
Abstract
Invasive plant species can drive ecosystem change, particularly on oceanic islands that are vulnerable to plant invasions and subsequent biodiversity loss. While invasive species management is vital for habitat restoration, efficacy of management efforts and the ability of native plants to regenerate varies among studies. The aim of this study was to examine the consequences of managing a thicket-forming woody plant species—Psidium cattleyanum Sabine (strawberry guava)—on subtropical forest regeneration, comparing spatial scales and management periods. We surveyed 15 locations on Norfolk Island, an isolated Pacific Island with a high proportion of endemic and threatened species, by establishing paired managed and unmanaged plots to assess changes in species-area relationships, abundance, richness, and composition of the plant community. Total plant richness was higher in managed plots at the largest scale examined (125 m2). However, there was no significant difference between managed and unmanaged plots in the slopes of species-area relationships when canopy, understory, and seedling strata were combined. Stratum-specific analyses revealed that management-driven changes were most evident in the subcanopy strata, which had significantly higher woody plant abundance and richness in managed plots. Compositional dissimilarity between managed and unmanaged plots was greater in locations with higher moisture levels, suggesting that post-management regeneration is related to environmental conditions. This study suggests that P. cattleyanum suppresses plant recruitment and its removal allows for forest regeneration, indicating that management of dominant invasive woody species facilitates plant recruitment on Norfolk Island if nearby native propagule sources are present.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Plant invasions can drive biodiversity declines and contribute to habitat degradation and ecosystem change (Hejda et al. 2009; Florens et al. 2016). Multiple studies have reported lower native plant richness or diversity after plant invasions, along with limited recruitment of native species (Mascaro et al. 2008; Jäger et al. 2009; Pyšek et al. 2012; Florens et al. 2017). Invasive plant species are more likely to negatively affect native species richness on islands than mainlands (Pyšek et al. 2012), as islands tend to have higher endemism, smaller population sizes, more available niche space, and therefore a relatively higher extinction risk (Caujapé-Castells et al. 2010; Kueffer et al. 2010; Ren et al. 2022). Rare and endemic plant species with low abundances are particularly vulnerable to the adverse impacts of invasive plants (Oostermeijer 2003; Jäger et al. 2009; Caujapé-Castells et al. 2010; Bellingham et al. 2018). For example, Jäger et al. (2009) found that endemic plant species in the Galápagos experienced a proportionally greater reduction in cover in response to woody plant invasion compared to non-endemic native species. An understanding of the impacts of invasive plants on native communities is therefore required to inform management and conservation, particularly in areas such as islands where plant communities are threatened (Ramula et al. 2008; Reid et al. 2009; Kettenring and Adams 2011).
The introduction of non-native plant species can reshape community structure, potentially impacting key metrics such as species composition, richness, and diversity. In some cases, non-invasive introduced species can coexist with native species without significantly changing the recipient community’s composition, richness, and diversity (Stohlgren and Rejmánek 2014; Lai et al. 2015; Diekmann et al. 2016). However, in other cases, dominant invasive plant species can substantially reduce native species richness and diversity (Mascaro et al. 2008; Jäger et al. 2009; Pyšek et al. 2012; Powell et al. 2013; Florens et al. 2017). While there is strong evidence that non-native plant species lead to ecosystem changes and have an overall detrimental effect on plant communities (Mascaro et al. 2008; Monty et al. 2013; Florens et al. 2017), specific impacts differ depending on the alien and native plant identities, cover/abundance, functional niches, life stages, time since invasion, and other environmental factors (Gooden et al. 2009; Kueffer et al. 2009; Vilà et al. 2011; Pyšek et al. 2012; Bradley et al. 2019).
Invasive species also differ in their scale-dependent impacts, which depend on a variety of environmental and anthropogenic factors as well as species-specific traits (Stohlgren and Rejmánek 2014; Valone and Weyers 2019; Hemp et al. 2021; Ren et al. 2022). The species area relationship (SAR) is used to understand the relationship between the size of an environment and the number of species found. Invader effects on SARs vary for different study systems and climates (Powell et al. 2013; Stohlgren and Rejmánek 2014; Valone and Weyers 2019; Lebbink et al. 2021). Most studies find that plant invasion reduces community richness at small scales (Powell et al. 2013; Valone and Weyers 2019; Lebbink et al. 2021). However, there are instances of higher small-scale richness or no significant difference post-invasion depending on the study system (Stohlgren and Rejmánek 2014). Results for species accumulation rates (SAR slopes) can also differ among study systems, as some studies find no difference in species accumulation with increased invasive plant cover (Stohlgren and Rejmánek 2014; Lebbink et al. 2021) and others find higher (Powell et al. 2013; Valone and Weyers 2019) or even lower (Stohlgren and Rejmánek 2014) species accumulation post-invasion depending on how the invasive species impacts richness at different scales. It is therefore important not to generalize invader effects on SARs across ecosystems.
It is informative to examine the composition of native species at different life stages and in different strata to determine whether they are differentially impacted by invasion and management and how this could affect the trajectory of forest regeneration (Mascaro et al. 2008; Baider and Florens 2011; Florens et al. 2017). Management typically increases recruitment and survival of native species, having a net positive impact on plant communities (Baider and Florens 2011). However, the impact of management can vary due to reinvasion or invasion by non-target introduced species (Reid et al. 2009) along with recruitment barriers including soil seed bank depletion, seed dispersal limitations, predation and competition, environmental conditions, and native population dynamics and demography (Erskine Ogden and Rejmánek 2005; Standish et al. 2007; Loh and Daehler 2008; Ramula et al. 2008; Nerfa et al. 2022).
Despite a wide range of potential regeneration outcomes, most studies focus on invasive species removal, while far fewer studies examine regeneration post-management (Kettenring and Adams 2011). A review of invasive plant control experiments showed that many experiments encompass only brief time frames (one growing season) and small plot sizes (< 1 m2), leading to results that are not always indicative of what happens long-term at the restoration-level scale, as well as to data that are more prone to errors of omission (Kettenring and Adams 2011). Additionally, grasslands are the predominant ecosystems studied (Schuster et al. 2018; Weidlich et al. 2020, 2021) and herbaceous invasive species are better studied than woody invasives (Schuster et al. 2018; Weidlich et al. 2020). Subtropical ecosystems are understudied (Carlucci et al. 2020; Weidlich et al. 2021), as are Pacific Islands (Kettenring and Adams 2011; Everingham et al. 2021), despite high endemism of flora on islands and substantial issues with invasive species (Caujapé-Castells et al. 2010; Kueffer et al. 2010; Pyšek et al. 2012; Ren et al. 2022) such as Psidium cattleyanum Sabine var. cattleyanum.
Psidium cattleyanum has been suggested as one of the world’s top 100 worst invasive species (GISD 2023). It particularly threatens flora on tropical/subtropical islands with histories of intense deforestation and many endemic flora (Macdonald et al. 1991; Auld and Hutton 2004; Shimizu 2006; Kueffer et al. 2009; Baider and Florens 2011; Takahashi et al. 2011; Director of National Parks 2018; Meyer et al. 2019). Examining how ecosystems recover after P. cattleyanum management can reveal if additional barriers to recovery exist that need to be addressed. In some cases, P. cattleyanum management alone can increase species richness and recovery of presumed extinct and critically endangered plant species (Baider and Florens 2011; Florens 2013; Florens et al. 2017). In other cases, seed predation and limited seed dispersal may hinder native forest regeneration following P. cattleyanum management (Mascaro et al. 2008; Nerfa et al. 2022), highlighting the need for additional active restoration actions.
Psidium cattleyanum is considered a primary threat to native vegetation on Norfolk Island (Director of National Parks 2010, 2018)—a small, remote island in the South Pacific—but there are no published studies about the impacts of P. cattleyanum and its management on native forest regeneration on this island. Norfolk Island has 43 endemic plant species, 15 of which are critically endangered, and many more of which are threatened (Environment Protection and Biodiversity Conservation—EPBC—Act 1999; Director of National Parks 2010, 2018). Its vegetation is not only threatened by P. cattleyanum but has also been impacted by several anthropogenic factors that are becoming increasingly common on tropical/subtropical islands globally such as deforestation, invasions by animals and other plant species, and climate change (Caujapé-Castells et al. 2010; Maurin et al. 2021; Hughes et al. 2022; Olsen and Cunningham 2022). Norfolk Island is a representative example of other P. cattleyanum-invaded islands with similar history and ecology (Macdonald et al. 1991; Shimizu 2006; Zimmerman et al. 2008; Tng et al. 2016; Florens et al. 2017; Meyer et al. 2019; GISD 2023).
The aim of this study was to document the consequences of P. cattleyanum removal on native subtropical forest regeneration across a range of spatial scales and spanning management times from 1993 to the present. Specifically, we were interested in addressing the following questions: (1) How does P. cattleyanum removal influence species-area relationships of woody plant species? (2) How does P. cattleyanum removal impact the abundance, richness, and composition of woody plant species in different vegetation strata (seedling, understory, and canopy)? and (3) What factors—including soil moisture, year since first management effort, and canopy cover—contribute to the magnitude of differences between managed and unmanaged plots?
Based on general trends in plant invasion studies, along with our field observations, we hypothesized that compared to unmanaged plots, managed plots would have higher richness at the smallest scale (the intercept of a species-area relationship) (Powell et al. 2013), higher overall richness at the largest scale (Baider and Florens 2011) and therefore no difference in accumulation rate (the slope of the species-area relationship) (Stohlgren and Rejmánek 2014; Lebbink et al. 2021), because if both small-scale and large-scale richness increase, the slope of the species-area relationship will not change. We also hypothesized that managed plots would have a higher abundance and richness of seedlings and understory plants (Baider and Florens 2011), as we expected dense P. cattleyanum to suppress the recruitment of some native species. We predicted strong compositional dissimilarity between managed and unmanaged plots, particularly in locations that were managed earlier where there has been more time for species to accumulate (O’Leary et al. 2018), and expected canopy cover (as a proxy for light) (Čuda et al. 2014; Depauw et al. 2021) and soil moisture (as a proxy for environmental favorability) (Takahashi et al. 2011) would be related to these compositional differences.
Methods
Study system
Psidium cattleyanum Sabine var. cattleyanum, commonly known as red guava or strawberry guava, is a woody plant species in the Myrtaceae family. Its native range extends from north-eastern Brazil to northern Uruguay (Tng et al. 2016; DeSisto et al. 2020). However, P. cattleyanum has invaded many other locations globally, introduced around the world largely for its edible fruits (Brown and Gurevitch 2004; Kueffer et al. 2010; Takahashi et al. 2011; Tng et al. 2016). It primarily invades tropical and subtropical ecosystems but can encroach into temperate regions (Kueffer et al. 2010; Patel 2012; Tng et al. 2016; Florens et al. 2016). Psidium cattleyanum has high reproductive capacity, including effective seed dispersal, as its attractive fruits are readily dispersed by birds and ungulates. It also reproduces vegetatively, forming dense stands and thick mats of roots that impede recruitment of other species (Christian 1999; Shimizu 2006; Tng et al. 2016). It alters soil chemistry and environmental conditions, producing allelopathic substances (Christian 1999; Patel 2012), modifying light levels (Christian 1999), and reducing groundwater recharge (Takahashi et al. 2011; Patel 2012). These traits enable P. cattleyanum to effectively depress recruitment of other plant species while simultaneously creating conditions more favorable to its own growth, making it a formidable invader (Christian 1999; Patel 2012). Although P. cattleyanum is especially successful at invading gaps after disturbance events (Christian 1999; Tng et al. 2016), it is also shade tolerant (Christian 1999; Tng et al. 2016), enabling it to colonize diverse ecosystems, and contributing to its effectiveness in obtaining dominance within a community (Fridley et al. 2023).
Norfolk Island (29° 02′ S 167° 57′ E) is an isolated subtropical volcanic island in the South Pacific (Fig. 1), with periodic Polynesian occupation from about 1200–1600 and English settlement since 1788 (Gibbs et al. 2017; Levin and Kark 2023). Prior to English settlement, Norfolk Island was covered primarily by dense subtropical rainforest, which was extensively cleared and grazed during the initial settlements to the extent that only about 460 hectares of remnant forest remains, and this forest is degraded, in part due to invasive species, particularly P. cattleyanum (Director of National Parks 2010, 2018). Norfolk Island is currently thought to contain approximately 182 native vascular plant species, 24% of which are endemic taxa, and a further 370 introduced species (over 60% of all plant species on the island) (Director of National Parks 2010; Mills 2010). About one-quarter of its native plant species are considered threatened (EPBC Act), and many of its endemic bird and invertebrate species rely heavily on native vegetation (Director of National Parks 2010).

Location of Norfolk Island in the South Pacific, the study region on the island (Norfolk Island National Park—outlined in yellow), and the setup of the managed and unmanaged paired plots in selected locations (red squares) within the National Park
Psidium cattleyanum was introduced to Norfolk Island in 1788 by the first European settlers for its edible fruits and spread rapidly (Christian 1999). Invasion was facilitated by deforestation and cattle grazing, which led to disturbance of native forest and dispersal of P. cattleyanum. The establishment of the National Park and Botanic Gardens in 1986 and the exclusion of cattle from the area (Director of National Parks 2018) along with ongoing weed management beginning in the 1990s served to decrease deforestation and cattle-facilitated invasion, and led to increased conservation efforts (Benson 1980; Director of National Parks 2010; Gibbs et al. 2017). P. cattleyanum, however, is still considered the most pervasive and problematic invasive plant species on the island and continues to pose a substantial threat to the native forest (Director of National Parks 2010). Norfolk Island, therefore, provides an ideal case study to examine P. cattleyanum invasion, management, and subsequent forest regeneration.
Plot design and survey methods
We collected field data between March and December 2020 from 15 locations in the Norfolk Island National Park (Fig. 1). At each location we identified two paired sites consisting of one unmanaged plant community and one managed plant community, where invasive species had been removed. We selected sites where P. cattleyanum was the dominant invasive plant species prior to management. Managed plots were selected to encompass a range of time intervals since the initiation of the first management efforts in 1993 up to the present. All invasive woody plants, regardless of the species, were removed from managed plots when first managed. The primary management method employed was ringbarking, which involved removing bark around the circumference of individual stems, disrupting water and nutrient transport. Herbicide application was used in conjunction with ringbarking to minimize re-shooting and ensure effective control. It is important to note that although the management efforts described were the general practice involved in P. cattleyanum removal, certain details may have varied over time and location. For example, most plots underwent a single, large-scale management intervention. However, a supplementary treatment may have taken place several years later. Additionally, the specific herbicides used, method of herbicide application, whether dead stems were retained or removed after intervention, and other management techniques may have varied. We selected pairs of plots with clear distinctions between areas where management did and did not occur, as the main goal of the study was to document how invasive species removal impacts the native plant community.
To select plot locations, we randomly assigned 15 points to a polygon map of managed areas in the National Park, then adjusted the locations as needed to encompass variation in environment and in time since management (from 1993 to the present). We further adjusted locations in the field to account for slight errors in the management shapefile and to better pair with unmanaged sites. We set up a total of 30 plots (15 pairs). Managed and unmanaged plot pairs were located as close together as possible to minimize potential environmental variation. We avoided managed sites that had been actively replanted. Plots were 125 m2 in overall size, with several smaller plots nested within for a total of two 25 m2 subplots, four 5 m2 subplots, and eight 1 m2 subplots to ensure sufficient replication at smaller scales (Fig. S1) (Peet et al. 1998; Dengler 2009; Powell et al. 2013).
We recorded all woody tree and shrub species in the 1 m2 subplots, and species absent from the smaller subplots were progressively recorded in the larger subplots. Both native and introduced species were included in the surveys and analyses, but the presence of introduced species (other than P. cattleyanum) was negligible. All vines, herbaceous species, and ferns except for tree ferns (Cyathea sp.) were excluded to concentrate solely on the woody species, as they represent a key structural and ecological aspect of forest ecosystems and are a common focus for forest restoration. This approach allowed us to explore the dynamics of woody plants in managed and unmanaged subtropical forest plots. To measure P. cattleyanum density in the managed and unmanaged plots, we laid a 11.2 m transect down the center of each plot and recorded the number of stems within 1 m either side of the transect (11.2 × 2 m total area). In addition, we recorded abundance counts and size class data for each woody tree and shrub species (except for P. cattleyanum, for which we calculated density) in the two 25 m2 subplots. We labelled any individual that formed the canopy or emerged above it “canopy” (anything equal or greater to the height of the P. cattleyanum stand, typically 7 + m tall), any individual smaller than 0.25 m a “seedling”, and any individual between those two heights an “understory” plant. For each plot we collected soil moisture data using a Fieldscout direct moisture probe (Spectrum Technologies, inc., USA), the number of years since first managed (if applicable) using a management shapefile, and canopy cover data using a spherical convex densiometer (Forestry Suppliers, inc., USA). Canopy cover and soil moisture data for all plots were collected in December 2020. Moisture measurements were collected during one week of dry weather (5 measurements per plot) and during one week of wet weather (5 measurements per plot) and measurements were then averaged per plot. Mean moisture and pH measurements, canopy cover, and P. cattleyanum stem count at the time of the study, as well as latitude/longitude, elevation, and year of first management activity can be found in Table S1.
Species-area analyses
All analyses were undertaken in R version 4.2.1 (R Core Team 2022). To test whether P. cattleyanum modified the scaling of species richness with area, we used the species-area equation S = cAz, where S is the number of species, A is area, and c and z are constants (intercept and slope, respectively). We transformed the values for richness and subplot size with a log10 transformation [log(S) = zlog(A) + log(c)] to calculate the slope (rate of species accumulation with area) and intercept (richness at small scales) of the species-area relationship (Figure S2). We tested for differences in the intercepts and slopes between managed and unmanaged plots using a linear mixed-effects model (lmer function from the lme4 package in R) (Bates et al. 2015), allowing each plot (nested in plot pair) to have its own intercept and slope as random effects. The main effect of treatment (i.e. management) was a test of differences in species richness between managed and unmanaged plots at small scales (the intercept). The interaction between treatment and subplot size was a test of the differences in slopes (the rate of species accumulation). This approach allowed us to account for uncertainty in the intercept and slope estimates while testing the effect of treatment. We also tested for the overall difference in richness between managed and unmanaged plots at the largest (125 m2) plot size (total plot richness) using a Mann–Whitney-U Test (Wilcoxon signed rank test with continuity correction in R).
Strata-based richness and abundance analyses
To conduct strata-based analyses we pooled individuals from the two 25 m2 subplots for a total survey area of 50 m2 per plot. We then subsetted the data by strata (canopy, understory, seedling). For each stratum, we tested the difference in richness (number of species) and the difference in abundance (count of individuals) between managed and unmanaged plots using Mann–Whitney-U Tests. Information about the tree ferns and woody tree and shrub species present in our study sites including species, stratum, status, EPBC listing, life form, max height, and lifespan (if available) can be found in Table S2. Additional information about strata-specific counts can be found in Table S3.
Drivers of dissimilarity
To calculate dissimilarity among our paired managed and unmanaged plots, we used the Bray–Curtis index. After ensuring our predictor variables were not correlated using the Pearson’s correlation coefficient, we used a multiple linear regression model to assess the contribution of soil moisture, time since first management effort, and canopy cover to the magnitude of pairwise dissimilarity between managed and unmanaged plot pair. We chose these predictors because of their biological relevance to plant management and regeneration. Soil moisture was used as a representation of environmental favorability for plant growth, years since first P. cattleyanum management as an indicator of how much time the managed site had to recover, and canopy cover as a proxy for light.
Additional compositional analyses
We further examined compositional differences between managed and unmanaged plots using presence-absence data of the species observed in the two combined 25 m2 subplots (the 50 m2 data) to assess patterns in the canopy and subcanopy, as well as using presence-absence data from the 125 m2 plots to assess overall patterns. We reduced dimensionality using non-metric Multi-Dimensional Scaling (nMDS) to extract two axes that best explained compositional variation among plots (using the metaMDS function in the vegan package) (Oksanen et al.), allowing us to visualize compositional differences between managed and unmanaged plots. We minimized the stress values to < 0.2, combining understory and seedling (referred to as “subcanopy”) data in the 50 m2 data for a more reliable fit. We calculated convex hulls to group the 50 m2 data by treatment (managed or unmanaged) and the 125 m2 plots by vegetation type. For vegetation type we used coastal forest (typically the driest), palm-gully forest (the wettest), and inland pine-hardwood forest (intermediate moisture availability) (Booth et al. 2021).
Results
Species-area relationships
We found no significant difference in richness at small scales (intercept of SAR, Fig. 2a, Table 1) and no significant difference in species accumulation with subplot size (slope of SAR, Fig. 2b, Table 1) between managed and unmanaged plots. Richness at the largest scale (125 m2) was significantly higher in managed plots compared to unmanaged plots (paired Mann–Whitney-U-Test, p = 0.003) (Fig. 2c), indicating that management increases richness at larger scales. The fact that richness did not differ significantly at small scales but was higher at the largest scale suggests that the slopes of SARs should have been steeper in managed plots than in unmanaged plots (because the SARs start at similar numbers of species at small scales but end at higher numbers of species for managed plots at the largest scale). We found some evidence for this with managed SAR slopes being steeper than unmanaged slopes for 11 out of 15 locations (Fig. 2b). When analyzed using a method that appropriately accounted for uncertainty in SAR estimates (a linear mixed-effects model), managed plots were found to have slightly steeper SAR slopes than unmanaged plots on average (small, positive estimate for the interaction term in Table 1) but the difference was only close to significant (p = 0.079).

Plots of a Richness at small scales (intercept estimates) for the species-area relationship (SAR) model, b species accumulation with subplot size (slope estimates) for the SAR model, c richness at the largest scale (125 m2 scale) for unmanaged and managed plots. Lines join pairs of managed and unmanaged plots. Statistical tests for a and b were conducted using a linear mixed-effects model, while a paired Mann–Whitney-U-Test was used for c. *p < 0.05, **p < 0.01, ***p < 0.001
Strata-based abundance and richness analyses
At the canopy level, managed and unmanaged plots did not significantly differ in richness (Fig. 3c, p = 0.495) or abundance (Fig. 3f, p = 0.393), indicating that the canopy stratum has not yet been impacted by P. cattleyanum. At both the understory and seedling levels, managed plots had significantly higher richness (Seedling: Fig. 3a, p = 0.033; Understory: Fig. 3b, p = 0.015) and significantly higher abundance (Seedling: Fig. 3d, p = 0.0111; Understory: Fig. 3e, p = 0.003).

Species richness in managed and unmanaged plots for seedling (a), understory (b), and canopy (c) strata; and abundance in managed and unmanaged plots for seedling (d), understory (e), and canopy (f) strata. For d–f abundance (+ 1) is plotted on the log10 scale only for visualization (small abundances were barely visible on the raw abundance scale). All statistical tests were paired Mann–Whitney-U-Tests: *p < 0.05, **p < 0.01, ***p < 0.001
Drivers of dissimilarity
Compositional dissimilarity in the subcanopy strata was greater when moisture levels were high (Fig. 4, p = 0.015; Table 2), suggesting post-management regeneration is enhanced when environmental conditions are more favorable for native plant recruitment or survival. Time since first managed and canopy cover were not significantly related to dissimilarity (Table 2).

The difference in Bray–Curtis dissimilarity indices between managed and unmanaged plot pairs in the subcanopy strata, plotted as a function of the mean soil moisture at each site (multiple linear regression, p = 0.015). Regression statistics are shown in Table 2
Additional compositional analyses
For the 50 m2 data, at the canopy level, there were strong compositional similarities between managed and unmanaged plots, indicated by high overlap between the two categories (Fig. 5a). At the subcanopy level, there was considerably less overlap between managed and unmanaged plots, indicating higher dissimilarity (Fig. 5b).

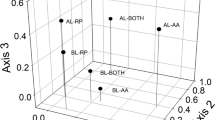
Non-metric multi-dimensional ordination scaling of a species present in the canopy for the 50 m2 data, with convex hulls around managed (black circles) and unmanaged (red triangles) plots; b species present in the subcanopy strata (seedling and understory species combined) for the 50 m2 data, also with convex hulls around managed and unmanaged plots; and c species present in the 125 m2 plots in managed (circles) and unmanaged (triangles) plots using species presence-absence data, with convex hulls around three different habitat types (coastal forest—orange hull, palm gully forest—green hull, and pine-hardwood forest—blue hull) and arrows indicating species that are correlated with a region in the ordination space. Dotted lines link managed and unmanaged plot pairs. Points closer together in ordination space represent plots with more similar species composition. All stress values were < 0.2
At the 125 m2 scale, the unmanaged plots grouped together, and the managed plots grouped together, with little overlap between the two categories, indicating compositional differences related to treatment type (Fig. 5c). Psidium cattleyanum was predictably associated with the unmanaged plots, whereas native species were more strongly associated with managed plots. The plots also grouped by vegetation type, with coastal forest (dry) and palm-gully (wet) having no overlap with each other, and pine-hardwood overlapping with the other two vegetation types. Rhopalostylis baueri (Norfolk Island/Niau Palm) and Cyathea tree ferns were strongly associated with gully vegetation types, Araucaria heterophylla (Norfolk Island Pine), Pittosporum bracteolatum and Alyxia gynopogon were strongly associated with pine-hardwood, and Myrsine ralstoniae, Elaeodendron curtipendulum, Lagunaria patersonia, and the invasive species Olea europaea were associated with coastal forests.
Discussion
Our study revealed that on Norfolk Island, subcanopy species richness and abundance increased after P. cattleyanum management, indicating the effectiveness of management efforts in promoting native woody species recruitment. Our findings highlight the importance of considering site-specific conditions such as moisture for strategic management, as our study showed that on Norfolk Island, successful native regeneration post-invasive species management is more likely in wetter environments conducive to native species recruitment. While this work focused on invasive plant management and subsequent forest regeneration, it is important to note that restoration encompasses a range of activities beyond invasive species control and passive regeneration, including active restoration and planting, which can further improve results.
Canopy species richness and abundance did not differ between managed and unmanaged plots, likely because remnant trees persisted in unmanaged plots, but the subcanopy showed significant differences. Psidium cattleyanum invasion reduces subcanopy abundance and richness in other areas (Zimmerman et al. 2008; Florens et al. 2017), and the results of our study suggest that management can lead to recovery if native species are capable of dispersing to the managed area. However, in situations where seed dispersal is limited and/or seed predation is high, native richness and abundance may not recover after management (Meyer et al. 2019; Nerfa et al. 2022), and many studies list dispersal limitations as a primary barrier to native forest regeneration (Holl 1999; Zimmerman et al. 2000; Baider et al. 2001; Standish et al. 2007; Loh and Daehler 2008; Palma et al. 2021; Nerfa et al. 2022). Therefore, management efforts should be prioritized in areas with remnant native species or where there are neighboring native species capable of dispersing into the invaded region after management for effective passive regeneration.
The implementation of useful management strategies additionally requires an understanding of which site features correspond with successful regeneration. Microhabitat—for example light and moisture—plays a crucial role in shaping the trajectory of forest restoration, including management and regeneration efficacy (Holmgren et al. 1997; Holl 1999; Zimmerman et al. 2000; Baider et al. 2001; Standish et al. 2007; Loh and Daehler 2008; Cabin et al. 2002; Čuda et al. 2014). Canopy cover and time since first management action did not significantly relate to dissimilarity, suggesting that they may not play a strong role in driving subcanopy recovery in this system. The highest compositional dissimilarity between managed and unmanaged sites occurred in areas with higher soil moisture, possibly because native plants are better at regenerating in higher moisture environments (Cabin et al. 2002). However, this is not always the case, as other studies have found a negative correlation between moisture availability and native plant regeneration (Funk et al. 2008; Schmidt et al. 2020), underscoring the importance of microsite and species trait assessments for effective management and regeneration strategies. These results suggest that P. cattleyanum management efforts should focus on areas with favorable conditions for native plant recruitment and growth to maximize natural regeneration success. Restoration outcomes may be improved in less suitable habitats through watering and actively planting seeds or seedlings suitable for the microsite (Nerfa et al. 2022).
Species-area relationships are useful for comparing invasive and native communities across spatial scales to determine whether there are fundamental differences between them. We found no significant difference in SARs between managed and unmanaged sites, despite many studies which find that invasion alters SARs (Powell et al. 2013; Valone and Weyers 2019; Lebbink et al. 2021). However, the relationship in this study may have been obscured or mitigated by the presence of remnant native canopy species, as differences were clearly seen in the subcanopy. SAR findings may not be consistent across different systems (Stohlgren and Rejmánek 2014) or across vertical strata within a system. Our results indicate that SARs are not always a useful metric for inferring management and regeneration success in highly structured environments such as forests unless strata-specific information is included.
Norfolk Island’s subtropical forests clearly benefit from P. cattleyanum management, showing passive regeneration in the subcanopy post-management. On Norfolk Island, and in other similar regions, resources should focus on invasive plant management where native species are still relatively abundant and the environment is favorable for native plant recruitment and growth. However, for rare native plant species and in regions where the seed bank may be depleted or there is little remaining native flora, out-planting of native seeds or saplings would be beneficial (Dann et al. 2021; Nerfa et al. 2022).
Conclusion
Useful management efforts must be informed by an empirical understanding of how plants respond to invasion. We found that species richness and abundance in the subcanopy increases as a result of P. cattleyanum management, with regeneration being most successful in high moisture environments. These results suggest that management allows for natural regeneration of native plant communities, even in highly invaded areas, if the growth conditions for native species are favorable and there is capacity for native recolonization through the soil seed bank or long-distance dispersal. Our study highlights the positive impact of invasive woody plant management on native plant regeneration and underscores the importance of strategically targeting management where the capacity for passive recovery is greatest.
Data availability
The datasets generated during and analysed during the current study are available from the corresponding author on reasonable request.
References
Auld TD, Hutton I (2004) Conservation issues for the vascular flora of Lord Howe Island. Cunninghamia 8:490–500
Baider C, Florens FBV (2011) Control of invasive alien weeds averts imminent plant extinction. Biol Invasions 13:2641–2646. https://doi.org/10.1007/s10530-011-9980-3
Baider C, Tabarelli M, Mantovani W (2001) The soil seed bank during Atlantic Forest regeneration in Southeast Brazil. Rev Bras Biol 61:35–44. https://doi.org/10.1590/S0034-71082001000100006
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting Linear Mixed-Effects Models Using lme4. J Stat Soft. https://doi.org/10.18637/jss.v067.i01
Bellingham PJ, Tanner EVJ, Martin PH et al (2018) Endemic trees in a tropical biodiversity hotspot imperilled by an invasive tree. Biol Cons 217:47–53. https://doi.org/10.1016/j.biocon.2017.10.028
Benson ML (1980) Dieback of Norfolk Island pine in its natural environment. Aust For 43:245–252. https://doi.org/10.1080/00049158.1980.10674278
Booth C, Nias R, Christian N, Mills K (eds) (2021) The native plant communities of Norfolk Island. Invasive Species Council and TierraMar. Katoomba, NSW
Bradley BA, Laginhas BB, Whitlock R et al (2019) Disentangling the abundance–impact relationship for invasive species. Proc Natl Acad Sci USA 116:9919–9924. https://doi.org/10.1073/pnas.1818081116
Brown KA, Gurevitch J (2004) Long-term impacts of logging on forest diversity in Madagascar. Proc Natl Acad Sci USA 101:6045–6049. https://doi.org/10.1073/pnas.0401456101
Cabin RJ, Weller SG, Lorence DH et al (2002) Effects of microsite, water, weeding, and direct seeding on the regeneration of native and alien species within a Hawaiian dry forest preserve. Biol Conserv 104:181–190. https://doi.org/10.1016/S0006-3207(01)00163-X
Carlucci MB, Brancalion PHS, Rodrigues RR et al (2020) Functional traits and ecosystem services in ecological restoration. Restor Ecol 28:1372–1383. https://doi.org/10.1111/rec.13279
Caujapé-Castells J, Tye A, Crawford DJ et al (2010) Conservation of oceanic island floras: present and future global challenges. Perspect Plant Ecol Evol Syst 12:107–129. https://doi.org/10.1016/j.ppees.2009.10.001
Christian N (1999) A study of the determinants of invasive success and management options for the weed species Psidium cattleianum sabine var. cattleianum (strawberry guava) in Norfolk Island.pdf. Honours, Southern Cross University
Čuda J, Skálová H, Janovský Z, Pyšek P (2014) Habitat requirements, short-term population dynamics and coexistence of native and invasive Impatiens species: a field study. Biol Invasions 16:177–190. https://doi.org/10.1007/s10530-013-0512-1
Dann L, Scott M, Guja L et al (2021) A guide to propagating Norfolk Island’s native plants and seeds. In: National environmental science program threatened species recovery hub. The University of Queensland, Brisbane
DeSisto CMM, Park DS, Davis CC et al (2020) An invasive species spread by threatened diurnal lemurs impacts rainforest structure in Madagascar. Biol Invasions 22:2845–2858. https://doi.org/10.1007/s10530-020-02293-7
Dengler J (2009) A flexible multi-scale approach for standardised recording of plant species richness patterns. Ecol Ind 9:1169–1178. https://doi.org/10.1016/j.ecolind.2009.02.002
Depauw L, Perring MP, Landuyt D et al (2021) Evaluating structural and compositional canopy characteristics to predict the light-demand signature of the forest understorey in mixed, semi-natural temperate forests. Appl Veg Sci 24:e12532. https://doi.org/10.1111/avsc.12532
Diekmann M, Effertz H, Baranowski M, Dupré C (2016) Weak effects on plant diversity of two invasive Impatiens species. Plant Ecol 217:1503–1514. https://doi.org/10.1007/s11258-016-0663-0
Director of National Parks (2010) Norfolk Island region threatened species recovery plan. Department of the Environment, Water, Heritage and the Arts, Canberra, ACT
Director of National Parks (2018) Norfolk Island National Park and Norfolk Island Botanic Garden Management Plan 2018–2028. Australian Government, Director of National Parks, Canberra, ACT
Erskine Ogden JA, Rejmánek M (2005) Recovery of native plant communities after the control of a dominant invasive plant species, Foeniculum vulgare: implications for management. Biol Conserv 125:427–439. https://doi.org/10.1016/j.biocon.2005.03.025
Everingham SE, Offord CA, Sabot MEB, Moles AT (2021) Time-traveling seeds reveal that plant regeneration and growth traits are responding to climate change. Ecology 102:e03272. https://doi.org/10.1002/ecy.3272
Florens FBV, Baider C, Martin GMN et al (2016) Invasive alien plants progress to dominate protected and best-preserved wet forests of an oceanic island. J Nat Conserv 34:93–100. https://doi.org/10.1016/j.jnc.2016.09.006
Florens FBV, Baider C, Seegoolam NB et al (2017) Long-term declines of native trees in an oceanic island’s tropical forests invaded by alien plants. Appl Veg Sci 20:94–105. https://doi.org/10.1111/avsc.12273
Florens FBV (2013) Conservation in Mauritius and Rodrigues: challenges and achievements from two ecologically devastated oceanic islands. In: Raven PH, Sodhi NS, Gibson L (eds) Conservation biology. Wiley, Oxford, pp 40–50
Fridley JD, Bellingham PJ, Closset-Kopp D et al (2023) A general hypothesis of forest invasions by woody plants based on whole-plant carbon economics. J Ecol 111:4–22. https://doi.org/10.1111/1365-2745.14001
Funk JL, Cleland EE, Suding KN, Zavaleta ES (2008) Restoration through reassembly: plant traits and invasion resistance. Trends Ecol Evol 23:695–703. https://doi.org/10.1016/j.tree.2008.07.013
Gibbs M, Duncan B, Varman R (2017) The free and unfree settlements of Norfolk Island: an overview of archaeological research. Aust Archaeol 83:82–99. https://doi.org/10.1080/03122417.2017.1404732
Global Invasive Species Database (GISD) (2023) Full species profile Psidium cattleianum. http://www.iucngisd.org/gisd/species.php?sc=59
Gooden B, French K, Turner PJ, Downey PO (2009) Impact threshold for an alien plant invader, Lantana camara L., on native plant communities. Biol Conserv 142:2631–2641. https://doi.org/10.1016/j.biocon.2009.06.012
Hejda M, Pyšek P, Jarošík V (2009) Impact of invasive plants on the species richness, diversity and composition of invaded communities. J Ecol 97:393–403. https://doi.org/10.1111/j.1365-2745.2009.01480.x
Hemp A, Del Fabbro C, Fischer M (2021) Area modulates the effect of elevation but not of land use or canopy on tropical plant species richness. Biodivers Conserv 30:4265–4277. https://doi.org/10.1007/s10531-021-02304-6
Holl KD (1999) Factors limiting tropical rain forest regeneration in abandoned pasture: seed rain, seed germination, microclimate, and soil. Biotropica 31:229–242. https://doi.org/10.1111/j.1744-7429.1999.tb00135.x
Holmgren M, Scheffer M, Huston MA (1997) The interplay of facilitation and competition in plant communities. Ecology 78:1966–1975. https://doi.org/10.1890/0012-9658(1997)078[1966:TIOFAC]2.0.CO;2
Hughes J, Petheram C, Taylor A et al (2022) Water balance of a small island experiencing climate change. Water 14:1771. https://doi.org/10.3390/w14111771
Jäger H, Kowarik I, Tye A (2009) Destruction without extinction: long-term impacts of an invasive tree species on Galápagos highland vegetation. J Ecol 97:1252–1263. https://doi.org/10.1111/j.1365-2745.2009.01578.x
Kettenring KM, Adams CR (2011) Lessons learned from invasive plant control experiments: a systematic review and meta-analysis: invasive plant control experiments. J Appl Ecol 48:970–979. https://doi.org/10.1111/j.1365-2664.2011.01979.x
Kueffer C, Daehler CC, Torres-Santana CW et al (2010) A global comparison of plant invasions on oceanic islands. Perspect Plant Ecol Evol Syst 12:145–161. https://doi.org/10.1016/j.ppees.2009.06.002
Kueffer C, Kronauer L, Edwards PJ (2009) Wider spectrum of fruit traits in invasive than native floras may increase the vulnerability of oceanic islands to plant invasions. Oikos 118:1327–1334. https://doi.org/10.1111/j.1600-0706.2009.17185.x
Lai HR, Mayfield MM, Gay-des-combes JM et al (2015) Distinct invasion strategies operating within a natural annual plant system. Ecol Lett 18:336–346. https://doi.org/10.1111/ele.12414
Lebbink G, Dwyer JM, Fensham RJ (2021) An invasive grass species has both local and broad-scale impacts on diversity: potential mechanisms and implications. J Veg Sci 32:e12972. https://doi.org/10.1111/jvs.12972
Levin N, Kark S (2023) From historical maps to remote sensing: reconstructing land use changes on Norfolk Island over the past 250 years. Cartogr J. https://doi.org/10.1080/00087041.2022.2150367
Loh RK, Daehler CC (2008) Influence of woody invader control methods and seed availability on native and invasive species establishment in a Hawaiian forest. Biol Invasions 10:805–819. https://doi.org/10.1007/s10530-008-9237-y
Macdonald IAW, Thébaud C, Strahm W, Strasberg D (1991) Effects of alien plant invasions on native vegetation remnants on La Reunion (Mascarene Islands, Indian Ocean). Environ Conserv 18:51–61. https://doi.org/10.1017/S0376892900021305
Mascaro J, Becklund KK, Hughes RF, Schnitzer SA (2008) Limited native plant regeneration in novel, exotic-dominated forests on Hawai’i. For Ecol Manage 256:593–606. https://doi.org/10.1016/j.foreco.2008.04.053
Maurin C, Lucas S, Ordens CM, et al (2021) Norfolk Island environmental assessment. In: A report by Monash University and The University of Newcastle for the Norfolk Island Regional Council (NIRC), and funded by the Australian Government
Meyer JY, Laitame T, Gaertner JC (2019) Short-term recovery of native vegetation and threatened species after restoration of a remnant forest in a small oceanic island of the South Pacific. Plant Ecol Divers 12:75–85. https://doi.org/10.1080/17550874.2019.1584651
Mills K (2010) Defining indigenous plants: some problematic species from Norfolk Island. Cunninghamia 11:407–412
Monty MLF, Florens FBV, Baider C (2013) Invasive alien plants elicit reduced production of flowers and fruits in various native forest species on the tropical island of Mauritius (Mascarenes, Indian Ocean). Tropic Conserv Sci 6:35–49. https://doi.org/10.1177/194008291300600107
Nerfa L, Hastings Z, Tsuneyoshi A et al (2022) Removal of non-native trees fosters but alone is insufficient for forest regeneration in Hawaiʻi. For Ecol Manage 517:120267. https://doi.org/10.1016/j.foreco.2022.120267
Olsen P, Cunningham R (2022) Influence of extreme and typical rainfall on nestling body condition of the endangered Norfolk Island Morepork: conservation implications of climate change. Emu-Austral Ornithol 122:176–185. https://doi.org/10.1080/01584197.2022.2105236
Oostermeijer JGB (2003) Threats to rare plant persistence. In: Brigham CA, Schwartz MW (eds) Population viability in plants. Springer, Berlin, pp 17–58
O’Leary B, Burd M, Venn SE, Gleadow R (2018) Integrating the passenger–driver hypothesis and plant community functional traits to the restoration of lands degraded by invasive trees. For Ecol Manage 408:112–120. https://doi.org/10.1016/j.foreco.2017.10.043
Palma AC, Goosem M, Fensham RJ et al (2021) Dispersal and recruitment limitations in secondary forests. J Veg Sci 32:e12975. https://doi.org/10.1111/jvs.12975
Patel S (2012) Exotic tropical plant Psidium cattleianum: a review on prospects and threats. Rev Environ Sci Biotechnol 11:243–248. https://doi.org/10.1007/s11157-012-9269-8
Peet RK, Wentworth TR, White PS (1998) A flexible, multipurpose method for recording vegetation composition and structure. Castanea 63:262–274
Powell KI, Chase JM, Knight TM (2013) Invasive plants have scale-dependent effects on diversity by altering species-area relationships. Science 339:316–318. https://doi.org/10.1126/science.1226817
Pyšek P, Jarošík V, Hulme PE et al (2012) A global assessment of invasive plant impacts on resident species, communities and ecosystems: the interaction of impact measures, invading species’ traits and environment. Glob Change Biol 18:1725–1737. https://doi.org/10.1111/j.1365-2486.2011.02636.x
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ramula S, Knight TM, Burns JH, Buckley YM (2008) General guidelines for invasive plant management based on comparative demography of invasive and native plant populations. J Appl Ecol 45:1124–1133. https://doi.org/10.1111/j.1365-2664.2008.01502.x
Reid AM, Morin L, Downey PO et al (2009) Does invasive plant management aid the restoration of natural ecosystems? Biol Conserv 142:2342–2349. https://doi.org/10.1016/j.biocon.2009.05.011
Ren H, Svenning J, Mi X et al (2022) Scale-dependent species–area relationship: Niche-based versus stochastic processes in a typical subtropical forest. J Ecol 110:1883–1895. https://doi.org/10.1111/1365-2745.13924
Schmidt AD, Castellani TT, De Sá DM (2020) Biotic and abiotic changes in subtropical seasonal deciduous forest associated with invasion by Hovenia dulcis Thunb. (Rhamnaceae). Biol Invasions 22:293–306. https://doi.org/10.1007/s10530-019-02089-4
Schuster MJ, Wragg PD, Reich PB (2018) Using revegetation to suppress invasive plants in grasslands and forests. J Appl Ecol 55:2362–2373. https://doi.org/10.1111/1365-2664.13195
Shimizu Y (2006) Population dynamics of an invasive plant (Psidium cattleianum) after two continuous disturbances in the Pinus-Schima secondary forest on Chichijima in the Ogasawara (Bonin) Islands. Reg Views 19:1–15
Standish RJ, Cramer VA, Wild SL, Hobbs RJ (2007) Seed dispersal and recruitment limitation are barriers to native recolonization of old-fields in western Australia: seed and recruitment limitations on old-fields. J Appl Ecol 44:435–445. https://doi.org/10.1111/j.1365-2664.2006.01262.x
Stohlgren TJ, Rejmánek M (2014) No universal scale-dependent impacts of invasive species on native plant species richness. Biol Lett 10:20130939. https://doi.org/10.1098/rsbl.2013.0939
Takahashi M, Giambelluca TW, Mudd RG et al (2011) Rainfall partitioning and cloud water interception in native forest and invaded forest in Hawai‘i Volcanoes National Park. Hydrol Process 25:448–464. https://doi.org/10.1002/hyp.7797
Tng DYP, Goosem MW, Paz CP et al (2016) Characteristics of the Psidium cattleianum invasion of secondary rainforests. Austral Ecol 41:344–354. https://doi.org/10.1111/aec.12319
Valone TJ, Weyers DP (2019) Invasion intensity influences scale-dependent effects of an exotic species on native plant diversity. Sci Rep 9:18769. https://doi.org/10.1038/s41598-019-55165-z
Vilà M, Espinar JL, Hejda M et al (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol Lett 14:702–708. https://doi.org/10.1111/j.1461-0248.2011.01628.x
Weidlich EWA, Flórido FG, Sorrini TB, Brancalion PHS (2020) Controlling invasive plant species in ecological restoration: a global review. J Appl Ecol 57:1806–1817. https://doi.org/10.1111/1365-2664.13656
Weidlich EWA, Nelson CR, Maron JL et al (2021) Priority effects and ecological restoration. Restor Ecol 29:e13317. https://doi.org/10.1111/rec.13317
Zimmerman N, Flint Hughes R, Cordell S et al (2008) Patterns of primary succession of native and introduced plants in lowland wet forests in eastern Hawai‘i. Biotropica 40:277–284. https://doi.org/10.1111/j.1744-7429.2007.00371.x
Zimmerman JK, Pascarella JB, Aide TM (2000) Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restor Ecol 8:350–360. https://doi.org/10.1046/j.1526-100x.2000.80050.x
Acknowledgements
We thank the Norfolk Island community and the Norfolk Island National Park staff for providing access and support for our research (permit NI 2018/27). We thank Melinda Wilson, Joel Christian, and Nigel Greenup for their assistance with planning and executing the fieldwork, reading drafts, and providing advice. We thank Andrew Rogers for the help with survey design and data collection. We acknowledge Parks Australia, the Australian National Botanic Gardens and Friends of the Australian National Botanic Gardens, and the National Environmental Science Program’s Threatened Species Recovery Hub for funding and support. We thank the editor and reviewers for the time and effort they took to review this manuscript and provide constructive suggestions.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This work was funded by the National Environmental Science Program Threatened Species Recovery Hub and Friends of the Australian National Botanic Gardens/Parks Australia to Salit Kark. Open access funding was provided by the University of Queensland.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Data collection was performed by Leah Dann. Data analysis was performed by Leah Dann, John Dwyer, and Lydia Guja. The first draft of the manuscript was written by Leah Dann. All authors edited and made comments on manuscript drafts. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Dann, L.E., Guja, L., Kark, S. et al. Comparative study reveals management of a dominant invasive plant facilitates subtropical forest regeneration. Biol Invasions 26, 299–313 (2024). https://doi.org/10.1007/s10530-023-03174-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-023-03174-5