, the Historical Heritage Service of the Cabildo de Gran Canaria requested an archaeological intervention. This, according to the historical cartography available at the time, affected the northeastern end of the old hospital. The property was bordered by the Cilla Episcopal, the Plaza de Los Álamos and buildings related to various types of craftworks (Tous Meliá 1995). Preservation of the archaeological remains of the old hospital thus led to a rescue operation carried out by the private company Tibicena, S.L. (Mendoza Medina et al. 2012).

A Map of the Canary Islands and (B-C) detail of Gran Canaria indicating the location of the Hospital de San Martín
The archaeological work identified several architectural features and an interior patio belonging to different construction phases of the HSM and later buildings. Ultimately three archaeological surveys focused on the areas threatened by the foundations of the new building (Fig. 2). These recintos or enclosed features (henceforth labelled R1, R2, and R3) were bordered by surface construction structures that were maintained to preserve the largest possible number of archaeological elements. The soundings sunk between the structures revealed stratigraphic sequences consisting of occupation layers and fills containing archaeological remains in secondary position.

A Section of the deeper sounding in R3. B Aerial view of the archaeological intervention of 2010 at the Hospital de San Martín with the position of the surveys at the Recintos (R1, R2 and R3)
The upper sedimentary layer was sealed by a level associated with the later buildings, which capped the most recent construction phase of the hospital. It is only represented in R2 and R3 soundings. R2 revealed several stratigraphic units such as SUs 43 and 81 rich in ash and coal containing numerous elements of fauna (Mendoza Medina et al. 2012) which appear to be rubbish dumps of the site’s most recent phase linked to consumption. R3, in turn, revealed a sequence 2.50 m thick starting with the natural substrate capped by the oldest phase of construction of the hospital. This was followed by a series of fills stemming from different architectural remodeling capped by the hospital’s most recent construction phase (see Fig. 2). The sequence also revealed three floor layers evidencing three different occupations. One is associated with the site’s most recent phase (SU37), while the other two (SU74 and SU83) formed part of the hospital’s initial construction which contained many artifacts, notably local and imported ware from the late fifteenth to sixteenth centuries.
R1 is characterized by a foundation (SU20), a stone paving (SU12), and a water well (now dry) forming part of the inner patio of the hospital dated to the most recent period (Mendoza Medina et al. 2012). The northern end of this space yielded a sequence 2.60 m thick attaining the natural substrate. It comprised successive fills of remodeling from the oldest phase. It is noticeable that this second phase also yielded numerous faunal remains in units SU52 deriving from consumption as well as imported ware from the sixteenth to seventeenth centuries. Below there were other layers such as SU88, a fill containing many artifacts from the late fifteenth to sixteenth centuries.
Structural evidence of the two construction phases and the relative chronology gleaned from the pottery’s stratigraphic contexts point to two main phases (Mendoza Medina et al. 2012). The earliest levels are dated by imported pottery from middle fifteenth to sixteenth centuries, whereas the latest are associated with a higher proportion seventeenth-eighteenth century imported ware (Sosa Suárez 2012).
Methods
Radiocarbon Dating and Bayesian Modeling
To corroborate the archaeological sequence of the site, six bone samples were radiocarbon dated by Accelerator Mass Spectrometry (AMS) at the SUERC Laboratory of the University of Glasgow (UK) (Table 1). The results were subjected to Bayesian modeling to offer a more precise chronological framework. The exact dates of the construction (1481) and abandonment of the hospital (1780) were used as priors of the Bayesian models. OxCal software version 4.4 (Bronk Ramsey 2009) and the IntCal 20 calibration curve (Reimer et al. 2020) served to design the calibrations, plots, and modelings (Table 1). A multiphase uniform Bayesian model (Bronk Ramsey 2009) was also developed to corroborate the potential two phases of occupation.
Sampling
Archaeological remains were manually recovered during the excavation and their stratigraphic position was recorded within the identified structures (Mendoza Medina et al. 2012). Sediments were not screened. Therefore, faunal remains of small size (i.e., teeth, carpals, tarsals, and phalanges) and from small vertebrates might have been lost. Consequently, the representation of these sort of skeletal elements and taxa may be negatively biased in our samples.
Zooarchaeological Analysis
All faunal remains were observed with led lighted magnifying glasses of different strengths (x3, x10). Taxonomic identification was carried out with the aid of Las Palmas Zooarchaeology collection (LPZ henceforth) of the Archaeology Laboratory at the Department of Historical Sciences at ULPGC. This collection includes modern specimens from the species represented in the San Martín samples. Taxonomic identification was complemented by consulting osteological manuals (Barone 1976; Cohen and Serjeantson 1996; Hillson 2005; Schmid 1972). Diagnostic characters among the caprine remains were analyzed to differentiate goat from sheep (Boessneck 1969; Meco Cabrera 1992; Prummel and Frisch 1986). Rabbits and pigeons were registered as wild taxa as the historical sources contemporary to the HSM note them as hunted species (Ruíz Melgarejo 1974 [1531]). However, they also might have been bred in domestic contexts in the city of Las Palmas.
The Number of analyzed Specimens (NSP) was divided into two groups: (1) Number of Identified Specimens (NISP) prioritizing the minor taxonomic category, and (2) Unidentified remains (NID). The NID were classified into general groups according to the characteristics of the cortical bone: medium mammals (MSM), large mammals (LSM), small vertebrates such as rodents and reptiles (SVER), birds (AVES), and splinters devoid of identifiable diagnostic features (UNI). NISP values served as the basic measure to quantify the assemblage and to make comparisons with other faunal studies in the discussion (Cannon 2013; Lyman 1994, 2008; O’Connor 2000, 2003). The Minimum Number of Elements (MNE) was established based on the sum of specimens according to the diagnostic regions of each bone element and by considering zygopodium elements as independent. The MNE was used to calculate anatomical distribution patterns, taxonomic frequency, Minimum Number of Individuals (MNI) and to estimate mortality profiles. The well-represented taxa (MNE = > 100) offered the option to calculate the Minimum Animal Units (MAU = MNEi/Number of times that bone element i appears in a whole animal) and their percentage (%MAU = MAU x 100/MAU maximum observed in the faunal group) (Lyman 1994). The %MAU was used to estimate the frequency of anatomical segments and to correct sample fragmentation (O’Connor 2000).
Identification of adult and non-adult animals was based on the degree of epiphyseal fusion (Silver 1969). Observations of cortical surfaces and entheses, as well as degenerative pathologies of the joints, were also used to differentiate them (Baker and Brothwell 1980). A higher level of ontogenetic definition was observed among non-adult caprines, suidae, and rabbit specimens bearing these diagnostic elements (Hillson 2005; Jones 2006; Prummel 1987; Silver 1969). Sex was estimated from anatomical regions (i.e., pelvis) for caprines, dentition was used for pigs, and the presence/absence of tarsometatarsal spurs for chicken (Hatting 1995; Zeder 2006).
Taphonomic conditions were evaluated based on LPZ and specific specialized manuals (Fernández-Jalvo and Andrews 2016). This involved recording the presence/absence of rodent incisor crenulated or carnivore puncture marks. Traces of burning and cutting were noted for all the skeletal elements in order to identify the handling and consumption of the carcasses. Thermal alteration of cortical bone surfaces was recorded following the five categories registered by Costamagno et al. (2009): 0 = no heat alteration; 1 = partial thermal disturbance; 2 = charred (black); 3 = charred (gray); 4 = charred (white). Finally, the analysis focused on butchery traces, distinguishing between knife cuts and chopping marks. This included recording the morphology, distribution, orientation, and relationship of the traces observed with different processing and consumption strategies (Greenfield 2000; Seetah 2018; Soulier and Costamagno 2017).
The data set was recorded in FileMaker TIPZOO® application which specifically serves to manage zooarchaeological information (Discamps 2021). Finally, the results were depicted graphically by means of vector schemes processed with Inkscape 1.0.2 (e86c8708) and QGis (Orton 2010; Yvinec et al. 2007).
Results
Radiocarbon Dating and Bayesian Modeling
The results place all the radiocarbon dates between the mid-fifteenth century and the outset of the twentieth century. The resulting model yielded good agreement indices (Fig. 3; Amodel = 77.9; Aoverall = 80.5). Hence the modeling delimits the absolute chronology of the two phases of construction: the first from 1480 to the first half of the seventeenth century (Start:1475–1620, End:1490–1640; 95.4%) and the second from the second half of the seventeenth century to 1780 (Start: 1535–1660, End: 1670–1780; 95.4%). These results also suggest that the transition between each of the construction phases was very brief due to the overlap between their ends and beginnings. There is likewise a chronological coherence between the dates of each phase indicating that the stratigraphic layers, despite containing older materials in secondary position, do reflect the two temporal sequences. Thus, integrating the data from the Bayesian model with the relative chronology garnered from the pottery yielded two chronocultural phases for the HSM during the Modern Era: Phase I (1480 - late sixteenth century) and Phase II (early seventeenth century − 1780).

Probability ranges of the radiocarbon datings of animal bones from the Hospital de San Martín. Each is represented by two lines: a light gray line indicating the 14 C calibration and a dark gray line representing the Bayesian modeling (posterior density estimates). The chart also contains the actual dates of the hospital’s initial construction and abandonment
Zooarchaeological Analysis
The faunal assemblage of the HSM consists of 2,390 remains (NSP) of which 1,929 could be identified to taxon level (NISP) (Tables 2 and 3). The domestic species comprise cattle, sheep, goat, pig, dog, cat, and chicken, whereas the wild taxa are rabbit, pigeon, and lizard (Tables 2 and 3). Most of the teeth are preserved still attached in the alveoli of their mandibles or maxillae (Table 4). A modest number of traces of gnawing linked to rodents (272/NSP = 11%) and carnivores (203/NSP = 8%) evidences the presence of these other species on site, too. This pattern is consistent with data collected during the archaeological intervention suggesting that most of the stratigraphic levels were sealed and hardly disturbed by postdepositional actions.
Phase I (fifteenth - sixteenth centuries)
This phase contains 1,362 NISPs representing 71% of the total sample (NISP = 1362/1929). Most are from the fills of levels such as SU52 (NISP = 315) and SU84 (NISP = 595) (see Table 2). Caprines (sheep/goats) are the best represented (62% MNE) (see Table 2). At the species level, a MNE of 44 Capra hircus and a MNE of 26 Ovis aries was identified. The medial portions of the hind and forelimbs are the best represented (38 MNE tibiae diaphyses and 34 MNE radii diaphyses; see Fig. 4). The frequency of immature animals calculated from the state of epiphyseal fusion attains 43%, with the peak of mortality of this group between 2.5 and 3.5 years (Table 5). Based on the morphology of a left acetabulum it was possible to recognize one female individual.

Schema of the frequency (%MAU) of caprines for Phase I
Anthropic modifications among the caprines include Grade 1 (MNE = 398) and Grade 2 (MNE = 19) thermal alterations mainly concentrated on metapodials (MNE = 42) and tibiae (MNE = 28). Knife cuts (n = 102) and chopping marks (n = 73) occur more often in the appendicular skeleton, particularly on the neck of scapulae, the proximal epiphyses of ulnae and femora. The axial skeleton, in turn, reveals knife cuts on the ventral area of ribs and longitudinally split vertebrae.
Pig, with 204 specimens and a MNE of 96, is the second-best represented taxon (15% NISP, 18% MNE). At the anatomical level, the eight ulnae recovered suggest a higher deposition of forelimbs. 57% of the elements with ontogenic diagnostic features correspond to immature specimens. This is highlighted by piglets at ages of less than 1.5 years (see Table 5). Furthermore, the assemblage shows two isolated canines of a male. Grade 1 thermal alteration, frequent among metapodials, humeri, and ulnae, is observed on more than half of the specimens (NISP = 123/204; 60%). Knife cuts (n = 13) and chopping marks (n = 16) mainly affect the appendicular skeleton. They were observed on the proximal epiphyses of ulnae and transversal chop marks were recorded on the distal epiphyses of three humeri. But they are also displayed on the axial skeleton (i.e., axis) and the skull (i.e., lingual and buccal sides of two jaws, respectively).
Cattle, the third most abundant species, is represented by 125 specimens (9% NISP) corresponding to a MNE of 67 (13% MNE, see Table 2). Although these parameters do not allow a more detailed analysis of anatomical distribution, a trend appears to emerge indicating a preference for forelimbs. The scant cases of epiphyseal fusion point to the preferential slaughter of adults (see Table 5). Anthropic modifications include Grade 1 thermal alterations (NISP = 64) as well as 19 chopping and 38 knife cuts. Noteworthy is the higher frequency of butchery marks among the phalanges in the form of six transversal cuts on three elements.
Other domestic taxa are chicken (5 NISP, one tarsometatarsal of a female), dog (NISP = 6; MNE = 2), and cat (NISP = 1; MNE = 1). Rabbit stands out as the most abundant wild species (93 NISP making up 29 MNE) (Table 2) with jaw and pelvis as the most common elements. Three bones of rabbit (2 humeri and 1 tibia) are estimated to be immature individuals slaughtered at between eight and nine months. Traces of thermoalteration and processing of the carcasses were recorded on several rabbit specimens (NISP = 28 and 6, respectively). Among the butchered bones (MNE = 6) a single transversal cut on an ilium and two diagonal cuts on tibiae stand out.
Phase II (seventeenth - eighteenth centuries)
Of the total 837 specimens collected from this phase, it was possible to identify 567, that is 29% of the total sample (NISP = 567/1929; see Table 3). Most derive from layers SU37 (NISP = 232) and SU71 (NISP = 222) of Recinto 3, a floor and its associated fill. Caprines are the most abundant taxa (NISP = 270; 48%) with a total of 115 MNE (41%), which can be broken down into 15 MNE of Capra hircus and 3 MNE of Ovis aries. The anatomical distribution pattern reveals a greater representation of medial hindlimbs (15 tibiae shafts) and forequarters (12 radii and 11 scapulae; Fig. 5). Immature animal frequency is estimated at 47%, with the highest mortality among the 1.5-2.5-year group. Two acetabula (left and right) are from female individuals.

Schema of the frequency (%MAU) of caprines for Phase II
Alterations of anthropic origin include burnt metapodials (MNE = 5) and femora (MNE = 4) (Grade 1), as well as butchery marks on radii (23 knife cuts) and humeri (22 chopping marks). Percussion traces are for the most part visible on the axial skeleton, concentrating on vertebrae.
Pig, with 68 NISP (12%) and 36 MNE (13%; see Table 3), is the second most common domestic taxon represented, most frequently by forelimbs. The scarce data on epiphyseal fusion suggest a balance in terms of the consumption of immature and adult animals (see Table 5). Thermal alteration is visible on 22 NISP with a special incidence among scapulae (MNE = 4). This sample also reveals 10 knife cuts and one chopping mark (NISP = 7; MNE = 6), highlighted by a proximal ulna shaft bearing four fine cuts.
The number of adult cattle remains is modest (NISP = 30; MNE = 21) rendering it impossible to establish the anatomical profile (see Table 3). This taxon also reveals longitudinal percussion cuts on vertebrae (MNE = 4) (Fig. 6E), two thin transversal cuts on carpals (MNE = 2) and three on the proximal epiphyses of tarsals (MNE = 2).

A Chicken synsacrum sectioned by a metal blade, dorsal and ventral view; B Left rabbit tibia bearing multiple knife cuts on the diaphysis; C Left rabbit tibia fresh fracture and evidence of consumption; D Right caprine metacarpal with transversal knife cuts on the anterior part of proximal epiphysis; E Longitudinal chopping mark on a cattle vertebra
Chicken is represented by 37 NISP (MNE = 32), notably adult wings and legs (ulnae 7 MNE; femora 3 MNE). None could be sexed. Apart from thermally altered elements, four bones present knife cuts and a synsacrum was chopped through (Fig. 6A).
Rabbit stands out. It is in general terms the second most common taxon (NISP = 140; 25%; MNE = 57; 20%) (see Table 3). Its anatomical distribution pattern reveals a higher number of forelimbs (humeri 9 MNE). NISP estimations place nine as young: three femora and three humeri would belong to individuals of less than four and eight months, respectively. They also bear evidence of thermal alteration (NISP = 21) as well as fine knife marks (NISP = 4) directly linking them to anthropic consumption (Fig. 6B and C). The spectrum of fauna is completed by the bones of cat (NISP = 4), pigeon (NISP = 17), and 1 NISP of lizard (Gallotia sp.) (see Table 3).
Discussion
The study of the faunal assemblage of the HSM offers the first perspective from an archaeological standpoint of the land animals exploited and consumed on the island of Gran Canaria during the Modern Era. It also yields a comparative framework to grasp the role of European colonization and that of terrestrial animals in the subsistence and consumption practices of the Canarian society by that moment. The islands were the only Macaronesian archipelago colonized by the Crown of Castile in its expansion across the Atlantic. The archipelago also played a remarkable role in this Atlantic dispersal as it was previously inhabited by agricultural populations of Berber origin that had access to a number of different domestic animals before the European conquest. They thus differed from other processes in the Caribbean and the American continent that benefitted from a greater access to resources serving the development of large ungulates. Nevertheless, it must be noted that the context of the finds, an old hospital, could reflect the diet of patients, a specific segment of the population (Huggon 2018), and not necessarily line up with the eating habits of the general Canarian population (Pigière et al. 2004). Even so, this study offers a preliminary assessment of the types of animals consumed and exploited in the Las Palmas urban context during the centuries after the European conquest.
The Animals Consumed at the Hospital de San Martín
The zooarchaeological analysis demonstrates that sheep and goats, followed by pigs and cattle, were the main sources of meat (Table 6). The anatomical distribution and slaughter profiles point to a preferential selection of adult and subadult individuals attaining their meat optimum (see Table 5). The faunal remains also reflect a greater abundance of goats over sheep throughout the entire sequence. According to the MNE for every element of sheep in Phase I there are two goats, a ratio that in Phase II increases to 1:5 (see Tables 2 and 3). This higher prevalence of goat contrasts with other late medieval assemblages elsewhere in Europe (Albarella 1997; Davis 2008; Grau Sologestoa 2020) and contemporary finds in the Iberian Peninsula where sheep usually outnumber goats (i.e., Puerto de Palos (Riquelme 2020), Castelo de Evoramonte (Costa 2009), and Silves (Cardoso and Gomes 1996; Valera Gomes et al. 1996). The pattern of San Martín also diverges from contemporary faunal associations in the Caribbean such as Puerto Real on Hispaniola or St. Agustine in Florida (Reitz 1992; Reitz and Waselkov 2015) where the numbers of native fauna and cattle outweigh that of caprines.
The relative predominance of goats among the HSM fauna complex resembles that of other islands and high mountain landscapes (Atici et al. 2017; Halstead 1996; Navarrete et al. 2022; Ramis 2018), and, above all, the type of livestock of the Indigenous population of Gran Canaria (Castellano Alonso et al. 2018; Pais Pais 1996). Both historical and ethnohistorical sources reveal an abundance of goats and that a significant portion of their management since the initial moments of the European colonization rested in Indigenous hands (Alberto-Barroso 2004; Betancor Quintana 2003; Ruíz Melgarejo 1974 [1531]). The key role of goats as a means of exchange, income, and inheritance as recorded in the wills of the Gran Canarian population in the early sixteenth century (Tabares and Santana Rodríguez 2018) needs to be highlighted. The availability of this taxon and the familiarity of the natives with its management explains its relative abundance among the finds of the oldest phase of San Martín. It must also be noted that goats were well adapted to the semi-arid environment and limited ecological resources of the Canary Islands (Castro et al. 2017). The continuity of caprines husbandry was certainly encouraged by the particular characteristics of these animals as appears to have also occurred with barley of indigenous origin whose adaptive capacities favored its adaptation and use – even to the present – in Gran Canaria’s environment (Hagenblad and Morales 2020).
Our results thus suggest a predisposition among European settlers to exploit both this well adapted animal and the knowledge of its management by the Indigenous populations. This is comparable to the breeding of camelids in South America after the Castilian conquest, although the introduction of sheep since the sixteenth century did lead to substantial changes in the composition and management of the Andean Altiplano herds (Wakild 2021; Wheeler et al. 1995). It is also noteworthy that the Canarian caprine livestock contributed to the dispersal of these animals throughout the American continent during the Spanish colonization (Álvarez et al. 2013; Amills et al. 2009; Wanjala et al. 2021). The great number of goats on the islands undoubtedly rendered this species profitable among the Atlantic colonies. From this perspective, it is hoped that future paleogenetic analyses on goat samples from the Modern Era will help to understand if goats were imported from Europe or North Africa after the conquest or if European colonizers continue resorting indigenous goats.
Pig is the second most abundant taxon, with a similar trend in both phases for the consumption of forelimbs of young individuals of less than 1.5 years of age (see Table 5). Despite the fact that the sample size is limited (MNE = < 100), the results do not change over time. Yet, the faunal assemblage of the HSM reflects a decrease in the relative frequency of pig with respect to the zooarchaeological record of late Gran Canaria’s Indigenous sites and their period of contact with the Europeans (see Table 6). The few sites studied to date suggest that in pre-European domestic contexts this taxon surpassed caprines indicating it was much more abundant than in the Modern Era (Alberto-Barroso et al. 2017; Castellano Alonso et al. 2016) (Fig. 7). The reduction in pig relative frequency may have begun in the period of contact as evidenced by the fifteenth century faunal samples from the Cueva Pintada (Castellano Alonso et al. 2018). This site contains the remains of Agaldar, one of the largest settlements at the time of the European arrival, located in the northwestern side of the island (Onrubia Pintado 2003). The decrease of pigs is also observed in the late faunal assemblage of the Hospital of San Martín. This trend is significant given the widespread role pig had in late medieval European culinary traditions (García García et al. 2021).

Frequency (%NISP) of the main domesticates by period. Data from Table 6
The present work confirms that the arrival of the Europeans led to the introduction of new faunal species. Finds of cattle, chicken, rabbit, and cat remains differ from the early to the late phase at the HSM (see Tables 2 and 3). With the exception of cat, these taxa are not present in Indigenous pre-European contexts. The few cases where they have been found have not benefitted from radiocarbon dating to assess whether they are intrusive elements (Alberto-Barroso et al. 2017; Castellano Alonso et al. 2016, 2018; Hernández Marrero et al. 2016; Rando Reyes et al. 1996,1997). Historical written sources document the importation of alien species (López de Ayala 1780 [1393]; Ruíz Melgarejo 1974 [1531]; Sosa 1994 [1678–88]). Recent genetic studies also highlight the introduction of large mammals, sheep, pigs, dogs, and brown rats (Álvarez et al. 2013; Ferrando et al. 2015; López et al. 2013; Olalde et al. 2015; Suárez et al. 2013). However, the precise timing of these animal translocations in the Modern Era remains unclear as no paleogenomic analyses on directly-dated specimens have been conducted so far (Table 7).
Cattle is the third type of livestock (see Table 6). Its low numbers contrast with the data available for Puerto de Palos, a contemporary site in the south of the Iberian Peninsula (fifteenth - sixteenth CE) where bovids are relatively abundant (24% NISP) (Riquelme 2020). They also differ from the frequency attested in Hispanic colonies in the Caribbean Greater Antilles, whose colonization took place after that of the Canary Islands, and where bovines stand out among the faunal assemblages (Reitz 1986; Delsol et al. 2023).
The proliferation of bovids throughout a territory like the island of Gran Canaria must have been complicated by its abrupt orography, scarce pastures, and limited sources of water (Capote et al. 2002). The early written sources likewise record the difficulty and high rate of mortality associated with transporting large mammals between the Iberian Peninsula and the archipelago (Ruíz Melgarejo 1974 [1531]). The disposal of their carcasses into the sea is in fact behind the epithet “Golfo de las Yeguas” (Gulf of the Mares) given to this maritime stretch along the African coastline (Fernández de Oviedo y Valsés 1851 [1535]). The parceling of the land, population growth, and the solvency of smaller livestock during the colonial period were factors that could have limited the development of bovine herds. This is reflected by the low proportion of cattle and absence of other large mammals (i.e., horses and asses) among the assemblage of San Martín and, consequently, their scant role in both occupation phases.
This trend is likewise bolstered by the absence of bovids in the analyzed contexts from Cueva Pintada (Castellano Alonso et al. 2018). However, it must be borne in mind that cattle were not only valued for their meat or milk, but for traction. In this sense, the low number of their remains and the large representation of adult specimens may indicate that they were mainly bred for working (and not for meat consumption) (Aznar Vallejo et al. 2000; Luxán et al. 2009; Ruíz Melgarejo 1974 [1531]; Tabares and Santana Rodríguez 2018).
This study advances the earliest clear archaeological evidence of the consumption of domestic fowl and rabbit in the Canary Islands, two species that accompanied the Europeans into the Atlantic as they colonized the islands of Macaronesia, the Caribbean, and the American mainland (Delibes and Delibes-Mateos 2015; Herrera et al. 2020; Traveset et al. 2009). An increase of chicken and rabbit remains from Phase I to II evidence their important role in the diet of the residents of the hospital complex (see Tables 2 and 3). Both taxa reveal signs of having been processed for human consumption (see Figs. 6 and 7). The prominence of these species as a food resource expands at the Hospital of San Martín as the Modern Era progresses, reflecting a diversification of the sources of meat, albeit not superseding that of caprines. The increase in chicken consumption seems to be a relatively recent phenomenon that was established during the Middle Ages (Best et al. 2022). Historical sources also point out the key role of white poultry in the treatment of Canarian patients (Coello Gómez et al. 1980) and in other European hospitals from both medieval (Barger 1998; Martínez García 1995; Wall 1980) and modern times (Morris 2004). White meat at the time was especially recommended for the treatment of the sick (Coello Gómez et al. 1980; Martínez García 1995; Villagrasa-Elías 2022). In this sense, the assemblage of San Martín could reflect practices serving for the care of patients.
Finally, it is remarkable the absence of autochthonous taxa at San Martín with the exception of lizard and probably pigeons. The lack of comparative osteological studies hampers the identification of the two autochthonous species of pigeons present in Gran Canaria (Martín et al. 2013) as well as of their domestic counterparts. That could relate to the scarcity of large native mammals in the insular ecosystem. In fact, old texts describe rabbit hunting in the island’s mountains since the outset of the sixteenth century, noting the prohibition of starting fires to corner them due to the great risk to forests (Ruíz Melgarejo 1974 [1531]). In other words, the main hunted animals were species that were introduced, not native to the island. However, it cannot be ruled out that the fragility of certain taxa and the recovery methods used during the excavation led to the absence of wild birds or small commensal mammals such as rats and mice which are otherwise certified by bones bearing gnawing traces.
Other Activities: Slaughter and Tanning
The assemblage of the HSM also sheds light on anthropogenic uses of animal carcasses. The representation of body parts, biological profiles, and processing marks suggest a selection intended for meat consumption. Butchery traces left on the bones are compatible with the metal tools introduced by the Europeans, implements that could not be manufactured on the archipelago due to the absence of metal ores (see Fig. 6). The recurring location of these marks suggest a standardized handling of the carcasses, at least in the case of caprines (Fig. 8). The evisceration and the use of tallow is gleaned from butchery traces in the ventral area of ribs and vertebrae. The latter also often present longitudinal chopping intended to divide the carcass along the spine. Moreover, most of the axes and atlases bear signs indicating the removal of the head. Traces of chopping and fine slicing marks are also frequent on both front and hind legs, with a particular concentration on the proximal epiphyses of radii and ulnae on both caprines and pigs. They are related to dismembering the lower part of the leg from the upper quarter and also to filleting their meat (Soulier and Costamagno 2017).

A Schema indicating the frequencies of thermal alterations per bones in Phase I and II. B Schema indicating the traces and number of cuttings per element according to hypothetical functions of the caprine slaughtering process in Phase I and II. CRA: cranium; MAN: mandibula; VERT: vertebrae; COS: ribs; SC: scapula; HUM: humerus; RA: radius; UL: ulna; MC: metacarpal; CARP: Carpals; PEL: pelvis; FEM: femur; TIB: tibia; MT: metatarsal; TAR: tarsal: PHX: phalanges
A similar pattern of animal carcass management is recorded in medieval and post-medieval European contexts (Grant 1987). Yet in the Canary Islands the lack of comparative datasets is challenging. Most zooarchaeological analyses have not systematically recorded butchery marks in order to detect a standardized pattern as it has been observed in HSM. The historical textual records from Gran Canaria indicate that butchery was carried out by specialized workers of a marginalized social group during the Indigenous Period (Alberto-Barroso 2004; Zurara 1978 [1452–1453]; Rodríguez-Rodríguez 1997; Seetah 2018). Therefore, the use of metal tools to process the animal carcass consumed at HSM is clearly linked to European traditions.
The skeletal remains also reveal knife marks among carpals, tarsals, metapodials, and phalanges of caprines and cattle that probably relate to skinning and tanning of hides (see Figs. 6D and 8B). Leather was an object of trade between Europeans and Natives from the outset of their contacts and it was fundamental to the local consumption of the newly implanted society in Gran Canaria (Carriazo Arroquia 1946; González Marrero 1996). The tanning of caprine hides rose to great heights in the sixteenth century substantiated by exports to continental America usually with wine cargoes (Lobo Cabrera 1994). During the sixteenth century in Las Palmas some ordinances reported butchery guilds. Meat could only be sold in butcher shops and “tocinerías” (shops specialized in the sale of lard) regulated by the colonial administration. This surveillance wanted to ensure a regular supply and control over the prices (Ruíz Melgarejo 1974 [1531]).
Conclusions
The zooarchaeological analysis of faunal remains from the old HSM in the city of Las Palmas sheds new light on the dietary practices of this specific site. It likewise gathers compelling data on the role of livestock in the local economy. The data collected provide for the first-time information on the consumption of animals translocated by Europeans during the expansion across the Atlantic. The results point to goats meat, and to a lesser extent pork and beef, as part of the diet of hospital patients. An increase in the consumption of chicken and rabbit from Phase I to Phase II appears to reflect a full integration of these two species into the diet of Canarian society as the Modern Era progressed. It cannot be excluded that the increase of chicken remains stems from the fact that white meat at the time was especially recommended for the treatment of the sick.
The relative frequency of domestic taxa at San Martín differs considerably from that of other contemporary sites under the Crown of Castile either in the Iberian Peninsula, the Caribbean or Continental America. The data, in combination with notions gleaned from primary written sources, suggest that the new European settlers took advantage of the availability of indigenous caprines to improve the stock over the following centuries. This decision was certainly encouraged by the particular characteristics of these animals as appears to have also occurred with barley of indigenous origin whose natural capacities favored its adaptation and use – even to the present – in the semi-arid environment of Gran Canaria.
The current study thus offers a comparative base for subsequent Canarian zooarchaeological analyses. Future lines of research should focus on other urban and rural contexts so as to evaluate if the faunal assemblage of the HSM reflect the eating habits of the majority of the population of the Modern Era, or if they are limited to the specific segment of the population, in this case hospital patients. In addition, it is hoped that zooarchaeological studies will be performed in other islands of the archipelago to explore the occurrence of similar patterns to those here described or if, on the contrary, they reveal other circumstances specific to each island. In any case, this study clearly validates the role of zooarchaeology in providing direct information on the relation between humans and animals in the Modern Era, and the singularities that characterize each region and period.
Data Availability
The authors confirm that all data generated or analysed during this study are included in the article.
References
Albarella, U. (1997). Size, power, wool and veal: zooarchaeological evidence for late medieval innovations. In De Boe, G. and Verhaeghe, F. (eds.), Environment and Subsistence in Medieval Europe - Papers of the Medieval Europe Brugge 1997 Conference. 9: 19–31.
Alberto-Barroso, V. (1998). Los otros animales: consumo de Gallotia goliath y Canariomys bravoi en la prehistoria de Tenerife. El Museo Canario 53: 59–84.
Alberto-Barroso, V. (2004). De carne y hueso: la ganadería en época prehispánica. El Pajar 18: 4–8.
Alberto-Barroso, V., Moreno-Benítez, M., Alamón-Núñez, M., Suárez Medina, I., and Mendoza Medina, F. (2017). Estudio zooarqueológico de La Restinga (Gran Canaria, España): datos para la definición de un modelo productivo. Actas XXII Coloquio de Historia Canario-Americana: las Ciudades del Mundo Atlántico, Pasado, Presente y Futuro. Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria, pp. 1–17.
Alcover, J. A., Rando Reyes, J. C., García-Talavera, F., Hutterer, R., Michaux, J., Trias, M., and Navarro Mederos, J. F. (2009). A reappraisal of the stratigraphy of Cueva del Llano (Fuerteventura) and the chronology of the introduction of the house mouse (Mus musculus) into the Canary Islands. Palaeogeography, Palaeoclimatology, Palaeoecology 277(3–4): 184–190.
Álvarez, I., Capote, J., Traoré, A., Fonseca, N., Pérez, K., Cuervo, M., Fernández, I., and Goyache, F. (2013). Mitochondrial analysis sheds light on the origin of hair sheep. Animal Genetics 44: 344–347.
Amills, M., Ramírez, O., Tomás, A., Badaoui, B., Marmi, J., Acosta, J., Sánchez, A., and Capote, J. (2009). Mitochondrial DNA diversity and origins of South and Central American goats. Animal Genetics 3: 315–322.
Armenteros Martínez, I. (2018). The Canary Islands as an Area of Interconnectivity between the Mediterranean and the Atlantic (Fourteenth-Sixteenth Centuries). In Jaspert, N., and Kolditz, S. (eds.), Entre mers - Outre-mer: Spaces, Modes and Agents of Indo-Mediterranean Connectivity. Heidelberg University, Heidelberg, pp. 201–216.
Arnaiz-Villena, A., Álvarez-Tejado, M., Ruíz-del-Valle, V., García-de-la-Torre, C., Varela, P., Recio, M. J., Ferre, S., and Martínez-Laso, J. (1999). Rapid radiation of Canaries (Genus Serinus). Molecular Biology and Evolution 16(1): 2–11.
Arnay-de-la-Rosa, M. (2009). La arqueología histórica en Canarias: el yacimiento sepulcral de la iglesia de Nuestra Señora de la Concepción de Santa Cruz de Tenerife. Arqueología Iberoamericana 1(3): 21–36.
Arnay-de-la-Rosa, M., Ordóñez, A. C., and Pérez Álvarez, A. R. (2015). Evidencias arqueológicas del movimiento de personas y productos en Canarias en el siglo XVIII: esclavos y tabaco en el yacimiento arqueológico de la Iglesia de Nuestra Señora de La Concepción de Santa Cruz de Tenerife. Vegueta: Anuario de la Facultad de Geografía e Historia 15: 37–64.
Atoche Peña, P. and Ramírez Rodríguez, M. Á. (2017). C14 references and cultural sequence in the proto-history of Lanzarote (Canary Islands). In Barceló, J. A., Bogdanovic, I., and Morell, B. (eds.), Actas Congreso de Cronometrías Para La Historia de La Península Ibérica (IberCronos 2016). Universidad Autónoma de Barcelona, Barcelona, pp. 272–285.
Atici, L., Pilaar Birch, S. E., and Erdoğu, B. (2017). Spread of domestic animals across Neolithic western Anatolia: New zooarchaeological evidence from Uğurlu Höyük, the island of Gökçeada, Turkey. PLoS One 12(10): e0186519.
Aznar Vallejo, E. (1983). La Integración de las Islas Canarias en la Corona de Castilla, 1478–1526: Aspectos Administrativos, Sociales y Económicos. Secretariado de Publicaciones de la Universidad de La Laguna, San Cristóbal de La Laguna, Tenerife.
Aznar Vallejo, E. (2008). Exploración y colonización en la configuración de la Europa Atlántica. Historia. Instituciones. Documentos 35:45–61.
Aznar Vallejo, E., Corbella, D., Pico, B., and Tejera Gaspar, A. (2006). Le Canarien: Retrato de Dos Mundos. Instituto de Estudios Canarios, San Cristóbal de La Laguna, Tenerife.
Aznar Vallejo, E., González Marrero, M. del C., and Larraz Mora, A. (2000). Las cuentas de la armada, fuente para el estudio de la vida cotidiana. Gran Canaria en 1496. In Morales Padrón, F. (ed.), Actas XIII Coloquio de Historia Canario-Americana, VIII Congreso Internacional de Historia de America (1998). Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria, pp. 2244–2259.
Baker, J. R. and Brothwell, D. (1980). Animal Diseases in Archaeology. Academic Press, London.
Barger, G. (1998). Animal bone. In Price, R. and Ponsford. M. (eds.), St Bartholomew’s Hospital, Bristol. The Excavation of a Medieval Hospital: 1976-8. Council for British Archaeology, London, pp. 181–192.
Barone, R. (1976). Anatomie Comparée des Mammifères Domestiques. Vigot Frères, Paris
Best, J., Doherty, S., Armit, I., Boev, Z., Büster, L., Cunliffe, B., Foster, A., Frimet, B., Hamilton-Dyer, S., Higham, T., Lebrasseur, O., Miller, H., Peters, J., Seigle, M., Skelton, C., Symmons, R., Thomas, R., Trentacoste, A., Maltby, M., Larson, G., and Sykes, N. (2022). Redefining the timing and circumstances of the chicken’s introduction to Europe and north-west Africa. Antiquity 96(388): 868–882.
Betancor Quintana, G. (2003). Los Indígenas en la Formación de la Moderna Sociedad Canaria: Integración y Aculturación de Canarios, Gomeros y Guanches (1496–1525). Doctoral dissertation, Universidad de Las Palmas de Gran Canaria, Gran Canaria.
Birkhead, T. R., Schulze-Hagen, K., and Kinzelbach, R. (2004). Domestication of the canary, Serinus canaria - the change from green to yellow. Archives of Natural History 31(1): 50–56.
Boessneck, J. (1969). Osteological differences between sheep (Ovis aries Linné) and goat (Capra hircus Linné). In Brothwell, D. and Higg, E. (eds.), Science in Archaeology. Thames and Hudson, London, pp. 2331–2358.
Bosch Millares, J. (1940). El Hospital de San Martín: Estudio Histórico desde su Fundación Hasta Nuestros Días. Minerva, Las Palmas de Gran Canaria.
Bronk Ramsey, C. (2009). Bayesian analysis of radiocarbon dates. Radiocarbon 51(1): 337–360.
Bruquetas de Castro, F. (2015). Los genoveses en Canarias: Siglos XVI y XVII. In Iglesias Rodríguez, J. J., Pérez García, R. M., and Fernández Chaves, M. F. (eds.), Comercio y Cultura en la Edad Moderna - XIII Reunión Científica de la Fundación Española de Historia Moderna. Editorial universitaria de Sevilla, Sevilla, pp. 57–68.
Cadamosto, A. (2017 [1456]). Los Viajes Africanos de Alvise Cadamosto (1455–1456). Aznar Vallejo, E., Corbella Díaz, D, Tejera Gaspar, A. (eds.), Instituto de Estudios Canarios, San Cristóbal de La Laguna.
Cannon, M. D. (2013). NISP, bone fragmentation, and the measurement of taxonomic abundance. Journal of Archaeological Method and Theory 20: 397–419.
Capote, J., Argüello Henríquez, A., López, J. L., Montesdeoca, M. C., Amills, M., and Tejera Gaspar, A. (2002). Introducción de caprinos en las islas Canarias y América: una visión desde punto de vista etnológico e histórico. In Peris Palau, B., Molina Pons, P., Lorente Alonso, M., and García Muñoz, Á. (eds.), XXVII Jornada Científicas y VI Jornadas Internacionales de la Sociedad Española de Ovinotecnia y Caprinotecnia. Servicio de Publicaciones de UCH-CEU, Valencia.
Cardoso, J. L. and Gomes, M. V. (1996). Contributo para o estudo das faunas encontradas no poço-cisterna de Silves (séculos XV-XVI). XELB: Revista de Arqueologia, Arte, Etnologia e História 3: 207–268.
Carriazo Arroquia, J. d. M. (1946). El capítulo de Canarias en la "Crónica de Juan II": (versión original, inédita, de Alvar García de Santa María). Revista de Historia 73(12): 1–9.
Castellano Alonso, P., Moreno García, M., Alberto-Barroso, V., Rodríguez-Rodríguez, A., Arencibia Espinosa, A., and Blanco Sucino, D. (2016). El Lomo de Los Melones (Telde). Explotación prehispánica del ganado doméstico en un enclave costero. In Acosta Guerrero, E. (ed.), Actas XXI Coloquio de Historia Canario-Americana. Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria, pp. 1–14.
Castellano Alonso, P., Moreno García, M., Rodríguez-Rodríguez, A., Sáenz Sagasti, J. I., and Onrubia Pintado, J. (2018). Gestión de la ganadería y patrones de consumo de una comunidad indígena expuesta al fenómeno colonial: el caso de la Estructura 12 de la Cueva Pintada (Gran Canaria, España). Archaeofauna 27: 37–56.
Castro, N., Argëllo, A., and Capote, J. (2017). The Canary islands’ goat breeds (majorera, tinerfeña, and palmera): an example of adaptation to harsh conditions. In Simões, J. and Carlos, G. (eds.), Sustainable Goat Production in Adverse Environments: Volume II. Springer, Cham, pp. 221–231.
Coello Gómez, M. I., Rodríguez González, M., and Parrilla López, A. (1980). Protocolos de Alonso Guitierrez (1522–1525). Aula de Cultura del Cabildo Insular de Tenerife, Instituto de Estudios Canarios, Santa Cruz de Tenerife.
Cohen, A. and Serjeantson, D. (1996). A Manual for the Identification of Bird Bones from Archaeological Sites. Archetype, London.
Costa, C. (2009). A fauna mamalógica do silo 1 do Castelo de Evoramonte (Estremoz). Vipasca. Arqueologia e História 3(2): 39–49.
Costamagno, S., Théry-Parisot, I., Castel, J. C., and Brugal, J.-P. (2009). Combustible ou non? analyse multifactorielle et modèles explicatifs sur des ossements brûlés paléolithiques. In Théry-Parisot, I., Costamagno, S. and Henry, A. (eds.), Gestion des Combustibles au Paléolithique et au Mésolithique: Nouveaux Outils, Nouvelles Interprétations. UISPP, XV congress (Lisbon, 4–9 septembre 2006). Archaeopress, Oxford, pp. 69–84.
Cullen del Castillo, P. (1947). Libro Rojo de Gran Canaria o Gran Libro de Provisiones y Reales Cédulas. Excmo. Ayuntamiento de Las Palmas de Gran Canaria, Las Palmas de Gran Canaria.
Davis, S. J. M. (2008). Zooarchaeological evidence for moslem and christian improvements of sheep and cattle in Portugal. Journal of Archaeological Science 35: 991–1010.
Delgado, J., Perezgrovas, R., Camacho, M., Fresno, M. and Barba, C. (2000). The wool-less canary sheep and their relationship with the present breeds in America. Animal Genetic Resources Information 28: 27–34.
Delibes, R. and Delibes-Mateos, M. (2015). Linking historical ecology and invasion biology: some lessons from European rabbit introductions into the new world before the nineteenth century. Biological Invasions 17(9): 2505–2515.
Delsol, N., Stucky, B. J., Oswald, J. A., Reitz, E. J., Emery, K. F., Guralnick, R., and Chiang, T.-Y. (2022). Analysis of the earliest complete mtDNA genome of a Caribbean colonial horse (Equus caballus) from 16th-century Haiti. PLoS One 17(7): e0270600.
Delsol N, Stucky BJ, Oswald JA, Cobb CR, Emery KF, Guralnick R. (2023). Ancient DNA confirms diverse origins of early post-Columbian cattle in the Americas. Scientific reports 13(1): 12444.
Díaz-Sierra, I. (2022). Hidden in plain sight? looking for the indigenous agricultural fields of Gran Canaria, Agüimes and Temisas, Canary Islands, fifteenth and sixteenth centuries. Internationa Journal of Historical Archaeology 26: 434–456.
Discamps, E. (2021). TIPZOO: a touchscreen interface for palaeolithic zooarchaeology: towards making data entry and analysis easier, faster, and more reliable. Peer Community in Archaeology 1: e67.
Fajardo Spínola, F. (2004). Portugueses en Canarias en el siglo XVII. Una relación de 1626. In Morales Padrón, F. (ed.), Actas XV Coloquio de Historia Canario-Americana. Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria, pp. 310–320.
Fernández de Oviedo y Valsés, G. (1851 [1535]). Historia General y Natural de las Indias, Islas y Tierra Firme del Mar Oceano. Imprenta de la Real Academia de la Historia, Madrid.
Fernández-Jalvo, Y. and Andrews, P. (2016). Atlas of Taphonomic Identifications: 1001 + Images of Fossil and Recent Mammal Bone Modification. Springer, Dordrecht.
Ferrando, A., Manunza, A., Jordana, J., Capote, J., Pons, A., Pais Pais, J., Delgado-Darias, T., Atoche Peña, P., Cabrera, B., Martínez, A., Landi, V., Delgado, J. V., Argüello, A., Vidal, O., Lalueza-Fox, C., Ramírez, O., and Amills, M. (2015). A mitochondrial analysis reveals distinct founder effect signatures in Canarian and Balearic goats. Animal Genetics 46(4): 452–456.
Fregel, R., Ordonez, A. C., and Serrano, J. G. (2021). The demography of the Canary Islands from a genetic perspective. Human Molecular Genetics 30(R1): R64-R71.
Frutuoso, G. (1964 [c. 1590]). Las Islas Canarias (De «Saudades da Terra»). Serra Rafols, E., Régulo, J., and Pestana, S. (eds.), Instituto de Estudios Canarios, La Laguna.
García García, M., Moreno García, M., Fuentes Santos, M. d. C., and Hidalgo Prieto, R. (2021). Por sus basuras los reconoceréis: La identificación arqueozoológica de una comunidad ḏimmí cristiana en la Qurṭuba tardoandalusí (Cercadilla, siglo XII). Arqueología y Territorio Medieval 28: 1–33.
González Marrero, M. d. C. (1996). El calzado y su manufactura en Tenerife en la primera mitad del siglo XVI. Revista de Historia Canaria 178: 94–105.
González Marrero, M. d. C. and Tejera Gaspar, A. (2011). La arqueología medieval en Canarias. Boletín de Arqueología Medieval 15: 127–164.
Grant, A. (1987). Some observations on butchery in England from the Iron Age to the Medieval Period. Anthropozoologica 1: 53–58.
Grau Sologestoa, I. (2020). Livestock size change and animal husbandry between the Late Middle Ages and the Modern Era in the Basque Country and its surroundings. In Grau Sologestoa, I. and Quirós Castillo, J. A. (eds.), Arqueología de la Edad Moderna en el País Vasco y su Entorno. Archaeopress, Oxford, pp. 140–164.
Greenfield, H. J. (2000). The origins of metallurgy in the central Balkans based on the analysis of cut marks on animal bones. Environmental Archaeology 5(1): 93–106.
Hagenblad, J. and Morales, J. (2020). An evolutionary approach to the history of barley (Hordeum vulgare) cultivation in the Canary Islands. African Archaeological Review 37: 579–595.
Halstead, P. (1996). Pastoralism or household herding? Problems of scale and specialization in early Greek animal husbandry. World Archaeology 28: 20–42.
Hatting, T. (1995). Sex-related characters in the pelvic bone of domestic sheep (Ovis aries). Archaeofauna 4: 71–76.
Henríquez-Valido, P., Morales, J., Vidal-Matutano, P., Moreno-Benítez, M., Marchante-Ortega, Á., Rodríguez-Rodríguez, A., and Huchet, J. B. (2020). Archaeoentomological indicators of long-term food plant storage at the prehispanic granary of La Fortaleza (Gran Canaria, Spain). Journal of Archaeological Science 120: 105179.
Hernández Marrero, J. C. (2001). Estructura y dinamismo en las relaciones sociales guanches tras la conquista de Tenerife. Tabona: Revista de Prehistoria y de Arqueología 10: 247–266.
Hernández Marrero, J. C., Navarro Mederos, J. F., Trujillo Mora, J. M., Cancel, S., Machado Yanes, C., Pais Pais, J., Morales, J., and Rando, J. C. (2016). An approach to prehistoric shepherding in La Gomera (Canary Islands) through the study of domestic spaces. Quaternary International 414: 337–349.
Hernández Suárez, S. (2021). La importación de trigo en las islas de realengo: el caso de la palma en la segunda mitad del siglo XVI. In Acosta Guerrero, E. (ed.), Actas XXIV Coloquio de Historia Canario-Americana, Congreso Internacional ADHILAC (2020) Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria, pp. 1–12.
Herrera, M. B., Kraitsek, S., Alcalde, J. A., Quiroz, D., Revelo, H., Alvarez, L. A., Rosario, M. F., Thomson, V., Jianlin, H., Austin, J. J., and Gongora, J. (2020). European and Asian contribution to the genetic diversity of mainland South American chickens. Royal Society Open Science 7(2): 191558.
Hillson, S. (2005). Teeth (Cambridge Manuals in Archaeology). 2nd ed. Cambridge University Press, Cambridge.
Huggon, M. (2018). Medieval medicine, public health, and the medieval hospital. In Gerrard, C. and Gutiérrez, A. (eds.), The Oxford Handbook of Late Medieval Archaeology. Oxford University Press, Oxford, pp. 836–855.
Hutterer, R. (1990). Remarks on a presumed record of Felis margarita from Tenerife, Canary Island. Vieraea 19: 169–174.
Jones, E. L. (2006). Prey choice, mass collecting, and the wild European rabbit (Oryctolagus cuniculus). Journal of Anthropological Archaeology 25(3): 275–289.
Ladero Quesada, M. Á. (1966). Las cuentas de la conquista de Gran Canaria. Anuario de Estudios Atlánticos 12: 11–104.
Lobo Cabrera, M. (1998). Formas de poder y economía canaria entre los siglos XV-XVII. Investigaciones Históricas: Época Moderna y Contemporánea 18: 13–28.
Lobo Cabrera, M. (2012) La Conquista de Gran Canaria (1478–1483). Cabildo insular de Gran Canaria, Las Palmas.
Lobo Cabrera, M. (1994). Los cordobanes canarios y su exportación a Indias. In Mancomunidad de Cabildos (eds.), Actas X Coloquio de Historia Canario-Americana (1992). Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria, pp. 195–215.
López de Ayala, P. (1780 [1393]). Cronicas de los Reyes de Castilla, Don Pedro, Don Enrique II, Don Juan I, Don Enrique III. Zurita, G., and Llaguno Amirola, E. (eds.). Don Antonio de Sancha’s Printing House, Madrid.
Le Canarien (B) and (G) (2003 [1488–1491] and [c. 1419]). Le Canarien: Manuscritos, Transcripción y Traducción. Pico, B., Aznar, E., and Corbella, D. (trans.), Instituto de Estudios Canarios, La Laguna, Santa Cruz de Tenerife.
Lobo Cabrera, M. (2016). La población esclava de Las Palmas en la primera mitad del siglo XVI. In Acosta Guerrero, E. (ed.), Actas XXI Coloquio de Historia Canario-Americana (2014). Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria, pp. 1–15.
Lobo Cabrera, M. (2019). Los Guanches y el ganado menor tras la conquista. In Chávez-Álvarez, M. E., Camalich Massieu, M. D., and Marín Socas, D. (eds.), Un Periplo Docente e Investigador: Estudios en Homenaje al Profesor Antonio Tejera Gaspar. Universidad de La Laguna, San Cristóbal de La Laguna, pp. 319–334.
López, M., Foronda, P., Feliu, C., and Hernández, M. (2013). Genetic characterization of black rat (Rattus rattus) of the Canary Islands: origin and colonization. Biological Invasions 15(11): 2367–2372.
Luxán, S. d., Viña Brito, A., Hernández, M., and Ronquillo Rubio, M. (2009). La Empresa Azucarera en Canarias: Siglos XV-XX. Destilerías Arehucas, Ayuntamiento de Los Llanos de Aridane, Las Palmas de Gran Canaria.
Lyman, R. L. (1994). Vertebrate Taphonomy. Cambridge University Press, Cambridge.
Lyman, R. L. (2008). Quantitative Paleozoology. Cambridge University Press, Cambridge.
Martín, A., Suárez-Pérez, A., Rodríguez, E., and Afonso, D. (2013). Sobre la antigua presencia de palomas endémicas en la isla de Gran Canaria (islas Canarias). Revista de la Academia Canaria de Ciencias 25:135–140.
Martínez García, L. (1995). La alimentación en el Hospital del Rey de Burgos, contribución a la historia del consumo en la Baja Edad Media. Cuadernos Burgaleses de Historia Medieval 3: 83–154.
McTavish, E. J., Decker, J. E., Schnabel, R. D., Taylor, J. F., and Hillis, D. M. (2013). New World cattle show ancestry from multiple independent domestication events. Proceedings of the National Academy of Sciences 110(15): E1398-E1406.
Meco Cabrera, J. (1992). Los Ovicaprinos Paleocanarios de Villaverde: Diseño Paleontológico y Marco Paleoambiental. Gobierno de Canarias, Dirección General de Patrimonio Histórico, Santa Cruz de Tenerife.
Mendoza Medina, F., Suárez Medina, I., and Moreno-Benítez, M. A. (2012). Trabajos de Restauración, Consolidación y Conservación en las Obras del Solar de la Calle San Marcial, Esquina Herrería. Tibicena Arqueología y Patrimonio SL., Servicio de Patrimonio Histórico del Cabildo insular de Gran Canaria, Las Palmas de Gran Canaria.
Mesa Hernández, E. (2016). Los Guanches y el Marisqueo: Aprovechamiento de los Recursos Malacofáunicos en la Prehistoria de Tenerife. Doctoral dissertation, Universidad de la Laguna, Tenerife.
Mitchell, P. J. (2023). Archaeological research in the Canary Islands: Island archaeology off Africa’s atlantic coast. Journal of Archaeological Research, https://doi.org/10.1007/s10814-023-09186-y.
Morales, J., Vidal-Matutano, P., Marrero-Salas, E., Henríquez-Valido, P., Lacave-Hernández, A., García-Ávila, J. C., Abreu-Hernandez, I., and Arnay-de-la-Rosa, M. (2021). High-mountain plant use and management: macro-botanical data from the pre-Hispanic sites of Chasogo and Cruz de Tea, 13-17th centuries AD, Tenerife (Canary Islands, Spain). Journal of Archaeological Science: Reports 35: 102730.
Morales, J., Rodríguez-Rodríguez, A., and Henríquez-Valido, P. (2017). Agricultura y recolección vegetal en la arqueología prehispánica de las Islas Canarias (siglos III-XV d.C.): la contribución de los estudios carpológicos. In Fernández Eraso, J., Mujika Alustiza, J. A., Arrizabalaga Valbuena, Á., and García Diez, M. (eds.), Miscelanea en Homenaje a Lydia Zapara Peña (1965–2015). Universidad del País Vasco, Bilbao, pp. 189–218.
Morales, J., Speciale, C., Rodríguez-Rodríguez, A., Henríquez-Valido, P., Marrero-Salas, E., Hernández-Marrero, J. C., López, R., Delgado-Darias, T., Hagenblad, J., Fregel, R., and Santana, J. (2023). Agriculture and crop dispersal in the western periphery of the Old World: the Amazigh/Berber settling of the Canary Islands (ca. 2nd-15th centuries CE). Vegetation History and Archaeobotany. https://doi.org/10.1007/s00334-023-00920-6.
Navarrete, V., Urquiza, S. V., Quiroga, L., and Puente, V. (2022). Introducción de animales domésticos exóticos en la economía pastoril andina (ca. 440–500 AP). Un caso de estudio de Antofagasta de la Sierra, Catamarca, Puna Austral de Argentina. Estudios Atacameños 68: e5816.
Morris, J. (2004). Explorations in anatomy: the remains from Royal London Hospital. Anthropozoologica 49(1): 109–120.
Nogales, M., Rodríguez-Luengo, J. L., and Marrero, P. (2006). Ecological effects and distribution of invasive non-native mammals on the Canary Islands. Mammal Review 36(1):49–65.
Núñez-Lahuerta, C., Moreno-Azanza, M., Pérez-Pueyo, M., Del-Arco-Aguilar, M. del C., Del-Arco-Aguilar, M., Siverio-Batista, C., Castillo-Ruiz, C., and Cruzado-Caballero, P. (2023). Shearwater eggs in Lobos 3, a holocene site of Fuerteventura (Canary Islands). Diversity 15(2). https://doi.org/10.3390/d15020144.
O’Connor, T. (2000). The Archaeology of Animal Bones. 2nd ed. Texas A & M University Press, College Station.
O’Connor, T. (2003). The Analysis of Urban Animal Bone Assemblages. Council for British Archaeology, York.
Olalde, I., Capote, J., Del-Arco, M.C., Atoche Peña, P., Delgado-Darias, T., González-Anton, R., País Pais, J., Amills, M., Lalueza-Fox, C., and Ramírez, O. (2015). Ancient DNA sheds light on the ancestry of pre-hispanic Canarian pigs. Genetics Selection Evolution 47(1). https://doi.org/10.1186/s12711-015-0115-7.
Onrubia Pintado, J. (2003). La Isla de los ”Guanartemes”: Territorio, Sociedad y Poder en la Gran Canaria Indígena:(Siglos XIV-XV). Cabildo de Gran Canaria., Las Palmas de Gran Canaria.
Onrubia Pintado, J. and González Marrero, M. d. C. (2016). The Archaeology of the early castilian colonialism in Atlantic Africa. The Canary Islands and Western Barbary (1478–1526). In Cruz Berrocal, M., Montón-Subías, S. and Ruiz Martínez, A. (eds.), Archaeologies of Early Modern Spanish Colonialism. Springer, Cham, pp. 119–151.
Orton, D. C. (2010). A new tool for zooarchaeological analysis: ArcGIS skeletal templates for some common mammalian species. Internet Archaeology 28. https://doi.org/10.11141/ia.28.4.
Pais Pais, J. (1996). La Economía de Producción en la Prehistoria de la Isla de La Palma. La Ganadería. Dirección General de Patrimonio Histórico, Gobierno de Canarias, Santa Cruz de Tenerife.
Pigière, F., Boone, I., Udrescu, M., Van Neer, W., and Vanpoucke, S. (2004). Status as reflected in food refuse of late medieval noble and urban households at Namur (Belgium). In O’Day, S. J., Van Neer, W., and Ervynck, A. (eds.), Behaviour Behind Bones: The Zooarchaeology of Ritual, Religion, Status and Identity 9th Conference of the International Council of Archaeozoology. Oxbow, Oxford, pp. 233–243.
Pliny The Elder, G. (1629 [77]). Historia Natural. Huerta, G. de (trans.), Iuan Gonçalez’s printing house, Madrid.
Prummel, W. (1987). Atlas for the identification of foetal skeletal elements of cattle, horse, sheep and pig. Archaeozoologia 12: 11–41.
Prummel, W. and Frisch, H.-J. (1986). A guide for the distinction of species, sex and body side in bones of sheep and goat. Journal of Archaeological Science 13(6): 567–577.
Quintana Andrés, P. C. (2003). Grupos urbanos y asistencia social: el Hospital de San Martín en Las Palmas en el seiscientos. Vegueta: Anuario de la Facultad de Geografía e Historia 7: 41–64.
Ramis, D. (2018). Animal exploitation in the early prehistory of the Balearic Islands. Journal of Island and Coastal Archaeology 13(2): 269–282.
Rando Reyes, J., Alcover, J. A., Michaux, J., Hutterer, R., and Navarro Mederos, J. F. (2012). Late-Holocene asynchronous extinction of endemic mammals on the eastern Canary Islands. Holocene 22(7): 801–808.
Rando Reyes, J. C., López, M., and Jiménez, M. C. (1997). Bird remains from the archaeological site of Guinea (El Hierro, Canary Islands). International Journal of Osteoarchaeology 7(4): 298–302.
Rando Reyes, J. C., Rodríguez-Rodríguez, A., Pais Pais, F. J., Navarro Mederos, J. F., and Martín, E. (1996). Los restos de aves del yacimiento arqueológico de «El Tendal» (La Palma, Islas Canarias). El Museo Canario 51: 87–102.
Recco, N. da (1830 [1342]). Della Canaria e Delle Altre Isole Oltre Ispania nell’Oceano Nuovamente Ritrovate. Boccaccio, G. and Sebastiano, C. (trans.), Paolo Andrea Molina’s Printing House, Milán.
Reimer, P. J., Austin, W. E., Bard, E., Bayliss, A., Blackwell, P. G., Ramsey, C. B., Butzin, M., Cheng, H., Edwards, L. E., Friedrich, M., Grootes, P. M., Guilderson, T. P., Hajdas, I., Heaton, T., Hogg, A. G., Hughen, K. A., Kromer, B., Manning, S. W., Muscheler, R., Palmer, J. G., Pearson, C., Plicht, J. van der, Reimer, R. W., Richards, D. A., Scott, E. M., Southon, J. R., Turney C. S. M., Wacker, L., Adolphi, F., Büntgen, U., Capano, M., Fahrni, S. M., Fogtmann-Schulz, A., Friedrich, R., Köhler, P., Kudsk, S., Miyake, F., Olsen, J., Reinig, F., Sakamoto, M., Sookdeo, A., and Talamo, S. (2020). The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62(4): 725–757.
Reitz, E. J. (1986). Vertebrate fauna from Locus 39, Puerto Real, Haiti. Journal of Field Archaeology 13(2): 317–328.
Reitz, E. J. (1992). The Spanish colonial experience and domestic animals. Historical Archaeology 26(1): 84–91.
Reitz, E. J. and Waselkov, G. A. (2015). Vertebrate use at early colonies on the southeastern coasts of Eastern North America. International Journal of Historical Archaeology 19(1): 21–45.
Riquelme, J. A. (2020). Análisis faunístico de los restos óseos de mamíferos recuperados en el puerto histórico de Palos de la Frontera (Huelva). In Campos Carrasco, J. M. (ed.), La Recuperación Geoarqueológica del Puerto Histórico de Palos de la Frontera (SS. XIV-XVI): Medio Natural e Instalaciones Portuarias. Universidad de Huelva, Huelva, pp. 217–235.
Rivero Suárez, B. (1991). El Azúcar en Tenerife, 1496–1550. Instituto de Estudios Canario, La Laguna.
Rodríguez-Rodríguez, A., Santana, J., Castellano Alonso, P., del Pino Curbelo, M., Francisco Ortega, I., Gómez de la Rúa, D., González Ruiz, M. d. C., Henríquez-Valido, P., Machado Yanes, M. d. C., Marlasca, R., Mesa Hernández, E., Morales, J., Moreno García, M., Rando Reyes, J. C., and Hernández Calvento, L. (2021). Un lugar entre las dunas: aprovechamiento oportunista de un espacio costero durante la etapa preeuropea de la isla de Gran Canaria (circa siglos VIII-XI AD). Trabajos de Prehistoria 78(2): 325–343.
Rodríguez Santana, C. G. (1994). La Pesca Entre los Canarios, Guanches y Auaritas: Las Ictiofaunas Arqueológicas del Archipiélago Canario. Doctoral dissertation, Universidad de La Laguna, Tenerife.
Rodríguez-Rodríguez, A. (1997). La tecnología de la piel y el cuero en la prehistoria de Canarias: una aproximación etnorqueológica. El Museo Canario 52: 11–31.
Ronquillo Rubio, M. (2008). Ingenios azucareros en la colonización canaria: 1487–1526: localización espacial y organización del espacio en Gran Canaria. In Viña Brito, A., Gambín García, M., and Chinea, C. (eds.), Azúcar: Los Ingenios en la Colonización Canaria (1487–1525). Museo de Historia y Antropología de Tenerife, Santa Cruz de Tenerife, pp. 99–112.
Ronquillo Rubio, M. and Aznar Vallejo, E. (1998). Repartimientos de Gran Canaria. Transcripción, Estudio y Notas. Cabildo insular de Gran Canaria, Las Palmas de Gran Canaria.
Ruíz Melgarejo (1974 [1531]). Ordenanzas del Concejo de Gran Canaria (1531). Morales Padrón, F (ed.), Ediciones del Excmo. Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria.
Santana, J., Fregel, R., Lightfoot, E., Morales, J., Alamón-Núñez, M., Guillen, J., Moreno-Benítez, M. A., and Rodríguez-Rodríguez, A. (2016). The early colonial Atlantic world: new insights on the African diaspora from isotopic and ancient DNA analyses of a multiethnic 15th-17th century burial population from the Canary Islands, Spain. American Journal of Physical Anthropology 159(2): 300–312.
Santana Pérez, G., Salas Pascual, M., and Cáceres Lorenzo, M. T. (2004). Historia de la incorporación de cultivos africanos en Canarias durante los siglos XV al XVIII. Revista de Historia Canaria 20: 219–234.
Santana-Pérez, J. M. (2005). Excluidos y Recluidos en el Antiguo Régimen. Anroart, Las Palmas de Gran Canaria.
Santana-Pérez, J. M. (2018). The African Atlantic islands in maritime history during the Ancien Régime. International Journal of Maritime History 30(4): 634–648.
Schmid, E. (1972). Atlas of Animal Bones. Elsevier, Amsterdam.
Seetah, K. (2018). Humans, Animals, and the Craft of Slaughter in Archaeo-Historic Societies. Cambridge University Press, Cambridge.
Silver, I. A. (1969). The ageing of domestic animals. In Brothwell, D. and Higg, E. (eds.), Science in Archaeology: A Survey of Progress and Research. Thames and Hudson, London, pp. 283–302.
Siverio-Batista, C. (2017). Estudio de las arqueofaunas terrestres en talleres de púrpura del Atlántico y Mediterráneo centro-occidental durante la Antigüedad. Arqueología y Territorio 14: 193–204.
Sosa, F. J. de (1994 [1678–88]). Topografía de la Isla Afortunada de Gran Canaria. Ronquillo Rubio, M., and Viña Brito, A. (eds.), Ediciones del Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria.
Sosa Suárez, E. (2012). Informe de las Cerámicas de Importación: Yacimiento: “San Marcial” Vegueta, 2011. Tibicena Arqueología y Patrimonio SL., Las Palmas de Gran Canaria.
Soulier, M.-C. and Costamagno, S. (2017). Let the cutmarks speak! experimental butchery to reconstruct carcass processing. Journal of Archaeological Science: Reports 11: 782–802.
Speller, C. F., Burley, D. V., Woodward, R. P., and Yang, D. Y. (2013). Ancient mtDNA analysis of early 16th century Caribbean cattle provides insight into founding populations of New World creole cattle breeds. PLoS One 8(7): e69584.
Suárez, N. M., Betancor, E., Fregel, R., and Pestano, J. (2013). Genetic characterization, at the mitochondrial and nuclear DNA levels, of five Canary Island dog breeds. Animal Genetics 44(4): 432–441.
Tabares, L. and Santana Rodríguez, L. (2018). Testamentos de Canarios, Gomeros y Herreños [1506–1550]. Instituto de Estudios Canarios, San Cristóbal de La Laguna, Tenerife.
Tejera Gaspar, A. (2020). Colón en las Islas Canarias: 1492–1502: La Gomera y Gran Canaria. LeCanarien Ediciones, Santa Cruz de Tenerife.
Tejera Gaspar, A. and Aznar Vallejo, E. (1991). El primer contacto entre europeos y canarios: ¿1312?-1477. In Morales Padrón, F. (ed.), Actas VIII Coloquio de Historia Canario-Americana (1988). Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria, pp. 17–37.
Torres Krupij, A., Morales Pérez, A. L., and Capote, J. (2018). The historical background of chickens in Canary Islands. In Torres, A. and Capote, J. (eds.), Traditional Breed of Canarian Hen. Instituto Canario de Investigaciones Agrarias. Gobierno de Canarias, San Cristóbal de La Laguna, Tenerife, pp. 9–21.
Torriani, L. (1592). Descrittione et Historia del Regno de Isole Canarie, già dette le Fortunate, con il Parere delle Loro Fortificationi. Self-published, Coimbra.
Tous Meliá, J. (1995). Las Palmas de Gran Canaria a través de la Cartografía (1588–1899). Casa de Colón y Museo Militar Regional de Canarias, Las Palmas de Gran Canaria.
Traveset, A., Nogales, M., Alcover, J. A., Delgado, J. D., López-Darias, M., Godoy, D., Igual, J. M., and Bover, P. (2009). A review on the effects of alien rodents in the Balearic (Western Mediterranean Sea) and Canary Islands (Eastern Atlantic Ocean). Biological Invasions 11: 1653–1670.
Valera Gomes, M., Gomes, R. V., and Cardoso, J. L. (1996). Aspectos do quotidiano numa casa de Silves, durante o século XV. XELB: Revista de Arqueologia, Arte, Etnologia e História 3: 33–78.
Velasco-Vázquez, J., Alberto-Barroso, V., and Quintana Andrés, P. C. (2003). La Mala Muerte: El Depósito Funerario de la Calle Rosarito de La Isleta. Cabildo de Gran Canaria, Las Palmas de Gran Canaria.
Velasco-Vázquez, J., Alberto-Barroso, V., Delgado-Darias, T., and Moreno-Benítez, M. (2021). A propósito del poblamiento aborigen en Gran Canaria. Demografía, dinámica social y ocupación del territorio. Complutum 32(1): 167–189.
Villagrasa-Elías, R. (2022). El Renacimiento Hospitalario en la Península Ibérica: Una Propuesta Desde la Cultura Escrita (1374–1549). Doctoral dissertation, Universidad de Zaragoza, Zaragoza.
Viña Brito, A. (2006). Canarias en el comercio atlántico de esclavos. In Viña Brito, A., Hernández González, M., and Alfaro Hardisson, E. (eds.), Esclavos. Gobierno de Canarias, San Cristóbal de La Laguna, pp. 15–25.
Viña Brito, A. (2012). Los flamencos en Canarias en el siglo XVI. Revista de Historia Canaria 194: 161–191.
Viña Brito, A. (2014). Azúcar y Mecenazgo en Gran Canaria: El Oro de Las Islas, Siglos XV-XVI. Cabildo insular de Gran Canaria, Las Palmas de Gran Canaria.
Wakild, E. (2021). Learning from the llama: on the broad contours of cultural contributions and geographic expansion. História, Ciências, Saúde-Manguinhos 28: 141–159.
Wall, S. M. (1980). The animal bones from the excavation of the Hospital of St Mary of Ospringe. Archaeologia Cantiana 96: 227–266.
Wanjala, G., Bagi, Z., and Kusza, S. (2021). Meta-Analysis of mitochondrial DNA control region diversity to shed light on phylogenetic relationship and demographic history of african sheep (Ovis aries) breeds. Biology 10(8): 762.
Wheeler, J. C., Russel, A. J. F., and Redden, H. (1995). Llamas and alpacas: pre-conquest breeds and post-conquest hybrids. Journal of Archaeological Science 22: 833–840.
Yvinec, J. H., Coutureau, M., and Carpentier, C. (2007). Corpus of digitalized mammals skeletons. ArchéoZoothèque. https://www.archeozoo.org/archeozootheque/.
Zeder, M. A. (2006). Reconciling rates of long bone fusion and tooth eruption and wear in sheep (Ovis) and goat (Capra). In Ruscillo, D. (ed.), Recent Advances in Ageing and Sexing Animal Bones. Oxbow, Oxford, pp. 87–118.
Zurara, G. E. (1978 [1452–1453]). Crónica dos Feitos Notáveis que se Passaram na Conquista de Guiné por Mandato do Infante D. Henrique. Sosa Soares, T. (ed.), Academia Portuguesa da Historia, Lisboa.
Acknowledgments
The authors would like to acknowledge Tibicena S. L. and El Museo Canario for providing access to fieldwork reports and materials. The authors also thank María del Carmen Cruz de Mercadal, Marco Moreno, David Naranjo, Manuel Lobo, María del Cristo González, Juan Manuel Santana, Jacob Morales, Sergio Hernández, Raúl Villagrasa-Elías, Simon-Pierre Gilson, E. M. Fidalgo, Saro Méndez, Pedro Henríquez, Aaron Morquecho, and the Seminars' members of the ULPGC for comments on an earlier version of this manuscript. We extend our sincere gratitude to the referees and the editor for their thoughtful suggestions aimed at enhancing the quality of this contribution.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This research was supported by the RTI2018-101923-J-I00, PID2020-117496GB-I00 and RYC2019-028346 projects (Ministerio de Ciencia e Innovación, Spain) and the ERC Starting Grant project IsoCAN (grant 851733, European Commission). The first author was supported by a PhD scholarship of the Agencia Canaria de Investigación, Innovación y Sociedad de la Información (ACIIS) (European Social Fund and the Consejería de Economía, Industria, Comercio y Conocimiento of the Government of the Canary Islands).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing Interests
The authors declare that there is no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brito-Mayor, A., Santana, J., Moreno-García, M. et al. Animal Consumption at Hospital de San Martín (Gran Canaria): First Zooarchaeological Analysis in the Modern Era of the Canary Islands (Fifteenth-Eighteenth Centuries CE). Int J Histor Archaeol 27, 1210–1242 (2023). https://doi.org/10.1007/s10761-023-00708-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10761-023-00708-4