
Abstract
The straying of hatchery salmon may harm wild salmon populations through a variety of ecological and genetic mechanisms. Surveys of pink (Oncorhynchus gorbuscha), chum (O. keta) and sockeye (O. nerka) salmon in wild salmon spawning locations in Prince William Sound (PWS), Alaska since 1997 show a wide range of hatchery straying. The analysis of thermally marked otoliths collected from carcasses indicate that 0–98% of pink salmon, 0–63% of chum salmon and 0–93% of sockeye salmon in spawning areas are hatchery fish, producing an unknown number of hatchery-wild hybrids. Most spawning locations sampled (77%) had hatchery pink salmon from three or more hatcheries, and 51% had annual escapements consisting of more than 10% hatchery pink salmon during at least one of the years surveyed. An exponential decay model of the percentage of hatchery pink salmon strays with distance from hatcheries indicated that streams throughout PWS contain more than 10% hatchery pink salmon. The prevalence of hatchery pink salmon strays in streams increased throughout the spawning season, while the prevalence of hatchery chum salmon decreased. The level of hatchery salmon strays in many areas of PWS are beyond all proposed thresholds (2–10%), which confounds wild salmon escapement goals and may harm the productivity, genetic diversity and fitness of wild salmon in this region
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Pacific salmon (Oncorhynchus spp.) are noted for their abilities to home to natal areas to spawn. This homing isolates populations from one another and often leads to adaptations to the environmental conditions of a particular watershed (e.g., Quinn 1993, 2005; Eliason et al. 2011). Homing is not perfect, however, and some wild fish stray into non-natal spawning areas. These background levels of natural straying may be adaptive by allowing populations to expand into new habitats and by supplementing populations with low genetic diversities (Quinn 2005). The straying of hatchery-reared salmon, on the other hand, may be disruptive to wild populations. A few generations of artificial breeding and rearing of salmon in a protected environment can lead to domestication, altered gene frequencies, and phenotypic changes that reduce the adaptive fitness of hatchery salmon (Fleming and Gross 1993; Berejikian et al. 2001; Myers et al. 2004; Mobrand et al. 2005; Araki et al. 2007; Naish et al. 2007; Araki et al. 2008; Grant 2012). Hence, hybridizations between hatchery and wild fish may also decrease the adaptive fitness of wild populations (McClelland et al. 2005; Ford et al. 2006; Wessel et al. 2006).
The focus of the present study is on estimating the extent that hatchery fish stray into wild spawning areas in Prince William Sound (PWS), Alaska. Previous studies of straying, based on coded-wire tags, indicate that increases in the hatchery production of pink salmon (Oncorhynchus gorbuscha) in the late 1980s resulted in large numbers of hatchery salmon in wild salmon streams (Sharp et al. 1994; Sharr et al. 1995). Prompted by these results, surveys of fish in 14 wild-spawning streams in 1997 for thermally marked otoliths (Volk et al. 2005) indicated that 26–97% of the pink salmon sampled were of hatchery origin (Joyce and Evans 1999). Some streams located more than 40 km from a hatchery release site contained more than 50% hatchery pink salmon. After hatchery production increased, surveys conducted by fix-winged aircraft indicated that escapement of chum salmon from the fishery into spawning areas in one district increased from an average of less than 200 fish (1974–1994) to more than 11,000 fish (1995–2007; Lewis et al. 2008). Observations at weirs also indicated an influx of chum salmon in streams where few had previously been observed.
Hatcheries in PWS presently release about 643 million pink, 146 million chum, and 8.5 million juvenile sockeye salmon into PWS each year (White 2007). This will increase to about 690 million pink, 162 million chum and 10.5 million juvenile sockeye salmon within the next few years. About 33.8 million pink, 2.6 million chum, and 0.7 million sockeye salmon of hatchery origin return to PWS annually (Ashe et al. 2005a, b; Hollowell et al. 2007). Most hatchery salmon are harvested in commercial fisheries from May to September.
Several threshold levels for straying or introgression have been proposed or can be implied from the literature. Based on reported straying of less than 2.2% for pink salmon (Boyd 1964; Blair 1968), the PWS Comprehensive Salmon Management Plan (PWS CRRPT 1994) sets a limit on the proportion of hatchery fish in PWS streams at 2%. From a genetic perspective, additional threshold percentages of hatchery strays in wild spawning systems have been suggested, including less than 5% (Mobrand et al. 2005). Additionally, a quantitative genetics model by Ford (2002) indicated that over a wide range of model parameters, exceeding 10% hatchery strays with introgression into a wild stock may produce a significant loss of fitness.
The goal of this study was to investigate the extent that hatchery fish are straying into wild-population spawning areas in PWS by estimating: 1) proportions of hatchery pink, chum, and sockeye salmon in historically important wild-salmon spawning areas, and 2) spatial and temporal extent of hatchery pink and chum salmon straying. Data generated from this study will be used to determine the degree to which hatchery strays may have contributed to the overestimation of wild salmon escapement estimates. We used thermally marked otoliths to identify hatchery salmon and compare results and modeling simulations to the proposed straying threshold levels of 2%, 5%, and 10%.
Methods
Study sites
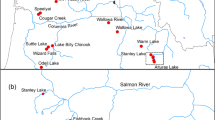
Prince William Sound, Alaska is about 8,800 km2 in size (Fig. 1) and has more than 1,000 streams with spawning salmon, mostly pink and chum salmon. Alaska Department of Fish and Game (ADFG) biologists surveyed 214 of these streams using fixed-wing aircraft approximately weekly during the spawning season (July–October). Collectively, these aerial index streams represent an estimated 75–85% of the wild pink salmon spawning escapement in PWS (Fried et al. 1998) and are believed to encompass most of the spawning chum and sockeye salmon systems (S. Moffitt, ADFG, pers. obs.). Many streams drain into PWS from steep basins so that spawning areas for pink and chum salmon are limited to the lower reaches of the streams or to intertidal areas (Bailey 1964; Helle 1970).

Commercial fishing districts (221–228) and release locations of hatchery pink, chum, and sockeye salmon in PWS, Alaska. District names are as follows: 221 = Eastern, 222 = Northern, 223 = Coghill, 224 = Northwestern, 225 = Eshamy, 226 = Southwestern, 227 = Montague, 228 = Southeastern, 229 = Unakwik District (not shown). Hatcheries and release locations correspond to the following numbers: 1 = Solomon Gulch Hatchery (SGH), 2 = Cannery Creek Hatchery (CCH), 3 = Wally Noerenberg Hatchery (WNH), 4 = Main Bay Hatchery (MBH), 5 = Armin F. Koernig Hatchery (AFK), 6 = Port Chalmers (PC) remote release location, 7 = Marsha Lake (MBH sockeye smolt release), 8 = Solf Lake (MBH sockeye smolt release)
Otoliths from pink salmon carcasses were sampled at 14 aerial index streams in 1997 (Joyce and Evans 1999), 25 streams in 1998, and 33 streams in 1999 (Table 1). In 1997, sampling was largely confined to southwestern PWS. In 1998, streams were sampled throughout PWS. In 1999, most samples came from streams in the Eastern and Northern Districts.
For the 2008–2010 hatchery pink salmon straying study, we used a stratified random design to select pink salmon streams for sampling to account for the distance from hatchery release sites (Fig. 1). We selected 10 circular distance bins around three pink salmon hatcheries: Cannery Creek (CCH), Wally Noerenberg (WNH), and Armin F. Koernig (AFK) to a distance of 44 km from each hatchery (straight-line distance). Distance bins were held to 44 km because this is about half the distance between the AFK and WNH hatcheries (Fig. 1). Samples collected within a distance bin also enabled us to assess straying at greater than 44 km from all other release facilities. We included aerial index streams in our selection process if they had annual average escapements greater than 10,000 pink salmon. Escapement was determined by area-under-the-curve of observed counts from weekly aerial surveys, adjusted for estimated stream life and observer efficiency (Fried et al. 1998). When a distance bin did not contain a stream with minimal escapement criteria, we randomly selected a stream from an adjacent distance bin. Additional streams were sampled for pink salmon carcasses during sampling for chum salmon (see below), by weir crews, and during visits to streams by state and federal biologists. Distance bins were not selected around the Solomon Gulch (SGH; Fig. 1) hatchery because flying restrictions have prevented aerial surveys around this facility during recent years. However, we did sample streams near SGH and throughout the Eastern District.
Streams for the chum salmon straying project were selected within 10 km distance bins around the two release sites (WNH and Port Chalmers; Fig. 1), if their annual average peak escapements indices were greater than 1,000 chum salmon. Chum salmon were also sampled at weirs and during visits to streams for other research projects. Otolith sampling of chum salmon in streams began with a 13-stream pilot study in 2004 and expanded to more than 20 streams sampled annually from 2005 to present.
Sockeye salmon carcasses were sampled in lakes and streams where sockeye salmon spawn, at weirs on Eshamy and Coghill Rivers, and during visits to streams for the pink and chum salmon straying projects.
Identification of hatchery fish
All hatchery salmon released into PWS have thermally marked otoliths. Pink and sockeye salmon thermal marks are specific to release locations; however, chum salmon with the same mark have been released at multiple locations as recently as 2007 (Lewis et al. 2008). Fish with hatchery marks were considered hatchery “strays” if they died naturally at a spawning location prior to otolith sampling. Otoliths collected at the Eshamy and Coghill weirs (streams 511 and 322, respectively) were generally, but not always, from sacrificed fish. These fish are reported as hatchery proportions within streams, but not as “strays”. All other otoliths were dissected from dead fish in the stream or stream bank and placed in a labeled tray. Both sagittal otoliths were collected, whenever possible, and the sex of the fish and GPS coordinate were noted (WGS 84). Otoliths were subsequently polished and inspected under a light microscope for the presence of hatchery thermal marks (Volk et al. 2005) by trained personnel who were tested with randomized blind tests of known origin fish to assess accuracy (Joyce and Evans 1999).
Sample size
During 1997–1999, otoliths from 48 pink salmon carcasses were sampled twice a week for up to 4 weeks (384 total; Joyce and Evans 1999). For the 2004–2008 chum salmon straying project, the goal was to sample 96 carcasses three times during the spawning season. Power analysis indicated that 128 otoliths from three samplings (384 total) were needed to estimate the percentage of hatchery salmon in a stream within 5% of the true percentage, with 95% confidence (Thompson 1992). Straying proportions and total population size for many streams were known; however, because they were highly variable we used the worst case sample size of 384, which assumes an infinite population size and a proportion of hatchery fish of P = 0.5. Thus, in 2008 we attempted to collect otoliths from 128 pink, chum, and sockeye salmon carcasses in a stream during the early, middle, and late portions of the spawning season. We used historical escapement timing and aerial surveys to determine when sample collections should take place for each stream. For the pink and chum salmon studies, we excluded from our analyses streams with sample sizes less than 50 carcasses.
Straying percentages
The percentage of hatchery pink salmon sampled in a stream was weighted by estimates of escapement to the stream. For non-weir sites, pink salmon escapement was determined by adjusting aerial survey observations for average observer efficiency (0.44) and by stream-specific estimates of stream life for pink salmon in PWS (Fried et al. 1998). Linear interpolation was used to estimate escapement between aerial observations, and cumulative escapement was calculated using an area-under-the-curve method. We subtracted one-half of stream duration (days) from our otolith sampling date and used the escapement estimate at that date to weight straying proportions, because we could not determine when sampled fish had died. Exact binomial confidence intervals (95%) were calculated for each sample. For streams with more than one sampling strata in a year, the overall confidence intervals were calculated by weighting with the strata escapement.
Pink salmon modeling
First-order exponential decay models were fit to the 2008–2010 data with Origin 8.1 (OriginLab Corporation) to characterize distance from hatcheries and percentage of pink-salmon strays within streams. Percentages of hatchery strays (y) were modeled for each facility (SGH, CCH, WNH, and AFK) and year (2008–2010) using
where y 0 = y offset (greater than or equal to 0%), A = amplitude (less than or equal to 100%), x = shortest water distance (km) between a hatchery release location and a sampled stream, and t = the decay constant. AIC was used to select the initial model (logistic or exponential decay) (Burnham and Anderson 2002). Distances were estimated with ArcGIS 9.3 as the shortest water distance between coordinates representing the mouth of a stream (Johnson and Klein 2009) and hatchery release sites. We used known proportions of hatchery salmon collected from brood stock to anchor the model, because we could not always sample streams within the nearest distance bin to hatchery release locations (Joyce and Evans 1999; ADFG unpublished). Convergence on the best model fit occurred after 10–500 iterations to minimize chi-square using the Levenberg-Marquardt algorithm (OriginLab Corporation). The fitted model for each release location and year was solved for the distance at which streams would contain specific percentages of hatchery salmon strays (e.g., 2%, 5%, 10%, etc.) from individual hatcheries (n = 4 models per year). Graphical representations of these percentages were plotted in ArcGIS 9.3. We also used model results to estimate the number of aerial index streams with these percentages of pink salmon in their escapement. Given the inconsistency of stream sampling during 1997–1999, we did not construct model results for these years.
Results
Pink salmon
Results from 1997 are published in Joyce and Evans (1999), but results for 1998 and 1999 have not previously been published (Table 1). Across these years, streams closest to release facilities generally contained the highest proportions of hatchery pink salmon strays (Joyce and Evans 1999). However, the Eshamy District (Fig. 1, 225), which does not contain a pink salmon hatchery, also had streams with relatively high levels of hatchery strays (14–88%; Table 1). In 1997, streams in the Southwestern District (Fig. 1, 226) contained 26–86% hatchery strays, while streams in the Northern (Fig. 1, 222) and Eastern (Fig. 1, 221) districts had 40–97% hatchery strays. In 1998, sampled streams contained 0–98% hatchery strays. In 1999, streams in Eastern and Northern district had 0–38% hatchery strays, with remaining districts containing 6–22% hatchery strays. The proportion of hatchery strays in streams increased throughout the spawning season, except for hatchery strays from SGH (Joyce and Evans 1999).
For the present study (2008–2010), complete samples could not be obtained from all streams during all 3 years (Table 2) due to small escapements and storm events that flushed carcasses out of intertidal areas. Of the 43 streams sampled, complete samples were obtained from 25 streams during 3 years and 10 streams during 2 years, while eight streams were sampled opportunistically during a single year as part of other research projects such as chum salmon straying or weir projects. Proportions of hatchery pink salmon within streams varied widely across districts and within individual streams during 2008–2010 (Table 2). Twenty-two (51%) streams contained more than 10% hatchery strays, and 15 (35%) streams had more than 20% hatchery strays in at least 1 year (Table 2). At the same time, 21 (49%) sampled streams contained less than 5% hatchery strays, and 17 (40%) streams had less than 2% hatchery strays during at least 1 year.
Streams within 20 km of hatcheries generally contained the highest proportions of hatchery pink salmon strays; however, strays were also frequently found in streams outside the districts containing the release facility. In fact, 77% of sampled streams contained hatchery pink salmon from at least three release locations, mostly from AFK, CCH, and WNH; while 86% of streams contained strays from at least two facilities. Straying outside districts was particularly evident for streams in the Eshamy District, which contained high proportions of pink salmon from WNH and AFK hatcheries and overall escapement estimates consisted of 5.5–88% hatchery strays (Tables 2 and 3). Strays in Northern District streams were mostly from CCH and WNH, where escapements consisted of 0–70% hatchery pink salmon (Tables 2 and 3).
Similar to the Joyce and Evans (1999) study, streams throughout the Southwestern District contained some of the highest percentages of hatchery pink salmon strays from 2008 to 2010 (2–89%; Table 2), which were mostly from AFK (2–74% of total escapement in individual streams). Significant proportions of hatchery strays were also from CCH and WNH, which contributed 1–7% and 2–14% to total stream escapement, respectively (Table 3).
In the Eastern District pink salmon escapement consisted of 0–98% hatchery strays, mostly from SGH (Tables 2 and 3). Few pink salmon strays from SGH were found in streams outside the Eastern District. More than 4% of the pink salmon sampled in three Eastern District streams were from CCH, with small proportions from WNH and AFK (Table 3).
Annual pink salmon escapement into streams sampled in the Coghill (Fig. 1, 223) and Northwestern (Fig. 1, 224) districts consisted of 0–28% hatchery pink salmon strays, mostly from WNH (Tables 2 and 3). Hatchery pink salmon represented 2–7% of the annual escapement into streams in the Montague District (Fig. 1, 227), and most were from AFK (Tables 2 and 3).
Hatchery pink salmon returning to CCH, WNH and AFK facilities arrive relatively late in the spawning season, and most strays from these facilities were found within streams after Julian day 230 in mid August (Fig. 2). Streams generally contained fewer than 10% strays prior to this date but after this date strays constituted as much as 93% of the fish sampled. Hatchery pink salmon return to SGH comparatively early in the summer spawning season and strays from this facility were most prevalent in streams prior to Julian day 245.

Percentages of hatchery pink salmon collected within PWS streams over time that originated from (a) SGH and (b) from a combination of AFK, CCH and WNH. Pink salmon data are combined from 2008–2010. Panel (c) is the percentage of hatchery chum salmon over time within PWS streams, pooled for 2004–2010
Chum salmon
From 2004 to 2010 chum salmon were sampled at 25 streams for which ADFG has long-term records of chum salmon spawning (Table 4). The proportion of hatchery chum salmon strays within streams varied greatly across years (0–63%; Table 4). The highest percentages of hatchery strays were in streams 264 (5–63%) and 214 (3–31%) in the Northern District, and stream 117 in the Eastern District (2–19%). Neither of these districts contains a chum salmon hatchery or remote release locations. In contrast, streams in the Coghill and Northwestern districts, which are substantially closer to WNH, generally had lower proportions of hatchery strays. We could not determine how straying proportions related to distance from release facility, since fish with the same thermal mark were released at multiple locations (AFK, Port Chalmers, and WNH).
Twelve of the historically important chum salmon streams sampled had more than 5% hatchery strays during at least 1 year, and 12 streams had no hatchery strays during at least 1 year. Samples from Southwestern and Montague districts, which were not historically important spawning locations, had 91–100% hatchery chum salmon. Samples of live fish from weirs contained 93% (Coghill River) and 98–100% (Eshamy River) hatchery chum salmon.
The total percentage of hatchery chum salmon in individual streams decreased during the course of the summer (Fig. 2). Before Julian day 240 (late August), as many as 100% of the fish in some streams were hatchery chum salmon, but no stream had more than about 20% hatchery chum salmon after this date.
Sockeye salmon
Due to the scant availability of sockeye salmon carcasses around many lakes, consistent sampling was limited to the weir sites on the Coghill and Eshamy rivers, where more than 150 sockeye salmon were sampled each year from streams 322 (Coghill River) and 511 (Eshamy River) (Table 5). For these locations, only Eshamy River had levels of hatchery sockeye salmon exceeding the 2% threshold and this was confined to 2007 (22% unweighted by escapement, 33% weighted). The Coghill River sample had less than 1% hatchery sockeye salmon across all years. Other historically important sockeye salmon spawning sites had 0–93% hatchery sockeye salmon but we were only able to sample small numbers of carcasses (n = 8–45) from four systems with some of the highest percentages of hatchery sockeye salmon. For streams without documented populations of wild sockeye salmon, 29 of 44 (66%) sockeye salmon carcasses sampled were of hatchery origin.
Pink salmon modeling
Similar to empirical results, model output showed high levels of hatchery pink salmon strays in streams far from the AFK hatchery facility and lower levels of hatchery strays surrounding the other release facilities (Fig. 3a–c; Table 6). Model fits were generally better for hatchery strays originating from CCH and SGH than for WNH and AFK (Table 6). Exponential decay model fits predict that 24–29% of the aerial index streams in PWS will have more than 10% hatchery pink salmon and that 39–55% of aerial index streams will have more than 5% hatchery pink salmon. For all years, model fits predicted that the percentage of hatchery pink salmon from CCH and/or WNH will be above the 2% level in stream for all distances from these facilities. While some streams in the Southwestern District contained more than 2% strays from these facilities, streams within the Eastern, Northwestern, and Montague districts had less than 2% hatchery pink salmon from these facilities. The chosen model underestimated straying from CCH, WNH, and AFK within Eshamy District streams but sometimes overestimated the level of straying to streams on Montague Island.


Predicted percentages of hatchery pink salmon within PWS streams at various distances (km) from hatcheries during (a) 2008 (b) 2009 and (c) 2010. Output is from first order exponential decay model runs for each facility (SGH, CCH, WNH, AFK) by year. Distances are the nearest water distances between the hatcheries and mouths of streams
Discussion
The results of this study show large amounts of hatchery salmon straying in some areas in PWS. Although we were unable to sample every stream each year of the study, the extensive geographic coverage of pink salmon spawning streams sampled allowed us to produce a model of the straying intensities around four hatcheries. We have some confidence in the results of this model, because samples were collected throughout the spawning season of multiple years to be able to detect seasonal and inter-annual variability in hatchery straying. The results of the current study, when combined with previous examinations of straying in PWS, provide a large database (more than 70,000 fish sampled since 1997) for describing straying behavior and for understanding the effects of hatchery strays on the management and health of wild salmon populations.
What are safe levels of hatchery salmon straying?
Low levels of natural straying among wild populations of salmon may be advantageous to the persistence of salmon populations in an area (Quinn 2005). However, an extensive body of literature suggests that hatchery rearing practices can result in changes to hatchery salmon including altered mating behavior, domestication and reduced fitness (Berejikian et al. 2001; Metcalfe et al. 2003; Campton 2004; Wessel et al. 2006; Araki et al. 2008; Williamson et al. 2011). From a genetic point of view, there are two concerns over interbreeding between wild and stray hatchery salmon. One is that interbreeding can decrease the fitness of wild fish. For example, hatchery techniques can shift the timing of spawning (Quinn et al. 2002), such that progeny from hatchery-wild mating do not spawn at an optimal time for reproduction (Ford et al. 2006). A second concern is that hatchery straying into wild populations can reduce genetic diversity among wild salmon populations. Even low, but persistent, rates of hybridization and introgression could alter the genetic diversity and traits of wild salmon populations (Ryman and Laikre 1991; Ryman et al. 1995; Ford 2002). This reduces the potential for the portfolio effect to enhance the persistence of a regional group of relatively distinct populations (Schindler et al. 2010).
The amount of straying that a wild population can tolerate is uncertain. The guiding management plan for hatcheries in PWS suggests an upper limit of hatchery strays of 2% (PWS CRRPT 1994), while other studies suggest that thresholds as high as 5% or 10% may be detrimental to wild salmon populations through mechanisms discussed above (Ford 2002; Mobrand et al. 2005). In our study, we found a wide range of straying proportions across years, but hatchery pink, chum, and sockeye salmon exceeded all proposed thresholds of straying into many wild salmon spawning locations. For hatchery pink salmon in particular, results and modeling simulations indicated that threshold levels were exceeded for large numbers of streams throughout PWS. Pink salmon are thought to have the highest natural straying rates of any Pacific salmon (Quinn 2005). However, relative to other species of salmon, the short generation time (2 years) and large number of hatchery pink salmon released may accelerate the genetic impacts to wild pink salmon in PWS (e.g., Ryman and Laikre 1991; Ford 2002). While stream proportions of hatchery pink salmon strays from individual hatcheries declined exponentially with distance from a given release facility, most streams received hatchery strays from two or three hatcheries. Corrective actions to lower straying proportions below proposed thresholds need to account for the overlapping zones of influence from multiple release sites. In addition, large hatchery salmon returns may make it difficult to reduce straying below threshold levels, even if only a fraction of the hatchery population strays.
Regardless of whether hatchery and wild fish interbreed, hatchery strays may negatively impact wild salmon by intensifying competitive interactions between adults on or near spawning locations (Essington et al. 2000). Stress and competition associated with crowding in streams can induce egg retention or mortality prior to spawning (Quinn et al. 2007). Spawning pink and chum salmon within PWS are often confined to a limited intertidal area and may be particularly vulnerable to density-dependent effects. The increase of stray hatchery pink salmon throughout the spawning season may increase the potential for hatchery strays to destroy redds from earlier spawning wild fish (Bailey 1964; McNeil 1964; Fukushima et al. 1998; Quinn 2005). Hatchery pink salmon strays may also displace earlier arriving wild fish on productive spawning sites. The magnitude of impacts to wild salmon from ecological interactions with their hatchery counterparts likely depends on the timing and density of wild and hatchery salmon. In the current study (2008–2010), many streams had greater than 20% hatchery pink salmon strays after Julian day 240, and 13 of 14 streams sampled by Joyce and Evans (1999) had greater than 50% hatchery strays near the end of the spawning season. The considerable spatial and temporal overlap of wild and hatchery pink salmon in spawning locations leads to concern that ecological interactions alone could impair the productivity of wild salmon in PWS.
Drivers of straying
The regional, yearly, and hatchery-specific differences in straying in our study suggest potential drivers of straying. Pink salmon spawn in diverse habitats extending from intertidal to upstream freshwater areas and juveniles migrate directly to marine waters soon after emerging from the gravel. As much as 70% of PWS pink salmon spawn in intertidal areas (Bailey 1964; Helle 1970). These intertidal populations show less genetic differentiation among one another than do upstream populations, indicating greater amounts of natural straying and gene flow (Seeb et al. 1999). Since homing ability is influenced by both genetic and environmental factors (Quinn 1984, 2005), the selection of broodstock for hatchery production from a particular population may determine how well hatchery-reared fish home to the hatchery. We hypothesize that some of the donor populations used for broodstock in PWS pink salmon hatcheries have an inherently low fidelity to natal spawning sites. The genetic and environmental components of homing ability could be influenced by three factors: 1) the distance of the donor wild population from a release site (Wertheimer et al. 1994), 2) the use of predominately intertidal spawning fish in donor stocks (Seeb et al. 1999), and 3) the intermingling of wild and hatchery fish during early marine and adult life stages.
These factors may explain why hatchery pink salmon stray more from some facilities (AFK, CCH, and WNH) than from others (SGH). If the genetic component of straying in pink salmon is large, then fish from some hatchery broodstocks potentially have greater tendencies to stray. Relatively few hatchery pink salmon strays from SGH were found in PWS streams (Table 3). Streams within 30 km of SGH contained fewer hatchery strays from this facility than from CCH, which is 90 km away. The broodstock for SGH originated from nearby streams, while broodstock for the other facilities was created from a combination of local and non-local sources (Habicht et al. 2000). We do not have evidence that SGH broodstock contained a greater “upstream” component compared to other facilities; however, this, in combination with the use of more localized broodstock for SGH, may be a factor in their greater apparent homing fidelity. Pink salmon from SGH also have earlier run timing than other hatchery and wild stocks in PWS and may not intermingle with fish homing to other areas. In contrast, the run timing of hatchery pink salmon with the highest proportions of strays (AFK, CCH, and WNH) overlaps with many wild pinks salmon stocks, resulting in abundant opportunities for schooling and straying.
Actions by managers and commercial fishermen may also contribute to straying. Reduced fishing effort in the Eshamy District and management concerns for wild sockeye salmon escapement to the Eshamy River could have contributed to the higher level of hatchery sockeye strays in 2007 (Table 5). Commercial fishing in the Eshamy District was restricted for much of the 2007 season to allow escapement of wild sockeye salmon into the Eshamy River. Large returns of salmon to the Copper River District in 2007 also resulted in about 100 fewer commercial fishing permits harvesting fish in the Eshamy District compared to 2008–2010 (Lewis et al. 2008). High proportions of hatchery chum salmon within several fishing districts in 2007 (Table 4) also followed a lack of fishing pressure. Thus, fisheries managers face the dilemma of restricting fisheries to allow wild salmon run entry while simultaneously maintaining effort on hatchery salmon to reduce straying.
Management implications
The immediate management impact of hatchery salmon straying is the inflation of wild salmon escapement estimates. Counts from aerial surveys and weirs are the primary tool available for ADFG biologists to evaluate sustainable escapement goals (SEG) of wild salmon in PWS (Bue et al. 2002). Commercial fishing periods are often coupled to the achievement of these goals, which have not always been met in recent years, even prior to accounting for hatchery strays (e.g., Botz et al. 2008). Currently, all fish observed in streams during aerial surveys and at weirs are counted as being of wild origin. Our results confirm that this is not a valid assumption and that any models developed to evaluate the extent to which wild salmon escapement goals are, or have been, achieved must account for spatial and temporal trends of hatchery salmon straying. It is unknown whether hatchery-origin fish contribute to wild population recruitment in PWS, but some studies indicate that the productivity of salmon populations can decline substantially when mature hatchery fish are present in streams (Buhle et al. 2009; Chilcote et al. 2011). Therefore, hatchery strays make it difficult for fisheries managers to evaluate whether wild salmon escapement goals are being met and could make it difficult to link spawning escapement and recruitment.
Conclusions
The straying of hatchery salmon into wild stock streams may harm the health and productivity of wild salmon through ecological and genetic mechanisms, and can hinder management through errors in estimating wild salmon escapement and recruitment potential. Wild salmon will likely experience variable amounts of introgression with hatchery strays due to genetic and life history differences (Utter 2000, 2003) and temporal patterns of straying. Thus, the use of a single threshold proportion of hatchery strays in a stream is unlikely to be useful from a genetic perspective. The consequences of genetic (e.g., loss of genetic diversity and fitness) and ecological (e.g., competition by adults and juveniles in marine and freshwater environments) interactions are unknown. Research into these topics should be of considerable importance for sustaining wild populations in PWS and other regions with large hatchery programs, and a precautionary approach (e.g., Pearsons 2008) should be used to manage hatchery salmon production.
References
Araki H, Cooper B, Blouin MS (2007) Genetic effects of captive breeding cause a rapid, cumulative decline in the wild. Science 318:100–103
Araki H, Berejikian BA, Ford MJ, Blouin MS (2008) Fitness of hatchery-reared salmonids in the wild. Evol Apps 1:342–355. doi:10.1111/j.1752-4571.2008.00026.x
Ashe D, Gray D, Lewis B, Merizon R, Moffitt S (2005a) Prince William Sound management area 2003 annual finfish management report. Fishery management report no. 05-54. Alaska Department of Fish and Game, Anchorage
Ashe D, Gray D, Lewis B, Merizon R, Moffitt S (2005b) Prince William Sound management area 2004 annual finfish management report. Fishery management report no. 05-65. Alaska Department of Fish and Game, Anchorage
Bailey JE (1964) Intertidal spawning of pink and chum salmon at Olsen Bay, Prince William Sound, Alaska. U.S Fish and Wildlife Service Manuscript Report MR 64-6
Berejikian BA, Tezak EP, Park L, LaHood E, Schroder SL, Beall E (2001) Male competition and breeding success in captively reared and wild Coho salmon (Oncorhynchus kisutch). Can J Fish Aquat Sci 58:804–810
Blair AA (1968) Pink salmon find new home in Newfoundland. Fish Can 21:9–12
Botz J, Brenner R, Hollowell G, Lewis B, Moffitt S (2008) 2006 Prince William Sound area finfish management report. Fishery management report no. 08-30. Alaska Department of Fish and Game, Anchorage
Boyd FC (1964) Return of pink salmon to Robertson Creek shows promise of success. Can Fish Cult 32:59–62
Bue B, Hasbrouck J, Evenson M (2002) Escapement goal review of Copper and Bering Rivers, and Prince William Sound Pacific salmon stocks, report to the Alaska Board of Fisheries. Regional information report no 2A02-35. Alaska Department of Fish and Game, Commercial Fisheries Division, Anchorage
Buhle ER, Holsman KK, Schaeuerell MD, Albaugh A (2009) Using an unplanned experiment to evaluate the effects of hatcheries and environmental variation on threatened populations of wild salmon. Biol Cons 142:2449–2455
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Campton DE (2004) Sperm competition in salmon hatcheries: the need to institutionalize genetically benign spawning protocols. Trans Am Fish Soc 133:1277–1289
Chilcote MW, Goodson KW, Falcy MR (2011) Reduced recruitment performance in natural populations of anadromous salmonids associated with hatchery-reared fish. Can J Fish Aquat Sci 68:511–522
Eliason EJ, Clark RD, Hague MH, Hanson LM, Gallagher ZS, Jeffries KM, Gale MK, Patterson DA, Hinch SG, Farrell AP (2011) Differences in thermal tolerance among sockeye salmon populations. Sci 332:109–112
Essington TE, Quinn TP, Ewert VE (2000) Intra- and inter-specific competition and the reproductive success of sympatric pacific salmon. Can J Fish Aquat Sci 57:205–213
Fleming IA, Gross MR (1993) Breeding success of hatchery and wild Coho salmon (Oncorhynchus kisutch) in competition. Ecol Appl 3:230–245
Ford MJ (2002) Selection in captivity during supportive breeding may reduce fitness in the wild. Cons Biol 16:815–825
Ford MJ, Fuss H, Boelts B, LaHood E, Hard J, Miller J (2006) Changes in run timing and natural smolt production in a naturally spawning Coho salmon (Oncorhynchus kisutch) population after 60 years of intensive hatchery supplementation. Can J Fish Aquat Sci 63:2343–2355. doi:10.1139/F06-119
Fried SM, Bue BG, Sharp D, Sharr S (1998) Injury to spawning areas and an evaluation of spawning escapement enumeration of pink salmon in Prince William Sound, Alaska, Exxon Valdez Oil Spill State/Federal Natural Resource Damage Assessment Final Report (Fish/Shellfish Study Number 1, Restoration Study Number 9, and Restoration Study Number 60B), Alaska Department of Fish and Game, Division of Commercial Fisheries, Anchorage, Alaska
Fukushima M, Quinn TJ, Smoker WW (1998) Estimation of eggs lost from superimposed pink salmon (Oncorhynchus gorbuscha) redds. Can J Fish Aquat Sci 55:618–625
Grant WS (2012) Understanding the adaptive consequences of ecological interactions between hatchery and wild salmon in Alaska. Environ Biol Fish (this issue)
Habicht C, Simpson EM, Seeb JE (2000) Broodstock acquisition and release sites for hatcheries producing pink salmon in Prince William Sound. Regional information report no. 5J00-07. Alaska Department of Fish and Game, Division of Commercial Fisheries, Anchorage
Helle JH (1970) Biological characteristics of intertidal and fresh-water spawning pink salmon at Olsen Creek, Prince William Sound, Alaska, 1963–1963. U.S. Fish and Wildlife Service Special Scientific Report-Fisheries 602
Hollowell G, Lewis B, Merizon R, Moffitt S (2007) 2005 Prince William Sound area finfish management report. Fishery management report no 07-33. Alaska Department of Fish and Game, Division of Sport Fish and Commercial Fisheries, Anchorage
Johnson J, Klein K (2009) Catalog of waters important for spawning, rearing, or migration of anadromous fishes–Southcentral region, effective June 1, 2009. Special Publication No. 09-03. Anchorage, Alaska
Joyce TL, Evans DG (1999) Otolith marking of pink salmon in Prince William Sound salmon hatcheries, Exxon Valdez oil spill restoration final report (Restoration Project 99188) Alaska Department of Fish and Game, Division of Commercial Fisheries, Cordova and Anchorage, Alaska
Lewis B, Botz J, Brenner R, Hollowell G, Moffitt S (2008) 2007 Prince William Sound area finfish management report. Alaska Department of Fish and Game, Fishery management Report No. 08-53, Anchorage, Alaska
McClelland EK, Myers JM, Hard JJ, Park LK, Naish KA (2005) Two generations of outbreeding in Coho salmon (Oncorhynchus kisutch): effects on size and growth. Can J Fish Aquat Sci 62:2538–2547
McNeil WJ (1964) Redd superimposition and egg capacity of pink salmon spawning beds. J Fish Res Board Can 21:1385–1396
Metcalfe NB, Valdimarsson SK, Morgan IJ (2003) The relative roles of domestication, rearing environment, prior residence and body size in deciding territorial contests between hatchery and wild juvenile salmon. J Appl Ecol 40:535–544
Mobrand LE, Barr J, Blankenship L, Campton DE, Evelyn TTP, Flagg TA, Mahnken CVW, Seeb LW, Seidel PR, Smoker WW (2005) Hatchery reform in Washington State: principles and emerging issues. Fisheries 30:11–39
Myers RA, Levin SA, Lande R, James FC, Murdoch WW, Paine RT (2004) Hatcheries and endangered salmon. Science 303:1980
Naish KA, Taylor JE III, Levin PS, Quinn TP, Winton JR, Huppert D, Hilborn R (2007) An evaluation of the effects of conservation and fishery enhancement hatcheries on wild populations of salmon. Adv Mar Biol 53:61–194. doi:10.1016/S0065-2881(07)53002-6
Pearsons TN (2008) Misconception, reality, and uncertainty about ecological interactions and risks between hatchery and wild salmonids. Fisheries 33:278–290
PWS CRRPT (1994) Prince William Sound-Copper River phase 3 comprehensive salmon plan. Prince William Sound–Copper River Regional Planning Team. Alaska Department of Fish and Game, Division of Commercial Fisheries, Cordova and Anchorage
Quinn TP (1984) Homing and straying in Pacific salmon. In: McCleave JD, Arnold GP, Dodson JJ, Neill WH (eds) Mechanisms of migration in fishes. Plenum, New York, pp 357–362
Quinn TP (1993) A review of homing and straying of wild and hatchery-produced salmon. Fish Res 18:29–44
Quinn TP (2005) The behavior and ecology of Pacific salmon and trout. American Fisheries Society, Bethesda, pp 85–104
Quinn TP, Peterson JA, Gallucci VF, Hershberger WK, Brannon EL (2002) Artificial selection and environmental change: countervailing factors affecting the timing of spawning by Coho and Chinook salmon. Trans Am Fish Soc 131:591–598
Quinn TP, Eggers DM, Clark JH, Rich HB Jr (2007) Density, climate, and the processes of prespawning mortality and egg retention in pacific salmon (Oncorhynchus spp.). Can J Fish Aquat Sci 64:574–582. doi:10.1139/F07-035
Ryman N, Laikre L (1991) Effects of supportive breeding on the genetically effective population size. Cons Biol 5:325–329
Ryman N, Jorde PE, Laikre L (1995) Supportive breeding and variance effective population size. Cons Biol 9:1619–1628
Schindler DE, Hilborn R, Chasco B, Boatright CP, Quinn TP, Rogers LA, Webster MS (2010) Population diversity and the portfolio effect in an exploited species. Nature 465:609–612
Seeb JE, Habicht C, Templin WD, Seeb LW, Shaklee JB, Utter FM (1999) Allozyme and mitochondrial DNA variation describe ecologically important genetic structure of even-year pink salmon inhabiting Prince William Sound, Alaska. Ecol Freshwat Fish 8:122–140
Sharp D, Sharr S, Peckham C (1994) Homing and straying patterns of coded wire tagged pink salmon in Prince William Sound. Proceedings of the 16th Northeast Pacific Pink and Chum Salmon Workshop. Juneau, AK
Sharr S, Peckham CJ, Sharp DG, Peltz L, Smith JL, Willette TM, Evans DG, Bue BG (1995) Coded wire tag studies on Prince William Sound salmon, 1989–1991: Fish/Shellfish Study Number 3, Final Report. Exxon Valdez Oil Spill State/Federal Natural Resource Damage Assessment Final Report: 1–52
Thompson SK (1992) Sampling. Wiley, New York
Utter F (2000) Patterns of subspecific anthropogenic introgression in two salmonid genera. Rev Fish Biol Fish 3:1–15
Utter F (2003) Genetic impacts of fish introductions. In: Hallerman EM (ed) Population genetics: principles and applications for fisheries scientists. American Fisheries Society, Bethesda, pp 357–378
Volk EC, Schroder SL, Grimm JJ (2005) Otolith thermal marking. In: Cadrin SX, Friedman KD, Waldman JR (eds) Stock identification methods. Elsevier, pp 447–463
Wertheimer AC, Heintz RA, Thedinga JF, Maselko JM, Rice SD (1994) Straying of adult pink salmon from their natal streams following embryonic exposure to weathered Exxon Valdez crude oil. Trans Am Fish Soc 129:989–1004
Wessel ML, Smoker WW, Fagen RM, Joyce J (2006) Variation of agonistic behavior among juvenile Chinook salmon (Oncorhynchus tshawytscha) of hatchery, hybrid, and wild origin. Can J Fish Aquat Sci 63:438–447
White B (2007) Fishery management report no. 07-04. Alaska Salmon Enhancement Program 2006 Annual Report. Alaska Department of Fish and Game, Division of Sport Fish and Commercial Fisheries. http://www.sf.adfg.state.ak.us/FedAidPDFs/fmr07-04.pdf
Williamson KS, Murdoch AR, Pearsons TN, Ward EJ, Ford MJ (2011) Factors influencing the relative fitness of hatchery and wild spring Chinook salmon (Oncorhynchus tshawytscha) in the Wenatchee River, Washington, USA. Can J Fish Aquat Sci 67:1840–1851
Acknowledgments
We thank three anonymous reviewers for many helpful comments. R. Merizon developed the chum salmon straying evaluation project. We are grateful for the assistance of J. Allen, D. Anderson, B. Behrends, R. Brewer, B.J. Canfield, K. Froning, T. Jewell, K. Jurica, E. Mejia-Rosa, J. O’Rourke, M. O’Rourke, K. Williams and others for sample collection and analysis. A. Bidlack and J. Syder provided the GIS analysis and associated graphics. X. Zhang assisted with statistical analyses. We also thank the pilots and personnel of Cordova Air and Alaskan Wilderness Air for transportation to field sites. The 1997–1999 Joyce and Evans pink salmon straying project was funded by the Exxon Valdez Oil Spill Trustees Council and ADFG. Some air charter funds during 1997 and 1998 were provided by PWSAC. The current studies (2004–2010) were funded by the Alaska Sustainable Salmon Fund and ADFG. This is contribution PP-271 of ADFG, Commercial Fisheries Division.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Table 1
Nearest water distances (km) from sampled locations to pink and chum salmon hatcheries in PWS, Alaska. Districts names are 221 (Eastern), 222 (Northern) 225 (Eshamy), 226 (Southwestern) 227 (Montague,) 228 (Southeastern). Hatcheries are SGH (Solomon Gulch) CCH (Cannery Creek), WNH (Wally Noerenberg) and AFK (Armin F. Koernig) hatcheries. (DOC 117 kb)
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Brenner, R.E., Moffitt, S.D. & Grant, W.S. Straying of hatchery salmon in Prince William Sound, Alaska. Environ Biol Fish 94, 179–195 (2012). https://doi.org/10.1007/s10641-012-9975-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-012-9975-7