
Abstract
The cherry laurel (Prunus laurocerasus) is a very popular garden plant and has been widely reported as a garden escape in Central Europe. However, it is still considered an unestablished neophyte in Germany. Few critical data are available on local population sizes, life history or reproductive biology outside its natural range. We carried out a study with 24 transects of 500 m × 6 m in the Kottenforst near Bonn, western Germany to fill these knowledge gaps. Along these transects we recorded the plant number and plant sizes and analysed their age structure. We found Prunus laurocerasus along 19 of 24 transects with up to 63 individual per transect. The largest plants covered areas of up to 50 m2 and were up to 30 years old. However, most of the plants were less than 20 years old and established since 2000. Since we observed high fruit set and old plants surrounded by many seedlings we conclude that Prunus laurocerasus has to be considered as an established neophyte in our study area and has invasive potential.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Invasive species are amongst the largest threats to biodiversity because these species have the potential to disrupt established assemblages and may lead to declining populations of less competitive species being replaced by exotics (Gurevitch and Padilla 2004). Many exotic plant species, such as Impatiens glandulifera (Balsaminaceae) or Solidago canadensis (Asteraceae), had been cultivated in gardens, in some cases for centuries, before they started establishing populations in the wild relatively recently (Zäch 2005; Gregor et al. 2012; Hetzel 2012; Scherrer et al. 2022). There has been a dramatic increase in the abundance of neophytes in Germany, and probably also in other Central European countries, since 1990 (Eichenberg et al. 2021). These colonisations occurred mainly in open and disturbed habitats (Scherrer et al. 2022). However, recent studies indicate that forests in Central Europe are also affected (Essl et al. 2011, 2012). For example, thermophilous, neophytic shrub species with evergreen, laurophyllous leaves have started to spread from gardens, a process called laurophyllisation (Walther 2000). The first evidence of these invasions dates back to 1956 in the forests of Ticino, Switzerland and adjacent northern Italy (Walther 1999). Today, the herb and shrub layers of some forests in this region are already dominated by exotic, evergreen species, such as Laurus nobilis, Prunus laurocerasus and Trachycarpus fortunei (Zäch 2005). These neophytes compete with native species and can have a negative impact on regeneration (Langmaier and Lapin 2020).
North of the Alps, the colonisation by laurophyllous species started a few decades later, in the 1980s (Brandes 2011; Nehring et al. 2013a).A number of laurophyllous, neophytic shrub species spread into forests situated close to settlements in many Central European regions (e.g. Hetzel 2012; Nehring et al. 2013a) as previously reported from southern Switzerland. However, it has remained largely unclear whether neophytic, laurophyllous shrub species establish stable, self-sustaining populations north of the Alps (Nehring et al. 2013a).
Cherry laurel (Prunus laurocerasus; Rosaceae) is a typical laurophyllous shrub species and particularly popular as a hedging plant in European gardens. Rusterholz et al. (2018) documented several established populations of cherry laurel in the forests of northern Switzerland, surviving for at least 17 years. Prunus laurocerasus is a fast-growing, robust, evergreen shrub or small tree up to 18 m tall in its native range, which extends from southeastern Europe to western Asia, particularly northern Turkey and western Georgia (Berger et al. 2007; Nakhutsrishvili 1999). In central Europe, the species is usually spreading, with older individuals shading out large areas of the forest floor with their dense foliage (Fig. S1A; pers. obs. by the authors; Rusterholz et al. 2018).
Rusterholz et al. (2018) documented a lower diversity of native herb and shrub species in forest plots associated with cherry laurel. Additionally, plots with older Prunus laurocerasus individuals had lower values in soil moisture, soil pH and phosphorus availability (Rusterholz et al. 2018). Vegetative plant parts of Prunus laurocerasus are largely avoided by herbivores because they contain toxic cyanogenic glycosides, potentially giving them a competitive advantage over more palatable plants. Prunus laurocerasus may thus have a negative impact on the ecosystem, but it is still considered as a non-established neophyte in Germany (Lauterbach and Nehring 2013) and neither listed on the “Warnliste invasiver Gefäßpflanzenarten in Deutschland” (Nehring et al. 2013a) nor on the “List of invasive alien species of Union concern” of the EU (EU document 32022R1203). In order to investigate the establishment of Prunus laurocerasus in Central Europe—which is the basis for ranking a species as invasive in a second step—it is necessary to document stable populations and reproduction in the wild (Nehring et al. 2013b). In Belgium Prunus laurocerasus is already on the watch list (www.ias.biodiversity.be/species/show/112) and in Switzerland it is even listed on the black list of invasive species that can not be traded anymore (www.infoflora.ch/de/neophyten/listen-und-infoblätter.html#liste-der-invasiven-neophyten-der-schweiz).
As Rusterholz et al. (2018) mainly focused on the competition between Prunus laurocerasus and native herbs and shrubs in their study in forests around Basel, they did not evaluate details of population size and structure. To further investigate the status of establishment of Prunus laurocerasus we conducted a study along 24 transects in a forest in western Germany. Along these transects we analysed: (1) How common is the cherry laurel in the forest? (2) Is there a gradient of abundance indicating an invasion from closed settlements into the forest? (3) What is the age structure of the cherry laurel in the forest and is it already generatively reproducing? (4) How large have individual plants already grown in the forest?
Material and methods
Study site
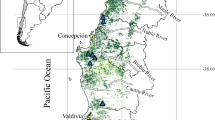
The Kottenforst is a forest complex, predominantly deciduous broadleaved forests of oak and beech with some conifers (e.g., spruce and douglas fir) southwest of Bonn, Germany, covering ⁓40 km2 (Krause and Vanberg 1997; Fig. 1). In terms of plant species composition and vegetation types this area can be considered as a typical example of a forest in western Germany as well as in neighbouring regions due to its climatic conditions and vegetation composition (Krause and Vanberg 1997). The forest is directly adjacent to residential areas of suburban Bonn and neighbouring villages with more or less extensive private gardens. Thus, it is appropriate for our study.

Overview map of the study area (Kottenforst, shown in green) south-west of Bonn (settlements shown in grey). Black bars represent the 24 study transects (500 m × 6 m); white dots represent documented individuals of Prunus laurocerasus; dark green forest area represent a 500 m boundary area to the next settlement area. Map projection: Universal Transverse Mercator (zone 32N); EPSG: 25,832
The Kottenforst lies at an elevation of 160 m to 190 m a.s.l. on loess loam. The mean annual temperature (1960–2021) is 10.2 °C, but comparing the mean annual temperatures between 1960–1999 and 2000–2021 there has been an increase of ⁓1 °C (9.8 °C vs. 10.8 °C; weather station Weilerswist Lommersum, www.cdc.dwd.de), paralleled by similar increases in mean monthly temperatures. The mean annual precipitation (1960–1999) is 607 mm, but comparing the mean annual precipitation between 1960–1999 and 2000–2021, precipitation has decreased by 52 mm (624 mm vs. 572 mm). The decrease in precipitation is most pronounced in spring, with up to 16 mm (− 33.2%) less precipitation in April.
Population density and structure
To collect data on the abundance of Prunus laurocerasus in the Kottenforst we recorded individuals along 24 transects, 500 m long and 6 m wide, in February 2022. All transects started at the edge of the forest next to the residential areas and continued into the forest at a 90° angle. As the forest edge is not running in a straight line we determined that at each point on all transects, the distance to other settlements was at least the distance to the starting point of the individual transect. The distance between transects was at least 300 m. We designed transects meeting all of the described characteristics using google earth aerial photos without visiting transects before. In three cases, transects crossed major roads, such as the highway A565 southeast of Röttgen (Fig. 1). Gardens 200 m around the starting point of each transect were visually inspected for planted Prunus laurocerasus plants without entering them and the approximate number of individuals per garden was recorded.

A Distribution of Prunus laurocerasus individuals in relation to the forest edge. B Distribution of cover classes (m2) among Prunus laurocerasus individuals in the Kottenforst. The maximum cover of each class is given by the number below the columns, the minimum cover by the number of the next lower class
Vegetative traits
For each individual of Prunus laurocerasus observed on the transects, we measured the height, the length and width of the plant, as well as the circumference of the main stem at 5 cm above the ground. We define the maximum length and width of a plant as the length and width observed from a bird’s eye view (Fig. S2). We recorded the distance to the forest edge using GPS. Near each transect, we cut five individuals of Prunus laurocerasus at 5 cm above the ground, and took a 15 cm piece of the main stem for later analysis. Stem circumference was also measured for these individuals. The investigated individuals covered the entire range of stem circumferences observed on the transects. To estimate the age of the collected Prunus laurocerasus stems we photographed each stem under a stereo microscope and counted the annual growth rings on the photographs.
At each transect we collected at least one herbarium specimen to document the morphological diversity of Prunus laurocerasus in the Kottenforst. Specimens are stored in the herbarium of the University of Bonn (BONN; herbarium numbers JO-01-02-22 to JO-16-02-22).
Reproductive traits and seed set
In late April to early May 2022, all transects were visited again and the number of flowering individuals of Prunus laurocerasus was recorded. Along twelve transects we marked a total of 34 plant individuals and counted the number of inflorescences/infructescences. We then counted the number of flowers/fruits on ten randomly selected inflorescences/infructescences of 31 of these individuals. At the end of July 2022, the transects were revisited and the number of developing fruits counted at the marked infructescences.
Total population size in the Kottenforst
To visualise the forest area, the study transects, the observed individuals and to estimate the total population size of Prunus laurocerasus in the Kottenforst, the Basis-DLM of the state of North Rhine-Westphalia (www.opengeodata.nrw.de) was used. The area of the Kottenforst was manually extracted from the forest layer of the Basis-DLM data set and the different layers for the settlements were visually inspected and merged into one layer combining all settlements. In order to visualise those sections of the forests that are up to 500 m away from the settlement edges, the boundary area was calculated using the settlement layer and intersected with the Kottenforst layer. Subsequently, the total area as well as the area of the 500 m forest boundary was calculated using the QGIS field calculator tool. The geodata were analysed and visualised using QGIS v.3.28 (QGIS.org 2023).
Statistical analyses
We used bar charts to document the number of Prunus laurocerasus individuals observed on the transects. By dividing the transects into ten subsections of 50 m, we documented the number of plant individuals and the distance of the individuals from the forest edge along the transects using bar charts. We also documented the age distribution of the plants, again using bar charts, showing the numbers of individual within five year subsections. To analyse the vegetative plant traits, including plant height, the length and width of the plant and the circumference of the main stem we calculated the mean and standard deviation for each trait separately. To obtain a rough estimate of the area covered by individual Prunus laurocerasus plants, we applied the standard formula for calculating the area of an oval:
By conducting regression analyses of plant age with main stem circumference, plant height and plant area, we tested for a vegetative trait that could be used to estimate plant age without counting stem rings. Applying a Spearman rank correlation (due to the non-normality of the data) we correlated the main stem circumference against the distance to the forest edge to check for a spatial age structure. Instead of using plant age we used the main stem circumference as a proxy because we have more data points for this trait covering the entire transects. Finally, we conducted a regression analysis between the number of Prunus laurocerasus individuals found on the transects and the number of individuals observed in the gardens close to the starting points of the individual transects. All statistical analyses were done in R (Development Core Team 2018).
Results
Population density, age structure and total population size
Prunus laurocerasus was observed in 19 out of 24 investigated transects. In total, 277 individual plants were documented, with a mean of 11.5 (± 17.2 [SD]) individuals per transect. On 10 transects only five or less individuals were detected. Conversely, on other transects up to 63 individuals were recorded. In the three cases, where transects crossed major roads, such as the A565, no individuals were found behind these roads. Most individuals were found within the first 50 m of the transects, and the number of Prunus laurocerasus individuals decreased with distance from the settlements. In a single case an individual of Prunus laurocerasus was found 500 m into the forest (Fig. 2A). A second peak in abundance was found between 200 and 249 m distance from the edge of the forest and a third peak between 300 and 349 m. These two peaks can be explained by our observation of individual mature plants surrounded by numerous seedlings and young plants. We found three cases where individual plants apparently established themselves vegetatively from twigs illegally discarded from gardens. In gardens close to starting points (with a maximum distance of 200 m) of 23 out of 24 of the transects we observed up to ⁓400 individuals (mean 189.5 ± 107.3 [SD]) of Prunus laurocerasus. The number of individuals in the gardens were positively related to the number of individuals found on the transects (R2 = 0.181, p = 0.022).
In total, we counted the growth rings of 71 Prunus laurocerasus individuals. Approximately 25.4% of the plants were up to 5 years old, 22.5% were between 6 and 10 years old, 28.2% between 11 and 15 years old, 12.7% between 16 and 20 years old, 4.2% between 21 and 25 years old and the remaining 7% between 26 and 30 years old. The oldest extant individuals in the study area dated to 1991, but the vast majority of individuals established themselves after 2006. Along the transect we found no linear age structure, i.e. a significant relation between plant age and distance from the forest edge (Spearman rank correlation, R = 0.0299, p = 0.6619).
The Kottenforst covers a forest area of approx. 45.36 km2 (45,356,960 m2), and 55.4% or 25.11 km2 (25,105,557 m2) lies within a 500 m range from the next settlement. Based on the mean number of Prunus laurocerasus individuals observed in the total area of the 24 transects (72,000 m2 in total) we estimated a total of 96,587 individuals in the 500 m boundary area closest to settlements (Fig. 1).
Vegetative traits
We measured vegetative traits, including plant height, length and width, as well as the circumference of the main stem of 221 Prunus laurocerasus individuals. The largest cherry laurel individuals in the Kottenforst are up to 4.8 m (mean 0.88 m ± 0.59 m [SD]) tall, with a stem circumference of up to 27 cm at 5 cm above the ground (mean 5.4 cm ± 3.5 cm [SD]), a maximum plant length of up to 8 m (mean 1.56 m ± 1.07 m [SD]) and a maximum plant width of 8 m (mean 1.23 m ± 0.86 m [SD]). The maximum area covered by a single plant was 50.3 m2 (mean 3.7 m2 ± 2.6 m2 [SD]; Fig. 2B) and the maximum total area covered by the species per transect was 244.4 m2 (8.13% of a total transect area). In the overgrown area we observed no other undergrowth species.
Main stem circumference, plant height and plant area are all positively correlated with plant age. The strongest correlation is between plant age and stem circumference (R2 = 0.767, p < 0.001; N = 71; Fig. S3), the correlations between plant age and plant height (R2 = 0.741, p < 0.001; N = 37) and area covered by the plants (R2 = 0.481, p < 0.001; N = 37) are similarly strong.
Reproductive traits
Of the 277 recorded individuals of Prunus laurocerasus in the Kottenforst, 100, mainly medium-sized to large individuals, were flowering. On these plants we observed an average of 218 (± 175 [SD]) inflorescences with a mean of 25.1 (± 6.9 [SD]) flowers each. When we visited the plants again in July, 77 plants were fruiting and we counted a mean of 83.9 (± 100.4 [SD]) infructescences per plant. Each infructescence produced a mean of 6.9 (± 6.1 [SD]) fruits.
Discussion
In this study, we analysed population structure and seed set of cherry, laurel Prunus laurocerasus, in the Kottenforst near Bonn, Germany. We demonstrate that cherry laurel is a common neophyte in the Kottenforst. Up to 97,000 individuals may be present within the 500 m boundary area close to settlements. Here, cherry laurel locally reaches considerable abundances with up to 63 plants per transect. However, due to variation in seed influx from adjacent gardens and differences in forest management the abundance of Prunus laurocerasus among transects may show considerable variation, but is still positively related to the number of individuals in the gardens. Individual plants are up to 30 years old and cover areas of up to 50 m2, but the majority of plants established themselves after 2000. The increased establishment of laurel cherry after 2000 may have been facilitated by the increased winter temperatures since then (www.cdc.dwd.de). Larger plants were surrounded by numerous seedlings and young plants leading to the conclusion that Prunus laurocerasus successfully reproduces in the forest, which clearly indicates that cherry laurel must be categorized as an established neophyte (Nehring et al. 2013b). Similar reports on the occurrence and local reproduction of Prunus laurocerasus come from observational data from other regions of western Germany (e.g. Brandes 2011; Hetzel 2012) as well as from a single other systematic study from adjacent Switzerland (Rusterholz et al. 2018), indicating that our local study documents a general trend.
Prunus laurocerasus is currently most abundant within the first 50 m of the forest edge, but individual plants are found up to 500 m deep in the forest. Surprisingly, we did not find a spatial age structure across the transects. This distribution is typical of alien species invading forests from the periphery (Essl et al. 2012). A similar distribution pattern has already been observed in the province of Varese, northern Italy (pers. comm. Michael Kleih, author of the Flora of the Lago Maggiore area). There, many plants have reached flowering and fruiting size, probably facilitating the further spread of cherry laurel into the forest. In the Kottenforst no other undergrowth species were observed in the overgrown areas, as the spreading habit and the dense, evergreen foliage of the laurel cherry appears to suppress all other plant species. Considering that about half of the plants observed were only a few years old—covering less than 1 m2—Prunus laurocerasus is likely to change overall vegetation (Langmaier and Lapin 2020).
Our data document that Prunus laurocerasus has very successfully colonized the Kottenforst and will spread more deeply into the forest, since the early colonizers already reached reproductive age and successfully fruit. The success of this exotic plant may—at first glance—appear puzzling. But, the fossil record indicates that Prunus laurocerasus respectively its fossil equivalent is part of the western European Pliocene palaeo-forest flora (Kvaček et al. 2020). These forests were characterized by the prevalence of tree genera such as Castenea, Carpinus and Fagus. In its present-day relict range cherry laurel is found in, e.g., Fagus orientalis (Akhalkatsi 2015) and Castanea sativa forests (Goginashvili et al. 2021). It is here associated with more or less the same tree, shrub and herb genera and sometimes species as found in the Kottenforst either as native components (Carpinus, Fagus, Ilex, Fragaria, Athyrium, Sanicula) or already successfully established taxa (Castanea, Taxus). Prunus laurocerasus—once introduced—thus finds itself in an ecosystem that is a close equivalent of the one it spent its early evolutionary history in and strikingly similar to its native relic habitat. In this context, the successful colonization of the Kottenforst is not particularly surprising.
Conclusions
Our study documents that Prunus laurocerasus is common. Established populations reproduce generatively in the study area in western Germany. On individual transects it has become the dominant understorey species and we observed an ongoing colonisation process of areas deeper in the forest. Thus, Prunus laurocerasus has to be considered as an established neophyte for the region. From both an ecological and palaeoecological perspective, it is colonising a forest community that closely resembles its ancestral, Pliocene habitat. Due to its excellent adaptation to the given ecology it has a high invasive potential and is likely to permanently alter forest structure and species composition.
References
Akhalkatsi M (2015) Forest habitat restoration in Georgia, Caucasus ecoregion. Mtsignobari, Tbilisi, p 101
Berger S, Söhlke G, Walther G-R, Pott R (2007) Bioclimatic limits and range shifts of cold-hardy evergreen broad-leaved species at their northern distributional limit in Europe. Phytocoenologia 37:523–539
Brandes D (2011) Neufunde von Neophyten im Stadtgebiet von Braunschweig. Braunschweiger Naturkundliche Schriften 10:79–96
Eichenberg D, Bowler DE, Bonn A, Bruelheide H, Grescho V, Harter D, Jandt U, May R, Winter M, Jansen F (2021) Widespread decline in Central European plant diversity across six decades. Glob Change Biol 27:1097–1110
Essl F, Milasowszky N, Dirnböck T (2011) Plant invasions in temperate forests: resistance or ephemeral phenomenon? Basic Appl Ecol 12:1–9
Essl F, Mang T, Moser D (2012) Ancient and recent alien species in temperate forests: steady state and time lags. Biol Invasions 14:1331–1342
Goginashvili N, Togonidze N, Tvauri I, Manvelidze Z, Memiadze N, Zerbe S, Asanidze Z (2021) Diversity and degradation of the vegetation of mountain belt forests of central Adjara (the Lesser Caucasus), Georgia. J for Sci 67:219–241
Gregor T, Bönsel D, Starke-Ottich I, Zizka G (2012) Drivers of floristic change in large cities–a case study of Frankfurt/Main (Germany). Landsc Urban Plan 104:230–237
Gurevitch J, Padilla DK (2004) Are invasive species a major cause of extinctions? Trends Ecol Evol 19:470–474
Hetzel I (2012) Ausbreitung klimasensitiver ergasiophygophytischer Gehölzsippen in urbanen Wäldern im Ruhrgebiet. Dissertationes Botanicae 411:1–205
Krause S, Vanberg C (1997) Flora und Vegetation des Kottenforstes bei Bonn. Teil: 1 Naturräumliche Grundlagen und Flora. Decheniana 150:169–189
Kvaček Z, Teodoridis V, Denk T (2020) The Pliocene flora of Frankfurt am Main, Germany: taxonomy, palaeoenvironments and biogeographic affinities. Palaeobiodivers Palaeoenviron 100:647–703
Langmaier M, Lapin K (2020) A systematic review of the impact of invasive alien plants on forest regeneration in European temperate forests. Front Plant Sci 11:524969
Lauterbach D, Nehring S (2013) Naturschutzfachliche Invasivitätsbewertung Prunus laurocerasus–Lorbeerkirsche; erstellt 30.06.2013. Bundesamt für Naturschutz, Bonn, p 2
Nakhutsrishvili G (1999) Evergreen broad-leaved vegetation in the Colchis. In: Klotzli F, Walther G-R (eds) Recent shifts in vegetation boundaries of deciduous forests, especially due to general global warming. Birkhauser, Basel, pp 167–179
Nehring S, Kowarik I, Rabitsch W, Essl F (2013a) Naturschutzfachliche Invasivitätsbewertungen für in Deutschland wild lebende gebietsfremde Gefäßpflanzen. BfN-Skripten 352:1–202
Nehring S, Essl F, Rabitsch W (2013b) Methodik der naturschutzfachlichen Invasivitätsbewertung für gebietsfremde Arten. Version 1.2. BfN-Skripten, p 340
R Development Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rusterholz HP, Schneuwly J, Baur B (2018) Invasion of the alien shrub Prunus laurocerasus in suburban deciduous forests: effects on native vegetation and soil properties. Acta Oecologica 92:44–51
Scherrer D, Bürgi M, Gessler A, Kessler M, Nobis MP, Wohlgemuth T (2022) Abundance changes of neophytes and native species indicate a thermophilisation and eutrophisation of the Swiss flora during the 20th century. Ecol Ind 135:108558
Walther G-R (1999) Distribution and limits of evergreen broad-leaved (laurophyllous) species in Switzerland. Bot Helv 109:153–167
Walther G-R (2000) Climatic forcing on the dispersal of exotic species. Phytocoenologia 30:409–430
Zäch R (2005) Ökologie und Ausbreitung von Neophyten auf dem Monte Caslano im Südtessin. Master’s thesis, ETH, Eidgenössische Technische Hochschule Zürich
Acknowledgements
We thank the Amt für Umwelt, Verbraucherschutz und Lokale Agenda of Bonn and the Regionalforstamt Rhein-Sieg-Erft for the permission to conduct our study in the Kottenforst. We further thank Dr. Jens Mutke (Nees Institute, University of Bonn) for helpful comments on the GIS analyses and Eliane Furrer-Abrahamczyk for the permission to use the photo of Prunus laurocerasus.
Funding
Open Access funding enabled and organized by Projekt DEAL. The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation and data collection was performed by Johanna Otto. Analyses were performed by Stefan Abrahamczyk and Tim Böhnert. The first draft of the manuscript was written by Stefan Abrahamczyk. All authors commented on previous versions of the manuscript and read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abrahamczyk, S., Otto, J., Böhnert, T. et al. Naturalization of Prunus laurocerasus in a forest in Germany. Biol Invasions (2024). https://doi.org/10.1007/s10530-024-03325-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10530-024-03325-2