
Abstract
Nursery habitats are larval or juvenile habitats that disproportionately contribute individuals to adult populations of a species. Identifying and protecting such habitats is important to species conservation, yet evaluating the relative contributions of different larval habitats to adult fish populations has proven difficult at best. Otolith geochemistry is one available tool for reconstructing previous habitat use of adult fishes during the early life history, thus facilitating the identification of nursery habitats. In this study, we compared traditional catch surveys of larval-stage longfin smelt (Spirinchus thaleichthys) occurring in habitats of different salinities to corresponding larval-stage salinity distributions of sub-adult/adult longfin smelt estimated using otolith geochemical techniques. This allowed us to evaluate the relative contribution of larvae from waters of various salinities to sub-adult/adult populations of longfin smelt. We used laser ablation MC-ICP-MS on otoliths and an empirically-derived relationship between strontium isotope ratios (87Sr/86Sr) of waters across the estuarine salinity gradient to reconstruct the larval salinity history of longfin smelt. Salinity values from the larval region of sub-adult/adult otoliths (corresponding to standard lengths of ca.10-mm) were compared to corresponding catch distribution of larval longfin smelt (≤ 10-mm) from 4 year-classes (1999, 2000, 2003 and 2006) in the San Francisco Estuary spanning a period when the population underwent a dramatic decline. Though the catch distribution of larval-stage longfin smelt was centered around 4-ppt and did not vary significantly among years, salinity distributions of sub-adult/adult were lower and narrower (ca. 2-ppt), suggesting that low-salinity habitats disproportionally contributed more recruits relative to both freshwater and brackish water habitats and, therefore, may function as important nursery areas. Furthermore, the relative importance of the low salinity zone (ca. 2-ppt) to successful recruitment appeared greatest in years following the longfin smelt population decline. Our results indicate that otolith strontium isotopes (87Sr/86Sr) are a powerful tool for identifying nursery habitats for estuarine fishes.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The Longfin Smelt (Spirinchus thaleichthys) was once one of the most abundant fishes in San Francisco Bay and Humboldt Bay and even supported a commercial bait fishery until the early 1970’s (Moyle 2002). Like several pelagic fishes of the upper San Francisco Estuary, the Longfin Smelt experienced a marked decline since 2001, which contributed to its listing as “threatened” under the State Endangered Species Act Footnote 1(Baxter et al. 2007; Sommer et al. 2007; California Fish and Game Commission 2009). Many factors have been associated with this “pelagic organism decline” (POD) including increased freshwater diversions, decreased low-salinity habitat, reduced prey abundance, and frequent toxic algal blooms. All of these impacts occur within larval habitats of the Longfin Smelt to various degrees (Armor et al. 2005; Feyrer et al. 2007b; Sommer et al. 2007; Lehman et al. 2008; Kimmerer et al. 2009). To better understand the potential consequences of these impacts to Longfin Smelt population dynamics, we first needed to develop a method to evaluate which larval habitat(s) contribute the most individuals to adult populations.
The Longfin Smelt are a short-lived species living only 2 to 3 years. Mature Longfin Smelt migrate upstream toward the confluence of the Sacramento and San Joaquin Rivers in winter (Dec-Feb) to spawn in freshwater and are thought to be benthic spawners, attaching their adhesive eggs to sandy substrates (Moulton 1974; Moyle 2002; Rosenfield and Baxter 2007). The larvae are pelagic and often found in surface waters at sizes up to 10-mm total length (TL). Upon swim bladder inflation (> 10-mm TL), the fish can vertically migrate and move deeper in the water column and occur in brackish habitats of Suisun and San Pablo Bay (Bennett et al. 2002; Hobbs et al. 2006). Longfin Smelt larvae (here we focus on those ≤ 10-mm TL) can be found in a wide range of salinities (0–20-ppt), but a majority of the catch occurs at salinities below 10-ppt, with the greatest densities occurring around 2-ppt, defined in this estuary as the “Low Salinity Zone” (Kimmerer et al. 2009). The location of this low salinity zone varies based on the magnitude of freshwater flowing into the upper estuary, shifting downstream in response to increased freshwater flows (Jassby et al. 1995) and the locations of Longfin Smelt larvae shift with it (Dege and Brown 2004). Thus, Longfin Smelt larval habitat can best be described by salinity rather than geography. With such a broad larval distribution, different factors associated with the POD may affect different segments of the population. For example, freshwater exports would impact fish living in freshwater habitats while reduced prey abundances in the low salinity zone would likely only affect fish rearing in those waters.
Previous research has used the concentrations of trace elements (e.g., strontium, barium and magnesium) in otoliths to infer the natal origins, stock structure and salinity history of fishes (Thorrold et al. 1998; Campana et al. 2000; Gillanders 2005; Hobbs et al. 2007). However, the incorporation of trace elements into otoliths from surrounding waters can be physiologically controlled and otolith trace element concentrations can vary significantly from that of the surrounding waters in response to variations in temperature, salinity and their effects on trace element incorporation into the otolith (Campana 1999). As a result, trace element concentrations may not accurately reflect environmental history for species that primarily use estuaries as nursery areas due to the high variability in temperature and salinity characteristic of these habitats (Elsdon et al. 2008). Though studies have found that Sr/Ca ratios in otoliths provided reasonable salinity proxies for reconstructing salinity history of some migratory fishes, the resolution of this technique is too coarse for discriminating among waters of low salinity (e.g., 0–5-ppt; Kalish 1989; Secor et al. 1995; Secor and Rooker 2000). In contrast, strontium isotope ratios (87Sr/ 86Sr) are not physiologically regulated and do not appear to fractionate when incorporated into fish otoliths; therefore, these ratios directly reflect the chemical signatures of the surrounding waters (Kennedy et al. 2000). Due to their conservative mixing properties, strontium isotope ratios (87Sr/ 86Sr) may provide a better proxy of salinity for fish living in low salinity, oligohaline regions of estuaries (Ingram and DePaolo 1993; Faure and Mensing 2005).
Otolith strontium isotope ratios (87Sr/86Sr) have been successfully used to determine natal origins of freshwater fishes from around the world including Barramundi (Lates calcarifer) and galaxiid Smelt in Australia (Woodhead et al. 2005; Milton et al. 2008), Dolly Varden (Salvelinus malma) in Alaska (Outridge et al. 2002), and Atlantic Salmon (Salmo salar) in Vermont (Kennedy et al. 2000). In California’s Central Valley, strontium isotope ratios (87Sr/86Sr) have been used to distinguish natal origins of Chinook Salmon (Oncorhynchus tshawytscha) (Barnett-Johnson et al. 2008), Sacramento Splittail (Pogonichthys macrolepidotus) (Feyrer et al. 2007a) and the endangered delta smelt (Hypomesus transpacificus) (Hobbs et al. 2005). Investigation of the spatial variability of strontium isotope ratios (87Sr/86Sr) in the central valley watershed and San Francisco Estuary resulted in an “isoscape,” (i.e., detailed map) of strontium isotope ratios (87Sr/86Sr) in this system, and evidence indicating the largest freshwater water sources to the upper estuary is distinctive in their strontium isotope ratios (87Sr/86Sr) (Ingram and Weber 1999; Barnett-Johnson et al. 2008). For example, consistently higher strontium isotope ratios (87Sr/86Sr; e.g., 0.707) occur in the San Joaquin River basin where older granites dominate, whereas lower values (e.g., 0.705) occur in the Sacramento River basin where younger volcanic rock is more abundant. Values reflecting a mixture of these two ratios occur in the delta where the rivers join (e.g., 0.706). As fresh and marine water mix toward the ocean, the strontium isotope ratios (87Sr/86Sr) approach the global marine value (0.7092) (Ingram and Sloan 1992; Ingram and DePaolo 1993; Ingram and Weber 1999; Barnett-Johnson et al. 2008).
In this study, we compared the larval salinity distributions derived from otolith strontium isotope ratios (87Sr/86Sr) of sub-adult/adult Longfin Smelt (i.e., those that survived to maturity) with “observed” larval salinity distributions from larval survey catch data to evaluate the relative contributions of larvae rearing in various habitats to sub-adult/adult Longfin Smelt populations. To retrospectively determine the salinity distributions from otolith strontium isotope ratios (87Sr/86Sr), we first modeled the relationship between the strontium isotope ratios (87Sr/86Sr) and salinity “parts per thousand” (ppt) for water samples collected throughout the San Francisco Estuary. We used this empirical relationship and otolith microchemistry to estimate larval salinity histories from otoliths of sub-adult/adult Longfin Smelt from three different surveys. We then compared retrospective larval-stage salinity distributions of older fish with field-measured salinity distributions from the corresponding (i.e., same year-class) larval surveys to examine the relative contribution of different larval habitats to sub-adult/adult Longfin Smelt populations.
Materials and methods
Water chemistry
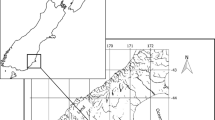
To characterize the variation in strontium isotope ratios (87Sr/86Sr) across the estuarine salinity gradient, water samples were collected in 2007 at 8 locations (the confluence of the Sacramento and San Joaquin Rivers near Sherman Island, the Pittsburg Marina, the Benicia Marina, Vallejo Harbor, Black Point, Point San Quentin, Sausalito, and Muir Beach) spanning a salinity gradient from 0.2 to 31.8-ppt (Fig. 1). Water samples (25-ml) were collected in acid rinsed polypropylene centrifuge tubes, filtered through 45-micron nylon filters and fixed in 1 M nitric acid in the field. Salinity and temperature where measured in the field at the time of water collections with a calibrated handheld YSI 90.

Map of the San Francisco Bay Estuary. Water sample sites denoted with numbers and details reported in Table 1
Strontium in water samples was purified using a standard micro-column chromatography technique to remove Rubidium. First, we evaporated 10–25-ml of water to obtain 0.3-micrograms of strontium. The dried strontium was then re-dissolved in 400-μl of 8-N nitric acid and loaded onto a wet chromatography column containing strontium “Spec resin” (Eichrom Inc.). Rubidium and other elements were removed from the column by manipulating the pH of the column by washing with 2 ml of 3 N nitric acid. The strontium was then eluted by again manipulating pH using 2.8-ml of 0.5-N nitric acid. This process was then repeated to ensure complete separation of rubidium from the strontium. The resulting pure strontium sample was then analyzed in solution mode on the Nu Plasma multi-collector ICP-MS. Replicate analyses of the SRM987 strontium standard over different analytical sessions since 2003 have yielded 87Sr/86Sr = 0.710220 ± 0.000035 (n = 45), within accuracy range of the accepted value of 87Sr/86Sr = 0.710248 for the SRM987 standard. Strontium isotope ratios (87Sr/86Sr) were normalized relative to 86Sr/88Sr = 0.1194 to correct for instrumental mass fractionation. Strontium isotope ratios (87Sr/86Sr) were then compared to corresponding field salinity measurements to develop a function for estimating salinity from strontium isotope ratio (87Sr/86Sr) values.
Otolith collection and processing
We analyzed otoliths of and sub-adult/adult Longfin Smelt collected by 3 fish surveys conducted by the Interagency Ecological ProgramFootnote 2: California Department of Fish and Game CDFG “Fall Midwater Trawl Survey” (FMWT), CDFG “Bay Study” and U.S. Fish and Wildlife Service (USFWS) “Chipps Island Study” (See Honey et al. 2004). The FMWT Survey samples 116 fixed stations located throughout the Delta, Suisun Bay and San Pablo Bay monthly from September-December (Fig. 1). A single oblique tow is conducted at each station with a midwater trawl net with a mouth opening of 3.7-m2 and a variable stretch mesh panel declining from 20.3-cm at the mouth to 1.3-cm at the cod end. The CDFG Bay Study uses the same midwater trawl gear and two methods to sample monthly, year-round at 52 fixed stations located throughout the San Francisco Estuary. The USFWS Chipps Island Survey samples monthly at 3 days per week, year-round at 3 parallel “trawl lanes” located in the channel adjacent to Chipps Island (i.e., at the confluence of the Sacramento and San Joaquin Rivers). Within a day ten 20-min tows are conducted alternating trawl lanes with each tow and using a surface trawl possessing a 9.1-m3 net mouth opening and mesh size graduated from 10-cm to 0.8-cm). Archived samples were obtained from the November and December 1999 and 2000 CDFG FMWT Surveys, January 2004 Bay Study, and the January and February 2008 Chipps Island Survey (Table 1). All Longfin Smelt used in otolith analyses were sub-adults (<75-mm FL) or adults (>75-mm FL) captured in Suisun Bay and preserved in 70% ethanol. Subsamples for otolith extraction and microchemical analysis were chosen randomly from the available catch.
Sagittal otoliths were removed from each fish and stored in 95% ethanol for clearing and whole-otolith imaging. Otoliths were mounted in Epoxicure® Footnote 3, cut in the frontal plane using a low-speed diamond tipped saw, and mounted on to glass slides using Crystal Bond thermoplastic glue (Crystalbond™ 509, Ted Pella Inc. Redding, CA). Otoliths were sanded with 1200 grit wet/dry sandpaper and polished with 0.3 micron alumina and a polishing cloth. Polished otoliths were washed with 1 M chemical grade nitric acid for 5 to 10 s, rinsed in an ultrasonic water bath for 5 min, and dried under a class 100 laminar flow hood.
Otolith microchemistry
Otolith strontium isotope ratios (87Sr/86Sr) were measured at the UC Davis Interdisciplinary Center for Plasma Mass Spectrometry. A multi-collector inductively coupled plasma mass spectrometer (Nu Plasma HR from Nu Instrument Inc.) was interfaced with a Nd:YAG 213 nm laser (New Wave Research UP213) for strontium isotope ratio (87Sr/86Sr) measurement by laser ablation (LA-MC-ICP-MS technique). A laser beam of 60 µm diameter traversed across the otolith from the core to the edge at 10 µm per second, with the laser pulsing at 10 Hz and at 75% power output. Helium was uses as the carrier gas to maximize sensitivity and minimize sample deposition at the ablation site, and was mixed with Argon gas between the laser sample cell and the plasma source. Gas blank and background signals were monitored until 84Kr and 86Kr stabilized after the sample change (i.e., exposing sample cell to the air) and were measured for 30 s. The laser was typically fired for 90–120 s and background signals were subtracted from the measured signals automatically. Strontium isotope ratios (87Sr/86Sr) were normalized relative to 86Sr/88Sr (0.1194) to correct for instrumental mass fractionation. The signal on mass 85 was monitored to correct for any 87Rb interference on 87Sr. The analytical accuracy was determined by using the results of replicate analyses of a modern marine mollusk at the beginning and end of several analytical sessions. Three replicate analyses yielded 87Sr/86Sr = 0.70919 ± 0.000096 (n = 28), consistent with modern seawater values of 0.70918. Data reduction was conducted off-line using Matlab 9.0 software.
Larval salinity distributions of sub-adult and adult fish
The larval salinity history of sub-adult/adult Longfin Smelt was calculated using the empirically-derived relationship between water salinity (ppt) and strontium isotope ratios (87Sr/86Sr) measured at our sampling sites (Fig. 1). This relationship was described (r2 0.98, p < 0.0001) by the asymptotic Boltzman function (Origin Lab 6.0 software). The resulting model was solved to estimate salinity from otolith strontium isotope ratios (87Sr/86Sr), assuming no isotopic fractionation. The strontium isotope ratio (87Sr/86Sr) relationships were modeled for each river (Sacramento and San Joaquin Rivers) and the common marine end-member (Pacific Ocean at Muir Beach) using a standard linear mixing model (Ingram and Sloan 1992). All Longfin Smelt in the San Francisco Estuary appear to spend time in the marine environment immediately prior to their spawning migration to freshwater (Rosenfield and Baxter 2007). To avoid potential bias from maternal contributions, otolith strontium isotope ratio (87Sr/86Sr) minima adjacent to the maternally derived core (ca. 50-µm from core) were used in conjunction with the salinity function to assign each fish a larval salinity value. This otolith location corresponds to a larva size of 8 to 10-mm total length (TL) and was used for comparison to field-caught larvae. Each fish was categorized as originating from fresh (< 0.3-ppt), low salinity zone (0.3–4.0-ppt) or brackish (> 4.0-ppt) waters according to previous descriptions of Longfin Smelt nursery habitats (Bennett et al. 2002; Hobbs et al. 2006). The relative proportions of Longfin Smelt from each salinity category were calculated for comparison among years and among sub-adult/adult and larval populations.
Larval salinity distributions from catch surveys
We obtained information on the salinity distributions of Longfin Smelt larvae (8–10 mm TL) from the CDFG “20-mm Survey”, a bi-weekly fish monitoring program sampling at 41 fixed stations located within the Delta and Suisun Bay and conducted from late March-July. The survey makes 3 oblique tows at each station using a sled-mounted net with a 1.5-m2 opening and a 5.1-m long body composed of a 1.6-mm mesh designed specifically to target larval and juvenile smelt (Honey et al. 2004). Pre- and post-tow readings of a calibrated General Oceanics flow meter suspended in the net mouth were used to determine distance traveled through the water for each tow. Volume of water filtered during each tow was calculated as product of distance traveled through the water and the net mouth area. The metric, Catch per unit effort (CPUE), was calculated for each station, using the total number of Longfin Smelt in the 8 to 10-mm TL size range collected, and dividing by total volume sampled for the 3 replicate tows conducted per station. This metric is hereinafter referred to simply as “catch”. To derive salinity, surface and bottom electrical conductivity measurements were taken once per station, averaged and transformed to mean salinity according to Poisson (1980). All catch data from a given station were grouped based on the station salinity value, then summed into one of the three larval habitats: fresh (<0.3-ppt), low salinity zone (0.3 to 4-ppt) or brackish (>4-ppt). For statistical comparisons among survey and otolith chemistry data, the catch for each salinity category was standardized to a proportion of the total annual catch for each year.
Analyses
Differences in the mean and variance of larval salinities were compared among years and within year-classes using bootstrapped (1,000×, n = 100) 95% confidence intervals (α = 0.05). Differences among years and between life stages within year-classes in the relative frequency of fish from the three larval habitats (fresh, low salinity zone and brackish waters) were compared using the Pearson chi-square test statistic (α = 0.05). Relative proportions were used to display these results. All statistical analyses were conducted using Systat 12.
Results
Strontium isotope ratios (87Sr/86Sr) vs. salinity
Strontium isotope ratios (87Sr/86Sr) in water samples varied across the estuary and served as a very good proxy for salinity from 0.2 to 5-ppt (Fig. 2). For example a 1-ppt difference in salinity resulted in approximately 4-ppm difference for strontium isotope ratios (87Sr/86Sr) up to 5-ppt salinity, and 1-ppm difference in strontium isotope ratios (87Sr/86Sr) up to a salinity of 13-ppt. Thus, variations in strontium isotope ratios (87Sr/86Sr) were well within the 2σ (± 0.0001) measurement error on the mollusk standard using the LA-MC-ICMPS (Table 2). However, the relationship approached an asymptote at salinities above 5-ppt, at which, salinity resolution was poor (Fig. 2). Due to the differences in main tributary isotope ratios (Sacramento River ca 0.705; San Joaquin River ca 0.707), there existed a ratio range for which we could not distinguish tributary mixing from freshwater-marine water mixing. Strontium isotope ratios (87Sr/86Sr) from 0.7064 to 0.7072 could represent either primarily San Joaquin River water or primarily Sacramento River water with a small mixture of San Joaquin River and marine waters. During both a wet year (2006) and a dry year (2007) strontium isotope ratios (87Sr/86Sr) for freshwaters at the confluence of the Sacramento and San Joaquin Rivers at Sherman Island were never below 0.7064; thus, values below this were definitively freshwaters (unpublished data). However, the strontium isotope ratio (87Sr/86Sr) at Mossdale on the San Joaquin River was 0.7072; and thus, freshwater signatures can be higher than 0.7064 if San Joaquin River outflow was relatively high (as in 1999). Therefore, for the purposes of this study, any values less than 0.7072 were considered freshwater (<0.3-ppt); though it is possible that values between 0.7064 and 0.7072 may also reflect very low salinity (<1-ppt) waters from the lower Sacramento River, thus potentially underestimating salinity values. However, this bias could not account for more than 1-ppt difference in salinity, assuming all freshwater entering the estuary was derived from Sacramento River water. Strontium isotope ratio (87Sr/86Sr) mixing models for the Sacramento River and Muir Beach and the San Joaquin River and Muir Beach were similar at salinities above 3-ppt, and demonstrate the variability in strontium isotope ratios (87Sr/86Sr) below 1-ppt (Fig. 2). Mixing curves and 95% confidence intervals encompassed all water strontium isotope ratio (87Sr/86Sr) values from this study and from previously published data (Ingram and Sloan 1992). The mixing curve exhibited slightly elevated strontium isotope ratios (87Sr/86Sr) between 2 and 6-ppt due to the very low strontium concentrations in Sacramento River water versus the delta and San Joaquin River. Overall, strontium isotope ratios (87Sr/86Sr) proved an excellent tool for estimating ambient salinities for low to moderate salinity waters inhabited by larval Longfin Smelt.

Strontium isotopes ratios (87Sr/86Sr) to salinity (ppt) relationships from water samples collected across the estuary (grey circles) and fit with a Boltzmann curve (dotted line) ±95% CI (black dashed lines). Dark grey line depicts a mixing curve between the Sacramento River (fresh) strontium isotope ratio (87Sr/86Sr) and the isotope ratio for Muir Beach (marine). Light grey line depicts a mixing curve between the San Joaquin River (fresh) strontium isotope ratio (87Sr/86Sr) and the Muir Beach sample
Salinity distributions from larval surveys and sub-adult/adult otoliths
Salinity distributions derived from catch of larval Longfin Smelt (<10 mm TL) had a broad range (ca. 0 to 15-ppt) with multiple peaks around 1 to 3-ppt and brackish waters >5-ppt (Fig. 3a-d). In contrast, salinity distributions derived from otolith strontium isotope ratios (87Sr/86Sr) for sub-adult/adult fish exhibited more narrow ranges (ca. 0 to >5-ppt) with single peaks between 1 and 3-ppt (Fig. 3e-h). We observed no fish with larval rearing strontium isotope ratios (87Sr/86Sr) for salinities greater than >5-ppt in 2003 and 2006 year classes (Fig. 3g-h). Bootstrapped mean larval salinities from larval catch data ranged from 4 to 5-ppt and did not vary significantly among years (Fig. 4a). In contrast, mean larval salinities of sub-adult/adult fish derived from strontium isotope ratios (87Sr/86Sr) were significantly lower in 2003 (1.2-ppt) than in all other years (approximately 2.5-ppt) and between larval catch and sub-adult/adult fish otolith strontium isotope ratios (87Sr/86Sr; Fig. 4a). The variance of larval salinity distributions was much higher for larval surveys than sub-adult/adults, with the narrowest distributions of sub-adult/adult fish occurring in 2003 and 2006 (Fig. 4b).

Proportional catch distribution of salinity residence for larval Longfin Smelt collected during a 1999, b 2000, c 2003 and d 2006 CDFG 20-mm Surveys. Also, the proportional distribution of larval salinity residence retrospectively derived from otolith strontium isotope ratios (87Sr/86Sr) of sub-adult/adult Longfin Smelt collected in e 1999 CDFG Fall Midwater Trawl Survey f 2000 CDFG Fall Midwater Trawl Survey g 2004 CDFG Bay Study and h 2008 Chipps Island Survey. Note: while the salinity distribution for catch are empirically derived and can take values between 0-35 ppt, the otolith strontium isotope ratios derived salinities are poorly resolved above 5 ppt and are only presented here for continuity with the catch distributions

a Bootstrapped mean salinity values and ±95% confidence limits for otolith strontium isotope ratio (87Sr/86Sr)–salinity (subadult) and CPUE-salinity data (larvae). b Bootstrapped mean variance and ±95% confidence limits for otolith strontium isotope ratio (87Sr/86Sr)–salinity (subadult) and CPUE-salinity data (larvae)
A comparison of the frequency distributions of the three larval habitats (fresh, low salinity, and brackish) among years for the 20-mm. Survey catch data, was statistically significant (Fig. 5a, Table 3), with a majority of larvae caught in brackish and low salinity zone waters and few in freshwaters. (χ2 = 18.5, p = 0.005). Pairwise, between year-class, comparisons of the frequency distribution of larvae across habitats revealed a significant difference between 2000 and 2006 year-classes (χ 2 = 15.2, p < 0.001), where 2006 had a majority of fish in brackish waters while 2000 had an equal distribution of fish in low salinity zone and brackish habitats. All other pair-wise comparisons among years were not significantly different (p > 0.05) (Fig. 5a, Table 3).

Proportion of Longfin Smelt from each habitat type (fresh, low salinity zone, brackish) for a larval catch and b adult otolith microchemistry. Salinity binned by categories, fresh = 0 to 0.3-ppt; low salinity 0.4 to 3-ppt; and brackish >4-ppt. Same letters indicate non-significant differences
Relative contributions of the three larval habitats derived from sub-adult/adult otolith strontium isotope ratios (87Sr/86Sr) data differed significantly among all years (Table 3), with a majority of fish with low salinity zone origins (χ 2 = 44.3, p < 0.001)(Fig. 5b). All inter-annual comparisons were significantly different (p < 0.05) with the exception of 2000 and 2006, where low salinity zone and brackish origin distribution were similar, but no freshwater fish were found in 2006 while a few were observed in 2000 (χ 2 = 2.8, p = 0.246) (Fig. 5b, Table 3). The clearest pattern observed was the large contribution of fish from low salinity waters to sub-adult/adult populations in 2000, 2003 and 2006, in contrast with the more even contribution of all habitats in 1999.
Comparisons of the contribution of the three larval habitats from sub-adult/adult otolith strontium isotope ratio (87Sr/86Sr) data with corresponding distributions from larval catch data for each corresponding year were all significantly different (Fig. 5a-b, Table 4). In 1999, it appeared that fish from brackish waters made a relatively lower contribution to the sub-adult population than the low salinity zone and freshwaters (χ 2 = 107, p = < 0.001) (Table 4). In 2000 and 2003 the brackish waters also exhibited a lower contribution to the sub-adult/adults, however the low salinity zone contributed a majority of successfully recruited sub-adult/adults and the freshwaters contributed fewer fish to the sub-adult/adult population (2000, χ 2 = 16, p = < 0.001 and 2003, χ 2 = 26, p = < 0.001). Similarly, 2006 experienced higher relative contribution of low salinity zone fish and low contribution of brackish fish, but no fish from freshwater were observed in the sub-adult/adult population (χ 2 = 107, p = < 0.001). If we used the more conservative freshwater cutoff of 0.7064, we observed no successfully recruited sub-adult/adults fish originating from freshwater habitats for all year-classes.
Discussion
In this study, strontium isotope ratios (87Sr/86Sr) proved a valuable tool for discriminating among low salinity habitats and reconstructing larval salinity origins for successfully recruited sub-adult/adult Longfin Smelt from the upper San Francisco Estuary. Compared to the Sr/Ca ratio technique, the strontium isotope ratios (87Sr/86Sr) were far more precise at salinities from 0–5-ppt (Secor et al. 1995), providing at a minimum of 1-ppt salinity resolution up to 5-ppt (Ingram and Sloan 1992). The strontium isotope ratios (87Sr/86Sr) changed little with salinity above 13-ppt, being apparently overwhelmed by marine sources strontium. For this reason strontium isotope ratios (87Sr/86Sr) may predict salinity history as well as Sr/Ca ratios for mesohaline and polyhaline environments. On the freshwater end of the distribution, the strontium isotope ratio (87Sr/86Sr) measured at Sherman Island was slightly higher than predicted by the Sacramento River—Marine water mixing model suggesting that substantial mixing of Sacramento and San Joaquin river waters had taken place by the time waters reached this location in the western Delta during this study. However data from Ingram and Sloan (1992) from the same location agree with the Sacramento River mixing model, highlighting the potential for inter-annual variability of strontium isotope ratios at this location. Water from the Sacramento and San Joaquin rivers mixes in Sherman Lake and several channels farther west, and local spring and neap tidal volume fluxes, which range from 1,800 to 5,900 m3·s-1, power this mixing (Kimmerer 2004). Nonetheless, the strontium isotope ratios (87Sr/86Sr) provided a very precise estimate of salinity in the upper estuary for salinities less than 5-ppt.
Strontium isotope ratios (87Sr/86Sr) in the larval region from otoliths of sub-adult/adult Longfin Smelt revealed very different pictures of favorable salinity habitats than did field observations of larval abundance. In 3 of the 4 years studied (2000, 2003 and 2006 year-classes), the highest catch of larval Longfin Smelt occurred at salinities greater than those estimated using strontium isotope ratios (87Sr/86Sr) of otolith cores for the same yearclass of fish collected as sub-adults/adults 10 months to 2 years after hatch. Furthermore, no successful recruits were observed with freshwater natal origins in 2006 and greater than 6-ppt in all years of the study. The majority of Longfin Smelt experienced the greatest recruitment success at salinities around 2-ppt (Fig. 3). Successful recruitment of larvae using the low salinity zone (0.5 to 6-ppt, Kimmerer 2004) is consistent with studies of larval Longfin Smelt that indicate the low salinity zone of the San Francisco Estuary is a region of high feeding success and good body condition (Bennett et al. 2002; Hobbs et al. 2006). Assuming the lack of fish in the sub-adult/adult population from salinity habitats (<1 and >6-ppt) is a reflection of mortality, larvae encountered in marginal salinity habitats may account for as much as 74% of the total larval population in some years, but a consistently small percentage of the sub-adult/adult population. Disproportionate contribution of larvae from the low salinity zone to the sub-adult/adult population highlights its potential role as nursery habitat (sensu Beck et al. 2000) for Longfin Smelt.
Though we believe the patterns presented represent the true shape of the salinity distributions of successfully recruited sub-adult/adult fish, we cannot rule out the possibility that temporal differences in salinity distributions were influenced by variation in sampling methods employed by the different sub-adult/adult fish surveys (e.g., surveys differed in the size and age distribution of specimens collected). For example, the 1999 and 2000 FMWT Surveys collected primarily age 0 fish, the 2004 Bay Study collected primarily age 1 fish and the 2008 Chipps Island survey collected primarily age 2 fish. To minimize the age effect, we selected individuals of the same year-class in each sub-adult/adult survey-year and compared them to larval catch data for that corresponding year-class only: age 0 for 1999 and 2000 (year-classes 1999 and 2000), age 1 for 2004 (year-class 2003) and age 2 for 2008 (year-class 2006). However, variability between years may reflect differences in cumulative mortality among different aged fish (0 for 1999 and 2000, 1 for 2004 and 2 for 2008) and not an inter-annual trend in larval salinity history. To minimize spatial effects (fish with different migration histories) on salinity distributions, all specimens selected for this study were collected within Suisun Bay during the winter spawning season; assuming all fish are spawning within the freshwaters of Suisun Bay (Moyle 2002). Despite the differences in survey methods and age classes examined, it is important to note that we observed disproportionate contributions of larvae from the low salinity zone to sub-adult/adult populations across all survey methods, strongly emphasizing the role of this zone as an important nursery area for Longfin Smelt.
We also could not rule out the potential for temporal bias (e.g., comparing the wrong region of the otolith for chemistry to catch) when comparing otolith-based salinity distributions with those from larval surveys. To minimize this potential artifact, we counted daily rings and measured the distance from the otolith core to the age at which fish average 10-mm TL. Thus, we used otolith chemistry data from regions of the otolith corresponding to a retrospective size of ca. 10-mm TL. However, in many fish we observed elevated strontium isotope ratios (87Sr/86Sr) at the core region of the otolith that extended into the time period when fish were 10-mm TL. This enriched region likely reflects a maternal marine influence, similar to those observed in otoliths of other migratory salmoniforms (e.g., Zimmerman and Reeves 2002). In these instances we used the lowest strontium isotope ratio (87Sr/86Sr) immediately following the elevated core region (∼30–50 µm), assuming this reflected the true (though conservative) larval salinity signature. An alternative interpretation of the enriched cores would entertain that larvae may have hatched at higher salinities, but very quickly made their way upstream into lower salinity water. We think this is unlikely given what is known about the migration strategies of Longfin Smelt, but cannot rule out this possibility. Studies are currently underway to examine maternal versus environmental signatures in the core region of Longfin Smelt otoliths. Preliminary data show that larval Longfin Smelt cannot tolerate salinities greater than 5-ppt for more than a few hours, making the elevated cores due to environmental exposure an unlikely prospect (unpublished data).
The use of strontium isotope ratios (87Sr/86Sr) for salinity history reconstruction in estuaries could have broad application in the field of estuarine research. Many estuarine fish have been previously shown to utilize the freshwater-low salinity waters for spawning and larval rearing in other systems. In the Chesapeake Bay Estuary Striped Bass (Morone saxatilits) spawn in freshwaters, while the larvae and juveniles rear in the low salinity waters. In addition, striped bass have been shown to exhibit multiple population contingents based on salinity residence during the early life (Secor 1999; Secor et al. 2001). The application of strontium isotope ratios (87Sr/86Sr) in this system could provide for a much more refined examination of the early life history dynamics in the upper estuary. In the St Lawrence River Estuary, Rainbow Smelt (Osmorus mordax), is ecologically similar to the Longfin Smelt and also uses low salinity to brackish water habitats as juveniles. Dodson et al. (1989), found that larval smelt exhibited variable vertical migration and retention strategies in response to variable hydrodynamics among three channel-island habitats between Ile d'orleans and Ile-aux-Coudres of the upper estuary. A parallel study found reduced growth rates for fishes occurring in downstream sites in higher salinity waters (Laprise and Dodson 1989). Moreover, fish residing in the north shore and south shore were genetically distinct and represented two sympatric populations, highlighting the importance of the larval retention strategies for maintaining population structure (Pigeon et al. 1998). The application of otolith strontium isotope ratios (87Sr/86Sr) could resolve the relationship between salinity residence and survival, and potentially provide new insights into inter-annual abundance patterns.
Conclusion
Our results demonstrate the utility of otolith strontium isotope ratios (87Sr/86Sr) in retrospectively providing new insights into the population dynamics of estuarine fishes. Strontium isotope ratios (87Sr/86Sr) served as excellent proxies for salinity up to 5-ppt and appears more reliable than traditional Sr/Ca ratios for low salinities (e.g., 0–5-ppt). We found that larvae from the 2-ppt low salinity zone were disproportionately represented in sub-adult/adult surveys, suggesting this low salinity habitat serves as the principle nursery habitat for the Longfin Smelt. Furthermore, this habitat appeared particularly important to the adult Longfin Smelt population in the most recent years of this study, during which the population abundance declined. As nursery habitat for this (and other) threatened species of fish, the 2-ppt low salinity zone should receive significant attention and protection by resource managers. The mechanism responsible for poor survival of larvae from fresh and brackish waters in 2003 and 2006 may correspond with the overall decline of several pelagic organisms in the estuary. Future studies should continue to examine the relative influence of potential mechanisms leading to differences in larval survival in these different habitats.
Notes
On March 4, 2009 the Fish and Game Commission (Commission) made a final determination that the listing of longfin smelt as a threatened species was warranted. The Commission has initiated a rulemaking process to officially add longfin smelt to the California Endangered Species Act (CESA) list of threatened species found in the California Code of Regulations (CCR), Title 14, section 670.5(b)(2). At the completion of this rulemaking process, the longfin smelt’s status will officially change from candidate to threatened.
References
Armor C, Baxter R, Bennett B, Breuer R, Chotkowski M, Coulston P, Denton D, Herbold B, Kimmerer W, Larsen K, Nobriga M, Rose K, Sommer T, Stacey M (2005) Interagency ecological program synthesis of 2005 work to evaluate the pelagic organism decline (POD) in the upper San Francisco Estuary. Rep. to the Interagency Ecol. Program. www.science.calwater.ca.gov/pod.
Barnett-Johnson R, Pearson TE, Ramos FC, Grimes CB, MacFarlane RB (2008) Tracking natal origins of salmon using isotopes, otoliths, and landscape geology. Limnol Oceanogr 53:1633–1642
Baxter R, Breuer R, Brown L, Chotkowski M, Feyrer F, Gingras M, Herbold B, Mueller-Solger A, Nobriga M, Sommer T, Souza K (2007) Pelagic organism decline progress report: 2007 synthesis of results. Rep. to the Interagency Ecol. Program. www.science.calwater.ca.gov/pod.
Beck MW, Heck KL Jr, Able KW, Childers DL, Eggleston DG, Gillanders BM, Halpern B, Hays CG, Hoshino H, Minello TJ, Orth RJ, Sheridan PF, Weinstein MP (2000) The identification, conservation and management of estuarine and marine nurseries for fish and invertebrates. Biosci 58(8):633–641
Bennett WA, Kimmerer WJ, Burau JR (2002) Plasticity in vertical migration by native and exotic estuarine fishes in a dynamic low-salinity zone. Limnol Oceanogr 47:1496–1507
California Fish and Game Commission (2009) California fish and game commission notice of findings: longfin smelt (Spirinchus Thaleichthys). Meeting: Woodland California, May 29, 2009.
Campana SE (1999) Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Mar Ecol Prog Ser 188:263–297
Campana SE, Chouinard GA, Hanson JM, Frechet A, Brattey J (2000) Otolith elemental fingerprints as biological tracers of fish stocks. Fish Res (Amsterdam) 46:343–357. doi:10.1016/S0165-7836(00)00158-2
Dege M, Brown LR (2004) Effect of outflow on spring and summertime distribution and abundance of larval and juvenile fishes in the upper San Francisco Estuary. In: Feyrer F, Brown LR, Brown RL, Orsi JJ (eds) Early life history of fishes in the San Francisco Estuary and watershed, 39th edn. American Fisheries Society, Bethesda, pp 49–65
Dodson JJ, Dauvin JC, Ingram RG, D’Anglejan G (1989) Abundance of larval rainbow smelt (Osmerus mordax) in relation to the maximum turbidity zone and associated macroplankton fauna of the Middle St. Lawrence Estuary Estuar 21:66–81
Elsdon TS, Wells BK, Campana SE, Gillanders BM, Jones CM, Limburg KE, Secor DH, Thorrold SR, Walther BD (2008) Otolith chemistry to describe movements and life-history parameters of fishes: hypotheses assumptions, limitations and inferences. Oceanography and Marine Biology: An Annual Review 46:297–330
Faure G, Mensing TM (2005) Isotopes: principles and applications, 3rd edn. Wiley, Hoboken
Feyrer F, Hobbs J, Baerwald M, Sommer T, Yin QZ, Clark K, May B, Bennett W (2007a) Otolith microchemistry provides information complementary to microsatellite DNA for a migratory fish. Trans Am Fish Soc 136:469–476. doi:10.1577/T06-044.1
Feyrer F, Nobriga ML, Sommer TR (2007b) Multidecadal trends for three declining fish species: habitat patterns and mechanisms in the San Francisco Estuary, California. USA Can J Fish Aquat Sci 64:723–734. doi:10.1139/F07-048
Gillanders B (2005) Using elemental chemistry of fish otoliths to determine connectivity between estuarine and coastal habitats. Estuar Coastal Shelf Sci 64:47–57. doi:10.1016/j.ecss.2005.02.005
Hobbs JA, Q-z Yin J, Burton WA Bennett (2005) Retrospective determination of natal habitats for an estuarine fish with otolith Sr isotope ratios. Mar Freshw Res 56:655–660
Hobbs JA, Bennett WA, Burton JE (2006) Assessing nursery habitat quality for native smelts (Osmeridae) in the low-salinity zone of the San Francisco estuary. J Fish Biol 69:907–922. doi:10.1111/j.1095-8649.2006.01176.x
Hobbs JA, Bennett WA, Burton J, Gras MA (2007) Classification of larval and adult delta smelt to nursery areas by use of trace elemental fingerprinting. Trans Amer Fish Soc 136:518–527. doi:10.1577/T06-087.1
Honey K, Baxter R, Hymanson Z, Sommer T, Gingras M, Cadrett P (2004) IEP Long-term monitoring program element review. Report to the Interagency Ecological Program for the San Francisco Bay/Delta Estuary. (http://www.water.ca.gov/iep/docs/IEP_FishMonitoring_final.pdf.)
Ingram BL, Sloan D (1992) Strontium isotopic composition in estuarine sediments as paleosalinity and paleoclimate indicator. Science 255:68–72
Ingram BL, DePaolo DJ (1993) A 4300 year Sr isotope record of estuarine paleosalinity in San Francisco Bay. California Earth Planetary Sci Let 119:103–119
Ingram BL, Weber PK (1999) Salmon origin in California’s Sacramento-San Joaquin river system as determined by otolith Sr isotopic composition. Geol 27:851–854
Jassby AD, Kimmerer WJ, Monismith SG, Armor C, Cloern JE, Powell TM, Schubel JR, Vendlinski TJ (1995) Isohaline position as a habitat indicator for estuarine populations. Ecological Applications 5(1):272–289
Kalish JM (1989) Otolith microchemistry: validation of the effects of physiology, age and environment on otoliths composition. J Exp Mar Biol Ecol 132:151–178
Kennedy BP, Blum JD, Folt CL, Nislow KH (2000) Using natural Sr isotopic signatures as fish markers: methodology and application. Can J of Fish Aquat Sci 57:2280–2292
Kimmerer WJ (2004) Open water processes of the San Francisco Estuary: from physical forces to biological processes. San Fran Est Watersh Sci 2(1):1–142
Kimmerer WJ, Gross ES, MacWilliams ML (2009) Is the response of estuarine nekton to freshwater flow in the San Francisco Estuary explained by variation in habitat volume? Estuar Coasts 32:375–389. doi:10.1007/s12237-008-9124-x
Laprise R, Dodson JJ (1989) Ontogeny and importance of tidal vertical migrations in the retention of larval smelt Omserus mordax in a well-mixed estuary. Mar Ecol Prog Ser 55:101–111
Lehman PW, Boyer G, Satchwell M, Waller S (2008) The Influence of environmental conditions on the seasonal variation of Microcystis cell density and microcystins concentration in the San Francisco Estuary. Hydrobiol 600:187–204. doi:10.1007/s10750-007-9231-x
Milton D, Halliday I, Sellin M, Marsh R, Staunton-Smith J, Woodhead J (2008) The effect of habitat and environmental history on otolith chemistry of barramundi Lates calcarifer in estuarine populations of a regulated tropical river. Estuar, Coas Shel Sci 78:301–315
Moulton LL (1974) Abundance, growth and spawning of the Longfin Smelt in Lake Washington. Trans Am Fish Soc 103(1):46–52
Moyle PB (2002) Inland fishes of California. University of California Press, Berkeley
Outridge PM, Chenery SR, Babaluk JA, Reist JD (2002) Analysis of geological Sr isotope markers in fish otoliths with subannual resolution using laser ablation-multicollector-ICP-mass spectrometry. Environ Geol 42:892–899. doi:10.1007/s00254-002-0596-x
Pigeon D, Dodson JJ, Bernatchez L (1998) A mtDNA analysis of spatiotemporal distribution of two sympatric larval populations of rainbow smelt (Osmerus mordax) in the St. Lawrence River estuary, Quebec, Canada. Can J Fish Aquat Sci 55:1739–1747
Poisson A (1980) Conductivity/salinity/temperature relationship of diluted and concentrated seawater. IEEE J Oceanic Eng 5(1):41–50
Rosenfield JA, Baxter RD (2007) Population dynamics and distribution patterns of Longfin Smelt in the San Francisco estuary. Trans Am Fish Soc 136:1577–1592. doi:10.1577/T06-148.1-x
Secor DH (1999) Specifying divergent migrations in the concept of stock: the contingent hypothesis. Fish Res 43:13–34
Secor DH, Rooker JR (2000) Is otoliths strontium a useful scalar of life cycles in estuarine fishes? Fish Res 46:359–371
Secor DH, Henderson-Arzapalo A, Piccoli PM (1995) Can otoltih microchemistry chart patterns of migration and habitat utilization in anadramous fishes? J Exper Mar Biol Ecol 192:15–33
Secor DH, Rooker JR, Zlokovitz E, Zdanowicz VS (2001) Identification riverine, estuarine, and coastal contingents of Hudson River striped bass based upon otolith elemental fingerprints. Mar Ecol Prog Ser 211:245–253
Sommer T, Armor C, Baxter R, Breuer R, Brown L, Chotkowski M, Culberson S, Feyrer F, Gingras M, Herbold B, Kimmerer W, Mueller-Solger A, Nobriga M, Souza K (2007) The collapse of pelagic fishes in the Upper San Francisco Estuary. Fisheries (Bethesda) 32:270–277. doi:10.1577/1548-8446-x
Thorrold SR, Jones CM, Campana SE, McLaren JW, Lam JWH (1998) Trace element signatures in otoliths record natal river of juvenile American shad (Alosa sapidissima). Limnol Oceanogr 43:1826–1835
Woodhead J, Swearer S, Hergt J, Maas R (2005) In situ Sr-isotope analysis of carbonates by LA-MC-ICP-MS: interference corrections, high spatial resolution and an example from otolith studies. J Anal Atom Spec 20:22–27. doi:10.1039/b412730g
Zimmerman CE, Reeves GH (2002) Identification of steelhead and resident rainbow trout progeny in the Deschutes River, Oregon, revealed with otolith microchemistry. Trans Am Fish Soc 131:986–993. doi:10.1577/1548-8659
Acknowledgements
The authors would like to thank the California Department of Fish and Game for sample collections and the Interagency Ecological Program for funding. This project was funded under the UCD Umbrella Contract to UCD for work on the Pelagic Organism Decline DWR Contract # 4600008137. Significant revisions to the manuscript were provided by Josh Israel, Jon Rosenfield and 2 anonymous reviewers. Special thanks to Peter Moyle for laboratory space and Bill Bennett for equipment loan.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Hobbs, J.A., Lewis, L.S., Ikemiyagi, N. et al. The use of otolith strontium isotopes (87Sr/86Sr) to identify nursery habitat for a threatened estuarine fish. Environ Biol Fish 89, 557–569 (2010). https://doi.org/10.1007/s10641-010-9672-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-010-9672-3