
Abstract
Salinisation has direct and detrimental physiological effects on freshwater organisms, yet little is known about its indirect effects. Here, we present a study that investigated the primary and secondary effects of salinisation on aquatic macroinvertebrates. We analysed macroinvertebrate samples from nine sites in river Wipper, Germany against nine stressor variables (water quality, salinisation, hydromorphological degradation). A Principal Component Analysis showed water quality deterioration and physical habitat degradation to constitute the main stressor gradients. Two macroinvertebrate community metrics (index of general degradation and percent Ephemeroptera-Plecoptera-Trichoptera specimens) revealed strong and significant positive correlations with water quality, but only weak and insignificant positive correlations with hydromorphological quality. High-resolution temporal measurements revealed notable oxygen deficits at a salinised and macrophyte-rich site that was dominated by the salt-tolerant macrophyte species Stuckenia [Potamogeton] pectinata (L.) Börner. At the site, oxygen levels frequently dropped below 6 mg/L during nighttime, suggesting an interaction of osmoregulatory stress (through salinisation) and respiratory stress (through plant mitochondrial respiration) that is caused by the mass development of a salt-tolerant macrophyte species. This biologically driven interaction of direct and indirect salinisation effects adds another level of complexity to the influence of multiple stressors in lotic systems and requires consideration in river management and restoration.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Salinisation of terrestrial and freshwater ecosystems is considered an emerging stressor (Kaushal et al., 2021), last but not least due to its connection with global and climate change (De Castro-Català et al., 2015). In semi-arid regions, crop production often requires intense irrigation, which can foster soil salinisation (Schuler et al., 2018) and then may impose negative effects on streams and rivers. With global warming, the thirst for irrigation water is likely to increase also in temperate regions. Large parts of central Europe, for example, witnessed extremely dry spring seasons between 2007 and 2020 (Ionita et al., 2020). The ten hottest summers in Europe have all occurred since 2003 (Copernicus, 2022). Hence, both global warming and a higher probability of extreme drought events are likely to put freshwater ecosystems further at risk.
Besides irrigation, the production and application of fertilizers—namely potash (potassium chloride, KCl)—constitutes another important source of salinisation (Cañedo-Argüelles et al., 2018; Schulz & Cañedo-Argüelles, 2018). Potash is a natural mineral usually obtained by underground mining. However, it is only the potassium ion (but not chloride) that renders potash an invaluable fertiliser, because potassium has been found to enhance the stress tolerance of many agricultural crops such as corn, wheat and rye (Wang et al., 2013). Manufacturing potassium fertilisers from salt rocks gives rise to considerable amounts of residues often containing several non-targeted salt ions (e g., chloride, sodium, sulphate). These residues are treated as waste and are usually dumped, for example, as open solid stock piles. Being exposed to precipitation, the salt is quickly dissolved, which renders open stock piles and their drainage a considerable source of brine. Disposal of brine into rivers usually has severe adverse effects on the riverine biodiversity (Cañedo-Argüelles et al., 2013; Kefford et al., 2016).
From an organismal perspective, salt ions are directly operating and physiologically highly active and hence may impose immediate osmoregulatory stress on many freshwater organisms, including aquatic macrophytes, macroinvertebrates and fish (Beisel et al., 2011). The stress level is dependent on the overall concentration of salt ions (i.e., the level of salinisation), but also on the composition of particular ions. Anions such as chloride (Cl−) and sulphate (SO4− −) have been found to impose higher stress levels as compared to cations such as potassium (K+), calcium (Ca++) and magnesium (Mg++) (Ziemann, 1967; Schuler et al., 2018). Halse et al. (2003) reported a strong, non-linear decline of macroinvertebrate richness under moderate levels of chloride, whereas recent studies from Germany imply the loss of many EPT (Ephemeroptera, Plecoptera and Trichoptera) taxa at chloride levels as low as 30–50 mg/L (Sundermann, 2015; Halle et al., 2017). For sulphate, Simmons (2012) found highly toxic effects on the duckweed Lemna minor L., which, however, were different in the presence of sodium and potassium. While sodium increased the toxicity of sulphate (supplied as Na2SO4), the presence of potassium and calcium enhanced the duckweed’s tolerance to sodium sulphate. A similar protective effect of calcium has been reported by Nanjappa et al. (2022) for the mayfly Austrophlebioides sp. AV2 Campbell & Suter. While sodium sulphate turned out to be most toxic, potassium chloride (KCl), calcium chloride (CaCl2) and potassium sulphate (K2SO4) were least toxic. These examples imply beneficial effects of several cations (but not sodium) on the salt tolerance of aquatic plants (Rout & Shaw, 2001). In contrast a review by Griffith (2016), which addressed the osmoregulatory and ionoregulatory effects of several major salt ions and estimated their toxicity for freshwater fish, arthropods and molluscs, concluded that potassium appeared to have the highest potential for causing toxic effects to freshwater animals under high concentrations in the water.
Besides salinisation levels and the particular ion’s identity, the biological response to salinisation is dependent on the osmoregulatory ability of particular species and species groups. The capability to control and regulate the inner osmolarity is well pronounced, for example, for many freshwater fish, but less so for freshwater plants and macroinvertebrates (Beisel et al., 2011; Wolfram et al., 2014). Therefore, salinisation can have immediate adverse effects on freshwater biodiversity and ecosystem functioning (Cañedo-Argüelles et al., 2013, 2018), especially among those species groups that lack a pronounced osmoregulatory capability. This is supported by the low tolerance of many riverine EPT taxa (Sundermann, 2015; Halle et al., 2017), which contribute notably to riverine macroinvertebrate biodiversity as well as to ecosystem functions such as organic matter decomposition (Wallace & Webster, 1996).
While the immediate and direct osmoregulatory effects of salinisation on freshwater organisms are well reported (Beisel et al., 2011; Silver & Donini, 2021, see also Cunillera-Montcusi et al., 2022 for a review), little is known about potential indirect effects. Personal observations from river Wipper in Thuringia, Germany, for example, suggest a boost of salt-tolerant macrophytes, namely Stuckenia pectinata (L.) Börner in heavily salinised sections of the river (up to 1500 mg/L chloride). There, the plant’s respiration during nighttime may impose additional respiratory stress on breathing organisms such as macroinvertebrates or fish. Following this causal inference, salinisation would indirectly cause oxygen depletion, with primary production being the potential link between primary and secondary effects. Here, we present a study that was driven by the attempt to disentangle the causes of degradation of benthic macroinvertebrate communities in the brine-loaded catchment of river Wipper, Thuringia, Germany. In particular, we investigated the role of salinisation (cations and anions) originating from potash mining and tried to disentangle the potential link between direct (osmoregulatory) and indirect (macrophyte-driven respiratory) effects of salinisation on macroinvertebrates.
Material and methods
Study area
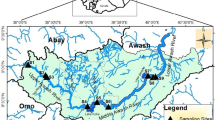
River Wipper is located south of the mountain range ‘Hainleite’ in Thuringia, Germany (Fig. 1). Its 92 km long course between the source at the city of Worbis in the west (330 m a.s.l.) and the mouth (into river Unstrut) at the city of Sachsenburg in the east (142 m a.s.l.) is characterised by calcareous minerals. Due to the calcareous geology, three different stream types can be found according to Pottgiesser (2018); type 6 (small fine substrate-dominated calcareous highland rivers), type 7 (small coarse substrate-dominated calcareous highland rivers) and type 9.1 (mid-sized fine to coarse substrate-dominated calcareous highland rivers). The carbonatic character of the catchment is distinctly pronounced by several salt springs at the river bottom. These springs constitute a natural source of salinisation, yet with much lower concentrations of salt ions as compared to the brine (Sommer et al., 2012). Because of the lack of sound data on natural salinisation, both natural and mining-related salinisation were not separated in this study. The 647 km2 catchment predominantly drains agricultural land (crops, pastures, meadows), with only a small proportion of forests and a few small cities located along the river course, of which Sondershausen is the biggest one (approx. 21,000 inhabitants).

Map of the Wipper basin with location of sampling sites
Due to intensive potash mining activities between the 1890s and 1990s along the middle part of the Wipper and one of its tributaries, river Bode, large parts of the Wipper system continue to be impacted by salinisation, with chloride concentrations exceeding 1500 mg/L at Hachelbich in the lower part of the catchment (Fig. 1). In addition to chloride, the main salt ions include sulphate, sodium, magnesium, calcium and potassium, the latter of which is the target ion for the production of potash fertiliser (KCl). For decades, non-targeted production residues were dumped at four dumps close to the rivers Wipper and Bode and since then act as a constant donor of brine. If mobilised by rainwater, the salt enters the Wipper system either through surface runoff or through groundwater infiltration. Moreover, brine from three dumps is stored in a retention basin and released to the lower Wipper depending on the river’s discharge. This constant pollution by salt has turned large parts of the Wipper into a degraded river system of poor to bad ecological status according to the WFD assessment (Fig. 1).
Data sources
Abiotic monitoring data of the Wipper
The following physical water quality variables were measured in the field using field probes, either in parallel or shortly before biological sampling in late spring 2016: electric conductivity (LF 196, WTW company Weilheim, Germany), pH (inoLab, WTW company Weilheim, Germany), oxygen concentration (Oxi 330, WTW company Weilheim, Germany) and water temperature (LF 196, WTW company Weilheim, Germany). In parallel to physical measurements, a water sample (1 L) was taken and kept cool for transport to the lab, where chemical analysis took place according to national standards. Lab analysis of chemical variables encompassed salt ions (chloride, sulphate, magnesium, calcium, sodium, potassium), nitrogen components (ammonia nitrogen, nitrate nitrogen) and phosphorous components (ortho-phosphate phosphorus, total phosphorus).
Fifteen physical habitat variables including hydrological (e.g., stagnation, presence of weirs/dams), morphological (e.g., bottom substrate conditions and diversity, bank modifications and special features) and riparian conditions were also recorded in the field in 2013 at the scale of 100 m intervals. For each interval, physical habitat variables were summarised in a physical habitat quality index (Gellert et al., 2014). For our analyses, only those 100 m river sections were considered that also included biological sampling sites. Because the physical habitat structure of the Wipper remained relatively stable in the past decade (no major modification like flood protection or restoration measures took place), the physical habitat data from 2013 are considered comparable with the data obtained in 2016.
Catchment land use/cover of biological sampling sites was determined by clipping ATKIS land cover data (ADV, 2021) with polygons of each site’s individual catchment using a Geographical Information System (GIS). Four land use/cover categories were distinguished: forest, grassland, crop and urban area. Other categories (e.g., surface waters, industrial areas) made up less than one percent coverage of the site’s catchment and thus were excluded from the analysis.
High-resolution abiotic measurements along the Wipper
At two sites (Fig. 1), automatic probes were installed to measure physical and chemical parameters online at a temporal resolution of 15-min intervals (except Ptotal: 60 min; Table 3). This allowed for a detailed comparison of physical and chemical conditions under the influence of brine. The site at Bleicherode was located close to the biological sampling site ‘Bleicherode Halde’ in the Wipper upstream of its confluence with river Bode. The site was moderately impacted by brine discharge. In contrast, the site at Hachelbich was located further downstream, close to the biological sampling site ‘Hachelbich’ (Fig. 1) and severely impacted by brine discharge. Online measurement ran from April 1st until October 31st, 2016. Prior to statistical analysis, non-plausible extreme measurements (e.g., due to malfunction of probes) were identified as lying outside the interval [25th quantile–1.5-fold interquartile range, 75th quantile + 1.5-fold interquartile range] and removed from the dataset. Unreliable values (e.g., data gaps and zero-values recorded during and shortly before/after several maintenance events) were graphically identified and removed from the dataset.
To enable separate statistics for daytime and nighttime measurements, data/time stamps of individual measurements were linked to daily sunrise/sunset times using the R package ‘photobiology’ (Aphalo, 2015), with time zone set to Europe/Berlin and longitude/latitude set to the city of Bleicherode (Fig. 1). Daytime (as the estimated start of daily primary production) was set to sunrise plus one hour, while nighttime start was set to sunset as the expected daily end of the photoperiod.
Biological monitoring data at the Wipper
Biological sampling was conducted at nine sites (Fig. 1) between April and August 2016. Macrophyte sampling followed the German standard protocol (Schaumburg et al., 2004), with in situ identification of all macrophyte species within a sampling stretch of 50 m and estimations of each species’ abundance according to Kohler (1978). Kohler’s abundance classes (1 = very rare … 5 = very abundant/mass development) were transferred to percent coverage according to Londo (1974) (1: < 1%, 2: 1–5%, 3: > 5–25%, 4: > 25–50%, 5: > 50–100%). Macrophyte taxalists were assessed with the German standard assessment system Phylib v5.3 (Schaumburg et al., 2012).
Macroinvertebrate sampling followed the German standard protocol ‘Perlodes’ (Meier et al., 2006), which is based on multi-habitat sampling (Hering et al., 2004). At each site, within a sampling stretch of 50–100 m, bottom substrate cover was estimated at 5% increments. A total of 20 sub-samples (20 × 5% = 100%) were then allocated according to the individual substrate’s bottom cover. For example, a substrate covering 30% of the bottom would be assigned six sub-samples. Sub-samples were taken with a kick-net (25 × 25 cm, mesh: 500 µm) that was placed on the bottom, while the substrate in front of the net was disturbed (by hand or heel) to move disclosed macroinvertebrates into the net. All sub-samples were then pooled, transferred into a container and fixed with ethanol (final concentration: 70%) prior to lab transport. Samples were stored at 4 °C in the dark in a refrigerator until further processing. Representative sub-samples comprising at least a sixth of the pooled sampling material and 350 macroinvertebrate specimens were taken from each pooled sample and sorted. After sorting, macroinvertebrates were identified to the taxon levels as defined by the operational taxalist (Haase et al., 2019), which is species level except for oligochaetes (worms) and dipterans (flies, midges). Eventually, macroinvertebrate taxalists were processed with the software ASTERICS v4.04 (ASTERICS, 2014) to obtain (i) the overall ecological status class (ECS), (ii) the ecological quality ratio (EQR) of the module ‘General Degradation’, which assesses the physical habitat quality (AD_EQR), and (iii) the ecological quality ratio of the proportion of EPT specimens in the community (EPT_EQR). EQRs are calculated by dividing the observed value by a stream type-specific expected value (O/E ratio). For a detailed description of the metrics, see Schaumburg et al. (2012) and Meier et al. (2006).
Additional monitoring data of further German rivers
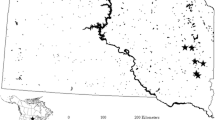
Due to the low number of available monitoring sites in the Wipper system, additional monitoring data from other watersheds in Germany were analysed to verify the results based upon the Wipper data. The dataset comprised macrophyte coverages and salt ions from 60 observations at 39 rivers of comparable stream types (type 6, 7 and 9.1 acc. to Pottgiesser, 2018), to ensure that the natural geo-morphological background conditions of the verification sites matched those in the Wipper. Sampling of biological and abiotic data of the verification dataset followed the same protocols as outlined above for the Wipper monitoring dataset.
Statistical analysis
All statistical analyses and graphical representations of results were obtained with R v4.0.5 (R Core Team, 2021). Statistical significance of observed differences between groups of sites were analysed using the Mann–Whitney U-test (function wilcox.test()) with the minimum type-I error set to P < 0.05, however, significant results could not be obtained due to the small sample size.
Because of the limited sample size, regression modelling (e.g., GLM) seemed inappropriate to derive the stressor hierarchy from the Wipper data. Therefore, we chose a combination of Principal Components Analysis (PCA) and subsequent correlation with the main PCA components to compare the relationship of stressor variables and macroinvertebrate metrics. First, physical, chemical, and physical habitat stressors were subjected to a variance inflation factor (VIF) analysis (package usdm; Naimi et al., 2014), to identify non-collinear stressor variables. Stressor variables with a VIF > 10 (Dormann et al., 2012) were excluded from further analyses. Also, land use variables were excluded, because they revealed largely similar (i.e., invariable) values at all sites. Second, a PCA using the function princomp() and a correlation matrix was run with five non-collinear stressor variables, to identify the main stressor gradients. And third, the relationship between the main PCA components and the two macroinvertebrate metrics was investigated using Spearman’s correlation coefficient ρ, to quantify the relationship. Correlations were tested for significance with R’s function cor.test().
Results
Ecological status of the Wipper system
Environmental monitoring revealed that the Wipper system is heavily impacted by several stressors (Table 1). Stressor levels not meeting the national or other published quality targets show strong impacts by nutrient enrichment, salinisation, physical habitat modification and land use. While nutrient enrichment was moderate, stressors of the remaining three categories (salinisation, physical habitat modification and land use) exceeded quality targets manifold.
Ecological quality ratios (EQR) of macrophytes and macroinvertebrates along the river continuum confirm pronounced ecological implications in the lower Wipper, which could be attributed mainly to water and habitat quality issues (Fig. 2). Water quality in the lower Wipper was predominantly deteriorated by enhanced concentrations of chloride, sulphate and potassium. The (daytime) oxygen deficits at Großfurra and Hachelbich (Fig. 2), together with rather inconsistent trends in ammonia, nitrate and ortho-phosphate along the continuum suggest interactions of physical and chemical stressors with macrophytes (mainly S. pectinata) and benthic algae [mainly Cladophora glomerata (L.) Kütz and Enteromorpha intestinalis (L.) Nees].

Nine stressor variables and two biological indices at seven sites (black dots) along the Wipper continuum. Sites are (from left to right): Worbis, Wülfingerode, Sollstedt, Bleicherode Halde, Großfurra, Hachelbich and Sachsenburg (see Fig. 1 for a site map). Except for oxygen, higher values on the y-axis represent higher stress levels. Dashed lines mark official German environmental quality targets (OGewV, 2016; red colour) or otherwise reported target values (Feld, 2013 for % crop in the catchment; UBA, 2003 for nitrate and potassium; black colour)
Effects of water and habitat quality on macrophytes
The comparison of stressor levels in macrophyte-poor (upstream) and macrophyte-rich (downstream) river sites shows mainly water quality variables to distinguish between both sections (Table 2), whereas differences in physical habitat quality and land use were less pronounced (see also Fig. S1 in the Supplementary Material for a comparison of salt vs. habitat as descriptors of macrophyte coverage). Nitrate dropped from levels above 5 mg/L at the sites upstream of Großfurra, to levels ranging 2–3 mg/L further downstream, thus implying an uptake of nitrate due to primary production. The levels of ammonia, ortho-phosphate and total phosphorus, however, were at least doubled in the lower Wipper and suggest the presence of active sources of pollution along the river continuum (see also Fig. 2), rather than an interaction with macrophytes. Macrophyte-rich sites were also characterised by warmer water, which – in part – may point at a stronger solar heating under reduced flow conditions in dense macrophyte stands. Yet, the warming of streams along the continuum in general is a natural phenomenon, which is difficult to disentangle from other (anthropogenic) causes of heating due to the lack of sufficient data in this study.
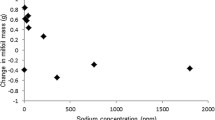
Notably, all six salt ions showed much higher mean concentrations at macrophyte-rich sites (significance not tested due to small sample size) (Table 2), hence confirming a pronounced tolerance of S. pectinata, which was the dominant species in the lower Wipper. The tolerance of S. pectinata towards salinisation was confirmed by data of 60 samples taken at 39 salinised river sites outside (but typologically comparable to) the Wipper system (Fig. 3). Apparently, the species can reach high a coverage (i.e., abundance class 4 or 5, translating to coverages of 25–50% or 50–100%, respectively, according to Kohler, 1978 and Londo, 1974) already at chloride levels around 250 mg/L. Furthermore, the species seems to respond fairly tolerant to enhanced concentrations of sulphate and potassium. It is important to note that both the findings in the Wipper system and at comparable sites outside the system do not imply a boost of S. pectinata by salt, but confirm the pronounced tolerance of the species towards (chloride) salinisation.

Coverage of Stuckenia [Potamogeton] pectinata against the concentration of chloride, sulphate, sodium and potassium in an external dataset of 60 samples from 39 river sites outside the Wipper system in Germany. External river sites were constrained to the same three river types that occurred in the Wipper system (6, 7 and 9.1 acc. to Pottgiesser, 2018). Coverage according to Londo (1974): 1 = < 1% coverage, 2 = 1–5%, 3 = > 5–25%, 4 = > 25–50%, 5 = > 50–100%
High-resolution time-series monitoring revealed strong oxygen deficits at the macrophyte-rich site Hachelbich (Table 3), with 49% of nigh-time measurements not meeting the (German) quality target of 7 mg/L oxygen concentration. Contrastingly, high-resolution oxygen measurements at the macrophyte-poor site Bleicherode did not exhibit notable phases with oxygen depletion during night-time, and > 93% of all measurements (day + night) met the quality target 7 mg/L. Diurnal differences in high-resolution measurements of other physical and chemical variables were negligible, except for water temperature, which on average was 1 centigrade (= 1 Kelvin) lower during nighttime at all sites (Table 3).
Effects of water and habitat quality on macroinvertebrates
Variance inflation factor analysis revealed five non-collinear stressors in the Wipper system (N = 9 sites) allocated to three stressor groups: (i) nutrients (ammonia, ortho-phosphate), (ii) salt (chloride, sulphate) and (iii) physical habitat (habitat quality index ‘GSG’). Percent catchment land uses (crop land, urban area) of sites were invariable along the river continuum and thus excluded from the analysis. When subjected to a PCA, two main components showed eigenvalues above 1 and sufficed to explain 70% of the variability in the stressor data (Fig. 4). PCA 1 (49.4%) described a water quality gradient subsuming the impacts of enhanced nutrient and salt concentrations. PCA 2 (20.6%) described a gradient of physical habitat degradation. Because PCA components are mutually orthogonal (uncorrelated) variables, both main stressor gradients were considered independent of each other.

Biplot of a PCA on values of five non-collinear stressor variables measured at nine sites in the Wipper system. Both main components PCA 1 and 2 have an eigenvalue above 1 and together explain 70% of the variability in the data. Stressor variables are ammonia–nitrogen (NH4-N), chloride (Cl), ortho-phosphate phosphorus (oPO4-P), sulphate (SO4) and physical habitat quality (GSG)
To contrast both stressor gradients with the organisms’ response, the sample sites’ PCA scores were extracted and correlated with two macroinvertebrate indices, namely the common degradation index (AD_EQR) and the proportion of EPT specimens (EPT_EQR) (Fig. 5). Correlations were notably stronger for the water quality component (r = − 0.78, P < 0.05 and r = − 0.81, P < 0.01, respectively) as compared to the physical habitat component (r = 0.27 and r = 0.20, respectively; both insignificant). The findings confirm water quality issues to be the major descriptor of macroinvertebrate ecological deterioration in river Wipper. However, although much smaller and insignificant, the trends of both biological metrics along the (uncorrelated) second PCA component suggest an indicative potential of macroinvertebrates for physical habitat degradation even under the pronounced influence of salinisation. The trend becomes particularly obvious for the common degradation index (AD_EQR) with samples of moderate to poor status (Fig. 5, yellow and orange dots in plot on the top righ-hand side).

Relationship of a macroinvertebrate common degradation index (AD_EQR) and the proportion of EPT (Ephemeroptera, Plecoptera, Trichoptera) specimens (EPT_EQR) with the two main principal components PC 1 (water quality) and PC 2 (habitat quality) as derived from the PCA (compare Fig. 4). Note that positive PC scores correspond to low water quality and high habitat quality, respectively. Spearman’s correlation coefficients are indicated in the top left of each plot. EQR stands for ecological quality ratio and ranges from 0 (poor quality) to 1 (high quality). The dots (sites) are colour coded according to their overall ecological classification
Discussion
River management requires management solutions that are tailored to the individual pressures and impacts of the water bodies of interest. This is one of the main conclusions of a recent pan-European project summarising nearly 70 studies on biological effects of a total of 174 stressor pairs in freshwater and marine ecosystems (Birk et al., 2020). Rivers across Europe turned out to be frequently impacted by a concert of impacts by, for example, nutrients, pesticides, oxygen depletion, warming, hydrological alteration, physical habitat degradation and land use, which eventually seem to drive complex biological responses.
Biological responses to water and habitat quality deterioration in the Wipper
Our study supports the conclusion by Birk et al. (2020) also for the Wipper system. The entire river course, including its major tributary, was affected by the influence of multiple chemical compounds originating from diffuse and point sources of pollution as well as from hydromorphological alterations of the river course. Yet, in addition the obvious mass development of salt-tolerant aquatic macrophytes, namely of S. pectinata, adds complexity to the environmental and biological deterioration of river Wipper, in that the plant species seems to link between direct (i.e., osmotic) and indirect (i.e., respiratory) adverse effects of salinisation on the macroinvertebrate community.
The most obvious direct effect is probably caused by a mixture of salt ions originating from the controlled and diffuse discharge of brine. Chloride levels, for example, quickly rose already in the upper part of the river and exceeded 1 g/L in the lower part. In light of current environmental quality targets in Germany (OGewV, 2016) and further proposed quality targets (Sundermann et al., 2015; Halle et al., 2017), strong adverse effects of chloride levels way beyond 1 g/L are very likely. Chloride, as many other salt ions, is known to impose direct and strong physiological stress on the aquatic stages of macroinvertebrates in that it enhances the water’s osmolarity. Because many macroinvertebrates possess only weak osmoregulatory capabilities (Wolfram et al., 2014), an increase of the osmolarity in the ambient medium is usually followed by a rapid increase of the osmolarity in the cell medium, which is lethal for many freshwater species. The toxicity of single ions (e. g., chloride, potassium) to freshwater macroinvertebrates has been described in detail by Schulz & Cañedo-Argüelles (2018).
Besides salinisation, our results revealed also enhanced nutrient levels, namely of phosphate and nitrate, to affect the water quality in the Wipper. Yet, in light of the observed concentrations of both compounds, direct physiological effects on benthic invertebrates seem rather unlikely (see Wang et al., 2013 for a discussion of direct vs. indirect nutrient effects). More likely, enhanced phosphorus and nitrogen levels may have driven primary production (Ouyang et al., 2018) and hence the mass development of aquatic macrophytes and algae in the lower Wipper. In this context, the decrease in the concentration of nitrate nitrogen in the lower Wipper may seem contradictory at first glance, yet may be explained by the plant’s uptake (retention) of nitrogen. In contrast, the increase of ortho-phosphate phosphorus in the lower Wipper suggests a continuous supply, for example, by point source pollution, which may have masked its retention in the plant biomass.
The direct influence of hydromorphological alterations of the Wipper on macroinvertebrates was less pronounced. Two macroinvertebrate community metrics revealed weak and insignificant correlations with a hydromorphological gradient, yet nevertheless suggest that structural deficiencies (e.g., lack of natural microhabitats, bank enforcement, lack of natural riparian vegetation) along the river course remain detectable by macroinvertebrates even under the influence of salinisation. At least for one metric (AD_EQR), a stronger trend would be observable with sites of moderate and poor ecological status (Fig. 5, plot of AD_EQR along PCA 2). The “bad” sites, however, may already have undergone a dramatic loss of species so that the remaining species pool was insufficient to track changes in the habitat quality. However, in light of the small sample size available for river Wipper, this assumption remains speculative and requires further evidence.
Macrophytes as connectors between osmoregulatory and respiratory stress on benthic macroinvertebrates
The strong positive relationship between salinisation and the mass development of macrophytes, namely of S. pectinata in the Wipper was striking and was supported by the analysis of data from similar rivers outside the Wipper system. Furthermore, because salt (Cl and K) was stronger correlated to macrophyte coverage than habitat quality (overall and bank quality) in the Wipper, it seems likely that salinity favoured the mass development of S. pectinata. There is sufficient evidence that salt-tolerant macrophytes (alike S. pectinata) can develop a high abundance even under heavily salinised conditions, with chloride concentrations beyond 1.4 g/L. While the tolerance of S. pectinata towards salinisation has been reported also by other studies (Wolfram et al., 2014), detailed studies on the physiological mechanisms controlling the salt tolerance of aquatic plants are sparse (e. g. by Simmons (2012) on the duckweed L. minor L. or by Rout & Shaw (2001) on some naiads Najas sp.). The studies suggest that the plant’s tolerance against chloride or sulphate may be controlled by potassium and other cations. Potassium-mediated stress tolerance is well reported for terrestrial plants, namely for crops such as barley (Wang et al., 2013), where potassium not only can help increase the plants tolerance against salinisation, but also its tolerance against other stressor, for example, drought stress. This explains, why potassium (as potash/potassium chloride, KCl) is supplied to agricultural crops. The high concentrations of potassium and chloride in river Wipper are directly linked to potash mining that took place mainly between the1890s and 1990s in the upper catchment. Nowadays, brine drainage from several open salt dumps in the catchment constantly convey salt into the Wipper—and will probably continue to do so for the next couple of decades (estimation by co-author CJS, State Office for the Environment, Mining and Nature Protection Thuringia).
The automatic probe measurements of oxygen levels at two sites in the Wipper revealed notable oxygen deficiencies at the macrophyte-rich site at Hachelbich during nighttime, as opposed to the macrophyte-poor site at Bleicherode. At Hachelbich, nocturnal oxygen concentrations frequently dropped below the German target level required to maintain a good ecological status (i.e., 7 mg/L; OGewV, 2016), while daytime oxygen levels at the same station ranged around 10 mg/L. This strong diurnal fluctuation of oxygen levels at Hachelbich can be explained by diurnal physiological patterns of primary producers. During daytime, photosynthesis is driven by sunlight and oxygen is produced. During nighttime, photosynthesis is replaced by mitochondrial respiration and the plants consume more oxygen than they produce. Oxygen consumption also appears during the aerobic degradation of plant biomass by microorganisms at the end of the vegetation period. Such plant-driven oxygen deficits, however, are likely to be overlooked by standard monitoring schemes, which usually involve oxygen measurements during daytime.
The frequent drops of oxygen levels below 6 mg/L that were observed for nearly 50% of the nighttime measurements at Hachelbich probably also occurred at other macrophyte-dominated sites in the lower Wipper. Hence, oxygen supply in the lower river was probably unfavourable for many macroinvertebrate species at nighttime during the summer period. Similar to salinisation, oxygen depletion has a direct physiological effect on the aquatic life stages of macroinvertebrates. Thereby, species without particular respiratory structures (e.g., gills) such as many plecopteran larvae respond more sensitive as opposed to ephemeropterans and trichopterans, whose larvae possess gills.
Implications for river management under the influence of salinisation
Due to its direct, osmo-regulatory effects on the physiology of both plants and animals salinisation constitutes a severe stressor of aquatic ecosystems (Beisel et al., 2011; Wolfram et al., 2014), whereas its effects might be boosted by climate change (Melles et al., 2023). Thereby, salt ions are not degradable and may accumulate in groundwater aquifers, sediments and biomass. This persistence of salt in ecosystems constitutes a long-term threat, which is likely to increase rapidly under the influence of global warming (Jeppesen et al., 2020), in particular in semiarid, but nevertheless agriculturally used regions around the world, such as the south-western USA, Mediterranean Europe, southern Africa, east Asia and Australia.
In concert, our findings suggest that a stressor (here: salt) can favour a biological response (here: boost of S. pectinata under salinisation) that in itself constitutes another stressor (here: oxygen depletion) with adverse effects on another organism group (here: macroinvertebrates). This complex interaction of osmoregulatory and respiratory stressors, which is caused by primary production, requires consideration in the management of salinised rivers.
Conclusion
Salinisation has direct and detrimental physiological effects on freshwater organisms. However, salinisation may favour particularly tolerant macrophyte species and then promote their mass development, which in turn may result in oxygen depletion in dense macrophyte stands during nighttime. Salt-tolerant species such as S. pectinata outcompete salt-sensitive species and can reach very high a coverage on the river bottom. Besides oxygen depletion during nighttime, the recent fish-kill in river Odra and the subsequent investigation of potential causes support such plant-driven effects of salinisation on other water quality parameters. In case of the recent fish-kill in river Odra, it was salinisation too that was assumed to have boosted the mass development of an golden algae (Prymnesium parvum N.Carter), whose toxicants are lethal for fish (IGB, 2022).
River management should acknowledge both the interaction of stressors and the role of biological responses as linkage between stressors. This includes the use of automatic probes, for example, to identify diurnal rhythms of particular stressors such as oxygen depletion, which tend to remain undetected by monitoring schemes applying daytime spot measures. In the long-term, however, a significant reduction of salt loads would be required to reduce the risk of unforeseen side effects and ultimately to achieve an ecological improvement of the aquatic communities.
Data availability
All data analysed for this article were obtained through national monitoring schemes in line with the demands of the European Water Framework Directive. Monitoring data are available on request at the Thüringer Landesamt für Umwelt, Bergbau und Naturschutz (TLUBN), Göschwitzer Str. 41, 07,745 Jena, Germany.
References
ADV, 2021. Amtliches Topographisch-Kartographisches Informationssystem (Official Topographic Information System). Retrieved from https://www.adv-online.de/icc/exteng/broker?uMen=4a1607d6-b048-65f1-80fa-29f08a07b51a (last access on 8 February 2023)
Allan, J. D., 2004. Landscapes and riverscapes: the Influence of land use on stream ecosystems. Annual Review of Ecology, Evolution and Systematics 35: 257–284.
Aphalo, P. J., 2015. The R photobiology suite. UV4Plants Bulletin 1, 21–29.
ASTERICS, 2014. ASTERICS and Perlodes Desktop Software version 4.0.4 (October 2014). Retrieved from https://www.gewaesser-bewertung.de/files/asterics_4.0.4-setup.zip (last access on 18 November 2023)
Beisel, J.-N., M.-C. Peltre & P. Usseglio-Polatera, 2011. Einfluss der Salzbelastung auf die aquatische Biozönose der Mosel (Impact of salt on aquatic communities of river Mosel). Bericht des Labors LIEBE im Auftrag der IKSMS, Metz, 1–62.
Birk, S., D. Chapman, L. Carvalho, B. M. Spears, H. E. Andersen, C. Argillier, S. Auer, A. Baattrup-Pedersen, L. Banin, M. Beklioglu, E. Bondar-Kunze, A. Borja, P. Branco, T. Bucak, A. D. Buijse, A. C. Cardoso, R. M. Couture, F. Cremona, D. de Zwart, D. de Zwart, C. K. Feld, M. T. Ferreira, H. Feuchtmayr, M. O. Gessner, A. Gieswein, L. Globevnik, D. Graeber, W. Graf, C. Gutiérrez-Cánovas, J. Hanganu, U. Işkın, M. Järvinen, E. Jeppesen, N. Kotamäki, M. Kuijper, J. U. Lemm, S. Lu, A. Lyche Solheim, U. Mischke, S. J. Moe, P. Nõges, T. Nõges, S. J. Ormerod, Y. Panagopoulos, G. Phillips, L. Posthuma, S. Pouso, C. Prudhomme, K. Rankinen, J. J. Rasmussen, J. Richardson, A. Sagouis, J. M. Santos, R. B. Schäfer, R. Schinegger, S. Schmutz, S. C. Schneider, L. Schülting, P. Segurado, K. Stefanidis, B. Sures, S. J. Thackeray, J. Turunen, M. C. Uyarra, M. Venohr, P. C. von der Ohe, N. Willby & D. Hering, 2020. Impacts of multiple stressors on freshwater biota across spatial scales and ecosystems. Nature Ecology & Evolution 4: 1060–1068.
Cañedo-Argüelles, M., B. J. Kefford, C. Piscart, N. Prat, R. B. Schäfer & C.-J. Schulz, 2013. Salinisation of rivers: an urgent ecological issue. Environmental Pollution 173: 157–167.
Cañedo-Argüelles, M., B. Kefford & R. B. Schäfer, 2018. Salt in freshwaters: causes, effects and prospects - introduction to the theme issue. Philosophical Transactions of the Royal Society B: Biological Sciences 374: 20180002.
Copernicus (2022) Seasonal review: Europe’s record-breaking summer. Retrieved from https://climate.copernicus.eu/seasonal-review-europes-record-breaking-summer (last access on 8 February 2023)
Cunillera-Montcusi, D., M. Beklioglu, M. Cañedo-Argüelles, E. Jeppesen, R. Ptacnik, C. A. Amorim, S. E. Arnott, S. A. Berger, S. Brucet, H. A. Dugan, M. Gerhard, Z. Horvath, S. Langenheder, J. C. Nejstgaard, M. Reinikainen, M. Striebel, P. Urrutia-Cordero, C. F. Vad, E. Zadereev & M. Matias, 2022. Freshwater salinisation: a research agenda for a saltier world. Trends in Ecology and Evolution 37: 440–453.
De Castro-Català, N., I. Muñoz, L. Armendáriz, B. Campos, D. Barcelo, J. Lopez-Doval, S. Pérez, M. Petrovic, Y. Pico & J. L. Riera, 2015. Invertebrate community responses to emerging water pollutants in Iberian river basins. Science of the Total Environment 503(504): 142–150.
Dormann, C. F., J. Elith, S. Bacher, C. Buchmann, G. Carl, G. Carré, J. R. G. Marquéz, B. Gruber, B. Lafourcade, P. J. Leitão, T. Münkemüller, C. McClean, P. E. Osborne, B. Reineking, B. Schröder, A. K. Skidmore, D. Zurell & S. Lautenbach, 2012. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36: 27–46.
Feld, C. K., 2013. Response of three lotic assemblages to riparian and catchment-scale land use: implications for designing catchment monitoring programmes. Freshwater Biology 58: 715–729.
Gellert, G., T. Pottgiesser & T. Euler, 2014. Assessment of the structural quality of streams in Germany – basic description and current status. Environmental Monitoring & Assessment 186: 3365–3378.
Griffith, M. B., 2017. Toxicological perspective on the osmoregulation and ionoregulation physiology of major ions by freshwater animals: teleost fish, Crustacea, aquatic insects, and Mollusca. Environmental Toxicology and Chemistry 36: 576–600.
Haase, P., A. Sundermann & K. Schindehütte, 2019. Operationelle Taxaliste als Mindestanforderung an die Bestimmung von Makrozoobenthosproben aus Fließgewässern zur Umsetzung der EU-Wasserrahmenrichtlinie in Deutschland (An operational taxalist as a minimum requirement for the identification of benthic macroinvertebrates from running waters for the implementation of the EU WFD in Germany). Retrieved from https://www.gewaesser-bewertung.de/files/operationelle_taxaliste_2019.xlsx (last access on 11 November 2022)
Halle, M. & A. Müller, 2014. Korrelationen zwischen biologischen Qualitätskomponenten und allgemeinen chemischen und physikalisch-chemischen Parametern in Fließgewässern (Correlation between biological quality components and general physical and chemical parameters in running waters). Endbericht zum Projekt O 3.12 des Länderfinanzierungsprogramms „Wasser, Boden und Abfall“ 2012, 190 S.
Halle, M. & Müller, A. (2015) Typspezifische Ableitung von Orientierungswerten für den Parameter Sulfat (Stream type-specific derivation of assessment values for the parameter sulphate). Abschlussbericht zum Folgeprojekt im Auftrag des Sächsischen Landesamt für Umwelt Landwirtschaft und Geologie (LfULG) zum Projekt O 3.12 des Länderfinanzierungsprogramms „Wasser, Boden und Abfall“ 2012, 200 S. Retrieved from https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwj73p2184X9AhXrSfEDHfwdBWwQFnoECAwQAQ&url=https%3A%2F%2Fgewaesser-bewertung.de%2Ffiles%2Flawa_acp_projekt_o3.12_endbericht_17apr2014.pdf&usg=AOvVaw0imNXZkVNMC7KKmIk_wYPe (last access on 8 February 2023)
Halle, M., A. Müller & E. Bellack, 2017. Schwellenwerte und Bioindikatoren zur gewässerbiologischen Beurteilung des Salzgehaltes von Fließgewässern gemäß EG-WRRL (Threshold values and bioindicators for the evaluation of salt in streams and rivers). Korrespondenz Wasserwirtschaft 9: 525–535.
Halse, S. A., J. K. Ruprecht & A. M. Pinder, 2003. Salinisation and prospects for biodiversity in rivers and wetlands of south-west Western Australia. Australian Journal of Botany 51: 673–688.
Hering, D., O. Moog, L. Sandin & P. F. M. Verdonschot, 2004. Overview and application of the AQEM assessment system. Hydrobiologia 516: 1–20.
IGB (Leibniz Institute of Freshwater Ecology and Inland Fisheries), 2022. Environmental disaster on the Oder: IGB researchers track potentially toxic algae. Press release by J. Graupner on 17 August 2022. Retrieved from https://www.igb-berlin.de/en/news/environmental-disaster-oder-igb-researchers-track-potentially-toxic-algae (last access on 7 February 2023)
Ionita, M., V. Nagavciuc, R. Kumar & O. Rakovec, 2020. On the curious case of the recent decade, mid-spring precipitation deficit in central Europe. Climate and Atmospheric Sciences 3: 49.
Jeppesen, E., M. Beklioglu, K. Özkan & Z. Akyürek, 2020. Salinization increase due to climate change will have substantial negative effects on inland waters: A call for multifaceted research at the local and global scale. The Innovation 1: 100030.
Kaushal, S. S., G. E. Likens, M. L. Pace, J. E. Reimer, C. M. Maas, J. G. Galella, R. M. Utz, S. Duan, J. R. Kryger, A. M. Yaculak, W. L. Boger, N. W. Bailey, S. Haq, K. L. Wood, B. M. Wessel, C. E. Park, D. C. Collison & B.Y. Aisin a. I., T.M. Gedeon, S.K. Chaudhary, J. Widmer, C.R. Blackwood, C.M. Bolster, M.L. Devilbiss, D.L. Garrison, S. Halevi, G.Q. Kese, E.K. Quach, C.M.P. Rogelio, M.L. Tan, H.J.S. Wald & S.A. Woglo, 2021. Freshwater salinization syndrome: from emerging global problem to managing risks. Biogeochemistry 154: 255–292.
Kefford, B. J., D. Buchwalter, M. Cañedo-Argüelles, J. Davis, R. P. Duncan, A. Hoffmann & R. Thompson, 2016. Salinized rivers: degraded systems or new habitats for salt-tolerant faunas? Biological Letters 12: 20151072.
Kohler, A., 1978. Methoden der Kartierung von Flora und Vegetation von Süßwasserbiotopen (Methods to record the flora and vegetation of freshwater biotopes). Landschaft Und Stadt 10: 73–85.
Londo, G., 1974. The decimal scale for relevés of permanent quadrats. In Knapp, R. (ed), Sampling methods in vegetation science W. Junk Publishers, The Hague/Boston/London: 45–49.
Meier, C., P. Haase, P. Rolauffs, K. Schindehütte, F. Schöll, A. Sundermann & D. Hering, 2006. Methodisches Handbuch Fließgewässerbewertung (Methodological handbook for the assessment of running waters). Retrieved from https://www.gewaesser-bewertung.de/files/meier_handbuch_mzb_2006.pdf (last access on 11 November 2022)
Melles, S. J., M. Cañedo-Argüelles & A. M. Derry, 2023. Documenting the impacts of increasing salinity in freshwater and coastal ecosystems: Introduction to the special issue. Limnology and Oceanography Letters 8: 1–7.
Naimi, B., N. A. Hamm, T. A. Groen, A. K. Skidmore & A. G. Toxopeus, 2014. Where is positional uncertainty a problem for species distribution modelling. Ecography 37: 191–203.
Nanjappa, V., S. Vink, J. Dunlop, M. N. Krosch & R. Mann, 2022. Influence of calcium on the toxicity of saline solutions to the mayfly, Austrophlebioides sp. AV II. Marine and Freshwater Research 73: 1499–1509.
OGewV (Oberflächengewässerverordnung), 2016. Oberflächengewässerverordnung vom 20. Juni 2016 (BGBl. I S. 1373), zuletzt durch Artikel 2 Absatz 4 des Gesetzes vom 9. Dezember 2020 (BGBl. I S. 2873) geändert. Bundesministerium der Justiz und für Verbraucherschutz und Bundesamt für Justiz Berlin, 87 S. Retrieved from http://www.gesetze-im-internet.de/ogewv_2016/ (last access on 8 February 2023)
Ouyang, Z., S. S. Qian, R. Becker & J. Chen, 2018. The effects of nutrients on stream invertebrates: a regional estimation by generalized propensity score. Ecological Processes 7: 21.
Pottgiesser, T., 2018. Zweite Überarbeitung der Steckbriefe der deutschen Fließgewässertypen (2nd Revision of the passports of the German stream types). FE-Vorhaben des Umweltbundesamtes „Gewässertypenatlas mit Steckbriefen“ (FKZ 3714 24 221 0). umweltbüro Essen, 24 S. + Anhang. Retrieved from https://www.gewaesser-bewertung.de/files/steckbriefe_fliessgewaessertypen_dez2018.pdf (last access on 11 November 2022)
R Core Team, 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Retreived from https://www.R-project.org/ (last access on 8 February 2023)
Rout, N. P. & B. P. Shaw, 2001. Salt tolerance in aquatic macrophytes: possible involvement of the antioxidative enzymes. Plant Science 160: 415–423.
Schaumburg, J., C. Schranz, J. Foerster, A. Gutowski, G. Hofmann, S. Schneider & U. Schmedtje, 2004. Ecological classification of macrophytes and phytobenthos for rivers in Germany according to the Water Framework Directive. Limnologica 34: 283–301.
Schaumburg, J., C. Schranz, D. Stelzer, A. Vogel & A. Gutowski, 2012. Verfahrensanleitung für die ökologische Bewertung von Fließgewässern zur Umsetzung der EG-Wasserrahmenrichtlinie: Makrophyten und Phytobenthos (Manual for the ecological assessment of running waters fort he implementation of the EU WFD: macrophytes and phytobenthos) - Phylib, Stand 13.02.2012. Retrieved from https://www.gewaesser-bewertung.de/files/verfahrensanleitung_fg.pdf (last access on 11 November 2022)
Schuler, M. S., M. Cañedo-Argüelles, W. D. Hintz, B. Dyack, S. Birk & R. A. Relya, 2018. Regulations are needed to protect freshwater ecosystems from salinization. Philosophical Transactions of the Royal Society B: Biological Sciences 374: 20180019.
Schulz, C.-J. & M. Cañedo-Argüelles, 2018. Lost in translation: the German literature on freshwater salinization. Philosophical Transactions of the Royal Society B: Biological Sciences 374: 20180007.
Silver, S. & A. Donini, 2021. Physiological responses of freshwater insects to salinity: molecular-, cellular- and organ-level studies. Journal of Experimental Biology 224: 20.
Simmons, J. A., 2012. Toxicity of major anions and cations (Na+, K+, Ca2+, Cl-, and SO42-) to a macrophyte and an algae. Environmental Toxicology and Chemistry 31: 1370–1374.
Sommer, T., D. Swaboda, U. Uhlig, T. Gottschalk & U. Stodolny, 2012. Gesamtkostenstudie zur wasserwirtschaftlichen Nachsorge der Haldenstandorte im Kali-Südharz-Revier. Studie im Auftrag der Gesellschaft zur Verwahrung und Verwertung von stillgelegten Bergwerksbetrieben mbH (GVV). Grundwasserforschungsinstitut (GFI) Dresden, 181 S. (Study on the monetary evaluation of the maintenance of potassium mining stock piles in the southern Harz region.) Retrieved from https://aktion-fluss.de/wp-content/uploads/ga-studie-bewirtschaftungsziele-kalibergbau-suedharz.pdf (last access on 8 February 2023)
Sundermann, A., M. Leps, S. Leisner & P. Haase, 2015. Taxon-specific physico-chemical change points for stream benthic invertebrates. Ecological Indicators 57: 314–323.
TLUG (Thüringer Landesanstalt für Umwelt und Geologie), 2011. Handbuch zur naturnahen Unterhaltung und zum Ausbau von Fließgewässern (Handbook for the near-natural maintenance and development of running waters). Schriftenreihe der TLUG Nr. 99, Jena, 157 S. Retrieved from https://tlubn.thueringen.de/fileadmin/content/wasser/45_handbuch_gewaesserunterhaltung.pdf (last access on 8 February 2023)
UBA (Umweltbundesamt), 2003. Leitbildorientierte physikalisch-chemische Gewässerbewertung – Referenzbedingungen und Qualitätsziele. (Leitbild-oriented physical and chemical freshwater assessment – reference conditions and quality targets.) UBA Texte 15/03, Berlin, 160 S. + Anhang. Retrieved from https://www.umweltbundesamt.de/publikationen/leitbildorientierte-physikalisch-chemische (last access on 7 September 2022)
Wallace, J. B. & J. R. Webster, 1996. The role of macroinvertebrates in stream ecosystem function. Annual Review of Entomology 41: 115–139.
Wang, M., Q. Zheng, Q. Shen & S. Guo, 2013. The critical role of potassium in plant stress response. International Journal of Molecular Sciences 14: 7370–7390.
Wolfram, G., J. Römer, C. Hörl, W. Stockinger, K. Ruzicska & A. Munteanu, 2014. Chlorid-Studie. Auswirkungen von Chlorid auf die aquatische Flora und Fauna, mit besonderer Berücksichtigung der Biologischen Qualitätselemente im Sinne der EU-WRRL (Chlorid study. Implications of chloride on the aquatic flora and fauna with particular emphasis on the biological components acc. to the EU WFD). Bericht Nr. 12/018-B01, Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft Österreich, Wien, 116 S. + Anhang.
Ziemann, H., 1967. Die Wirkung der Kaliabwässer auf die Flora und Fauna der Gewässer unter besonderer Berücksichtigung der Werra und Wipper (Effects of potash brines on the flora and fauna of waters, focusing on the Werra and Wipper rivers). Fortschritt Wasserchemie 7: 50–80.
Acknowledgements
We would like to thank the colleagues at the Thüringer Landesamt für Umwelt, Bergbau und Naturschutz (TLUBN) in Sondershausen, Germany, for assistance during field sampling, lab processing and data generation. The research underlying this article was funded by the Thüringer Landesamt für Umwelt, Bergbau und Naturschutz (TLUBN) Jena, Germany. We are grateful to two anonymous reviewers, whose comments helped improve the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL and the University of Duisburg-Essen. The research leading to the results presented in this article received funding by the Thüringer Landesamt für Umwelt, Bergbau und Naturschutz (TLUBN) Jena, Germany, between January 1st and December 31st, 2017. Two authors (MP and CJS) are employees of the funding agency.
Author information
Authors and Affiliations
Contributions
CKF: conceptualized the study, developed the analytical methodology, statistically analysed the data, lead the manuscript writing and drafted the manuscript. AL: developed the analytical methodology, compiled and statistically analysed the data, contributed to the draft manuscript and further editing of the manuscript. CJS and MP: contributed to the conceptualization of the study, to the manuscript writing and editing. MF: provided and curated the time-series data and contributed to the manuscript writing and editing.
Corresponding author
Ethics declarations
Conflict of interest
All authors declare that they have no competing interests in any material discussed in this article.
Ethical approval
The authors have no ethical conflict to state.
Consent to participate
The research leading to the results presented in this article did not involve animal experiments or humans.
Additional information
Handling editor: Verónica Ferreira
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Feld, C.K., Lorenz, A.W., Peise, M. et al. Direct and indirect effects of salinisation on riverine biota: a case study from river Wipper, Germany. Hydrobiologia 850, 3043–3059 (2023). https://doi.org/10.1007/s10750-023-05229-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-023-05229-z