
Abstract
Cdkn1c, a member of the Cip/Kip cyclin-dependent kinase inhibitor family, is critically involved in regulating cell cycle and cellular differentiation during development in mammals. However, the functional role of Cdkn1c and the underlying mechanisms by which Cdkn1c affects odontogenesis remain largely unknown. In our study, we found that Cdkn1c expression dynamically changes from embryonic day 11.5 (E11.5) to postnatal day 3 (P3), and exhibits tissue-specific expression profiles. Evaluation of CDKN1C protein by immunohistochemistry and western blot, revealed that CDKN1C protein expression peaks at P3 and then is reduced at P5 and P7. Interestingly, we observed that CDKN1C expression is higher in immature odontoblasts than preodontoblasts, is lower in mature odontoblasts, and is practically absent from ameloblasts. We evaluated cell cycle progression to further investigate the mechanisms underlying CDKN1C-mediated regulation of odontogenesis, and found that pRB, cyclin D1 and CDK2 expression decreased from P1 to P3, and reduced at P5 and P7. In addition, we observed increased methylation of KvDMR1 at P1 and P3, and reduced KvDMR1 methylation at P5 and P7. However, the methylation levels of Cdkn1c-sDMR were relatively low from P1 to P7. In summary, we demonstrated that Cdkn1c expression and methylation status may be involved in early postnatal tooth development through regulating the cell cycle inhibition activity of Cdkn1c. Notably, Cdkn1c expression and methylation may associate with cell cycle exit and differentiation of odontoblasts.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Tooth development involves a continuous and reciprocal series of interactions between dental epithelium and mesenchyme (Soukup et al. 2008; Lee et al. 2016); this cycle depends on the tightly regulated spatiotemporal expression of specific genes (Zhang et al. 2009; Zhou et al. 2017; Du et al. 2016). Cell-cycle-associated genes, including genes of the Cip/Kip family of cyclin-dependent kinase inhibitors, are critical mediators and regulators of odontogenesis (Kumamoto et al. 2001). Cip/Kip family genes are known to be important in differentiation of odontoblasts and ameloblasts (Lee et al. 2009; Iwamoto et al. 2017) and facilitate dental mineralization (Yin et al. 2014). However, little is known about the role of Cdkn1c, a recently discovered Cip/Kip family member, in odontogenesis.
p57KIP2, encoded by the Cdkn1c gene, is a major regulator of cell cycle progression (Hildebrand et al. 2012), and is vital in cell proliferation and differentiation (Pateras et al. 2009). p57KIP2 inhibits cyclin/cyclin-dependent kinase (CDK) complexes in the G1 phase, leading to the inhibition of phosphorylation of the retinoblastoma (Rb) protein, and resulting in suppression of E2F family members in the mid-G1 phase (Tury et al. 2012). In addition to its effect on growth arrest, p57 plays distinct functions in the development of several organs (Chung and Park 2016; Mademtzoglou et al. 2017; Stantzou et al. 2017), especially hair follicles (Purba et al. 2017), which share a similar formation process as teeth. Mice deficient for Cdkn1c expression display hyperplasia in several organs, and are not viable (Yan et al. 1997). In humans, abnormal expression of the CDKN1C gene is primarily associated with growth disorder syndromes, including Beckwith–Wiedemann syndrome (Bastaki et al. 2016) and Silver–Russell syndrome (Nakashima et al. 2015). Notably, these diseases are always accompanied by dental abnormalities, such as enamel defects, delayed dental age, and diminished mandibular development (Abeleira et al. 2011; Bergman et al. 2003).
In addition to genetic factors, epigenetics may also affect odontogenesis, as suggested by differences in tooth morphology seen in monozygotic twins (Townsend et al. 2009; Hughes et al. 2014). DNA methylation of gene loci is an epigenetic modification (Deaton and Bird 2011) that has profound and widespread effects on cellular differentiation, genomic imprinting, and tissue-specific gene expression (Suelves et al. 2016; Wu et al. 2016). Su et al. (2016) reported that DNA methylation participates in odontogenesis; they identified 2469 genes that are differentially methylated during pig tooth development from embryonic day 50–60 (E50–E60). Treatment of human dental pulp cells with 5-Aza-2′-deoxycytidine, an inhibitor of DNA methyltransferase activity, decreased cell proliferation and enhanced odontogenic differentiation (Zhang et al. 2015). Moreover, Liu et al. reported that DNA methylation facilitates the osteogenic differentiation of human periodontal ligament stem cells (Liu et al. 2016). The expression of Cdkn1c is also regulated by methylation, and is mainly associated with two differentially methylated regions (DMR) in mice: Cdkn1c-sDMR and KvDMR1 (Beatty et al. 2006; Bhogal et al. 2004). Therefore, investigating the methylation levels of Cdkn1c during odontogenesis may elucidate mechanisms of tooth development.
Cdkn1c gene is a maternally expressed imprinted gene (Matsuoka et al. 1996) and is essential for normal development (Bhogal et al. 2004). The spatiotemporal expression of the imprinted genes Dlk1 and Igf2 coincide with their methylation levels during tooth development (Khan et al. 2012). This suggests that imprinted genes and their methylation levels may be closely involved in odontogenesis. However, the expression and methylation status of Cdkn1c during odontogenesis remain virtually unexplored.
In this study, we characterize the expression of Cdkn1c gene during the early postnatal stages of the mouse first mandibular molar, and we investigate the underlying mechanisms connecting Cdkn1c expression with tooth development.
Materials and methods
Gene expression omnibus (GEO) data acquisition and processing
Cdkn1c gene expression data during tooth development was obtained from datasets available in the NCBI GEO database: GSE32321 (O’Connell et al. 2012), GDS4453 (Lachke et al. 2012), GSE76316 (Pantalacci et al. 2017) and GSE19488 (Sasaki et al. 2010) (http://www.ncbi.nlm.nih.gov/geo/). Cdkn1c mRNA expression z-Score was used for statistical analysis of differential gene expression (Sun et al. 2014).
Animals
Young postnatal (P1, P3, P5, and P7) BALB/c mice were obtained from Hunan SJA Laboratory Animal Co. The Animal Ethics Committee of the Central South University provided the approval for all experimental procedures. Animals were humanely euthanized by cervical dislocation. Mandibles were isolated and immediately fixed overnight in 4% paraformaldehyde (Biosharp, China) buffered with 0.1 M PBS (Zhongshan Goldenbridge Biotechnology, China), pH 7.4, at 4 °C. The first mandibular molar tooth germ was dissected using a stereomicroscope.
Histological and immunohistochemical analysis
After fixation for 24 h, the mandibles were decalcified in 10%, pH 7.4 EDTA for 5–7 days. Mandibles were placed into in a graded ethanol series to dehydrate, and were then embedded in paraffin. Finally, serial sagittal sections were obtained at 4 µm thickness; standard hematoxylin and eosin staining (HE) was employed to examine tissue morphology.
The process of immunohistochemical staining was previously described (Guo et al. 2016). In brief, antigen retrieval was performed by deparaffinizing the sections and incubating the sections with sodium citrate buffer (pH 6.0). Sections were blocked with 3% hydrogen peroxide and subsequently with 5% (v/v) bovine serum albumin (BSA) for 30 min, respectively. Sections were then incubated overnight at 4 °C with a monoclonal antibody to p57KIP2 (#ab75974, 1:1000, Abcam, USA). Sections were then incubated sequentially with agent 1 and agent 2 (#PV-9000-D, Zhongshan Goldenbridge Biotechnology, China). Finally, sections were visualized using a DAB staining kit (#ZLI-9018, Zhongshan Goldenbridge Biotechnology, China). Photomicrographs were obtained for further analysis. PBS was used as a negative control.
Western blot analysis
We isolated 4–20 first mandibular molar tooth germs from each postnatal stage (P1, P3, P5, P7). Total protein was isolated as previously described (Guo et al. 2017; Liu et al. 2018). In brief, total protein lysates were obtained using a lysis buffer composed of 180 µl of RIPA Buffer (Thermo Scientific, USA), 20 µl protease inhibitor cocktail (Bimake, USA), and 2 µl phosphatase inhibitor cocktail (Bimake, USA). Protein concentration was detected using the BCA method. Total protein lysates from each postnatal stage were resolved by SDS-polyacrylamide gel electrophoresis on an 8% w/v gel (Amresco, USA), and were then transferred onto PVDF membranes (Millipore, USA). After transfer, membranes were blocked in 5% fresh skim milk (Yili, China) and incubated overnight at 4 °C in TBST + 5% BSA supplemented with primary antibodies against p57KIP2 (#ab75974, 1:1000, Abcam, USA), Rb (#ab181616, 1:2000, Abcam, USA), Phospho-Rb (Ser780) (#D59B7, 1:1000, Cell Signaling Technology, USA), CDK2 (#78B2, 1:1000, Cell Signaling Technology, USA), Cyclin D1 (#92G2, 1:1000, Cell Signaling Technology, USA), or α-tubulin (#sc-23948, 1: 2000, Santa Cruz, USA). Membranes were then incubated for 1 h at 37 °C with an anti-rabbit peroxidase-conjugated secondary. Protein bands were visualized using an Immobilon western chemiluminescent HRP substrate (Millipore, USA).
Quantification of global DNA methylation
Genomic DNA was extracted from tooth germ (n = 4–10 from each stage) using a DNeasy Blood and Tissue Kit (#69504, Qiagen, Germany). Global methylation levels were evaluated using a MethylFlash™ Methylated DNA Quantification Kit (Colorimetric) (#P-1034, Epigentek, USA). Unmethylated (negative) control DNA (20 ng), a graded series of methylated (positive) control DNA, and purified sample DNA (about 100 ng) were incubated with a bonding solution in strip wells in duplicate. The methylated fraction of DNA was measured with dilute 5-mC capture and detection antibodies. A color developing solution was added and the absorbance at 450 nm was measured using a microplate spectrophotometer. The average of the duplicate measures was used for analyses. A standard curve was using the positive control series and optimal slope was calculated. The amount of methylated DNA was calculated according to the formula provided by the kit manufacturer.
Bisulfite sequencing PCR
Bisulfite conversion of DNA was carried out with the EpiTect Fast DNA Bisulfite Kit (Qiagen, Germany). The methylation status of two differentially methylated regions (Cdkn1c-sDMR and KvDMR1) associated with Cdkn1c imprinting in mice was analyzed in this study. These methylated regions were amplified using the following specific primer pairs, as previously reported (Van de Pette et al. 2017): sDMR-F: 5′-GATTAGTATAATGTAGTATTTTTAGTTT-3′; sDMR-R: 5′-AACTATACCCAACTCCATAATC-3′; KvDMR1-F: 5′-TTAAGGTGAGTGGTTTAGGATA-3′; KvDMR1-R: 5′-AAACCACTATAAACCCAC ACA-3′. PCR products were separated by electrophoresis on a 1.5% agarose gel; bands were recovered with a gel extraction kit (Sangon, China). Extracted fragments were cloned it into T-vectors (Tiangen, China), and transformed into competent bacteria (DH-5a Escherichia coli). DNA from single bacterial colonies was amplified by PCR using vector-specific primers; DNA methylation was analyzed by sequencing the PCR products.
Statistical analysis
All statistical analyses were performed using SPSS 20.0 software (IBM SPSS; Armonk, NY, USA). Data are expressed as means ± standard deviation (SD). Significant differences between two groups were analyzed using an independent t-test; multiple comparisons were performed using the least-significant difference (LSD) test. A p-value of P < 0.05 indicated a statistically significant result.
Results
Cdkn1c expression changes dynamically during molar tooth development
To investigate whether the Cdkn1c gene is involved in tooth development, we analyzed four GEO datasets: GSE32321 (O’Connell et al. 2012), GDS4453 (Lachke et al. 2012), GSE76316 (Pantalacci et al. 2017) and GSE19488 (Sasaki et al. 2010) (Fig. 1). Mouse tooth formation starts morphologically at embryonic day 11.5 (E11.5), and involves an interaction between dental epithelium and mesenchyme at the site of the future tooth; the region progresses through the bud stage, cap stage, bell stage, and secretory stage (O’Connell et al. 2012). The expression levels of Cdkn1c in dental epithelial and mesenchymal tissue began to increase at E11.5–E13.5, and were found to differ from each other at specific stages (Fig. 1a). However, Cdkn1c expression was still significantly lower than that of the whole embryo body without tooth germ at E13.5 (Fig. 1b). To further explore the existence of spatiotemporal expression of Cdkn1c in tooth development, we found the expression levels of molar Cdkn1c constantly changed during E14–E18, and were different between the upper and lower molars (Fig. 1c). After birth, Cdkn1c expression increased (Fig. 1d). These data indicate that Cdkn1c gene may be involved in odontogenesis.

Analysis of Cdkn1c expression in tooth germ from the gene expression omnibus (GEO) database. a Expression levels of Cdkn1c in dental epithelial and mesenchymal tissue at embryonic day 11.5–13.5 from C57BL/6 mice (GSE32321). b Expression levels of Cdkn1c in tooth germ tissue from ICR mice at embryonic day 13.5 and matched whole embryo body without tooth germ (GDS4453). c Expression levels of Cdkn1c in CD1 mouse lower and upper first molar for eight consecutive stages (embryonic day 14.5–18) (GSE76316). d Expression levels of Cdkn1c in ICR mice at pre- and post-natal stages (embryonic day 16 to postnatal day 3) from dental papillae of mandibular first molar tooth germs (GSE19488). E embryonic day, P postnatal day, *P < 0.05
Involvement of p57 in early postnatal tooth development
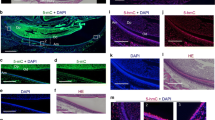
From the HE staining (Fig. 2a–d) we can know that the tooth germ was in the late bell stage at P1 and P3, and paved the way for the morphogenesis of the entire dental structure. At P5 and P7, the first molar entered the secretory stage, and began to produce a large amount of hard tooth tissue. The p57 expression was highly expressed at P1 and P3, and was reduced at P5 and P7 (Fig. 2a1–d1). Almost no positive staining for p57 was observed in ameloblasts from P1 to P7, except for some undifferentiated inner enamel epithelial cells at P1 (Fig. 2a5). p57 was primarily expressed in odontoblasts. Odontoblasts can be classified in three subtypes (Quispe-Salcedo et al. 2012): preodontoblasts, immature odontoblasts, and mature odontoblasts. p57 was intensely expressed in immature odontoblasts (Fig. 2a4–c4, d3), although other pulpal cells also showed weak positive staining. In contrast, p57 expression was greatly reduced in mature odontoblasts (Fig. 2b5, c5, d4), and was almost absent from the mature odontoblasts of the pulp horn at P7 (Fig. 2d5). However, p57 was nearly negative in preodontoblasts (Fig. 2a3–c3). These data suggest that p57 function might be critically required during the early phases of odontoblast differentiation.

Developmental localization of p57 during mouse molar development. Immunohistochemistry for p57 in the lower first molar at postnatal day 1–7 (P1–P7). a–d HE staining, a1–d5 Immunohistochemistry staining. a2–d2 higher magnified views of boxed areas indicated as 2 in a1–d1; the column at the right side is higher magnified views of boxed areas indicated as 2, 3, and 4 in a1–d1 and a2–d2, respectively. The tooth germ was in the late bell stage at P1 and P3, and entered the secretory stage at P5 and P7. p57 protein is intensely expressed in odontoblasts, although other pulpal cells also show weak positive reactions a1–d1. There was almost no positive p57 expression in ameloblasts a2–d2 and preodontoblasts a3–c3. p57 expression was highest in immature odontoblasts a4–c4, d3, and was lowest in mature odontoblasts b5, c5, d4; p57 expression may have already been down-regulated in the mature odontoblasts of the pulp horn at P7 when primary dentin formation is almost completed d5. Note the positive expression in inner enamel epithelium at P1 a5. IEE inner enamel epithelium, DP dental pulp, AB ameloblast, pAB preameloblast, pOB preodontoblast, iOB immature odontoblast, mOB mature odontoblast, EN enamel, P postnatal day. Scale bars = 200 µm in a for a–d, a1–d1, 80 µm in a2 for a2–d2, 10 µm in a3 for a3–d5
p57 participates in tooth development via its activity as a cell cycle inhibitor
To explore the mechanisms underlying the involvement of p57 in tooth development, we performed western blot analysis to evaluate the activation of signaling pathways related to the cell cycle at P1, P3, P5, and P7 (Fig. 3). The expression of p57 was consistent with the results from the IHC staining, increasing from P1 to P3, peaking at P3, then sharply falling at P5 and P7 (Fig. 3a). In a similar fashion to p57, expression levels of cyclin D1, CDK2, and pRB were the lowest at P5 and P7. However, expression patterns of cyclin D1, CDK2, and pRB decreased from P1 to P3 (Fig. 3b–d), suggesting that p57 negatively regulates the expression of CDK2, cyclin D1, and RB phosphorylation from P1 to P3, and is no longer effective from P5 to P7.

Expression of downstream signaling pathways related to cell cycle and p57 during mouse molar development. a Expression levels and relative abundance of p57, cyclin D1, CDK2, RB, and pRB in the mandibular first molar germ at postnatal day 1–7. b–e From postnatal day 1–3, the expression of p57 was increased, while expression of cyclin D1, CDK2, and pRB are decreased. Expression of p57, cyclin D1, CDK2, RB, and pRB was decreased at postnatal day 5 and 7. α-tubulin was used as a control. Data were pooled from three independent experiments with error bars designating standard deviation of the mean. P postnatal day, *P < 0.05
Cdkn1c methylation regulate Cdkn1c expression during mouse tooth development
To analyze whether the DNA methylation is associated with tooth development, we evaluated global DNA methylation levels in mouse first molar germ at P1, P3, P5, and P7. The overall levels of DNA methylation during tooth development were low (Fig. 4a), but there was still a dynamic change at each stage, although the average DNA methylation level showed no significant differences between each stage. This suggests that DNA methylation may be involved in odontogenesis.

Global and Cdkn1c specific DNA methylation analysis during mouse tooth development. a Quantification of 5-mC in genomic DNA isolated from postnatal day 1–7 mouse first molar tooth germ. The data are average values ± standard deviation (SD) from three different assays. The LSD test revealed no statistically significant differences between the two groups. b Schematic diagram of two differentially methylated regions (DMRs) which associate with Cdkn1c imprinting in the mouse. The horizontal bars indicate the positions of the two DMR; the narrow vertical bars represent the position of CpG dinucleotides. For Cdkn1c sDMR, we analyzed 361 bp containing 46 CpGs; the KvDMR1 region contains 341 bp and 17 CpGs. c Bisulfite sequence analysis showing the methylation status of the two DMRs that regulate Cdkn1c imprinted expression (KvDMR1 and Cdkn1c sDMR). Each circle indicates an individual CpG dinucleotide. Each row of circles corresponds to an individual clone of the bisulfite-PCR product. Empty and filled circles indicate unmethylated and methylated CpGs, respectively. The numbers beside the bisulfite-sequencing profiles indicate the percentage of methylated CpG sites. P: postnatal day
Since global DNA methylation level does not reflect the methylation level of a specific gene, we further examined the methylation level of two differentially methylated regions (DMRs) of the Cdkn1c gene (Fig. 4b, c): Cdkn1c-sDMR and KvDMR1. The KvDMR1 region contains 17 CpGs within a 341-bp sequence. KvDMR1 showed dense methylation at P3, but sparse methylation on P5 and P7; this was consistent with the p57 expression levels on those days. Maternal methylation of KvDMR1 suppresses the expression of Kcnq1ot1 and subsequently allows for the expression of Cdkn1c (Fitzpatrick et al. 2002). Together, the methylation levels of KvDMR1 region may play a role in regulating the expression of the Cdkn1c gene during odontogenesis. The Cdkn1c sDMR region is a 361-bp region containing 46 CpGs. The methylation levels of the Cdkn1c sDMR region were relatively low in P1, P3, and P5, and the region was un-methylated in P7. Since the methylation levels at Cdkn1c sDMR are not a precondition for the imprinted expression of Cdkn1c (Bhogal et al. 2004), we concluded that methylation at this region does not participate in tooth development.
Discussion
The present study demonstrated a role for p57 in murine tooth development at early postnatal stages. Data from the GEO database showed that the expression pattern of p57 changes dynamically during different stages of tooth development (Fig. 1). In our experiment, we suggest that p57 and the methylation status of the KvDMR1 region might participate in tooth development, and especially odontoblast differentiation, during the early postnatal stages via the cell cycle inhibitory activity of p57 (Figs. 2, 3, 4).
The Cip/Kip family members are regarded as critical regulators of odontogenesis (Kumamoto et al. 2001), and are prominently implicated in cell cycle regulation as inhibitors of CDKs (Besson et al. 2008). The Cip/Kip family contains three members: p21, p27 and p57. p21 is mainly expressed in the inner enamel epithelium during late cap and in the initial bell stages (Bloch-Zupan et al. 1998). p21 is also regarded as a marker of the early phases of odontoblast differentiation (Nakatomi et al. 2013), while p27 was reported strongly expressed in highly differentiated odontoblasts in humans (Klinz et al. 2013). p57 was reported to play a key role in mammalian development by regulating cell proliferation and differentiation in a number of different tissues (Pateras et al. 2009). Nevertheless, the functional role of p57 in odontogenesis remains unexplored. The GEO data we analyzed showed that the expression of p57 in developing tooth germ was spatiotemporal and tissue-specific (Fig. 1). This finding indicated that p57 might play a distinct role in tooth development.
There are five stages of tooth development: thickening stage, bud stage, cap stage, bell stage, and secretory stage (Tucker and Sharpe 2004; Zhang et al. 2018). In the Balb/c model, first molars at P1 and P3 were at the late bell stage, and at P5 and P7 were at the secretory stage (Fig. 2a–d), similar to a previous report (Lv et al. 2011; Jiang et al. 2017). The bell stage is a crucial stage where epithelial-mesenchymal interactions culminate in dramatic morphological and functional changes (Liu et al. 2017). During the bell stage, undifferentiated mesenchyme and epithelium differentiate into dentin-secreting odontoblasts and enamel-secreting ameloblasts, respectively (Nakasone et al. 2006). Odontoblasts are a monolayer of cells at the periphery of the dental pulp, and are divided into three types: preodontoblasts, immature odontoblastas, and mature odontoblasts (Quispe-Salcedo et al. 2012). Preodontoblasts are the polarized cells beginning to stop their proliferation and acquire differentiation, while mature odontoblasts are matrix-producing cells with their differention almost completed. It is generally thought that there exists an inverse relationship between cell proliferation and differentiation, and p57 is a cell cycle arrest gene and is associated with exit from the cell cycle and entry into differentiation (Rossi et al. 2018). Thus it is explicable that we observed intensely p57 expression in immature odontoblasts, and reduced expression in mature odontoblasts (Fig. 2), similar to the expression pattern of p21 (Nakatomi et al. 2013). This result indicated that p57 could serve as a potential marker of early odontoblast differentiation. However, we observed almost no positive staining of p57 in ameloblasts at any stage from P1 to P7, except for some undifferentiated inner enamel epithelium cells at P1 (Fig. 2a5), in contrast to the expression pattern of p21 (Bloch-Zupan et al. 1998). This result suggested that p57 may not participate in the regulation of ameloblast differentiation.
p57 regulates cell differentiation primarily by inhibition of the cell cycle (Martinez et al. 1999). For example, p57 participated in cell cycle exit and differentiation of an oligodendrocyte precursor (Dugas et al. 2007). In addition, during human hair follicle matrix differentiation, p57 is expressed in post-mitotic keratinocytes, accompanying down-regulation of cyclin A and B1 (Purba et al. 2017). In our study, we also observed that p57 expression was negatively correlated with pRB, cyclin D1, and CDK2 during the late bell stage (Fig. 3). However, we discovered both p57 and activity of downstream pathways related to cell cycle were relatively low at P5 and P7. This indicated that p57 might play no significant role in regulating tooth development at P5 and P7.
DNA methylation also plays a vital role in tooth development (Wan et al. 2018; Yoshioka et al. 2015). Our data, however, demonstrate that global methylation levels were relatively low in the tooth germ at P1–P7 (Fig. 4a). In concordance with our results, the global methylation levels were also quite low in the deciduous tooth germ in miniature pigs at the bell stage (E50) and secretory stage (E60); however, many differentially methylated genes were identified (Su et al. 2016). Cdkn1c lies within a complex imprinted domain, and is regulated by an imprinting center, KvDMR1, in both humans and mice. Methylation at this locus inhibits the expression of Kcnq1ot1 but allows the expression of Cdkn1c (Eggermann et al. 2014). The absence or loss of methylation at this locus results in Beckwith-Wiedemann syndrome leading to growth disorders (Bastaki et al. 2016; Singh et al. 2017). In addition, Cdkn1c is also regulated by the DMR of Cdkn1c in mice, which is located at the promoter and gene body (Bhogal et al. 2004). Methylation at this locus leads to inhibition of Cdkn1c expression. In the present study, we discovered that differential methylation of KvDMR1, not Cdkn1c sDMR, regulates the expression of Cdkn1c in tooth development (Fig. 4c). Similar expression patterns were also found in the liver, spleen, and lung of cattle (Wang et al. 2015). Since the methylation levels at the Cdkn1c sDMR are not a precondition for its imprinted expression, but likely are essential to maintain its imprinting (Bhogal et al. 2004), we suggest that methylation of the KvDMR1 region, but not the Cdkn1c sDMR region, is involved in tooth development.
In conclusion, the present study sheds light on the role of p57 in the early postnatal stage of tooth development. We report a detailed observation of the spatiotemporal expression of p57 in tooth germ, and we explore the potential molecular mechanisms by which it governs tooth development. We also examine the methylation status of the two DMR regions of Cdkn1c, and we demonstrate that Cdkn1c plays a crucial role in odontoblast differentiation. However, since the direct molecular mechanisms and functional analysis did not investigate in our studies, the elucidation of a potential role of Cdkn1c in tooth development, especially in odontoblasts, requires further more complex studies.
References
Abeleira MT, Seoane-Romero JM, Outumuro M, Caamaño F, Suárez D, Carmona IT (2011) A multidisciplinary approach to the treatment of oral manifestations associated with Beckwith-Wiedemann syndrome. J Am Dent Assoc 142(12):1357–1364. https://doi.org/10.14219/jada.archive.2011.0136
Bastaki F, Saif F, Al Ali MT, Hamzeh AR (2016) Molecular and clinical characterization of a nonsense CDKN1C mutation in an Emirati patient with Beckwith-Wiedemann syndrome. Saudi Med J 37(2):215–216. https://doi.org/10.15537/smj.2016.2.13593
Beatty L, Weksberg R, Sadowski PD (2006) Detailed analysis of the methylation patterns of the KvDMR1 imprinting control region of human chromosome 11. Genomics 87(1):46–56. https://doi.org/10.1016/j.ygeno.2005.05.015
Bergman A, Kjellberg H, Dahlgren J (2003) Craniofacial morphology and dental age in children with Silver-Russell syndrome. Orthod Craniofac Res 6(1):54–62
Besson A, Dowdy SF, Roberts JM (2008) CDK inhibitors: cell cycle regulators and beyond. Dev Cell 14(2):159–169. https://doi.org/10.1016/j.devcel.2008.01.013
Bhogal B, Arnaudo A, Dymkowski A, Best A, Davis TL (2004) Methylation at mouse Cdkn1c is acquired during postimplantation development and functions to maintain imprinted expression. Genomics 84(6):961–970. https://doi.org/10.1016/j.ygeno.2004.08.004
Bloch-Zupan A, Leveillard T, Gorry P, Fausser JL, Ruch JV (1998) Expression of p21(WAF1/CIP1) during mouse odontogenesis. Eur J Oral Sci 106(Suppl 1):104–111
Chung H, Park S (2016) Ghrelin regulates cell cycle-related gene expression in cultured hippocampal neural stem cells. J Endocrinol 230(2):239–250. https://doi.org/10.1530/joe-16-0126
Deaton AM, Bird A (2011) CpG islands and the regulation of transcription. Genes Dev 25(10):1010–1022. https://doi.org/10.1101/gad.2037511
Du J, Wang Q, Yang P, Wang X (2016) FHL2 mediates tooth development and human dental pulp cell differentiation into odontoblasts, partially by interacting with Runx2. J Mol Histol 47(2):195–202. https://doi.org/10.1007/s10735-016-9655-6
Dugas JC, Ibrahim A, Barres BA (2007) A crucial role for p57(Kip2) in the intracellular timer that controls oligodendrocyte differentiation. J Neurosci 27(23):6185–6196. https://doi.org/10.1523/JNEUROSCI.0628-07.2007
Eggermann T, Binder G, Brioude F, Maher ER, Lapunzina P, Cubellis MV, Bergada I, Prawitt D, Begemann M (2014) CDKN1C mutations: two sides of the same coin. Trends Mol Med 20(11):614–622. https://doi.org/10.1016/j.molmed.2014.09.001
Fitzpatrick GV, Soloway PD, Higgins MJ (2002) Regional loss of imprinting and growth deficiency in mice with a targeted deletion of KvDMR1. Nat Genet 32(3):426–431. https://doi.org/10.1038/ng988
Guo Y, Tang CY, Man XF, Tang HN, Tang J, Wang F, Zhou CL, Tan SW, Feng YZ, Zhou HD (2016) Insulin receptor substrate-1 time-dependently regulates bone formation by controlling collagen I 2 expression via miR-342. FASEB J 30(12):4214–4226. https://doi.org/10.1096/fj.201600445RR
Guo Y, Tang CY, Man XF, Tang HN, Tang J, Zhou CL, Tan SW, Wang M, Feng YZ, Zhou HD (2017) Insulin-like growth factor-1 promotes osteogenic differentiation and collagen I alpha 2 synthesis via induction of mRNA-binding protein LARP6 expression. Dev Growth Differ 59(2):94–103. https://doi.org/10.1111/dgd.12342
Hildebrand J, Sporl F, Korge S, Gallinat S, Wenck H, Deppert W, Knott A, Blatt T (2012) Cell cycle regulator Cdkn1c (p57/KIP2) shows distinct expression in epidermal differentiation. Eur J Dermatol 22(5):694–696. https://doi.org/10.1684/ejd.2012.1828
Hughes TE, Townsend GC, Pinkerton SK, Bockmann MR, Seow WK, Brook AH, Richards LC, Mihailidis S, Ranjitkar S, Lekkas D (2014) The teeth and faces of twins: providing insights into dentofacial development and oral health for practising oral health professionals. Aust Dent J 59(Suppl 1):101–116. https://doi.org/10.1111/adj.12101
Iwamoto T, Nakamura T, Ishikawa M, Yoshizaki K, Sugimoto A, Ida-Yonemochi H, Ohshima H, Saito M, Yamada Y, Fukumoto S (2017) Pannexin 3 regulates proliferation and differentiation of odontoblasts via its hemichannel activities. PLoS ONE 12(5):e0177557. https://doi.org/10.1371/journal.pone.0177557
Jiang T, Liu F, Wang WG, Jiang X, Wen X, Hu KJ, Xue Y (2017) Distribution of cathepsin K in late stage of tooth germ development and its function in degrading enamel matrix proteins in mouse. PLoS ONE 12(1):e0169857. https://doi.org/10.1371/journal.pone.0169857
Khan QE, Sehic A, Skalleberg N, Landin MA, Khuu C, Risnes S, Osmundsen H (2012) Expression of delta-like 1 homologue and insulin-like growth factor 2 through epigenetic regulation of the genes during development of mouse molar. Eur J Oral Sci 120(4):292–302. https://doi.org/10.1111/j.1600-0722.2012.00976.x
Klinz FJ, Korkmaz Y, Cho B, Raab WH, Addicks K (2013) Nuclear expression of p27(Kip1) is associated with in vivo differentiation of adult human odontoblasts. J Endod 39(8):995–1000. https://doi.org/10.1016/j.joen.2013.04.026
Kumamoto H, Kimi K, Ooya K (2001) Detection of cell cycle-related factors in ameloblastomas. J Oral Pathol Med 30(5):309–315
Lachke SA, Ho JW, Kryukov GV, O’Connell DJ, Aboukhalil A, Bulyk ML, Park PJ, Maas RL (2012) iSyTE: integrated systems tool for eye gene discovery. Investig Ophthalmol Vis Sci 53(3):1617–1627. https://doi.org/10.1167/iovs.11-8839
Lee D-S, Park J-T, Kim H-M, Ko JS, Son H-H, Gronostajski RM, Cho M-I, Choung P-H, Park J-C (2009) Nuclear factor I-C is essential for odontogenic cell proliferation and odontoblast differentiation during tooth root development. J Biol Chem 284(25):17293–17303. https://doi.org/10.1074/jbc.M109.009084
Lee HK, Park JW, Seo YM, Kim HH, Lee G, Bae HS, Park JC (2016) Odontoblastic inductive potential of epithelial cells derived from human deciduous dental pulp. J Mol Histol 47(3):345–351. https://doi.org/10.1007/s10735-016-9676-1
Liu Z, Chen T, Sun W, Yuan Z, Yu M, Chen G, Guo W, Xiao J, Tian W (2016) DNA demethylation rescues the impaired osteogenic differentiation ability of human periodontal ligament stem cells in high glucose. Sci Rep 6:27447. https://doi.org/10.1038/srep27447
Liu L, Chen W, Li L, Xu F, Jiang B (2017) Inhibition of chondroitin sulfate glycosaminoglycans incorporation affected odontoblast differentiation in cultured embryonic mouse molars. J Mol Histol 48(5–6):337–345. https://doi.org/10.1007/s10735-017-9732-5
Liu J, Qian L, Guo L, Feng Y (2018) Studying hepcidin and related pathways in osteoblasts using a mouse model with insulin receptor substrate 1loss of function. Mol Med Rep 17(1):350–357. https://doi.org/10.3892/mmr.2017.7876
Lv H, Fu S, Wu G, Yan F (2011) PHEX neutralizing agent inhibits dentin formation in mouse tooth germ. Tissue Cell 43(2):125–130. https://doi.org/10.1016/j.tice.2010.12.008
Mademtzoglou D, Alonso-Martin S, Chang TH, Bismuth K, Drayton-Libotte B, Aurade F, Relaix F (2017) A p57 conditional mutant allele that allows tracking of p57-expressing cells. Genesis. https://doi.org/10.1002/dvg.23025
Martinez LA, Chen Y, Fischer SM, Conti CJ (1999) Coordinated changes in cell cycle machinery occur during keratinocyte terminal differentiation. Oncogene 18(2):397–406. https://doi.org/10.1038/sj.onc.1202300
Matsuoka S, Thompson JS, Edwards MC, Bartletta JM, Grundy P, Kalikin LM, Harper JW, Elledge SJ, Feinberg AP (1996) Imprinting of the gene encoding a human cyclin-dependent kinase inhibitor, p57KIP2, on chromosome 11p15. Proc Natl Acad Sci USA 93(7):3026–3030
Nakashima S, Kato F, Kosho T, Nagasaki K, Kikuchi T, Kagami M, Fukami M, Ogata T (2015) Silver-Russell syndrome without body asymmetry in three patients with duplications of maternally derived chromosome 11p15 involving CDKN1C. J Hum Genet 60(2):91–95. https://doi.org/10.1038/jhg.2014.100
Nakasone N, Yoshie H, Ohshima H (2006) An immunohistochemical study of the expression of heat-shock protein-25 and cell proliferation in the dental pulp and enamel organ during odontogenesis in rat molars. Arch Oral Biol 51(5):378–386. https://doi.org/10.1016/j.archoralbio.2005.09.007
Nakatomi M, Ida-Yonemochi H, Ohshima H (2013) Lymphoid enhancer-binding factor 1 expression precedes dentin sialophosphoprotein expression during rat odontoblast differentiation and regeneration. J Endod 39(5):612–618. https://doi.org/10.1016/j.joen.2012.12.016
O’Connell DJ, Ho JW, Mammoto T, Turbe-Doan A, O’Connell JT, Haseley PS, Koo S, Kamiya N, Ingber DE, Park PJ, Maas RL (2012) A Wnt-bmp feedback circuit controls intertissue signaling dynamics in tooth organogenesis. Sci Signal 5(206):ra4. https://doi.org/10.1126/scisignal.2002414
Pantalacci S, Gueguen L, Petit C, Lambert A, Peterkova R, Semon M (2017) Transcriptomic signatures shaped by cell proportions shed light on comparative developmental biology. Genome Biol 18(1):29. https://doi.org/10.1186/s13059-017-1157-7
Pateras IS, Apostolopoulou K, Niforou K, Kotsinas A, Gorgoulis VG (2009) p57KIP2: “Kip"ing the cell under control. Mol Cancer Res 7(12):1902–1919. https://doi.org/10.1158/1541-7786.mcr-09-0317
Purba TS, Brunken L, Peake M, Shahmalak A, Chaves A, Poblet E, Ceballos L, Gandarillas A, Paus R (2017) Characterisation of cell cycle arrest and terminal differentiation in a maximally proliferative human epithelial tissue: lessons from the human hair follicle matrix. Eur J Cell Biol 96(6):632–641. https://doi.org/10.1016/j.ejcb.2017.03.011
Quispe-Salcedo A, Ida-Yonemochi H, Nakatomi M, Ohshima H (2012) Expression patterns of nestin and dentin sialoprotein during dentinogenesis in mice. Biomed Res 33(2):119–132
Rossi MN, Andresini O, Matteini F, Maione R (2018) Transcriptional regulation of p57(kip2) expression during development, differentiation and disease. Front Biosci (Landmark Ed) 23:83–108
Sasaki H, Muramatsu T, Kwon HJ, Yamamoto H, Hashimoto S, Jung HS, Shimono M (2010) Down-regulated genes in mouse dental papillae and pulp. J Dent Res 89(7):679–683. https://doi.org/10.1177/0022034510366844
Singh VB, Sribenja S, Wilson KE, Attwood KM, Hillman JC, Pathak S, Higgins MJ (2017) Blocked transcription through KvDMR1 results in absence of methylation and gene silencing resembling Beckwith-Wiedemann syndrome. Development 144(10):1820–1830. https://doi.org/10.1242/dev.145136
Soukup V, Epperlein HH, Horacek I, Cerny R (2008) Dual epithelial origin of vertebrate oral teeth. Nature 455(7214):795–798. https://doi.org/10.1038/nature07304
Stantzou A, Schirwis E, Swist S, Alonso-Martin S, Polydorou I, Zarrouki F, Mouisel E, Beley C, Julien A, Le Grand F, Garcia L, Colnot C, Birchmeier C, Braun T, Schuelke M, Relaix F, Amthor H (2017) BMP signaling regulates satellite cell-dependent postnatal muscle growth. Development 144(15):2737–2747. https://doi.org/10.1242/dev.144089
Su Y, Fan Z, Wu X, Li Y, Wang F, Zhang C, Wang J, Du J, Wang S (2016) Genome-wide DNA methylation profile of developing deciduous tooth germ in miniature pigs. BMC Genom 17:134. https://doi.org/10.1186/s12864-016-2485-9
Suelves M, Carrio E, Nunez-Alvarez Y, Peinado MA (2016) DNA methylation dynamics in cellular commitment and differentiation. Brief Funct Genom 15(6):443–453. https://doi.org/10.1093/bfgp/elw017
Sun Y, Gu X, Zhang E, Park MA, Pereira AM, Wang S, Morrison T, Li C, Blenis J, Gerbaudo VH, Henske EP, Yu JJ (2014) Estradiol promotes pentose phosphate pathway addiction and cell survival via reactivation of Akt in mTORC1 hyperactive cells. Cell Death Dis 5:e1231. https://doi.org/10.1038/cddis.2014.204
Townsend G, Hughes T, Luciano M, Bockmann M, Brook A (2009) Genetic and environmental influences on human dental variation: a critical evaluation of studies involving twins. Arch Oral Biol 54(Suppl 1):S45–S51. https://doi.org/10.1016/j.archoralbio.2008.06.009
Tucker A, Sharpe P (2004) The cutting-edge of mammalian development; how the embryo makes teeth. Nat Rev Genet 5(7):499–508. https://doi.org/10.1038/nrg1380
Tury A, Mairet-Coello G, DiCicco-Bloom E (2012) The multiple roles of the cyclin-dependent kinase inhibitory protein p57(KIP2) in cerebral cortical neurogenesis. Dev Neurobiol 72(6):821–842. https://doi.org/10.1002/dneu.20999
Van de Pette M, Abbas A, Feytout A, McNamara G, Bruno L, To WK, Dimond A, Sardini A, Webster Z, McGinty J, Paul EJ, Ungless MA, French PM, Withers DJ, Uren A, Ferguson-Smith AC, Merkenschlager M, John RM, Fisher AG (2017) Visualizing changes in Cdkn1c expression links early-life adversity to imprint mis-regulation in adults. Cell Rep 18(5):1090–1099. https://doi.org/10.1016/j.celrep.2017.01.010
Wan M, Li H, Zhou Y, Du W, Xu X, Ye L, Zhou X, Zheng L (2018) DNA methylation: a frontier in tooth organogenesis and developmental dental defects. Curr Stem Cell Res Ther 13(2):151–158. https://doi.org/10.2174/1574888X13666171206124940
Wang M, Li D, Zhang M, Yang W, Cui Y, Li S (2015) Methylation of KvDMR1 involved in regulating the imprinting of CDKN1C gene in cattle. Anim Genet 46(4):354–360. https://doi.org/10.1111/age.12297
Wu H, Zhao M, Tan L, Lu Q (2016) The key culprit in the pathogenesis of systemic lupus erythematosus: aberrant DNA methylation. Autoimmun Rev 15(7):684–689. https://doi.org/10.1016/j.autrev.2016.03.002
Yan Y, Frisen J, Lee MH, Massague J, Barbacid M (1997) Ablation of the CDK inhibitor p57Kip2 results in increased apoptosis and delayed differentiation during mouse development. Genes Dev 11(8):973–983
Yin Y, Wang Q, Sun W, Wang Y, Chen N, Miao D (2014) p27(kip1) deficiency accelerates dentin and alveolar bone formation. Clin Exp Pharmacol Physiol 41(10):807–816. https://doi.org/10.1111/1440-1681.12276
Yoshioka H, Minamizaki T, Yoshiko Y (2015) The dynamics of DNA methylation and hydroxymethylation during amelogenesis. Histochem Cell Biol 144(5):471–478. https://doi.org/10.1007/s00418-015-1353-z
Zhang Z, Lan Y, Chai Y, Jiang R (2009) Antagonistic actions of Msx1 and Osr2 pattern mammalian teeth into a single row. Science 323(5918):1232–1234. https://doi.org/10.1126/science.1167418
Zhang D, Li Q, Rao L, Yi B, Xu Q (2015) Effect of 5-Aza-2′-deoxycytidine on odontogenic differentiation of human dental pulp cells. J Endod 41(5):640–645. https://doi.org/10.1016/j.joen.2014.12.006
Zhang F, Jiang L, He Y, Fan W, Guan X, Deng Q, Huang F, He H (2018) Changes of mitochondrial respiratory function during odontogenic differentiation of rat dental papilla cells. J Mol Histol 49(1):51–61. https://doi.org/10.1007/s10735-017-9746-z
Zhou M, Guo S, Yuan L, Zhang Y, Zhang M, Chen H, Lu M, Yang J, Ma J (2017) Blockade of LGR4 inhibits proliferation and odonto/osteogenic differentiation of stem cells from apical papillae. J Mol Histol 48(5–6):389–401. https://doi.org/10.1007/s10735-017-9737-0
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant Number 81773339).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest with the contents of this article.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Li, Q., Guo, Y., Yao, M. et al. Methylation of Cdkn1c may be involved in the regulation of tooth development through cell cycle inhibition. J Mol Hist 49, 459–469 (2018). https://doi.org/10.1007/s10735-018-9785-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10735-018-9785-0