
Abstract
This study aims at reconstructing foodways in the north-east (NE) of the Iberian Peninsula, focusing on lipid residue analysis of utilitarian vessels and using as case studies the sites of Puig Castellar of Biosca (180–120 BCE) and Guissona (120 BCE-third century CE). In total, fifty vessel fragments of different types and origins were analysed with techniques such as gas chromatography-mass spectrometry (GC-MS) and gas chromatography-combustion-isotope ratio mass spectrometry (GC-C-IRMS). As some vessel fragments were recovered from waterlogged contexts, they had excellent lipid preservation, which enabled the detection of various compounds that are rarely reported in archaeological contexts in Iberia. Analyses revealed both animal and plant products in the vessels, suggested that a variety of food resources was consumed, and that vessels had multiple uses. The detection of levoglucosan in some extracts, along with other heat markers, further suggested the presence of cellulose or starchy products in contact with fire as well as the heating of animal fats. Combined with available bioarchaeological and palaeoenvironmental studies from both sites, the analyses indicated a continuity in diet across the time periods during which the sites were occupied. Some differences were also observed, including the possible use of tubers until the first century CE. The results are then contextualised and compared with other available organic residue studies from the Roman Iberian Peninsula. The study demonstrates how the combination of multiple bioarchaeological proxies and biomolecular approaches can provide a holistic means to approach Roman foodways.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Iberian Peninsula was one of the first areas to be conquered by the Romans outside the Italian region (Morillo and Aurrecoechea-Fernández 2006), bringing significant changes to all aspects of life, including foodways. The latter do not merely reflect the availability of ingredients and subsistence practices, but all activities involving food, from production and preparation to modes of consumption and discard. Importantly, they can reveal social relations, identities, economic strategies and more (e.g., Livarda 2013, 2018; Hastorf 2017; Twiss 2019). Significant advances to understand the food culture during the Roman period in the Iberian Peninsula have been made through several bioarchaeological studies, which have contributed important information on the types of plants (e.g., Peña Chocarro et al. 2019) and animals (e.g. Colominas et al. 2019) that were consumed and their management.
This study seeks to add to Roman food studies using a different approach for the area, offering the first study of organic residue analysis on Roman ceramics from Catalonia. Beyond Catalonia, residue analyses of Roman pottery in the Iberian Peninsula are limited with most of them centring on the detection of wine, olive oil or fish-based products in Roman amphorae (e.g., Blanco-Zubiaguirre et al. 2019; Dorrego et al. 2004; Cau Ontiveros et al. 2008; Pecci et al. 2021; Morais et al. 2016). Other studies have focused on the examination of single vessels (Cosano et al. 2023; Guerrero and Sánchez Vizcaíno 1995) while specific types of amphorae have been examined for type of content, specifically the African (Woodworth et al. 2015) and the Lusitanian amphorae (Morais et al. 2016), including material from the Iberian Peninsula. Only two studies examine various types of pottery with the aim to trace their contents and determine dietary and cooking habits. In particular, Pecci and Cau Ontiveros (2014) studied samples of cooking pots and different types of amphorae from the Sa Mesquida site in Mallorca (Balearic Islands), dated to the Late Roman period (fifth-seventh century CE). Their results indicated the presence of ruminant and vegetable fats and possibly fish in cooking pots, as well as vegetable oil, grains, and degraded wine or other substances in the amphorae. Parras et al. 2020 analysed 16 vessels from the site of Cerro de Las Albahacas (Santo Tomé, Jaén), a Roman camp built after the battle of Baécula (208 BCE), providing new insights into both the soldiers’ consumption practices and the productive capacity of the Iberian population that presumably supplied the vessels and their contents during the Second Punic War. The chemical markers found indicated the presence of beeswax, plant oils, such as those from nuts (almonds, walnuts), and fleshy fruits (olive) as well as animal fats, while GC-C-IRMS analysis of a single vessel indicated non-ruminant animal fats, most probably from pigs (Parras et al. 2020). Our current investigation focuses on the sites of Puig Castellar of Biosca and Guissona, which cover periods dating to the second century BCE through to the third century CE, allowing a substantial temporal resolution of food practices over a chronological timespan not covered before in the Iberian Peninsula through organic residue analysis (ORA).
Study sites
Puig Castellar (DMS: WGS84:148,514.590784, 5,134,997.013133) was a Roman Republican military fortress dating to between 180 and 120 BCE and was probably used to control the region and supply logistical equipment to the troops in the mainland (Rodrigo et al. 2022, 2014). It was built on top of a hill known as Puig Castellar de Biosca or Les Guixeres de Talteüll, surrounded by three river streams. The location itself allows visual control of the whole area, providing a strategic advantage in controlling the roads leading into the interior of the country. Additionally, the site was naturally protected, as access from the northern slope is impossible (Rodrigo Requena et al. 2022; Pera Isern et al. 2019). Puig Castellar (Fig. 1) was surrounded by a defensive wall which included square towers and bastions and occupied an area of approximately 1.6 hectares (Rodrigo et al. 2022; Rodrigo Requena et al. 2022). Attached to the interior of the southern and western sides of the wall, a series of barracks were documented, which may have served as living, storage and working quarters for the troops. In this same area, evidence of an open space used for metalworking was also detected. At the small plain located at the top of the hill, a large headquarters building was identified, showing clear Italian construction techniques, decorations, and measurements (Rodrigo et al. 2022; Romaní Sala et al. 2020). According to the excavators, this building is probably one of the first military headquarters (principia) of the Roman Republican army in this province (Rodrigo Requena et al. 2022). Bioarchaeological analyses of palaeoenvironmental remains at Puig Castellar are limited, however, the few animal bones recovered during the excavations indicated the presence of cattle (Bos taurus), suiformes, sheep/goats (Ovis aries/ Capra hircus), red deer (Cervus Elaphus) and horse (Equus caballus) (Colominas 2022 unpublished report).

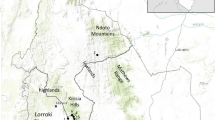
Plan of the site of Puig Castellar of Biosca (photo made by: Iñaki Matias, UDG-ICAC)
Soon after the abandonment of Puig Castellar, the ancient town of Iesso (under the modern town of Guissona, DMS: WGS84:143,595.548912, 5,129,219.920341) was founded about 6 km away, at the centre of the Guissona Plain (Rodrigo et al. 2022; Fig. 2). The abundant groundwater resources seem to have been precisely one of the main reasons why the new town was established in this area: the presence of a high-water table, which extends about 2000 km2 around Guissona, seems to have been crucial to ensure constant water availability (Guitart Duran 2007). The site was surrounded by a wall about 2.5–3 m wide and occupied a total area of 15–18 hectares. It was inhabited from the end of the 2nd c. BCE until at least the 7th c. CE, undergoing several transformations through time (Guitart et al. 2004).

The position of Puig Castellar of Biosca and Guissona in the map of Iberian Peninsula and the distance between them (photo made by: Institut Cartogràfic i Geològic de Catalunya (ICGC), licensed under CC BY 4.0)
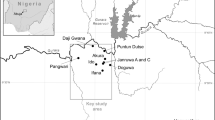
The Roman structures excavated at Guissona include five streets in a grid layout, a Republican residential neighbourhood progressively transformed into a large domus during the 1st and 2nd c. CE, a public bath and, outside the wall, an inn that provided meals, drink and lodging for travellers (Romaní Sala et al. 2022; Fig. 3). In addition, a total of five water wells, built between the 1st c. BCE and the 2nd c. CE, were identified (see Table 1).

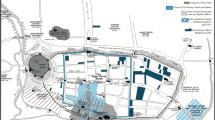
Aerial photograph of the site of Guissona (photo made by: Guissona Research Team)
Zooarchaeological material was studied from only two wells (wells 1 and 4, see Table 1). The remains of cattle (Bos taurus), dog (Canis familiaris), goat/sheep, pig (Sus domesticus) and rabbit (Oryctolagus cuniculus) were identified in well 1, located to a residential area (Padrós 2007). The next well studied was associated with the inn and included remains of dogs, oxen, horse (Equus caballus), sheep (Ovis aries), goats (Capra hircus), pigs, rabbits, small rodents, small birds, turtles, snails and amphibians (Larumbe 2020, 2022, 2023 unpublished report; see S1 Appendix). Archaeobotanical studies were conducted for the wells of the residential area (wells 1 and 2, see Table 1) revealing the presence of mainly fruits and nuts, with fig (Ficus carica), grape (Vitis vinifera) and walnut (Juglans regia) being the most abundant remains (Buxó et al. 2004, p. 229;242; Baniou et al. 2024; for a full list see S1 Appendix).
A palynological study from well 4 associated with the inn showed the presence of mainly Chenopodiaceae, Poaceae and Quercus coccifera, suggesting that the landscape was probably semi-open and dry, although, the presence of taxa such as Betula and Alnus indicated relative proximity to more humid areas (Currás 2018 unpublished report). Some indirect evidence for the existence of cultivated areas were also present. Vitis sp. pollen was also identified, which could be from both domesticated and wild varieties. The presence of coprolitic fungi was indicative of livestock activity in the area (Currás 2018 unpublished report; for a full list see S1 Appendix).
Finally, wood and charcoal remains from wells 1 and 2 of the residential area (Table 1) indicated the dominance of Pinus sylvestris/nigra and Quercus sp. caducifoli in the assemblage, while well 4 of the inn area had mainly Artemisia sp. and Sambucus sp. (Buxó et al. 2004 for well 1 and 2; López-Bultó 2020 unpublished report for well 4).
Methodology
Sample selection and lipid extraction
A total of 50 vessel fragments (n = 30 Guissona and n = 15 Puig Castellar of Biosca) were selected for lipid residue analysis. The chronological period ranges from the second century BCE to the third century CE. One potsherd had a visible charred residue (G12-48IR) that was encrusted onto the inner surface of the sherd, which was analysed separately. Most of the samples (n = 37) selected were fragments of cooking vessels, while the rest were vessels for processing (n = 6), serving or consumption (n = 6) and storage (n = 1) (Table 2). Potsherds from different locations on the vessel profile (e.g., base, body, rim) were selected. Both locally or regionally-produced and imported potsherds were analysed (North Africa; n = 13; Italy; n = 6). Twenty-six samples, originating from various contexts, had charring marks on their external surface indicating contact with fire. Of the selected samples, 6 fragments were from two of the wells found at Guissona (wells 4 and 5, associated with the Inn and the public baths respectively), preserved under waterlogged conditions. The rest of the fragments derived from various contexts from both sites. For detailed contextual information for each fragment, see S2 Appendix. Sediment control samples from the sites were not available as the pottery analysed was excavated in the past between 2012–2021, when ORA studies were not anticipated for the site. However, contamination from the surrounding environment is expected to be minimal as it has previously been observed that migration of lipids from the sedimentary matrix into pottery is highly restricted due to the hydrophobic properties of lipids that limit their transfer from the soil to the clay wall of the pottery (and vice versa) (Heron et al. 1991).
In addition, to minimise contamination, all surfaces of the selected potsherd fragments were removed with a modelling drill (cleaned prior with dichloromethane/methanol; 2:1 v/v) to remove exogenous lipids that may have originated from the storage or handling of the potsherds. Then the fragments were crushed to obtain a fine powder (2–3 g) in a mortar and pestle. Lipids were extracted using established protocols; 6 potsherds from waterlogged contexts from Guissona were extracted using the conventional solvent extraction protocol (Evershed et al. 1990), while for the rest (n = 44) the acidified methanol extraction protocol was used (Craig et al. 2013; Correa-Ascencio and Evershed 2014). Several other measures were adopted to avoid contamination. All solvents used were GC grade and the reagents were analytical grade (typically > 98% of purity). All glassware and the mortars and pestles used for crushing pottery fragments were sterilised in a muffle furnace at 450˚C for 8 h. For each lipid extraction batch, analytical blanks were prepared in order to detect potential contamination from solvents or reagents. Clean, nitrile gloves were worn while handling the potsherds and while preparing the extracts. For full details of the extraction protocols, see S3 Appendix.
Due to financial restrictions, all samples could not be analysed via every instrumental method. Thus, the following approach was adopted to ensure maximisation of the obtained results: all samples (n = 50) were analysed via Gas Chromatography-Flame Ionisation-Detector (GC-FID); then, samples with high lipid concentrations and/or with complex chromatographic profiles were further analysed via Gas Chromatography-Mass Spectrometry (GC–MS) (n = 31). The obtained GC–MS data were then used to identify compounds in the extracts, and compounds such as fatty acids were matched in other non-analysed extracts based on identical retention time. In addition, some of the samples that were not analysed via GC–MS (such as G12-4, G13-10, G14-11, G13-14, G13-15, G16-25, G16-30, G12-48) were analysed via Gas Chromatography-Combustion-Isotope Ratio Mass Spectrometry (GC-C-IRMS) to identify fatty acids and obtain their δ13C values to discriminate the source of the fats.
Instrumental analysis: GC-FID, GC–MS and GC-C-IRMS
GC-FID analysis was performed on 50 samples using an Agilent Technologies 7820A GC series with a Flame Ionisation Detector System with an Automatic Liquid Sampler and equipped with a 15 m × 0.25 mm × 0.15 µm DB-5 MS capillary column (J&W Scientific, CA, USA) and 5 m of guard column. Sample injection was performed in splitless mode. The oven temperature programme started at 50ºC (held for 2 min) then increased to 320ºC at a rate of 6ºC min−1 and held for 10 min. Injector and detector temperatures were 310ºC and 320ºC respectively. Hydrogen was used as the carrier gas at constant flow of 2 mL min−1. For full details, see S3 Appendix.
GC–MS analyses were performed on thirty-one samples (31/50 = 62%, see S2 Appendix) as they demonstrated high lipid content and complex chromatographic profiles in the GC-FID profiles. An Agilent Technologies 7890A GC System coupled to an Agilent 5975C MS and an Automatic Liquid Sampler was used, operated in electron impacted ionization mode (EI, 70 eV), equipped with a 30 m × 0.25 mm × 0.25 µm DB-5MS capillary column (J&W Scientific, CA, USA) and a 5 m guard column. Sample injection was performed in splitless mode. The oven temperature programme started at 50ºC (held for 2 min) then increased to 320ºC at a rate of 6ºC min−1 and held for 10 min. Injector and detector temperatures were 310ºC and 320ºC respectively. Helium was used as the carrier gas at constant flow of 1.5 mL min−1. Injector, transfer line, and ion source temperatures were 310ºC, 320ºC and 250ºC, respectively. The GC–MS was set to operate in a mixed mode. That means that a combination of acquisition in selective ion monitoring (SIM) and scan mode was programmed. The SIM mode was included to enhance the analytical sensitivity for ω − (o-alkylphenyl) alkanoic acids (APAAs) by targeting the ions with m/z 105 (as base peak), 262, 290, 318 and 346. Full details are provided in S3 Appendix. Compound identification was performed by comparing MS spectral data with the NIST database combined with the relative retention time.
The selection of samples analysed via GC-C-IRMS was made with the following criteria: samples from both archaeological sites and with different chronologies were analysed. An attempt was also made to analyse different types of vessels (cooking pot, mortar, bowl, patina, shallow casserole) of both imported and local origin in order to assess any differences in vessel type, dating or origin. Full details about the GC-C-IRMS analysis are provided in S3 Appendix.
Results
Lipid preservation
The range of lipid concentration in the vessels studied was wide and all samples except two (G21-22; G1-46) had interpretable lipid concentrations (above 5 µg g−1) (Evershed et al. 1999; Evershed 2008; see Table 2). None of the extracts had lipid profiles typical of lipids from sediments (i.e., high concentrations of n-alkanes). The average lipid concentration of vessels from Guissona was 84.4 µg g−1 (solvent extracts; n = 6) and 730.3 µg g−1 (acid extracts; n = 29), while the average lipid concentration of vessels from Puig Castellar was 105.1 µg g−1 (acid extracts, n = 15) (see Table 3). No relationship between lipid concentration and the type or the origin of vessel was observed, with patinae and cooking pots retaining both the highest and lowest amounts of lipids (Table 4; see S2 Appendix).
A range of compounds was detected in the samples, including saturated fatty acids ranging from C8:0 to C30:0 (up to C34:0 in samples G16-28 and G12-48), branched-chain fatty acids, unsaturated fatty acids, n-alcohols, α,ω-dicarboxylic acids, mid-chain ketones and sterols. Levoglucosan was detected in 3 extracts from waterlogged contexts. The different types of compounds detected and the types of organic substances identified are detailed below.
Animal fats
Overall, the extracts from Guissona (n = 35) are dominated by a wide range of fatty acids (with relatively low abundance of long-chain fatty acids (C20:0-C30:0)). Extracts from Puig Castellar (n = 15) demonstrate a narrower range of fatty acids (C14:0-C24:0). Fatty acids C16:0 and C18:0 had the highest relative abundance in extracts from both sites. Branched-chain fatty acids (C14:0br, C15:0br, C16:0br, C17:0br, C18:0br, C19:0br) were identified in almost all samples (32/35 91.4%- from Guissona and 14/15–93.3%- from Puig Castellar). These compounds have bacterial origin and might be indicative of ruminant fats (Christie 1981; Dudd and Evershed 1998). Thirteen samples from Guissona and 7 from Puig Castellar contained cholesterol derivatives (Fig. 4a, d). Sixteen samples from Guissona and 7 from Puig Castellar contained acyl glycerols (Fig. 4a-d). The above compounds (cholesterol derivatives and acyl glycerols) suggest the presence of degraded animal fats (Dudd and Evershed 1999; Whelton et al. 2018).

Chromatograms of extracts from Guissona (a-c) and Puig Castellar of Biosca (d-e). a) is a solvent extract and indicates the presence of plant oils, degraded animal fats, heat markers (mid-chain ketones and levoglucosan), dehydroabietic and 7-oxo-dehydroabietic acid; b-e) are acidified methanol extracts and indicate the presence of: b) degraded animal fats and plant products, c) degraded animal fats (heated), plant products and dehydroabietic and 7-oxo-dehydroabietic acid, d) degraded animal fats, heat markers, plant products and dehydroabietic and 7-oxo-dehydroabietic acid and e) plant oils and degraded animal fats. Key: Circles = fatty acids; Hexagon = unsaturated fatty acids; Stars = Acyl glycerols; Diamonds = DHA and 7- oxo-DHA; Squares = ketones; Triangles = cholesterol derivatives; Arrow = Diacids, * = plasticizer contamination; LV = Levoglucosan; OH = Alcohol; I.S. = Internal Standard)
The determination of the stable isotopic values (δ13C) of the major fatty acids (C16:0 and C18:0) in eighteen samples (13 from Guissona; 5 from Puig Castellar) enabled characterisation of the degraded animal fats. The δ13C16:0 values obtained (ranging from -32‰ to -25‰) suggest that the animals from which the fats were derived were grazing exclusively on C3 vegetation (Copley et al. 2003; Fig. 5). A scatterplot of the δ13C16:0 and δ13C18:0 values reveals that none of the samples fall within confidence ellipses for modern references for ruminant carcass fats or porcine fats, but rather in-between, reflecting the mixture of these products in vessels (Fig. 5). Similarly, Δ13C values (δ13C18:0-δ13C16:0), suggested that 6 samples (33.3%) fall within the range of ruminant adipose fats, while 12 samples (66.7%) fall within ranges for plant oils and in-between ranges of non-ruminant origin and ruminant adipose fats, reflecting mixtures (Fig. 5). Interestingly, despite the presence of short-chain fatty acids (C8:0-C10:0) in the lipid extracts, which have been reported in ruminant dairy fats (Dudd and Evershed 1998; Evershed et al. 1992), none of the obtained δ13C values fell within established ranges for ruminant dairy products.

Graphs showing A: δ13C18:0 versus δ13C16:0 values and B: Δ13C values (δ13C18:0-δ13C16:0) versus δ13C16:0 values from the vessels from Guissona (orange circles) and Puig Castellar (purple circles). In A, the confidence ellipses relate to modern fats of animals raised on a purely C3 diet in Britain (Copley et al. 2003). In B, ranges represent the mean ± 1 s.d. of the Δ13C values for a global database comprising modern reference animal fats from various regions including the UK, regions in Africa, central and south-west Asia, as well as plant oils (Dudd and Evershed 1998; Spangenberg et al. 2006; Gregg et al. 2009; Steele et al. 2010; Dunne et al. 2012)
Plant products
There is a variable amount of evidence pointing to the presence of plant products in most of the samples from both sites. Unsaturated fatty acids (C14:1, C16:1, C18:1, C18:2, C20:1 and C22:1; Fig. 4a-e) were identified in thirty-three samples from Guissona and 8 from Puig Castellar. The presence of oleic acid (C18:1) in higher or equal abundance to stearic acid (C18:0) in 4 extracts (G17-16, G11-47 PCB14-12, and PCB22-5; see S2 Appendix; Fig. 4a,e) is an indication of the presence of plant oils in these vessels (Evershed et al. 1990; Rageot et al. 2019). Additionally, the presence of erucic acid (C22:1) in samples G13-9, G14-12, G1-33 and PCB14-9 may be indicative of plants belonging to the Brassicaceae family (Colombini and Modugno 2009).
Hydroxy acids and a large range of α,ω-dicarboxylic acids (diacids), from C8 to C20, were found in thirteen extracts from Guissona and 5 extracts from Puig Castellar (Fig. 4b-e). Short-chain diacids are formed as degradation products of unsaturated fatty acids (Regert et al. 1998), which are present in higher quantities in plant products (Colombini and Modugno 2009). However, diacids with longer-chain lengths from C16 to C26 are diagnostic of suberin, which is found in the outer part of underground plants, such as tubers (Kolattukudy 1980, 1981; Dunne 2022) and may suggest processing of these products in samples G16-28, G1-34, G04-52, PCB16-19 and PCB20-26 (Fig. 4b).
Finally, plant sterols (such as stigmasterol, β-sitosterol and campsterol) were identified in sixteen samples (10/35 from Guissona, 28.6% and 6/15, 40% from Puig Castellar), which are undoubtedly of plant origin (Colombini and Modugno 2009; Briggs et al. 2023; Fig. 4c-d).
Diterpenoid biomarkers (Fig. 4a, c-d) can derive from the deliberate use of conifer resins (Regert 2004; Colombini et al. 2005), the firing process through the use of wood (Reber et al. 2018), reheating in an oxidising atmosphere (Whelton et al. 2021), or even bacteria (Costa et al. 2016). Some of the samples (22/50, 44%) from both sites contain diterpenoids, such as dehydroabietic acid and 7-oxodehydroabietic acid. In this context they might suggest the heating of coniferous resins in an oxygenated environment, the use of products that may have been in contact with resins, or a bacterial source (Whelton et al. 2021). Finally, the presence of levoglucosan in samples G17-16; G17-19 and G21-23 from waterlogged contexts suggests the presence of cellulose or starchy products that have been in contact with fire (see also below) (Shoda et al. 2018; Heron et al. 2016; Simoneit et al. 1999; Fig. 4a).
Heating markers and other compounds
Mid-chain ketones (C29; C31; C33; C35) were identified in twelve extracts (7 cooking pots, 1 cooking pot or casserole, 1 mortar, 2 shallow casseroles and 1 patina) from Guissona and 4 extracts (3 cooking pots, 1 mortar) from Puig Castellar. The heating of animal fats above 300 °C causes the decarboxylation and condensation of fatty acids and the formation of mid-chain ketones dominated by the C31, C33 and C35 homologues (Fig. 4a, c-d) (Evershed et al. 1995; Raven et al. 1997). This suggests that animal fats were heated in these vessels. Additionally, the homologue C29 is derived from the leaf waxes of Brassica sp. (Dudd and Evershed 1999; Evershed et al. 1995) and was found in 4 extracts from Guissona (Fig. 4a, c).
It is interesting to note that 9 of these samples show evidence of charring on the surface of the vessel. Twelve extracts (cooking pots, shallow casseroles, patinae, bowl) were found to contain polyaromatic hydrocarbons (PAHs) which are considered to be smoke indicators (Simoneit et al. 2000; Reber et al. 2018), possibly suggesting exposure to campfire smoke. However, the formation of PAHs can also be caused by environmental factors. In particular, it can be caused by endogenous synthesis of micro-organisms or highly developed plants that have come into contact with high temperatures (Suess 1976).
Three waterlogged samples contained levoglucosan, which has been identified as a pyrolysis product of cellulose or starch and might be consistent with the processing of starchy materials under heat in the vessels (Shoda et al. 2018; Simoneit et al. 1999; Caseiro and Oliveira 2012). Additionally, the presence of dehydroabietic acid and 7-oxodehydroabietic acid in seventeen samples (mostly in cooking pots, but also in a shallow casserole, bowl, mortar and storage container) indicate the presence of coniferous resins in contact with fire that could suggest the use of pine wood as fuel (Simoneit et al. 2000). However, as the abundance of these compounds in the extracts is quite low, it is possible that instead of indicating the direct use of wood or coniferous products in vessels, it rather shows the use of products that had previously been stored in resin-lined vessels (e.g., Cramp et al. 2011).
Discussion
Overall, organic residue analysis of a range of vessels from the two studied sites point to a variable diet from the second century BCE to the third century CE that combined different alimentary sources. Analysis of the relationship between the contents of vessels and their type, origin and form did not reveal clear patterns as similar products were found in all of them (Supplementary file 4).
The results show a mixture of ingredients in each vessel, which is indicative either of 1) their use multiple times, 2) the use of different products each time, and/or 3) the combination of products for the creation of meals.
At Puig Castellar, all available information on food resources is based on a limited number of animals remains that were found at the site. No flotation had been carried out until recently and the few samples available are currently under study. The results of the organic residue analysis suggest that a mixture of plant and animal products formed the diet of the people inhabiting the fortress. Concerning animal products, the fatty acid-specific stable isotopic results suggest that non-ruminant animals and ruminant meat was consumed, which is consistent with the dominance of ovicaprid remains followed by suiformes at the site (Colominas 2022 unpublished report). The zooarchaeological analysis indicated that the age of the animals was linked to the usual age at which animals were slaughtered for meat consumption. At the same time, the anatomical parts of animals at the site suggest that meat was brought into rather than prepared on-site (Colominas 2022 unpublished report). The supply of meat in Roman military camps was usually from the local area or from further away when in processed form (e.g., salted, dried, smoked) (Thomas and Stallibrass 2008). According to zooarchaeological studies, pig was the prevalent meat in military sites of the Roman world (King 1999), although local variation has also been argued for (e.g. MacKinnon 2019).
Regarding plant products consumed at Puig Castellar it is not possible to add much detail on the species due to the lack of completed archaeobotanical studies. However, as erucic acid was detected (in an imported Italian mortar), it is possible that species of the Brassicaceae family were used and consumed. The presence of mid-chain ketones, charring marks on vessels, products derived from heating (levoglucosan, PAHs) and other heat markers suggest that it is likely that meat was cooked, and that 4 vessels (3 cooking pots and 1 mortar) came into contact with high temperatures, shedding light on food preparation practices.
More detail can be achieved for the reconstruction of the foodways at Roman Guissona. Overall, the diet of the inhabitants of Guissona does not seem to change dramatically from the first century BCE to the third century CE. Faunal remains and lipid residue analysis point to the consumption of animal meat. Regarding the macrofauna remains from well 1 (1st c. CE to second half of the 2nd c. CE), ovine/caprine, cattle, pigs and rabbits have been identified with some of the bones showing colour changes in their surface due to contact with fire, as well as cutting marks (Padrós 2007). In well 4 (mid-1st c. CE- end of 2nd c. CE/ beginning of 3rd c. CE) some animals were found whole (dogs and horse) with no signs of processing, while others in bone portions (cattle, ovine/caprine). Some of the bones showed traces of thermal changes and intentional fractures linked to food consumption (Larumbe 2023 unpublished report). The presence of red deer (Cervus elaphus) is confirmed by the presence of 2 bones, one of which had probably been turned into an object of utility purpose (Larumbe 2023 unpublished report).
Based on the GC-C-IRMS results, the inhabitants of Roman Guissona consumed both ruminant and non-ruminant animals, which is also corroborated by the zooarchaeological analysis. Four of the vessels extracts from well 4 have mid-chain ketones, indicating that animals were cooked in these vessels (cooking pots, patina, shallow casserole). Given that only ruminant animals with traces of consumption were found in this well, it can be speculated that ruminant animals were processed in these vessels (note, however, that GC-C-IRMS results are not available for these samples). In contrast, both ruminant and non-ruminant animals with cut-marks were identified in well 1, fitting well with the lipid residue results from vessels from a similar chronological period. The consumption of cooked meat is also attested by the presence of ketones in twelve samples of different types of vessels (cooking pot, shallow casserole, patina and mortar) of various origin and chronology, while the main animals cooked and consumed could be cattle, goat, sheep, pig and rabbit, based on the animal bones that have been found at the site.
Diet in Roman Guissona also included legumes, cereals, fruits and nuts, according to archaeobotanical studies from two of the wells, and may have been supplemented by the consumption of vegetable oils, other plants, as well as starchy products, as suggested by the lipid residue results. In terms of cereal consumption, few barley (Hordeum vulgare), free-threshing wheat (Triticum aestivum/durum) and emmer (Triticum dicoccum) grains were identified (Buxó et al. 2004). The presence of legumes was more limited, with only common vetch (Vicia sativa) and pea (Pisum sativum) present. Many of the findings were fruits, mainly grapes and figs. At the same time, nuts were also identified, including walnuts (Juglans regia) and hazelnuts (Coryllus avellana) (Buxó et al. 2004). The presence of these species was not identified through organic residue analysis and could have been consumed through other culinary pathways (such as raw, or through flour-based preparations).
Evidence for plant oils were detected in four extracts from Guissona and Puig Castellar. The macrobotanical results suggest the presence of olive stones, the derivatives of which may have been used in the cooking process, although, specific evidence for olive oil was not found. The presence of erucic acid was attested in 4 other extracts, possibly suggesting the use of oils from the Brassicaceae family, which was also reported in palynological analyses (Currás 2018 unpublished report). The diet of the inhabitants was also possibly supplemented by tubers, as indicated in 5 extracts (3 from Guissona and 2 from Puig Castellar) containing long-chain diacids (C16-C20). Interestingly, the extracts with long-chain diacids are from cooking pots and a dish/flat bowl dating to between the second century BCE (PCB16-19, PCB20-26) and the first century BCE-beginning 1st c. CE (G16-28, G1-34, G04-52) but none of the later samples contain evidence of underground plant organs. Whether this is an artefact of sampling or a real indication of a change in dietary habits after the first century CE can be only assessed with larger-scale study of more vessels and of plant macro- and micro-remains.
Plant products were found in all the vessel types under consideration (cooking pots, shallow casserole, deep casserole, patina, mortars and dish/flat bowl) at both Guissona and Puig Castellar of Biosca, except for a single storage container which had low lipid concentration. Moreover, the presence of plant products was observed in vessels of both local/regional and imported origin from all study periods. Therefore, it seems that regardless of the type or origin of the vessel, the consumption of plant species in combination with animal fats was a standard cooking practice. The same appears to apply to the use of plant oils, as they have been identified in vessels of local origin or imported from Africa; in serving platters or patinae, shallow casseroles, cooking pots and dishes/flat bowls.
The case of fish at both sites is rather intriguing as a search for relevant ions for biomarkers of aquatic products using SIM mode on the GC–MS in the extracts (4,8,12-trimethyltridecanoic acid (TMTD): m/z 87, 213, 270; pristanic acid: 88, 101, 312; phytanic acid: m/z 101, 171, 326); ω − (o-alkylphenyl) alkanoic acids (APAAs): m/z 262, 290, 318 and 346, and the fragment ion of the base peak m/z 105) did not reveal any evidence for their presence. This is consistent with the absence of fish bones from at least all the -relatively few- studied contexts, although amphorae associated with the storage of salted fish and fish sauces were found at Guissona. Future injection of samples onto a longer column (e.g., 60 m × 0.32 mm) (e.g., Cramp and Evershed 2014; Lucquin et al. 2016) may further elucidate the role of fish in diet. The consumption of fish, especially fresh fish, in the Roman period was related to geographical location, period and social status (Marzano 2019). Fresh and salted fish were more spatially limited while sauces, such as garum, were more widespread socially and spatially (Bekker-Nielsen 2005). The absence of fish-related compounds may therefore be due to the geography of the sites away from the sea.
In a similar vein, dairy products were not detected in any of the samples from either site. In general, the Romans included dairy products in their diet; mainly cheese, as milk needed a special preservation method (MacKinnon 2019, 151). Dairy products could have also been part of the diet of the Roman troops (Davies 1971, 127–128). Zooarchaeological studies carried out on material from the North-East of the Iberian Peninsula, indicate that from the 1st to the 3rd century AC, sheep, goats, and cattle were not bred for their meat but mainly for their secondary products (Colominas et al. 2019; Colominas 2013, 136). In fact, it has been suggested that from the first century AC onwards, cattle served mainly as draught animals, goats for their milk, sheep for wool and pigs for their meat (Colominas 2013, 137). The zooarchaeological analysis carried out at Puig Castellar of Biosca did not produce any evidence for livestock breeding and instead it was identified as a consumption site (Colominas 2022 unpublished report). This may mean that cheese/milk production was not carried out at the site and thus dairy consumption may have been more limited. As far as Guissona is concerned, the study of zooarchaeological remains is so far limited to only a few finds from the wells. No extensive zooarchaeological study has been carried out in the different contexts of the site, and animal husbandry practices have not yet been explored. Furthermore, the absence of dairy products does not necessarily mean that such products were not consumed as their absence may be due to the specific vessels studied not having been used for their processing or consumption. Yet, as a variety of vessel types were studied, it is possible that dairy products were not a ‘staple’ of cooking practices but were rather consumed in more restricted or specific settings. Overall, the absence of dairy products could be the result of sampling, the limited presence of dairy products at the sites, or a combination of both.
This study further provides new insights into the cooking methods of the period. Τhe analysed material indicated that different kind of vessels (cooking pots, platters and mortars) were heated to temperatures above 300 °C, a practice seemingly common at both sites. The identification of the diterpenes typical of coniferous resin in some vessels, may be an indication for its use as fuel in cooking fires. These taxa were present in the area, as shown by the wood, anthracological and palynological analyses. In Guissona, the most common species among the wood remains were Pinus sylvestris-nigra type and Quercus sp. deciduous. Although both species were used among others to produce artefacts, they are also among the few species found in charred form (Piqué et al. 2016). Anthracological studies of the Roman city of Lleida (first century BCE-fourth century CE) as well as of other Roman and Late Antiquity sites of Spain also indicated its use as firewood, among other taxa (Piqué 2009).
Studies have shown that Roman mortars were used to mix and grind different kind of products such as plant, meat and dairy products (e.g., Cramp et al. 2011). Of the six mortars included in our study, in all of them traces of degraded animal fats were found, three of which had fatty acid isotopic results that suggest the presence of mixtures of ruminant and non-ruminant fats. In sample G14-8 ketones, hydroxy acid and dehydroabietic acid were identified, suggesting the presence of heated coniferous resins and the heating of animal fats in the vessels. However, the presence of traces of resin may originate from the use of products in the mortar that were transferred from resin-lined vessels (Cramp et al. 2011). It cannot be explained with absolute certainty why ketones are found in mortars. However, it has been suggested that mortars could have been placed directly on the fire during the preparation of specific recipes or used as a lid on cooking pots (Cramp 2008, 39).
Direct comparisons of the results of this study with those of other sites of the Iberian Peninsula are not straightforward as they do not align chronologically. However, the Roman military camp of Cerro de Las Albahacas (Santo Tomé, Jaén, Spain) dating to the late third century BCE (Parras et al. 2020) is relatively close chronologically to Puig Castellar (2nd c. BCE). At both sites non-ruminant animal fats were found, together with plant oils and probably plant waxes, with Puig Castellar providing additional evidence for the consumption of tubers as well as ruminant carcass fats.
From the first century BCE to the third century CE, our results from Guissona, point to a variety in cooking and consumption practices, including ruminant and non-ruminant animal fats, plant products, plant oils, and probably starchy products. While evidence from the fourth century CE is lacking, the site of Sa Mesquida (Mallorca, Balearic Islands) dating to the fifth-seventh century CE has evidence of the consumption of ruminant and non-ruminant animals, vegetal products and oils, with indications of potential fish consumption in the late Roman period (Pecci and Cau Ontiveros 2014). Based on the data from the examined sites, it appears that the diet of individuals in the military camps was relatively more limited, while in the residential sites was more variable, although this may be an artefact of sampling.
Conclusion
This study provides the first study of organic residue analysis from Roman ceramics in Catalonia. It is also one of the few studies in the Iberian Peninsula that deals with the analysis of different types of vessels at a settlement level and the first one for the period second century BCE-third century CE, opening up new windows to a better understanding of diet and cooking methods in the Roman past. According to the results, the diet during these centuries in the north-east of the Iberian Peninsula was characterised by a variety of animal and vegetable products and their potential combinations. The combination of plant and animal products in the preparation of meals was seemingly a common cooking practice as shown both by the results of the organic residues and by bibliographical sources. Fish and dairy products seem absent or limited based on current evidence at both sites. The potential consumption of tubers could be traced in vessels that date up to the first century CE. At both sites, evidence of the heating of vessels and the direct or indirect use of coniferous wood products was attested.
At Puig Castellar it appears that all cooking pots, dishes, and mortars, regardless of the context where they were collected, were used to process both animal fats and plant products, with a possible preference for non-ruminant animal fats as shown by their dominance in the GC-C-IRMS results. In Guissona, mixtures of products in vessels can be explained by the fact that the inhabitants combined meat and vegetable products in their diet. Combining the various archaeological analyses and the results of the organic analysis, it becomes clear that the consumption of ruminant and non-ruminant meat was a common practice. At the same time, the diet was supplemented by plant products in cooked form (based on residue analysis), but also in raw and/or flour-based form (based on archaeobotanical analysis). Finally, continuity in the eating habits seems to be delineated throughout the time span occupied within each site.
Although there is still a long way to go to fully understand Roman dietary habits, this study, through the analysis of organic residues and its combination with other proxies, has provided new insights into Roman dietary habits and cooking technologies, focusing its attention to a poorly studied area of the Iberian Peninsula in regard to organic residue analysis.
Data Availability
All the authors confirm that all data analysed during this study are included within the manuscript and the supplementary information files.
References
Baniou Th, Livarda A, Romaní Sala N, Rodrigo Requena E (2024) The diet in the Roman period through the study of sites with waterlogged assemblages: the case study of Guissona (Catalonia, Spain). In: Chatzikonstantinou Y (ed) Archaeozooms: Aspects and potentials of modern archaeological research. Propylaeum, pp 234–255. https://doi.org/10.11588/propylaeum.1319.c19013
Bekker-Nielsen T (2005) Ancient fishing and fish processing in the Black sea region. Aarhus University Press, Denmark
Blanco-Zubiaguirre L, Olivares M, Castro K, Carrero JA, García-Benito C, García-Serrano JA, Pérez-Pérez J, Pérez-Arantegui J (2019) Wine markers in archeological potteries: detection by GC-MS at ultratrace levels. Anal Bioanal Chem 411:6711–6722. https://doi.org/10.1007/s00216-019-02044-1
Briggs L, Cooper J, Craig OE, Heron C, Lucquin A, Martínez Milantchi MM, Samson A (2023) Molecular evidence for new foodways in the early colonial Caribbean: organic residue analysis at Isla de Mona, Puerto Rico. Archaeol Anthropol Sci 15(70):1–19. https://doi.org/10.1007/s12520-023-01771-y
Buxó R, Canal D, Guitart J, Pera J, Piqué R (2004) Excavació de dos pous d'època romana a Guissona: l’explotació dels recursos vegetals a la ciutat de Iesso als segles I a. C. - II d. C. In: Guitart Duran J, Pera Isern J (eds) Iesso I: miscel·lània arqueològica. Patronat d’Arqueologia de Guissona, pp 213–277
Caseiro A, Oliveira C (2012) Variations in wood burning organic marker concentrations in the atmospheres of four European cities. J Environ Monit 14(8):2261–2269. https://doi.org/10.1039/C2EM10849F
Cau Ontiveros MA, Martínez Farreras V, Pecci A, Mas Florit C, Fantuzzi L (2008) Archaeometric analysis for provenance and content of Roman amphorae from the site of Sa Mesquida (Mallorca, Spain). Mediter Archaeol Archaeom 18(2):87–105. https://doi.org/10.5281/zenodo.1297151
Christie WW (1981) Lipid metabolism in ruminant animals. Pergamon Press, Oxford
Colombini MP, Modugno F (2009) Organic mass spectrometry in art and archaeology. Wiley, West Sussex
Colombini MP, Modugno F, Ribechini E (2005) Direct exposure electron ionization mass spectrometry and gas chromatography/mass spectrometry techniques to study organic coatings on archaeological amphorae. J Mass Spectrom 40:675–687. https://doi.org/10.1002/jms.841
Colominas L (2013) Arqueología y Romanización. In: Producción, distribución y consumo de animales en el nordeste de la Península Ibérica entre los siglos V ane-V dne, vol. 2480. BAR International Series
Colominas L (2022) Estudi arqueozoològic de les restes de fauna recuperades al castellum de Biosca (Lleida). Campanyes 2014–2016. Unpublished report, Insitut Català d’Arqueologia Clàssica
Colominas L, Antolín F, Ferrer M, Castanyer P, Tremoleda J (2019) From Vilauba to Vila Alba: changes and continuities in animal and crop husbandry practices from the Early Roman to the beginning of the Middle Ages in the north-east of the Iberian Peninsula. Quat Int 499 Part A:67–79. https://doi.org/10.1016/j.quaint.2017.12.034
Copley MS, Berstan R, Dudd SN, Docherty G, Mukherjee AJ, Sraker V, Payne S, Evershed RP (2003) Direct chemical evidence for widespread dairying in prehistoric Britain. Proc Natl Acad Sci 100(4):1524–1529. https://doi.org/10.1073/pnas.0335955100
Correa-Ascencio M, Evershed RP (2014) High throughput screening of organic residues in archaeological potsherds using direct acidified methanol extraction. Anal Methods 6:1330–1340. https://doi.org/10.1039/C3AY41678J
Cosano D, Manuel Román J, Lafont F, Ruiz Arrebola JF (2023) Archaeometric identification of a perfume from Roman times. Heritage 6:4472–4491. https://doi.org/10.3390/heritage6060236
Costa MS, Rego A, Ramos V, Afonso TB, Freitas S, Preto M, Lopes V, Vasconcelos V, Magalhães C, Leão PN (2016) The conifer biomarkers dehydroabietic and abietic acids are widespread in Cyanobacteria. Sci Rep 6(23436):1–11. https://doi.org/10.1038/srep23436
Craig OE, Saul H, Lucquin A, Nishida Y, Taché K, Clarke L, Thompson A, Altoft DT, Uchiyama J, Ajimoto M, Gibbs K, Isaksson S, Heron C, Jordan P (2013) Earliest evidence for the use of pottery. Nature 496:351–354. https://doi.org/10.1038/nature12109
Cramp LJE, Evershed RP (2014) Reconstructing aquatic resource exploitation in human prehistory using lipid biomarkers and stable isotopes. In: Holland HD, Turekian KK (eds) Treatise on geochemistry: archaeology and anthropology, vol 12, 2nd edn. Elsevier, Amsterdam, pp 319–339
Cramp LJE, Evershed RP, Eckardt H (2011) What was a mortarium used for? Organic residues and cultural change in Iron Age and Roman Britain. Antiquity 85(330):1339–1352
Cramp LJE (2008) Foodways and Identity: organic residue analysis of Roman mortaria and other pottery. Dissertation, University of Reading
Currás A (2018) Informe dels resultats de l’anàlisi pol·línica del pou al jaciment del Parc Arqueològic de Guissona. Unpublished report. Instituto de las Ciencias del Patrimonio, CSIC
Davies RW (1971) The Roman military diet. Britannia 2:122–142 http://www.jstor.org/stable/525803
Dorrego F, Carrera F, Luxán MP (2004) Investigations on Roman amphorae sealing system. Mater Struct 37:369–374. https://doi.org/10.1007/BF02481686
Dudd S, Evershed RP (1998) Direct demonstration of milk as an element of archaeological economies. Science 282(5393):1478–1481. https://doi.org/10.1126/science.282.5393.1478
Dudd S, Evershed RP (1999) Evidence for varying patterns of exploitation of animal products in different prehistoric pottery traditions based on lipids preserved in surface and absorbed residues. J Archaeol Sci 26:1473–1482. https://doi.org/10.1006/jasc.1998.0434
Dunne J (2022) Gone to seed? Early pottery and plant processing in Holocene north Africa. Quatern Int 608–609(20):178–193. https://doi.org/10.1016/j.quaint.2021.02.004
Dunne J, Evershed RP, Salque M, Cramp L, Bruni S, Ryan K, Biagetti S, Lernia S (2012) First dairying in green Saharan Africa in the fifth millennium BC. Nature 486(7403):390–394. https://doi.org/10.1038/nature11186
Evershed RP (2008) Organic residue analysis in archaeology: the archaeological biomarker revolution. Archaeometry 50:895–924. https://doi.org/10.1111/j.1475-4754.2008.00446.x
Evershed RP, Heron C, Goad LJ (1990) Analysis of organic residues of archaeological origin by high-temperature gas chromatography and gas chromatography-mass spectrometry. Analyst 115(10):1339–1342. https://doi.org/10.1039/AN9901501339
Evershed RP, Heron C, Charters S, Goad LJ (1992) The survival of food residues: new methods of analysis, Interpretation and Application. Proc Br Acad 77:187–208
Evershed RP, Stott AW, Raven A, Dudd SN, Charters S, Leyden A (1995) Formation of long-chain ketones in ancient pottery vessels by pyrolysis of acyl lipids. Tetrahedron Lett 36:8875–8878. https://doi.org/10.1016/0040-4039(95)01844-8
Evershed RP, Dudd SN, Charters S, Mottram H, Stott AW, Raven A (1999) Lipids as carriers of anthropogenic signals from prehistory. Philos Trans R Soc Lond Ser B-Biol Sci 354:19–31. https://doi.org/10.1098/rstb.1999.0357
Gregg MW, Banning EB, Gibbs K, Slater GF (2009) Subsistence practices and pottery use in Neolithic Jordan: molecular and isotopic evidence. J Archaeol Sci 36(4):937–946. https://doi.org/10.1016/j.jas.2008.09.009
Guerrero ML, Sánchez Vizcaíno A (1995) Análisis de indicadores bioquímicos del contenido de recipientes arqueológicos. Complutum 6:281–291
Guitart Duran J (2007) L’utilisation de l’eau en Catalogne romaine: le cas de Iesso (Guissona). In: Jean-Pierre B, Jean-Luc F (eds) Énergie hydraulique et machines élévatrices d’eau dans l’Antiquité. Publications du Centre Jean Bérard, Naples, pp 33–49
Guitart J, Pera J, Ros J (2004) Arqueologia a l’antiga ciutat romana de Iesso (Guissona, Lleida). In: Primer Simposi Patrimoni i Turisme Cultural (ed) Arqueologia viva de les ciutats de l’antiguitat. Lleida, 4th-6th of October 2001, pp. 174–180
Hastorf CA (2017) The social archaeology of food: thinking about eating from prehistory to the present. Cambridge University Press
Heron C, Evershed RP, Goad LJ (1991) Effects of migration of soil lipids on organic residues associated with buried potsherds. J Archaeol Sci 18:641–659. https://doi.org/10.1016/0305-4403(91)90027-M
Heron C, Shoda S, Breu Barcons A, Czebreszuk J, Eley Y, Gorton M, Kirleis W, Kneisel J, Lucquin A, Müller J, Nishida Y, Son J, Craig OE (2016) First molecular and isotopic evidence of millet processing in prehistoric pottery vessels. Sci Rep 6(38767):1–9. https://doi.org/10.1038/srep38767
King A (1999) Diet in the Roman world: a regional inter-site comparison of the mammal bones. J Roman World 12:168–202. https://doi.org/10.1017/S1047759400017979
Kolattukudy PE (1980) Biopolyester membranes of plants: cutin and suberin. Science 208(4447):990–1000. https://doi.org/10.1126/science.208.4447.990
Kolattukudy PE (1981) Structure, biosynthesis, and biodegradation of cutin and suberin. Annu Rev Plant Physiol 32:539–567. https://doi.org/10.1146/annurev.pp.32.060181.002543
Larumbe M (2020) Variabilitat dels conjunts faunístics recuperats al pou de l’edifici extramurs del jaciment de Iesso (mitjans segle I - inicis segle III dC): reconstitució de la dinàmica de rebliment i accions implicades en la deposició de les restes de fauna. BA Thesis, Autonomous University of Barcelona
Larumbe M (2022) Ús social i processos de treball del conjunt faunístic recuperat al pou de l’edifici extramurs del jaciment de Iesso. M.Sc. Dissertation, Autonomous University of Barcelona
Larumbe M (2023) Estudi arqueozoològic de les restes de microfauna recuperades al pou de l’edifici extramurs del jaciment de Iesso (Guissona). Unpublished report, Laboratori d’Arqueozoologia, Autonomous University of Barcelona
Livarda A (2013) Date, rituals and socio-cultural identity in the northwestern Roman provinces. Oxf J Archaeol 32(1):101–117. https://doi.org/10.1111/ojoa.12004
Livarda A (2018) Tastes in the Roman provinces: an archaeobotanical approach to socio-cultural change. In: Rudolph KC (ed) Taste and the ancient senses. Routledge, New York, pp 179–196
López-Bultó O (2020) Informe de l’anàlisi dendrològic i morfològic de les fustes del pou de Iesso (Guisona). Unpublished report. Laboratori d’arqueobotànica. Autonomous University of Barcelona
Lucquin A, Gibbs K, Uchiyama J, Saul H, Ajimoto M, Eley Y, Radini A, Heron CP, Shoda S, Nishida Y, Lundy J, Jordan P, Isaksson S, Craig OE (2016) Ancient lipids document continuity in the use of early hunter-gatherer pottery through 9,000 years of Japanese prehistory. Proc Natl Acad Sci USA 113(15):3991–3996. https://doi.org/10.1073/pnas.1522908113
MacKinnon M (2019) Meat and other animal products. In: Erdkamp P, Holleran C (eds) The Routledge handbook of diet and nutrition in the Roman world. Routledge, New York, pp 150–162
Marzano A (2019) Fish and seafood. In: Erdkamp P, Holleran C (eds) The Routledge handbook of diet and nutrition in the roman world. Routledge, New York, pp 163–173
Morais R, Oliveira C, Araújo A (2016) Lusitanian amphorae of the Augustan Era and their contents: organic residue analysis. In: Vaz Pinto I, Roberto de Almeida R and Martin A (eds) Lusitanian amphorae: production and distribution. Roman and late antique Mediterranean pottery 10. Archaeopress Series, pp 105–109
Morillo Α, Aurrecoechea-Fernández J (2006) The Roman army in Hispania. An archeological guide. University of León, León
Padrós N (2007) Estudi arqueozoològic del pou 1 de la ciutat romana de Iesso: Guissona, Segarra. M.Sc. Dissertation, Autonomous University of Barcelona
Parras DJ, Sánchez A, Luque Á, Lechuga MA, Rueda C (2020) The diet of Romans during 2nd Punic War in their Camp Cerro De Las Albahacas (Santo Tomé, Spain) from Organic Residue Analysis in ceramic vessels by GC-MS, HPLC-APCI-MS and GC-C-IRMS methods. Mediter Archaeol Archaeom 20(1):127–143. https://doi.org/10.5281/zenodo.3674091
Pecci A, Cau Ontiveros MA (2014) Residue analysis of late Roman cooking pots and amphorae from SA Mesquida (Mallorca, Balearic Islands). In: Poulou-Papadimitriou N, Nodarou E, Kilikoglou V (eds) LRCW 4 Late Roman Coarse Wares, Cooking Wares and Amphorae in the Mediterranean Archaeology and archaeometry The Mediterranean: a market without frontiers, Archaopress - BAR International Series 2616 (II), pp. 833–841.
Pecci A, Reynolds P, Mileto S, Vargas Giron JM, Bernal-Casasola D (2021) Production and transport of goods in the Roman period: Residue analysis and wine derivatives in Late Republican Baetican ovoid amphorae. Environ Archaeol 1–13 https://doi.org/10.1080/14614103.2020.1867291
Peña Chocarro L, Pérez-Jordà G, Alonso N, Antolín F, Teira-Brión A, Tereso JP, Montes Moya EM, López Reyes D (2019) Roman and medieval crops in the Iberian Peninsula: a first overview of seeds and fruits from archaeological sites. Quatern Int 499:49–66. https://doi.org/10.1016/j.quaint.2017.09.037
Pera Isern J, Rodrigo Requena E, Romaní Sala N, Carreras Monfort C (2019) Puig Castellar de Biosca (Lleida). Una fortificación romana del siglo II a. C. en el noreste de la Hispania Citerior. Gladius 39:19–43. https://doi.org/10.3989/gladius.2019.02
Piqué R (2009) Wood in the ancient Mediterranean: forests, fuelwood and raw materials. Stud Rural World Roman Period 4:61–72
Piqué R, Ravotto A, López-Bultó O (2016) Roman wells of north-eastern Iberian Peninsula: Landscape and use of wooden resources. Quat Int 404 Part A:104–113. https://doi.org/10.1016/j.quaint.2015.08.011
Rageot M, Mötsch A, Schorer B, Bardel D, Winkler A, Sacchetti F (2019) New insights into Early Celtic consumption practices: organic residue analyses of local and imported pottery from Vix-Mont Lassois. PLoS ONE 14(6):1–19. https://doi.org/10.1371/journal.pone.0218001
Raven AM, van Bergen PF, Stott AW, Dudd SN, Evershed RP (1997) Formation of long-chain ketones in archaeological pottery vessels by pyrolysis of acyl lipids. J Anal Appl Pyrol 40–41:267–285. https://doi.org/10.1016/S0165-2370(97)00036-3
Reber EA, Kerr MTHL, Whelton MT, Evershed RP (2018) Lipid residues from low-fired pottery. Archaeometry 61:131–144. https://doi.org/10.1111/arcm.12403
Regert M (2004) Investigating the history of prehistoric glues by gas chromatography-mass spectrometry. J Sep Sci 27(3):244–254. https://doi.org/10.1002/jssc.200301608
Regert M, Bland HA, Dudd SN, Bergen PFV, Evershed RP (1998) Free and bound fatty acid oxidation products in archaeological ceramic vessels. Proc R Soc 265:2027–2032. https://doi.org/10.1098/rspb.1998.0536
Rodrigo E, Romaní N, Carreras C, Pera J, Catarineu L (2022) The Roman Republican castellum of Puig Castellar (Biosca): the earliest evidence for military headquarters in Hispania Citerior (180–120 BCE). J Roman Archaeol 35(2):818–847. https://doi.org/10.1017/S104775942200037X
Rodrigo E, Carreras C, Pera J, Guitart J (2014) La presencia romana en el NE de la Provincia Citerior durante el siglo II a.C. Aproximación arqueológica a partir de los yacimientos de Can Tacó (Montmeló, Barcelona) y Puig Castellar (Biosca, Lleida). Anejos Archivo Español de Arqueología, LXX:191–209
Rodrigo Requena E, Romaní Sala N, Carreras Monfort C, Pera Isern J, Catarineu Iglesias L (2022) Darreres novetats al castellum de Puig Castellar: l’organització i funcions dels espais de la tropa. Treballs D’arqueologia 25:175–203. https://doi.org/10.5565/rev/tda.144
Romaní Sala N, Rodrigo Requena E, Pera Isern J, Gutiérrez García-Moreno A (2020) El material constructiu de la fortificació militar de segle II ac de Puig Castellar de Biosca (Lleida). Paviments, revestiments parietals i cobertes. In Aquilué J, Beltrán de Heredia J, Caixal A, Fierro X, Kirchner H (eds) Estudis sobre ceràmica i arqueologia de l’arquitectura. Homenatge al Dr. Alberto López Mullor. Barcelona: Diputació de Barcelona, pp 391–403
Romaní Sala N, Rodrigo Requena E, Pera Isern J, Guitart Duran J (2022) La ocupación periurbana de la zona norte de la ciudad romana de Iesso. Novedades Arqueológicas. In Mateos Cruz P, Olcina M, Pizzo A, Schattner TG (eds) Small Towns, una realidad urbana en la Hispania romana, Mytra, pp 283–292
Shoda S, Lucquin A, Sou CI, Nishida Y, Sun G, Kitano H, Son JH, Nakamura S, Craig OE (2018) Molecular and isotopic evidence for the processing of starchy plants in Early Neolithic pottery from China. Sci Rep 8(17044):1–9. https://doi.org/10.1038/s41598-018-35227-4
Simoneit BRT, Schauer JJ, Nolte CG, Oros DR, Elias VO, Fraser MP, Rogge WF, Cass GR (1999) Levoglucosan, a tracer for cellulose in biomass burning and atmospheric particles. Atmos Environ 33:173–182. https://doi.org/10.1016/S1352-2310(98)00145-9
Simoneit BRT, Rogge WF, Lang Q, Jaffé R (2000) Molecular characterization of smoke from campfire burning of pine wood (Pinus elliottii). Chemosphere: Glob Chang Sci 2(1):107–122. https://doi.org/10.1016/S1465-9972(99)00048-3
Spangenberg JE, Jacomet S, Schibler J (2006) Chemical analyses of organic residues in archaeological pottery from Arbon Bleiche 3, Switzerland- evidence for dairying in the late Neolithic. J Archaeol Sci 33:1–13. https://doi.org/10.1016/j.jas.2005.05.013
Steele VJ, Stern B, Stott AW (2010) Olive oil or lard?: distinguishing plant oils from animal fats in the archaeological record of the eastern Mediterranean using gas chromatography/combustion/isotope ratio mass spectrometry. Rapid Commun Mass Spectrom 24(23):3478–3484. https://doi.org/10.1002/rcm.4790
Suess MJ (1976) The environmental load and cycle of Polycyclic Aromatic Hydrocarbons. Sci Total Environ 6:239–250. https://doi.org/10.1016/0048-9697(76)90033-4
Thomas R, Stallibrass S (2008) For starters: producing and supplying food to the army in the Roman north-west provinces. In: Thomas R (ed) Stallibrass S. Oxbow books, Oxford, pp 1–17
Twiss KC (2019) The archaeology of food: identity, politics, and ideology in the prehistoric and historic past. Cambridge University Press
Whelton HL, Rofet-Salque M, Kotsakis K, Urem-Kotsou D, Evershed RP (2018) Strong bias towards carcass product processing at Neolithic settlements in northern Greece revealed through absorbed lipid residues of archaeological pottery. Quatern Int 496:127–139. https://doi.org/10.1016/j.quaint.2017.12.018
Whelton HL, Hammann S, Cramp LJE, Dunne J, Roffet-Salque M, Evershed RP (2021) A call for caution in the analysis of lipids and other small biomolecules from archaeological contexts. J Archaeol Sci 132:105397. https://doi.org/10.1016/j.jas.2021.105397
Woodworth M, Bernal D, Bonifay M, Garnier NB, Keay S, Pecci A, Poblome J, Pollard M, Richez F, Wilson A (2015) The content of African Keay 25/Africana 3 amphorae: initial results of the CORONAM project. In: Morais R, Morillo A (eds) Oliveira C. pp 41–57
Acknowledgements
We would like to thank the Institute of Environmental Science and Technology (ICTA-UAB) where we carried out the laboratory analyses. Additionally, we would like to thank Dr. Colominas for sharing unpublished zooarchaeological work. We also thank the excavation team of Guissona and Puig Castellar of Biosca for providing unpublished reports.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. The organic residue laboratory analysis was funded by the GIAP Archaeobotany Research Team funds and the Quadrennial Archaeological Project 2022–2025 ‘Transformacions urbanístiques, dinàmiques constructives i vida urbana a la ciutat romana de Iesso (Guissona, Segarra) (CLT009/22/00039)’ funded by the Department of Culture of the Government of Catalonia and the Guissona City Council. Alexandra Livarda was funded by the Agencia Estatal de Investigación (Ramón y Cajal RYC-2017–22105). Joan Villanueva was funded by the ICTA-UAB “María de Maeztu” Programme of the Spanish Ministry of Science and Innovation (CEX2019-000940-M). The Catalan Institute of Classical Archaeology (ICAC) funded the research of Theoni Baniou. Akshyeta Suryanarayan was funded by the EUTOPIA-SIF programme (European Union’s Horizon 2020 research and innovation programme under the Marie Sklodowska Curie grant agreement number 945380).
Author information
Authors and Affiliations
Contributions
TB: conceptualization of research, writing original draft, sample selection, performance of laboratory analysis (organic residue sample preparation and analysis), data analysis and investigation, preparation of figures 2 and 4. AS: methodology protocol set-up, sample selection, supervision of the laboratory work (organic residue sample preparation and analysis), revisions and corrections of the manuscript drafts, preparation of figure 5. AL: conceptualization and planning of the research, revisions, corrections and editing of the manuscript drafts, funding acquisition. NR: excavation and recovery of material under study, sample selection, revisions and corrections of the manuscript drafts, preparation of figures 1 and 3, funding acquisition. NM: laboratory analysis operation (GC-C-IRMS), laboratory analysis advisor, revision of methodology draft and writing up of the GC-C-IRMS methodology. JV: methodology protocol set-up, laboratory analysis advisor, revision of methodology draft and writing up of the GC-C-IRMS methodology. ER: excavation and recovery of material under study, sample selection, preparation of figures 1 and 3. All authors thoroughly revised the final draft and authorised its submission.
Corresponding author
Ethics declarations
Data declaration
All the authors confirm that all data analysed during this study are included within the manuscript and the supplementary information files.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Baniou, T., Suryanarayan, A., Livarda, A. et al. Tracing culinary practices in the western provinces of the Roman Empire using Organic Residue Analysis. Archaeol Anthropol Sci 16, 103 (2024). https://doi.org/10.1007/s12520-024-02011-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-024-02011-7