
Abstract
In the absence of high-quality biodiversity data, land-use planners and conservationists often rely on biodiversity surrogates. Many studies have proposed surrogates based on assumptions about the environmental niches of species. However, the use of such assumptions is not always useful because biological processes and ecological interactions can operate at different scales due to the non-uniform geographical distribution of environmental conditions. In such cases, compositional heterogeneity across the same region can be expected but is often hidden by broad-scale environmental data. Furthermore, these assumptions may obscure important relationships between species and their environment. To elucidate this issue, we asked whether biotic interactions between two taxonomic groups are more important than other factors in reflecting the distribution of unsampled species. To do this, we compared the relationship between the distribution of bird species and the distribution of the five most-abundant palm species which are often considered indicators of bird communities. These species include Lepidocaryum tenue, Oenocarpus bataua, Oenocarpus bacaba, Mauritiella aculeata, and Attalea speciosa. Additionally, we considered environmental factors (precipitation, water-table levels, sand and clay contents) and the ecoregions along the Purus-Madeira interfluve as drivers of bird species composition. Our results show that bird-assemblage composition was strongly correlated with changes in palm-species abundance. The presence-absence data for bird-species showed that palm-species alone explained 25% and 19% of composition of all birds and only canopy birds, respectively. These palm species are abundant and can be easily identified and monitored by non-specialists, such as citizen scientists. Citizens are often involved in data acquisition but may not have the experience to sample large assemblages consisting of hundreds of species; thus, these five most-abundant palms species could serve as a cost-effective and efficient biodiversity surrogate for birds.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the absence of high-quality biodiversity data, land-use planners and conservationists often rely on biodiversity surrogates (Margules and Pressey 2000; Pinto et al. 2008), which are species, groups of species, or environmental variables used as proxies to represent broader aspects of biodiversity (Margules and Pressey 2000; Pinto et al. 2008). Use of biodiversity surrogates requires a trade-off between the surrogate’s ability to reflect the distribution of unsampled species and the cost of sampling species in the field (Gardner et al. 2008; Lansac-Tôha et al. 2022). This approach has been applied to numerous taxonomic groups in terrestrial (Corcos et al. 2021; Verdinelli et al. 2022) and aquatic ecosystems (Pakulnicka et al. 2015; Spigoloni et al. 2022). If a surrogate is identified, the results could be utilized to prioritize locations or actions for conservation purposes (Nogueira et al. 2023).
Evaluating the effectiveness of surrogates in conservation planning is challenging (Fattorini et al. 2012; Covre et al. 2022) since it necessitates the availability of the same extensive datasets whose absence initially justified the use of surrogates. For example, in the tropics, where endemic species are common and little information is available regarding the life history and ecology of most taxa, the surrogate approach has been recommended, even for relatively species-poor communities and relatively small areas (Wiens et al. 2008). However, studies often propose surrogates without considering who will use them and why they are better than direct measures (Caro 2010).
In cases where surrogates appear to underestimate complementarity, assessing the effectiveness of frequently used surrogates or establishing new ones may be necessary. In the Amazon, data on the distributions of various taxonomic groups are still too incomplete to serve as the foundation for systematic conservation planning (Carvalho et al. 2023). As a result, conservation targets aiming to maximize complementarity between protected areas have been established based on untested assumptions about the environmental niches of most species. These targets typically rely on ecoregions, vegetation types, and soil types as biodiversity surrogates (Peres 2005). However, the use of general classifications, such as broadscale environmental data, is not always useful because biological processes and ecological interactions can operate at different scales due to the non-uniform geographical distribution of environmental conditions (McGill 2010; Lu and Jetz 2023). Moreover, relying solely on broadscale environmental data may obscure important relationships between species and their environment (Qian and Kissling 2010; Monteiro et al. 2023).
This appears to be the case for the Purus-Madeira interfluve located in southwestern Amazonia; it is a large, forested area extending over 800 km where maps of vegetation types and geology show a relatively homogeneous landscape (Olson et al. 2001). However, biological surveys have revealed compositional heterogeneity across the region in many taxonomic groups, including ants (Baccaro et al. 2013), bats (Marciente et al. 2015), trees (Schietti et al. 2016), lizards (Peixoto et al. 2019), fishes (Stegmann et al. 2019), and dung beetles (Salomão et al. 2022). These results suggest that the effectiveness of using broadscale environmental data as biodiversity surrogates in large forested areas, such as the Amazon, could be inconsistent. Therefore, it becomes necessary to identify alternative and effective biodiversity surrogates to support conservation efforts in the region.
One potential alternative is considering the biotic interactions between taxonomic groups (Cosentino et al. 2023; Bazzato et al. 2023). For example, if biotic interactions between birds and palms are significant, then variations in bird species composition - defined by the presence or absence of specific bird species - would correspond with changes in the abundance of palm species. This expectation could be justified because birds interact with palm species in various ways, utilizing them as food resources, nesting, roosting and perching sites (Snow 1981; Brightsmith and Cáceres 2017; van der Hoek et al. 2019; Glória and Tozetti 2021). The extent to which birds exploit different plant species depends on their specific characteristics, such as whether they are canopy or understory birds (Bradfer-Lawrence et al. 2018; Lima et al. 2023). On the other hand, the composition of palm species changes in response to environmental variation (Sousa et al. 2020; Muscarella et al. 2020), with some species showing close associations with specific environmental characteristics. For instance, the presence of Oenocarpus bacaba palm species indicates well-drained soils, while Lepidocaryum tenue is more commonly found in wet conditions (Cohn-Haft et al. 2007).
To assess the potential of using data on palms as a substitute for bird assemblages, we compared the relationship between the distribution of 26 bird species, which can be easily-detected either visually or by ear, and which belong to different taxonomic groups, with the distribution of the five most abundant palm species along the Purus-Madeira interfluve (Lepidocaryum tenue, Attalea speciosa, Mauritiella aculeata, Oenocarpus bataua, Oenocarpus bacaba). To determine whether the composition of bird assemblages is more strongly correlated with changes in palm-species abundance than with environmental factors (such as precipitation, water-table levels, sand and clay content) that commonly affect species distributions in this region, we also compared these relationships to the ecoregions proposed by Ximenes et al. (2021), who considered the region to be relatively heterogeneous. If such a relationship is found, palm species, which are sedentary and potentially easier to sample, could serve as cost-effective and efficient biodiversity surrogates for birds. Moreover, these palm species are abundant in the region and can be easily identified and monitored by non-specialists, such as those in local communities.
Methods
Study area
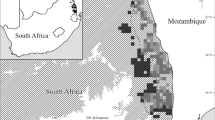
The study area is located between the Purus and Madeira rivers, in Brazilian Amazonia (Fig. 1). The northern part of the interfluve is comprised of three geomorphological units: alluvial terraces, where streams are dammed by the large rivers during the high-water season; mega-slopes, which are deeply dissected by stream meanders and flood temporarily after heavy rainfall; and mega-plateaus, which are higher, poorly drained areas featuring irregular microtopography and temporary pools (Brasil 1978). Vegetation type varies from dense forests in the north to more open, palm-dominated forests in the south (Brasil 1978). According to the ecoregions map by Olson et al. (2001), the Purus-Madeira interfluve region is dominated by the Purus-Madeira Moist Forests. For a more detailed classification incorporating bioclimatic, spatial, and topographic variations, see Ximenes et al. (2021).

Location and vegetation types of the Purus-Madeira interfluve in Brazilian Amazonia and data collection points along the BR-319 highway (M01-M11)
Sampling design
We collected data at 11 sites (M01 – M11, Fig. 1) along the BR-319 road, located in the Purus-Madeira interfluve. Sites were spaced roughly 60 km apart, forming a 670 km transect. At each site, a 5 km trail extended perpendicularly from the highway. Five 250 m-long transects were located at 1 km intervals along this trail, with the long axis of the plots following topographic contour lines. Bird data were collected at a point in the center of each transect and palm data were collected throughout each transect.
Water-table level, sand, and clay content were collected in situ and followed the sampling protocols applied by the Biodiversity Research Program in Western Amazonia (PPBio-AmOc). Water-table level was based on the mean of seven measurements of distance from the surface to ground water during the months of March, July and November 2011; March, August, October and December 2012; and March 2013. Positive values indicate that the water level was above ground. Soil samples (sand and clay content) were collected to a depth of 0.30 m every 50 m along the 250 m-long transects, giving six samples per transect. Sand and clay content in each plot was obtained following the standard protocol proposed by Donagema et al. (2011). Precipitation data were based on the annual precipitation (BIO12) obtained from the WorldClim database (Fick and Hijmans 2017).
The ecoregions proposed by Ximenes et al. (2021), which clustered the Purus-Madeira interfluve into 14 regions (Supplementary Information – Fig. S1), were based on latitude, altitude, drainage density, and soil characteristics. These variables were identified by the author as the most descriptive of environmental heterogeneity in the region relevant to biodiversity.
Due to the region’s rich bird diversity, we focused on 26 species (Supplementary Information – Table S1) easily-detected visually and by ear that covered a broad range of feeding guilds, body sizes, and use of different forest strata (canopy and understory). These species include nearly all guilds in the regional avifauna. We collected data between June and November 2010, a time of year when birds in the region reproduce and vocalize more frequently. Each bird-sampling point was visited for four consecutive days, and the order in which sites were visited was modified each day. Sampling each point repeatedly increased the probability of detecting species (Dorazio and Royle 2005; Mackenzie and Royle 2005), while sampling points in a different sequence each day removed the bias of detecting species that only call early in the morning. Between 5:30 AM and 9:30 AM, peak hours for bird activity, the same observer recorded all species detected via visual or auditory records at each sampling point for five minutes in an unlimited detection radius (Blake 1992; Vielliard 2000; Dorazio et al. 2006). Audio recordings were taken at all sampling points and deposited in the Sound Library (Acervo Sonoro) of the Bird Collection of the Instituto Nacional de Pesquisas da Amazônia.
We measured the abundance of five palm species that are often considered to be indicators of bird communities: Lepidocaryum tenue, Oenocarpus bataua, Oenocarpus bacaba, Mauritiella aculeata, and Attalea speciosa (Cohn-Haft et al. 2007). We collected palm-composition data in the same 250 m-long transects where the bird-sampling points were located. For clonal palm species, we considered each stem to be an individual.
Data analyses
We applied different statistical techniques to test whether the overall bird-species groups (all species, canopy birds, and understory birds) composition can be more accurately predicted by palm-species abundance (i.e., the five most abundant palm species), than environmental factors (precipitation, water-table level, sand, and clay contents) or ecoregions (Ximenes et al. 2021).
We fitted multivariate models based on the overall presence-absence data of bird-species, assuming that the response variable follows a binomial distribution, and used the “cloglog” link function to relate the predictors (palm-species abundance, environmental factors and ecoregions) to the response variable. To quantify the potential use of palm data as a substitute for bird assemblages, we compared the reduction in covariation among models. This metric is based on measurement of the sizes of the factor loadings and correlations in each model, estimated via the co-occurrence patterns using the Copula model (Popovic et al. 2022); a model-based approach that allows the construction of an ordination of the observations, where the taxa correlation is described using latent factors (i.e., unobserved variables that are inferred from the observed data) and modeled considering the data properties.
Additionally, we used Procrustes analysis(Jackson 1995; Peres-Neto and Jackson 2001) to statistically compare the similarity of overall bird-species ordination results with palm-species and environmental-factor ordinations. If such similarity is found, the superimposed ordination could potentially serve as effective biodiversity surrogates for birds. To characterize the variation in bird-species composition and palm-species abundance, we also used ordinations from the Copula model to visualize the main patterns in the multivariate datasets (Warton 2022; Popovic et al. 2022). For bird species, we fitted the multivariate models as previously described. For palm-species abundance, we used the same method, but assumed that the response variable follows a negative-binomial distribution and used the “log” link function. To summarize the variation in environmental factors, we used Principal Components Analysis (Pearson 1901). To assess the statistical significance of the Procrustes analysis, we used a permutation test (Number of permutation = 1000).
All analyses were undertaken in R version 4.2.2 (R Core Team 2022) using the packages ecoCopula (Popovic et al. 2019), mvabund version 4.2.1 (Wang et al. 2012) and vegan version 2.6-4 (Oksanen et al. 2022).
Results
The multivariate models fitted based on the overall presence-absence data for bird-species showed that palm-species abundance was the best predictor for the composition of all birds (BIC: 1266.24) and for canopy birds only (BIC: 704.98) (Table 1). For understory birds, environmental factors provided the best model (BIC: 684.69). All these models performed better than the ecoregions proposed by Ximenes et al. (2021). These results are qualitatively similar to those comparing the reduction of covariation among the analyses. The sizes of the factor loadings and correlations were in general lower for models with palm-species abundance as the predictor than other models, but only for all and canopy birds (Table 1); for understory birds, environmental factors were the best predictors of covariation in bird-species composition (Table 1).
The Procrustes analysis (Fig. 2; Table 2) showed that distributions of palm-species abundance (Supplementary Information – Fig. S2) could potentially serve as effective biodiversity surrogates for the distribution of bird-species (Supplementary Information – Fig. S3), but with a few limitations. The palm-species reflected the distributions of the dataset including all birds and only understory birds (Supplementary Information – Fig. S3). This analysis did not detect a significant relationship between the variation in the environmental factors (Supplementary Information – Fig. S2) and any bird functional group (Fig. 2; Table 2).

The relationships between the ordinations of bird-species, palm-species co-occurrence and environmental factors expressed by Procrustes analysis. The color represents the samples taken in dense forest (dark green) and open forest (light green) along the Purus-Madeira interfluve. The circles and diamonds represent the 250-m plots for birds and palms, respectively. The lines represent the connection between the target matrix and the rotated matrix in the analysis. The letters A to F represent the relationships between bird-species (i.e., target matrix), palm-species and environmental factors (i.e., rotated matrix): A) All birds and environmental factors; B) All birds and palm-species abundance; C) Canopy birds and environmental factors; D) Canopy birds and palm-species abundance; E) Understory birds and environmental factors; F) Understory birds and palm-species abundance
Discussion
Our results reveal an alternative and generally effective biodiversity surrogate in the Amazon that could support conservation efforts. This indicates that biotic interactions between two taxonomic groups may serve as a promising strategy to identify biodiversity surrogates. The observed variation in bird composition in the region was better predicted by palm-species abundance than environmental predictors or ecoregions. While this study did not test other surrogates that imply the study region to be homogeneous, they would be incapable of predicting variation in bird composition across the study area.
Palms may represent an important resource for vertebrates and invertebrates (Zona and Henderson 1989; Kahn and Granville 1992), especially when other forest resources are scarce (Peres 1994). Palms are also an important structural element in the region, where they can reach densities approximately four times higher than in other areas of Central Amazonia. The variation in palm composition between localities likely mirrors finer-scale environmental differences. Such local, fine-scale environmental variations, as indicated by palm composition, may have a more profound influence on bird populations than the environmental-surrogates-based on broad-scale-environmental data.
The importance of considering biotic interactions in studies for conservation planning should be emphasized, mainly because several studies have shown that biotic interactions (e.g., consumer–resource) have a significant effect on predicting the species distribution, often lowering the relative importance of environmental variables (Wang et al. 2018; Bazzato et al. 2023). Moreover, other studies have demonstrated that including biotic features can improve model performance to identify priority areas for conservation (Jenkins et al. 2020; Nogueira et al. 2023). The identification and comprehension of these relationships are urgent, because many conservations decisions guided by species distribution models (Guisan et al. 2013) underestimate the importance of biotic interactions in shaping species distributions.
Although there is a well-documented strong relationship, using palms as surrogates for birds in systematic conservation planning on a large scale in the Amazon has its limitations. Knowledge about the distribution and abundance of palm species is not available for the entire Amazon. However, alternative methods, such as remote sensing, have improved palm inventories and show promise as a strategy for understanding their distribution and diversity (Wagner et al. 2020). This strategy could highlight discrepancies between the actual number of species in a given area and the number expected based on habitat suitability, resources, and environmental conditions. Nonetheless, the taxonomy of palm species is relatively well understood, and local people are capable of conducting common palm species surveys efficiently.
Land-use planning is not only a central-government activity. Citizen scientists are often involved in data acquisition and subsequent planning (Garretson et al. 2023; Jesus et al. 2023), but they often do not have the experience to sample assemblages potentially consisting of hundreds of species. Involvement of local communities is facilitated if they can initially focus on species which they are already familiar with and use as resources, such as the palm species. The inclusion of these people in monitoring biodiversity could provide an inexpensive and potentially large labour force (Fa and Luiselli 2023). Initiatives that consider collaborative sampling by amateurs could be used in conjunction with traditional methods to evaluate the distribution of endangered species (Adamantopoulou et al. 2023) and conservation planning (Soteropoulos et al. 2021). The data provided by knowledgeable amateurs, could also reveal ecological patterns, enhancing our understanding in how biodiversity relates to environment (Devictor et al. 2010; Campbell et al. 2023; Garretson et al. 2023; Jesus et al. 2023). Furthermore, local people are often interested in sampling biodiversity in a purposeful manner, especially if the sample will contribute to scientific knowledge and decisions about land use (Thompson et al. 2023).
Given that the areas in our study have similar geomorphology, but showed different bird composition among the sites, using geomorphology as a surrogate to determine bird conservation priorities in the region will tend to underestimate the number of unique assemblages meriting protection. Even the ecoregions proposed by Ximenes et al. (2021), who considered the region to be relatively heterogeneous, are not good indicators of bird-species distributions. These surrogates frequently fail to track variation in conditions that are important for bird species, perhaps because such conditions vary at different spatial scales than the surrogates. Geomorphology and ecoregions may be useful for determining conservation priorities at large scales (Sano et al. 2019), but they can fail to capture between-site variation at fine scales, such as that captured by the biotic interaction between bird and palm species that we have studied.
In conclusion, identifying biodiversity surrogates, considering biotic interactions and incorporating local people in monitoring biodiversity programs, could be a useful tool for enhancing conservation and land-use planning across the Amazon basin and potentially in other parts of the world; especially when surrogates established based on assumptions about the environmental niches of most species have proved to be ineffective in predicting the species distributions at fine scales. Future studies should also incorporate other potential biotic interactions (Morales-Castilla et al. 2015) that are recognized to play a role in shaping the community composition and may have important consequences at an ecosystem level. Additionally, the reliability of conclusions regarding the identification of biodiversity surrogates may depend on both the taxonomic reorganization of species assemblages (e.g., into guilds) and the statistical methods employed. Taking these factors into consideration can improve our knowledge about biodiversity surrogates and could avoid inappropriate conservation recommendations.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Adamantopoulou S, Karamanlidis AA, Dendrinos P, Gimenez O (2023) Citizen science indicates significant range recovery and defines new conservation priorities for Earth’s most endangered pinniped in Greece. Anim Conserv 26:115–125. https://doi.org/10.1111/acv.12806
Baccaro FB, Rocha IF, del Aguila BEG et al (2013) Changes in ground-dwelling ant functional diversity are correlated with water-table level in an amazonian Terra Firme Forest. Biotropica 45:755–763. https://doi.org/10.1111/btp.12055
Bazzato E, Lallai E, Caria M et al (2023) Focusing on the role of abiotic and biotic drivers on cross-taxon congruence. Ecol Indic 151:110323. https://doi.org/10.1016/j.ecolind.2023.110323
Blake JG (1992) Temporal variation in Point counts of birds in a Lowland Wet Forest in Costa Rica. Condor 94:265–275. https://doi.org/10.2307/1368816
Bradfer-Lawrence T, Gardner N, Dent DH (2018) Canopy bird assemblages are less influenced by habitat age and isolation than understory bird assemblages in neotropical secondary forest. Ecol Evol 8:5586–5597. https://doi.org/10.1002/ece3.4086
Brasil (1978) Folhas SB 20 Purus: geologia, geomorfologia, pedologia, vegetação e uso potencial da terra. Ministério das Minas E Energia. Departamento Nacional de Produção Mineral (DNPM), Levantamento de Recursos Naturais, Rio de Janeiro
Brightsmith DJ, Cáceres A (2017) Parrots consume sodium-rich palms in the sodium‐deprived landscape of the Western Amazon Basin. Biotropica 49:921–931. https://doi.org/10.1111/btp.12479
Campbell J, Yakimishyn J, Haggarty D et al (2023) Fisheries (Bethesda) 48:8–19. https://doi.org/10.1002/fsh.10831. Citizen Science Surveys Provide Novel Nearshore Data
Caro T (2010) Conservation by Proxy: Indicator, Umbrella, Keystone, Flagship, and Other Surrogate Species
Carvalho RL, Resende AF, Barlow J et al (2023) Pervasive gaps in amazonian ecological research. Curr Biol. https://doi.org/10.1016/j.cub.2023.06.077
Cohn-Haft M, Pacheco A, Bechtoldt C et al (2007) Inventário ornitológico. Biodiversidade do médio Madeira. Bases científicas para propostas de conservação
Corcos D, Lami F, Nardi D et al (2021) Cross-taxon congruence between predatory arthropods and plants across Mediterranean agricultural landscapes. Ecol Indic 123:107366. https://doi.org/10.1016/j.ecolind.2021.107366
Cosentino F, Seamark ECJ, Van Cakenberghe V, Maiorano L (2023) Not only climate: the importance of biotic interactions in shaping species distributions at macro scales. Ecol Evol 13. https://doi.org/10.1002/ece3.9855
Covre AC, Lourenço-de-Moraes R, Campos FS, Benedito E (2022) Spatial relationships between fishes and amphibians: implications for conservation planning in a neotropical hotspot. Res Sq
da Glória CM, Tozetti AM (2021) Bird visits and resource use in Butia odorata (Arecaceae) palm groves in southern Brazil. Iheringia Ser Zool 111. https://doi.org/10.1590/1678-4766e2021032
Devictor V, Whittaker RJ, Beltrame C (2010) Beyond scarcity: citizen science programmes as useful tools for conservation biogeography. Divers Distrib 16:354–362. https://doi.org/10.1111/j.1472-4642.2009.00615.x
Donagema G, Campos D, Calderano S et al (2011) Manual de Métodos de Análise de Solo (vol 2). Embrapa, Embrapa Solos
Dorazio RM, Royle JA (2005) Estimating size and composition of Biological communities by modeling the occurrence of species. J Am Stat Assoc 100:389–398. https://doi.org/10.1198/016214505000000015
Dorazio RM, Andrew RJ, Söderström B, Glimskär A (2006) Estimating species Richness and Accumulation by modeling species occurrence and detectability. Source: Ecol 87:842–854. https://doi.org/10.1890/0012-9658(2006)87[842:ESRAAB]2.0.CO;2
Fa JE, Luiselli L (2023) Community forests as beacons of conservation: enabling local populations monitor their biodiversity. Afr J Ecol. https://doi.org/10.1111/aje.13179
Fattorini S, Dennis RLH, Cook LM (2012) Use of Cross-taxon congruence for hotspot identification at a regional scale. PLoS ONE 7:1–6. https://doi.org/10.1371/journal.pone.0040018
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int J Climatol 37:4302–4315. https://doi.org/10.1002/joc.5086
Gardner TA, Barlow J, Araujo IS et al (2008) The cost-effectiveness of biodiversity surveys in tropical forests. Ecol Lett 11:139–150. https://doi.org/10.1111/j.1461-0248.2007.01133.x
Garretson A, Cuddy T, Duffy AG, Forkner RE (2023) Citizen science data reveal regional heterogeneity in phenological response to climate in the large milkweed bug, Oncopeltus fasciatus. Ecol Evol 13. https://doi.org/10.1002/ece3.10213
Guisan A, Tingley R, Baumgartner JB et al (2013) Predicting species distributions for conservation decisions. Ecol Lett 16:1424–1435. https://doi.org/10.1111/ele.12189
Jackson DA (1995) PROTEST: a PROcrustean randomization TEST of community environment concordance. Écoscience 2:297–303. https://doi.org/10.1080/11956860.1995.11682297
Jenkins DA, Lecomte N, Andrews G et al (2020) Biotic interactions govern the distribution of coexisting ungulates in the Arctic Archipelago – a case for conservation planning. Glob Ecol Conserv 24:e01239. https://doi.org/10.1016/j.gecco.2020.e01239
Jesus LMG, Guedes JJM, Moura MR et al (2023) Environmental drivers of tropical forest snake phenology: insights from citizen science. Ecol Evol 13. https://doi.org/10.1002/ece3.10305
Kahn F, Granville J-J (1992) Palms in Forest ecosystems of Amazonia, Ecological studies. Springer-, Berlin
Lansac-Tôha FM, Heino J, Bini LM et al (2022) Cross-taxon Congruence of Taxonomic and functional Beta-diversity facets across spatial and temporal scales. Front Environ Sci 10:1–12. https://doi.org/10.3389/fenvs.2022.903074
Lima GR, Borges SH, Anciäes M, Cornelius C (2023) Ecomorphology and functional diversity of generalist and specialist bird assemblages in amazonian white-sand ecosystem habitat patches. Acta Amazon 53:141–153. https://doi.org/10.1590/1809-4392202203280
Lu M, Jetz W (2023) Scale-sensitivity in the measurement and interpretation of environmental niches. Trends Ecol Evol 38:554–567. https://doi.org/10.1016/j.tree.2023.01.003
Mackenzie DI, Royle (2005) Designing occupancy studies: general advice and allocating survey effort. J Appl Ecol 42:1105–1114. https://doi.org/10.1111/j.1365-2664.2005.01098.x
Marciente R, Bobrowiec PED, Magnusson WE (2015) Ground-Vegetation Clutter affects phyllostomid Bat assemblage structure in Lowland amazonian forest. PLoS ONE 10:e0129560. https://doi.org/10.1371/journal.pone.0129560
Margules CR, Pressey RL (2000) Systematic conservation planning. Nature 405:243–253. https://doi.org/10.1038/35012251
McGill BJ (2010) Matters of scale. Science (1979) 328:575–576. https://doi.org/10.1126/science.1188528
Monteiro J, Vieira C, Branquinho C (2023) Bryophyte assembly rules across scales. J Ecol 111:1531–1544. https://doi.org/10.1111/1365-2745.14117
Morales-Castilla I, Matias MG, Gravel D, Araújo MB (2015) Inferring biotic interactions from proxies. Trends Ecol Evol 30:347–356. https://doi.org/10.1016/j.tree.2015.03.014
Muscarella R, Emilio T, Phillips OL et al (2020) The global abundance of tree palms. Glob Ecol Biogeogr 29:1495–1514. https://doi.org/10.1111/geb.13123
Nogueira JG, Lopes-Lima M, Beja P et al (2023) Identifying freshwater priority areas for cross-taxa interactions. Sci Total Environ 864:161073. https://doi.org/10.1016/j.scitotenv.2022.161073
Oksanen J, Simpson GL, Blanchet FG et al (2022) vegan: community ecology package. R Package Version 2.6-2. https://CRAN.Rproject.org/package=vegan
Olson DM, Dinerstein E, Wikramanayake ED et al (2001) Terrestrial ecoregions of the World: a New Map of Life on Earth. Bioscience 5:933–938. https://doi.org/10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Pakulnicka J, Buczyńska E, Buczyński P et al (2015) Are beetles good indicators of insect diversity in freshwater lakes? Oceanol Hydrobiol Stud 44:487–499. https://doi.org/10.1515/ohs-2015-0046
Pearson K (1901) LIII. On lines and planes of closest fit to systems of points in space. Lond Edinb Dublin Philosophical Magazine J Sci 2:559–572. https://doi.org/10.1080/14786440109462720
Peixoto G, Leitão P, Kaefer IL, Lima AP (2019) The lizards along the road BR-319 in the Purus-Madeira interfluve, Brazilian Amazonia (Squamata, Lacertilia). Herpetol Notes 12:689–697
Peres CA (1994) Composition, density, and fruiting phenology of Arborescent palms in an amazonian Terra Firme Forest. Biotropica 26:285–294
Peres CA (2005) Why we need megareserves in Amazonia. Conserv Biol 19:728–733. https://doi.org/10.1111/j.1523-1739.2005.00691.x
Peres-Neto PR, Jackson DA (2001) How well do multivariate data sets match? The advantages of a procrustean superimposition approach over the Mantel test. Oecologia 129:169–178. https://doi.org/10.1007/s004420100720
Pinto MP, Diniz-Filho JAF, Bini LM et al (2008) Biodiversity surrogate groups and conservation priority areas: birds of the Brazilian Cerrado. Divers Distrib 14:78–86. https://doi.org/10.1111/j.1472-4642.2007.00421.x
Popovic GC, Warton DI, Thomson FJ et al (2019) Untangling direct species associations from indirect mediator species effects with graphical models. Methods Ecol Evol 10:1571–1583. https://doi.org/10.1111/2041-210X.13247
Popovic GC, Hui FKC, Warton DI (2022) Fast model-based ordination with copulas. Methods Ecol Evol 13:194–202. https://doi.org/10.1111/2041-210X.13733
Qian H, Kissling WD (2010) Spatial scale and cross-taxon congruence of terrestrial vertebrate and vascular plant species richness in China. Ecology 91:1172–1183. https://doi.org/10.1890/09-0620.1
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing,Vienna. https://www.R-project.org/
Salomão RP, Pires D, de Baccaro A FB, et al (2022) Water table level and soil texture are important drivers of dung beetle diversity in amazonian lowland forests. Appl Soil Ecol 170:104260. https://doi.org/10.1016/j.apsoil.2021.104260
Sano EE, Rodrigues AA, Martins ES et al (2019) Cerrado ecoregions: a spatial framework to assess and prioritize Brazilian savanna environmental diversity for conservation. J Environ Manage 232:818–828. https://doi.org/10.1016/j.jenvman.2018.11.108
Schietti J, Martins D, Emilio T et al (2016) Forest structure along a 600 km transect of natural disturbances and seasonality gradients in central-southern Amazonia. J Ecol 104:1335–1346. https://doi.org/10.1111/1365-2745.12596
Snow DW (1981) Tropical Frugivorous birds and their food plants. A World Survey
Soteropoulos DL, De Bellis CR, Witsell T (2021) Citizen Science contributions to address Biodiversity loss and Conservation Planning in a rapidly developing region. Divers (Basel) 13:255. https://doi.org/10.3390/d13060255
Sousa TR, Schietti J, Coelho de Souza F et al (2020) Palms and trees resist extreme drought in Amazon forests with shallow water tables. J Ecol 108:2070–2082. https://doi.org/10.1111/1365-2745.13377
Spigoloni ZA, Bernardy JV, Schlemmer Brasil L et al (2022) Odonata Concordance amongst aquatic taxa in Brazilian savanna streams. Int J Odonatol 25:80–88. https://doi.org/10.48156/1388.2022.1917178
Stegmann LF, Leitão RP, Zuanon J, Magnusson WE (2019) Distance to large rivers affects fish diversity patterns in highly dynamic streams of Central Amazonia. PLoS ONE 14:e0223880. https://doi.org/10.1371/journal.pone.0223880
Thompson MM, Moon K, Woods A et al (2023) Citizen science participant motivations and behaviour: implications for biodiversity data coverage. Biol Conserv 282:110079. https://doi.org/10.1016/j.biocon.2023.110079
van der Hoek Y, Álvarez Solas S, Peñuela MC (2019) The palm Mauritia flexuosa, a keystone plant resource on multiple fronts. Biodivers Conserv 28:539–551. https://doi.org/10.1007/s10531-018-01686-4
Verdinelli M, Pittarello M, Caria MC et al (2022) Congruent responses of vascular plant and ant communities to pastoral land-use abandonment in mountain areas throughout different biogeographic regions. Ecol Process 11:35. https://doi.org/10.1186/s13717-022-00379-9
Vielliard JME (2000) Bird community as an indicator of biodiversity: results from quantitative surveys in Brazil. Acad Bras Cienc 72:323–330. https://doi.org/10.1590/S0001-37652000000300006
Wagner FH, Dalagnol R, Tagle Casapia X et al (2020) Regional Mapping and spatial distribution analysis of Canopy palms in an Amazon Forest using deep learning and VHR images. Remote Sens (Basel) 12:2225. https://doi.org/10.3390/rs12142225
Wang Y, Naumann U, Wright ST, Warton DI (2012) ) < tt > mvabund - an < tt > R package for model-based analysis of multivariate abundance data. Methods Ecol Evol 3:471–474. https://doi.org/10.1111/j.2041-210X.2012.00190.x
Wang F, Zhao Q, McShea WJ et al (2018) Incorporating biotic interactions reveals potential climate tolerance of giant pandas. Conserv Lett 11:e12592. https://doi.org/10.1111/conl.12592
Warton DI (2022) Eco-stats: Data Analysis in Ecology. Springer International Publishing, Cham
Wiens JA, Hayward GD, Holthausen RS, Wisdom MJ (2008) Using Surrogate species and groups for Conservation Planning and Management. Bioscience 58:241–252. https://doi.org/10.1641/B580310
Ximenes AC, Amaral S, Monteiro AMV et al (2021) Mapping the terrestrial ecoregions of the Purus-Madeira interfluve in the Amazon Forest using machine learning techniques. Ecol Manage 488. https://doi.org/10.1016/j.foreco.2021.118960
Zona S, Henderson A (1989) A review of animal-meidated seed dispersal of palms. Selbyana 11:6–21
Acknowledgements
We would like to thank all local people who supported us on field data collection along the BR-319 and extend special thanks to Mario Cohn-Haft for advising during bird data collection and for insisting that palm species would be good indicators of Amazonian bird habitats. Our sample effort would not be possible without the PPBio network infrastructure, guidance and technical support. We thank the Long-Term Ecological Research Program in the Southwest of Amazonas (PELD – PSAM) for providing the infrastructure that allows sampling along the BR-319 and collaboration among researchers in different institutions. Permits for field research were granted by the Instituto Chico Mendes de Conservação da Biodiversidade (birds permit Nº 21421-2, palms permit Nº 26079-1).
Funding
Brazilian Science Funding Agency CNPq awarded stipends to authors J.M. and T.E. Bird data collection was financed by PRONEX (003/2009), a partnership between CNPq and State of Amazonas Research Foundation (FAPEAM). Palm data collection was financed by PRONEX (006/2006), Hidroveg Universal CNPq (473308/2009-6), Projeto Cenários FINEP/CNPq (52.0103/2009-2) and FAPESP/FAPEAM (465/2010). This work would not be possible without the infrastructure and support of CENBAM (Center for Integrated Studies of Biodiversity in the Amazon, MCT/CNPq 015/2008) and PPBio (Research Program in Biodiversity, CNPq 60/2009 and 558318/2009-6). Bird field equipment was donated by Idea Wild. Author S.S.J. is supported by Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM/PROFIX – RH – Edital/009/2021). Author T.E. is supported by Centro de Pesquisa em Biodiversidade e Mudanças Climáticas (CBICLIMA FAPESP #2021/10639-5).
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
All authors contributed to the conception and design of the study. Material preparation and data collection were executed by Juliana Menger and Thaise Emilio. Data analysis was carried out by Sergio Santorelli Junior, with statistical contributions from William E. Magnusson. The first draft of the manuscript was written by Juliana Menger. All authors provided critical revisions and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by Suraj Upadhaya.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Menger, J., Santorelli Junior, S., Emilio, T. et al. Palms predict the distributions of birds in southwestern Amazonia and are potential surrogates for land-use planning by citizen scientists. Biodivers Conserv (2024). https://doi.org/10.1007/s10531-024-02895-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10531-024-02895-w