
Abstract
Agroforestry systems provide important ecosystem functions and services. They have the potential to enrich agricultural monocultures in central Europe with structural elements otherwise absent, which is expected to be accompanied by a surplus of ecosystem functions. Here we used quantitative measures derived from terrestrial laser scanning in single-scan mode to describe the structural complexity, the canopy openness, the foliage height diversity and the understory complexity of four common agroforest systems in central Europe. We accessed silvopasture systems with grazing ponies and cattle as well as fellow deer, short rotation forests with agricultural use between the tree rows, tree orchards with grazing sheep and Christmas tree plantations on which chickens forage. As a reference, we used data for 65 forest sites across Germany, representing different forest types, various dominant tree species, stand ages and management systems. We found that overall stand structural complexity is ranked as follows: forest > silvopasture systems > short rotation forest > tree orchard > Christmas tree plantation. Consequently, if overall structural complexity of an agricultural landscape shall be enriched, there is now strong evidence on how this may be achieved using agroforests. However, if the focus lies on selected structures that serve specific functions, e.g. dense understory to provide animal shelter, specific types of agroforests may be chosen and the ranking in overall structural complexity may be less important.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In many countries of the world the landscape has been converted from grasslands or forests into cropland or pastures (e.g. Goldewijk et al. 2011). In the European Union, the share of agricultural land is decreasing but management intensity on the remaining area is increasing (Reidsma et al. 2006). In agricultural landscapes there can be little room for animals, particularly when fields are large, production is heavily mechanized, and insecticides are used. Faunal diversity is therefore often reduced in agricultural landscapes (Heikkinen et al. 2004; Bengtsson et al. 2005; Habel et al. 2019). Not only losses in insect diversity are related to intensive agriculture (Seibold et al. 2019; Cardoso et al. 2020) but also mammals do not find suitable habitats in agricultural environments (e.g. Laliberte and Ripple 2004). They rely on specific structures that often involve woody vegetation for breeding, feeding and hiding (Putman 1997).
It is therefore not surprising that even small refugees like hedgerows and tree groves, or even uncropped areas under power line pylons are important retreats for mammals on a landscape scale (e.g. Michel et al. 2006; Bates and Harris 2009; Šálek et al. 2020). So-called ‘trees outside forests’ (TOF) include hedgerows, tree groves, forest patches and other perennial wooden vegetation outside forests (FAO 2015). The existence of the tree component in such landscape elements is crucial, since vegetation vertically extending beyond the height of shrubs and bushes is known to have beneficial effects on many ecosystem services and functions (e.g. Ringler et al. 1997; Manning et al. 2006, 2009; Harvey et al. 2006; Bates and Harris 2009).
Agroforests, or agroforestry systems, are also considered TOF (FAO 2015). They may be useful alternatives to agricultural monocultures since they can combine the benefits of agricultural crops with those of trees and are generally known to provide many beneficial functions and services to humans and animals (e.g. Sheppard et al. 2020). This may include an increased structural complexity and hence greater habitat diversity for animals in landscapes that are otherwise dominated by agricultural systems (Manning et al. 2009). A single definition for agroforestry is still to be agreed on since the existing types of systems are as diverse as the ecological zones they are practiced in (Böhm et al. 2017). Generally, the term describes land use systems in which woody vegetation (trees or bushes) is combined with arable crops and/or animal husbandry so that ecological and economic benefits are created between the participating components (Nair 1993). It is the intentional combination of agriculture/ or livestock and forestry.
In Europe, traditional agroforestry systems mostly comprise of trees in combination with livestock or cereals (Reisner et al. 2007) while modern agroforests in Europe comprise of agricultural crops cultivated under or in-between fast growing tree species that are eventually harvested in short rotation cycles (Nerlich et al. 2013).
When considering tropical agroforestry systems, there are agroforests that comprise of perennial arable crops, like cacao, coffee, vanilla, rubber or others that are cultivated in the shade of forest trees (e.g. Tscharntke et al. 2011) while others are mixtures of oil palms and forest trees (e.g. Zemp et al. 2019).
The combining element of all types of agroforests is the existence of the woody vegetation component (e.g. Somarriba 1992). The presence of trees induces a structural component that makes agroforests different from agricultural cropland with annual rotations. This creates new and persistent types of habitats unavailable in agricultural fields but important to many species (Manning et al. 2009).
So far, it has rarely been quantified how different agroforestry systems compare with regard to their structural characteristics. We are aware of only one study that compared structural characteristics of tropical forest with that of oil palm plantations in which trees were planted to enrich the structural heterogeneity (Zemp et al. 2019). To our knowledge, past research did not aim for a quantitative comparison of the ‘structural complexity’ of different agroforestry systems since there was no methodology to do so. However, such means have been developed for forests lately based on terrestrial laser scanning in single-scan mode (e.g. Seidel et al. 2016, 2019; Ehbrecht et al. 2017; Willim et al. 2019). There is no reason to assume that such measures cannot be applied to agroforestry systems as well, as long as the vegetation has a minimum height of at least 80 cm, which is the lower limit in the assessment based on the aforementioned indices. If vegetation would be smaller, it would not be captured by any of the laser-based indices.
Here, we used four measures derived from highly efficient single-scan terrestrial laser scanning to assess structural characteristics of different agroforests in Germany and compared them with forest references.
We hypothesized that (1) agroforestry systems have a lower structural complexity, (2) a more open canopy, (3) less diverse vertical structures and (4) a lower complexity in the understory when compared to a forest reference. Our main focus was to provide a ranking of different agroforestry systems with regard to their structural characteristics. Such information may aid landscape management. For example, it would allow using agroforestry systems as a means to create specific structures that provide distinct ecosystem functions and services that are otherwise absent in agriculturally dominated landscapes.
Material and methods
We decided to investigate those agroforestry systems that are most often found in Germany. In the vicinity of 40 km around Göttingen, we were able to identify at least two replicates of all respective types, namely short rotation forestry (abbrev. SRF; n = 2), Christmas tree plantations (abbrev. CTP; n = 2), silvopasture systems (n = 2) and tree orchards (n = 3), for which the owners gave us permissions to conduct measurements in winter 2019/2020.
As a reference for forest structure, we used winter scans from forests across Germany acquired in winter 2014/15 in the framework of the Biodiversity Exploratories (see https://www.biodiversity‐exploratories.de/ or Fischer et al. 2010 for further detail). Accordingly, our study captures all sites in leaf-less status if deciduous trees were present. We used the public datasets 18268, 18269, 18270, 20055 and 17486, all available at the biodiversity explanatory database (https://www.bexis.uni-jena.de/sws/PublicDataLink/Index), for information on the stand characteristics as listed in Table 1.
All forest sites were located in three geographical areas, namely the Hainich-Dün region, the Schorfheide-Chorin or the Swabian Alb. Therefore, these sites cover three major climatic regions of Germany and various typical forest types, including stands dominated by Fagus sylvatica L., Picea abies H. Karst (L.), Pinus sylvestris L. and Quercus robur L. The forest stands ranged from managed age-class forest, over shelterwood forests to unmanaged forest (National Park or Biosphere reserve), and from young stands (immature timber) to mature stands. Table 1 provides an overview on the study sites, the number of scans taken per site, the geographical location and some key characteristics of the sites. A map of the locations is provided in Fig. 1.

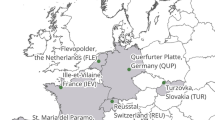
Map of the study site locations with the forest reference sites located in the Schorfheide Chorin (NE-Germany), the Hainich-Dün (Central) and the Swabian Alb (SW-Germany) and the agroforestry sites located within or in close-proximity to the municipality of Göttingen, Lower Saxony, Germany. Exact geolocations of the agroforest sites are provided in Table 1. Study site numbers in the inlay refer to: 1- CTP “Thüringen 1”; 2- CTP “Thüringen 2”; 3- Silvopasture “Hardegsen”; 4- Silvopasture “Solling”; 5- SRF “Göttingen”; 6- SRF „Reiffenhausen “; 7- Orchard „Am Lohberge “; 8- Orchard „Elkershausen “; 9- Orchard “Totenhäuser Gräben “ as visualized in Fig. 2
Terrestrial laser scanning
On each agroforest site, we conducted a minimum of eight scans with a Faro Focus M70 laser scanner (Faro Technologies Inc., Lake Marry, Florida, USA) mounted on a tripod at 1.3 m above ground in December 2019 (winter). Scans were placed on a quadratic grid with 33 m width that was randomly (orientation and start) placed over the sites. At each grid point (intersection of grid lines), the scanner was set to scan a field of view of 300° vertically and 360° horizontally with an angular step width of 0.035 degrees. This setup resulted in over 40 million measurements per scans, with each measurement corresponding to a laser beam that is emitted into the scanners vicinity and received back by the instrument if reflected by an object within 70 m distance or less. Each scan was converted into an xyz-file using the software Faro Scene (Faro Technologies, Lake Marry, Florida, USA). In Faro Scene, all scans were filtered for erroneous points using the software’s standard settings. Finally, before export from Faro Scene, each scan was reduced in resolution to 1/16th of the original scan for faster processing and as required for the calculation of the various indices described below. Co-registration was not required as a results of the single-scan approach.
In order to capture the variation of structures of various forests types we sampled 65 different forest sites of the Biodiversity Exploratories, each using one single laser scan position, except for four sites were we conducted two scans due to a large within plot heterogeneity (see details in Table 1, total of 69 scans in forest reference). Figure 2 shows an exemplary scan taken on each agroforestry site and one example from a forest reference site.

2D representation of an exemplary 3D point cloud from a single-scan from each study site and an exemplary forest site (here: Hainich forest in Thuringia, Germany)
Point cloud processing
Each xyz-file of a scan can be visualized as a so called point cloud for verification of the scan procedure (see Fig. 3). Here we used the xyz-files as input for a fully automated assessment of the structural complexity of the scene captured by the scanner. To do so, we used the so called Stand Structural Complexity Index (abbr.: SSCI), introduced in 2017 (Ehbrecht et al. 2017; R-code available here: https://github.com/ehbrechtetal/Stand-structural-complexity-index–SSCI). This index uses the distribution of hits to assess stand structural complexity solely from a mathematical point of view and hence without the operators involvement or expert knowledge.

2D representation of the 3D point cloud of an exemplary forest reference scene obtained from a single scan in leaf-less condition
Originally designed for forests, the index is calculated using Mathematica (Wolfram Research, Campaign, USA) based on 1280 vertical cross-sectional polygons through the stand, with each cross section being analyzed in view of its shape complexity. While details are provided in Ehbrecht et al. (2017), we here shortly summarize the procedure as follows: if the structure of the surrounding forest is complex, laser hits of consecutive beams will occur in a large variety of distances to the scanner. By combining all hits with a line a polygon is created and evaluated for its circumference-to-area ratio as introduced by McGarigal and Marks (1995) to assess complexity of polygons in landscape analysis. The average complexity of all 1280 polygons is used as a measure of complexity of the forest scene. However, since in forests complexity has little meaning without scale, the stand height is used to scale the average complexity with the vertical extent of the stand investigated. This ensures that tiny but complex thickets do not result in higher SSCI values than a multi-layered old-growth forest. The index proved to be valuable in various earlier studies. For example, it was successfully used to quantify impacts of forest management on forest structure (Ehbrecht et al. 2017; Stiers et al. 2018), to quantify the effect of mixture on forest structure (Juchheim et al. 2020), to assess tree microhabitats (Frey et al. 2020), it was closely related to the forest microclimate (Ehbrecht et al. 2019), and it was even a good predictor of ant species richness (Greve et al. 2018) or conventional expert ratings on stand structural complexity from visual assessment (Frey et al. 2019). In a recent study focusing on primary forests across the globe, the index was used to relate the stand structural complexity of forests to the climatic conditions of different biomes (Ehbrecht et al. 2021) and it was shown to correlate to conventional measures of structural heterogeneity (Ehbrecht et al. 2017).
The second measure obtained from the laser scans was canopy openness. It was calculated according to Zheng et al. (2013) and can be interpreted as the percent vegetation coverage in an upward facing canopy image simulated for the scanners position. The viewing angle is hereby restricted to a 60° opening angle (upside-down cone). Canopy openness is strongly related the microclimatic conditions in a stand (Ehbrecht et al. 2019) and it is related to the leaf area and light regime of a stand (e.g. Woodgate et al. 2015).
As a third measure, we calculated the Foliage Height Diversity (FHD) for each scan as introduced in Seidel et al. (2016). FHD is a measure of vertical distribution of plant material and was first introduced in 1961, when it was still determined based on visual assessments (MacArthur and MacArthur 1961). When based on laser scanning data, FHD is derived from the density of laser hits in horizontal layers of 1 m thickness relative to the density of the full stand profile. A low FHD indicates that there are great differences in the filling of different horizontal layers, while a high FHD indicates that all layers are more or less similarly filled with plant material. FHD is a particularly useful predictor of bird species diversity (MacArthur and MacArthur 1961).
Finally, we calculated the Understory Complexity Index (UCI) as introduced in Willim et al. (2019). The UCI is a measure of the three-dimensional architectural and spatial distribution of plant material in the shrub layer of a forest. It is calculated based on a horizontal layer ranging from 0.8 to 1.8 m through the stand. For this layer, all laser hits are vertically projected to a plane and the outline of the scanned area, defined by the innermost laser hits in any azimuthal direction, are connected by a line. This line forms a polygon if connections are made clockwise and the area and perimeter of this polygon are used to determine the circumference-to-area ratio as introduced by McGarigal and Marks (1995) and mentioned earlier for the analog calculation of SSCI.
Statistical analysis
The free software R (Vers.3.5, R Development Core Team) was used for all statistical analyses.
We conducted a Shapiro–Wilk’s Test for normality of the data and Levene’s test for variance homogeneity. Since neither normality of data nor variance homogeneity could be expected, we performed the non-parametric Kruskal–Wallis test to test for significant differences among the groups. For post-hoc testing we used the Wilcoxon-Rank-Sum test for pairwise comparisons based on a confidence interval of 95%. For all tests, we pooled the data of the respective systems, e.g. all scans taken in the agroforestry systems, to derive means, medians and standard deviations. Unpooled data per site can be found in the SI.
Results
The results of the Kruskal–Wallis test and the Wilcoxon-Rank-Sum test are summarized in Table 2 an will be discussed below.
Stand structural complexity
The stand structural complexity of the investigated systems decreased from the forest (mean: 3.77) to silvopasture (mean: 3.2) to SRF (mean: 3.02) to the tree orchard (mean: 2.46) and the CTP with lowest mean (1.95). While on average the forest systems where significantly more complex than the SRF, the tree orchard and the Christmas tree plantation, there were no significant differences between the forests and the silvopastures. Despite the ranking, silvopastures where also not significantly more complex than SRF or tree orchards. Figure 4 depicts the SSCI data of the different systems.

Box-and-whisker plots of the stand structural complexity index (SSCI) of the forest reference in comparison to the agroforestry systems. SRF: Short rotation forest, CTP: Christmas tree plantation. Different lower-case letters above the boxes indicate significant differences among the systems. White horizontal lines indicate the median, while the upper and lower end of the boxes indicate the 75% and 25% quantile respectively
The standard deviations were large for both forests (± 1.41) and silvopastures (± 1.49) and comparably small for SRF (± 0.57), tree orchards (± 0.59) and the CTP (± 0.70).
Canopy openness
For canopy openness we observed a ranking of means from forest (28%) to silvopasture (59%) to tree orchard (81%) to SRF (86%) to CTP (99%). While forests were significantly more closed than all other systems, CTP had a significantly higher openness than all other systems. Silvopastures, SRFs and tree orchards had similar median values, but strongly differing ranges of values. The mean values however differed more, resulting in significant differences between silvopasture and tree orchard, but not between any of the two and SRFs. Figure 5 illustrates the described pattern.

Box-and-whisker plots of canopy openness (in percent) of the forest reference in comparison to the agroforestry systems. Different lower-case letters below the boxes indicate significant differences among the systems. White horizontal lines indicate the median, while the upper and lower end of the boxes indicate the 75% and 25% quantile respectively
The standard deviations differed strongly between the groups, with the CTP being most homogeneous (standard deviation: ± 0.02), followed by the SRFs (± 0.18), the forest reference (± 0.15) and the tree orchards (± 0.27), and finally the silvopastures with the largest variability (± 0.41) of openness values.
Foliage height diversity
The foliage height diversity of the two systems containing forest trees, namely the forest reference and the silvopastures, had significantly higher means (3.04 and 2.74 respectively) than the other three systems, namely the SRFs (1.61), the tree orchards (1.68), and the CTP (1.78). Within these two groups there were no significant differences. Figure 6 provides a graphical overview of the data.

Box-and-whisker plots of foliage height diversity of the forest reference in comparison to the agroforestry systems. Different lower-case letters above the boxes indicate significant differences among the systems. White horizontal lines indicate the median, while the upper and lower end of the boxes indicate the 75% and 25% quantile respectively
The standard deviations of the silvopastures (± 0.75) and CTP (± 0.70) were larger compared to those of the tree orchards (± 0.29), the SRFs (± 0.24) or the forest reference (± 0.35).
Understory complexity
The understory of the forest reference (mean: 2.44) and the SRFs (mean: 3.96) were not significantly different from one another (see Fig. 7) but significantly more complex than those of the other systems (silvopasture: 1.69; tree orchard: 1.48; CTP: 1.68).

Box-and-whisker plots of the understory complexity index of the forest reference in comparison to the agroforestry systems. Different lower-case letters above the boxes indicate significant differences among the systems. White horizontal lines indicate the median, while the upper and lower end of the boxes indicate the 75% and 25% quantile respectively
The variability of the understory structures, assessed via the standard deviation of the understory complexity index of all scans taken in a system, differed largely between the systems. While the forest reference and the SRFs showed large variations (std. dev.: ± 1.31 and ± 3.00), the silvopastures (± 0.43), the tree orchards (± 0.49), and the CTP (± 0.26) were much more homogenous across scans.
Discussion
All measures used in our study to describe vegetation structures were based on single scans and are therefore prone to occlusion effects (e.g. Anderson et al. 2017). It is documented that a reliable structural characterization of different forest types can be performed using the SSCI obtained from single scans when about nine scans are taken per hectare (Ehbrecht et al. 2017). Since our aim was to characterize the different management systems, we followed the nine-scans-per-hectare protocol, resulting in at least 18 scans per system that were used to characterize structures.
According to the SSCI, reaching as far as 70 m in case of absence of plant material in the direct vicinity of the scanner, the observed ranking of systems with regard to their structural complexity is surprising. While forests clearly provide the greatest complexity in terms of the overall highest mean and median SSCI value detected, they also showed the greatest variability. This can be explained by the large number of different forest management systems included in our study in order to provide a solid reference across stand ages, dominating tree species, and management regimes. A large variation in forest structures due to different stand managements and forest types have already been documented for our sites in earlier studies using laser scanning-based measures (cf. Ehbrecht et al. 2017; Seidel et al. 2020). The silvopastures were statistically not significantly different from the forest reference and only had a slightly lower mean, median and lower maximum complexity. We argue, this is due to the fact that both silvopastures contained patches that appeared to be “forest like”, along with larger openings shaped by intensive animal grazing. Also, the two silvopastures differed strongly in their characteristics. While the Solling site was dominated by an open broad-leaved forest with grazing patches being spatially intermingled, the Hardegsen site only had trees in one part of the area and was dominated by a larger grazing patch in the other part. This also explains the larger standard deviation found in the silvopastures when contrasted to the forest reference.
A look at the canopy openness data supports the impression that the silvopastures contained some very open areas while also closed “forest-like” patches were present. Here, it is evident that it is worth to consider different aspects of complexity when comparing the different systems, since some characteristics may not differ strongly while others depict important dissimilarities (McElhinny et al. 2005). Additionally, the general availability of resources other than complexity, e.g. the existence of light or shade, which may be crucial in agroforestry systems (e.g. Bohn Reckziegel et al. 2021), may not be assessed by only one measure.
Surprisingly, despite the absence of larger trees, the stand structural complexity of the SRFs was higher than that of tree orchards or CTP. We argue this was due to the rather sparse setting of the latter two systems when compared to the densely planted rows in the alley-like short rotation system. Again, this is supported by the results of the canopy openness, showing very high mean values for both, CTP and tree orchards. Outliers towards the lower end of canopy openness (closed canopies) in the SRFs were simply caused by scans that were taken inside the vegetation rows. When compared to forests, SRFs may be of less divers structure, thereby possibly having a lower species diversity (e.g. Gallé et al. 2017), but when compared to agricultural cropland they may hold a large potential to provide structures that help increasing the biodiversity of agricultural landscapes (e.g. Verheyen et al. 2014). Also coppice forests attract a species portfolio quite different from that of high forest systems.
We observed little variability in the openness of the CTP due to the shape of Christmas trees, preventing a significant proportion of the trees from reaching into the range of the cone used to calculate the canopy openness. While Christmas trees are cultivated to look all the same and to have this cone-like shape by avoiding crown closure, for the tree orchard, the variability in canopy openness was much greater. Here the focus is not on symmetrical trees. Also, trees were of different ages among different systems, with the tree orchard “Elkershausen” being only ten to twelve years old by the time of the measurements, while the trees on both other orchards were around 70 years old. Therefore, the “Elkershausen” site was more open and tree crowns were far from being in contact with each other. A large variability of openness values points towards a high heterogeneity of light conditions within the systems. From these, more light-demanding species benefit. These are often pollinated by insects and the presence of these plant species is often accompanied by an abundance increase of the insects (e.g. Nerlich et al. 2013).
When it comes to the vertical diversity of structures (FHD), the two systems containing tall forest trees, namely the forest reference and the silvopastures, clearly provide the greatest diversity of structures, or, in other words, the greatest habitat heterogeneity. SRFs, tree orchards or CTP are characterized by a lower amount of vertical structures. Regularly planted trees of homogenous age resulted in very similar and simple vertical structures for all individuals. For FHD, as already seen for canopy openness, the variation between scans was largest in the silvopastures. Again, this can be explained by the large structural differences between the grazing patches and the more forest-like patches in the silvopastures. Very low values of FHD in the silvopastures may be explained by a homogenous filling on a very low level. This is because FHD does unfortunately not differentiate between sites with high or low filling if the filling pattern are the same between the layers (equally high or equally low).
The complexity of the understory in a height of 0.8 to 1.8 m is strongly affected by the number of tree stems in the vicinity of the scans (cf. Willim et al. 2019) and also by any kind of agricultural plants, herbs, or shrubs growing at a minimum height of 80 cm. Trees have a large effect particularly when they are small, due to their branches and leaves still being located in the understory, thereby increasing understory complexity. Large forest trees, with long branch-free boles, may have a lower effect, particularly since they are often more widely spaced. Therefore it is little surprising, that the SRFs yielded the highest UCI values, with their densely planted tree rows. In cases when a scan was taken between two rows, the distance to the next row would determine the complexity at this sample point. This resulted in low complexity for scans taken between rows and high complexity when scans were inside the planted alleys, explaining the large variability of UCI in the SRFs. Only in two cases, when scans were taken in-between two rows of the SRFs, the UCI failed and could not be determined. The polygon used to determine the complexity could not be created as not enough vegetation reached into that part of the scan that is actually considered for determining UCI (maximum distance: 20 m; cf. Willim et al. 2019). We argue this should be interpreted as absence of sufficient structures (no hits) and hence absence of structural complexity. In contrast, for the forest scans, the UCI variability is likely caused by the absence or presence of juvenile trees or regeneration patches. Grazing (cattle, ponies and deer in silvopastures, sheep in tree orchard) or removal of competing vegetation between the trees through mowing (CTP) most likely resulted in very homogeneous and overall low UCI-values.
Here, we used forest structures as a reference, as the used indices were originally designed to address those structures. These indices are, however, technically applicable to agroforests as well. With forest serving as one reference, it would be interesting to determine comparable indices used here for agricultural crops as well in order to have the other side of the structural reference. However, this is not possible unless the agricultural fruits are taller than 0.8 m for the UCI, 1.3 m for SSCI and canopy openness and at least several meters in height for the calculation of FHD. A direct consequence of this “forest-based” perspective is that comparisons with forest structures may result in interpretations that emphasize the structural deficits of agroforests. If, however, one pictures the benefits they provide when compared to agricultural monocultures, the outcome may be very different. Also, it is important to keep in mind that the main intention was to compare the different approaches within the group of agroforestry systems to one another. Technically, all measures used in our study would yield zero or no values (error) for typical agricultural crops like wheat, rye, barley or oilseed rape cultivated on large scale in Germany. A widely cultivated crop that may be an exception is corn, since this plant could reach two meters or more in height prior to harvest. Changing the perspective from forest as a reference to cropland as a reference would hence mean an increase in structural complexity.
The consequences of such a change in the reference system and correspondingly the change in perspective, are far-reaching. For example, we interpreted the FHD, a measure particularly important for bird species (cf. MacArthur and MacArthur 1961), of CTP, SRFs and tree orchards as very low compared to forests. When compared to agricultural land, however, positive effects on the bird species assemblages were found for CTP (e.g. Gailly et al. 2017), SRFs (e.g. Christian et al. 1997) as well as tree orchards (e.g. Kajtoch 2017), most likely because they simply add structures to the system. Since measures of cropland structural complexity are unavailable so far, we could only use indices designed for forest. However, we hypothesize that the inverse order of our findings may indicate the ranking of structural benefits to be expected if agricultural monocultures are ‘enriched’ with components of the investigated agroforestry systems. Future research in agroforests may address changes of structure over time and changes due to agricultural interventions, as well as an assessment of the different effects caused by the variety of agricultural/silvicultural interventions possible.
Conclusions
We acquired quantitative 3D data on vegetation structure from highly efficient single-scan terrestrial laser scanning. We could show that selected attributes of complexity may be more pronounced in agroforestry systems than in forests, such as the understory complexity in SRFs, and thereby promote specific ecosystem functions or services. For other measures, such as foliage height diversity, only the silvopastures provide the relevant structures to resemble forests. Canopy openness is a measure that makes the contrast particularly clear, with the agroforests all being significantly more open than the forest reference. It can be expected that the light regime, the microclimate and hence habitat conditions are vastly different in the studied agroforests when compared to forests. We conclude that agroforestry systems in central Europe, with the exception of silvopastures, do not exactly mimic ‘forest structures’ and project them into agricultural landscapes, but rather represent new structural formations, different from forests, but with their very own benefits. Hence, we hypothesize that there is a significant improvement in structural complexity if agricultural systems are converted to agroforests by the inclusion of woody perennials.
We suggest that future research should focus on an assessment of structural complexity of agroforest systems in other geographical location and of different kind, such as tropical agroforestry systems, in order to provide objective information on the structural pattern in those systems as well. It may be considered to quantify the differences among structural pattern always with respect to local silvicultural systems, as conducted here, in order to provide some reference. Finally, we argue it may be important to scan sites in locations with seasonal climates in summer to avoid effects introduced by coniferous vs. deciduous species and to ensure better comparability with sites in the evergreen forests.
Availability of data and material
Data is available at the corresponding author upon request.
Code availability
Mathematica code can be made available upon request. Please contact the corresponding author. R-code for SSCI is available at https://github.com/ehbrechtetal/Stand-structural-complexity-index-SSCI
References
Anderson KE, Glenn NF, Spaete LP, Shinneman DJ, Pilliod DS, Arkle RS et al (2017) Methodological considerations of terrestrial laser scanning for vegetation monitoring in the sagebrush steppe. Environ Monit and Assess 189(11):578
Bates FS, Harris S (2009) Does hedgerow management on organic farms benefit small mammal populations? Agr Ecosyst Environ 129(1–3):124–130
Bengtsson J, Ahnström J, Weibull A-C (2005) The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J Appl Ecol 42:261–269
Böhm C, Tsonkova P, Albrecht E, Zehlius-Eckert W (2017) Zur Notwendigkeit einer kontrollfähigen Definition für Agroforstschläge. Agrar- und Umweltrecht 1(2017):7–12
Bohn Reckziegel R, Larysch E, Sheppard JP, Kahle HP, Morhart C (2021) Modelling and comparing shading effects of 3D tree structures with virtual leaves. Remote Sens 13(3):532
Cardoso P, Barton PS, Birkhofer K, Chichorro F, Deacon C, Fartmann T et al (2020) Scientists’ warning to humanity on insect extinctions. Biol Conserv 242:108426
Christian DP, Collins PT, Hanowski JM, Niemi GJ (1997) Bird and small mammal use of short-rotation hybrid poplar plantations. J Wildl Manage 61(1):171–182
Ehbrecht M, Schall P, Ammer C, Seidel D (2017) Quantifying stand structural complexity and its relationship with forest management, tree species diversity and microclimate. Agr Forest Meteorol 242:1–9
Ehbrecht M, Schall P, Ammer C, Fischer M, Seidel D (2019) Effects of structural heterogeneity on the diurnal temperature range in temperate forest ecosystems. For Ecol Manage 432:860–867
Ehbrecht M, Seidel D, Annighöfer P, Kreft H, Köhler M, Zemp CD, Puettmann K, Nilus R, Babweteera F, Willim K, Stiers M, Soto D, Boehmer HJ, Fisichelli N, Burnett M, Juday G, Stephens SL, Ammer C (2021) Global patterns and climatic controls of forest structural complexity. Nat Commun 12:519
FAO (2015) FRA 2015. Forest ressource assessment. Terms and definitions. Food and agricultural organization of the United Nations, Rome, Italy, 31 p
Fischer M, Bossdorf O, Gockel S, Hansel F, Hemp A, Hessenmoller D, Korte G, Nieschulze J, Pfeiffer S, Prati D et al (2010) Implementing large-scale and long-term functional biodiversity research. Biodivers Explorat Basic Appl Ecol 11:473–485
Frey J, Joa B, Schraml U, Koch B (2019) Same viewpoint different perspectives—a comparison of expert ratings with a TLS derived forest stand structural complexity index. Remote Sens 11(9):1137
Frey J, Asbeck T, Bauhus J (2020) Predicting tree-related microhabitats by multisensor close-range remote sensing structural parameters for the selection of retention elements. Remote Sens 12(5):867
Gailly R, Paquet JY, Titeux N, Claessens H, Dufrêne M (2017) Effects of the conversion of intensive grasslands into Christmas tree plantations on bird assemblages. Agr Ecosyst Environ 247:91–97
Gallé R, Gallé-Szpisjak N, Torma A (2017) Habitat structure influences the spider fauna of short-rotation poplar plantations more than forest age. Eur J For Res 136(1):51–58
Goldewijk K, Beusen A, Van Drecht G, De Vos M (2011) The HYDE 3.1 spatially explicit database of human-induced global land-use change over the past 12,000 years. Global Ecol Biogeogr 20(1):73–86
Grevé ME, Hager J, Weisser WW, Schall P, Gossner MM, Feldhaar H (2018) Effect of forest management on temperate ant communities. Ecosphere 9(6):e02303
Habel JC, Ulrich W, Biburger N, Seibold S, Schmitt T (2019) Agricultural intensification drives butterfly decline. Insect Conserv Diver 12:289–295
Harvey CA, Medina A, Sánchez DM, Vílchez S, Hernández B, Saenz JC et al (2006) Patterns of animal diversity in different forms of tree cover in agricultural landscapes. Ecol Appl 16(5):1986–1999
Heikkinen RK, Luoto M, Virkkala R, Raino K (2004) Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural–forest mosaic. J Appl Ecol 41:824–835
Juchheim J, Ehbrecht M, Schall P, Ammer C, Seidel D (2020) Effect of tree species mixing on stand structural complexity. Forestry Int J Forest Res 93(1):75–83
Kajtoch Ł (2017) The importance of traditional orchards for breeding birds: the preliminary study on Central European example. Acta Oecol 78:53–60
Laliberte AS, Ripple WJ (2004) Range contractions of North American carnivores and ungulates. Bioscience 54(2):123–138
MacArthur RH, MacArthur JW (1961) On bird species diversity. Ecology 42(3):594–598
Manning AD, Fischer J, Lindenmayer DB (2006) Scattered trees are keystone structures—implications for conservation. Biol Conserv 132:311–321
Manning AD, Gibbons P, Lindenmayer DB (2009) Scattered trees: a complementary strategy for facilitating adaptive responses to climate change in modified landscapes? J Appl Ecol 46(4):915–919
McElhinny C, Gibbons P, Brack C, Bauhus J (2005) Forest and woodland stand structural complexity: its definition and measurement. For Ecol Manage 218(1–3):1–24
McGarigal K, Marks BJ (1995) Spatial pattern analysis program for quantifying landscape structure. Gen. Tech. Rep. PNW-GTR-351. US Department of Agriculture, Forest Service, Pacific Northwest Research Station, pp 1–122
Michel N, Burel F, Butet A (2006) How does landscape use influence small mammal diversity, abundance and biomass in hedgerow networks of farming landscapes? Acta Oecol. 30(1):11–20
Nair PR (1993) An introduction to agroforestry. Kluwer Academic Publishers, Netherlands, p 499p
Nerlich K, Graeff-Hönninger S, Claupein W (2013) Agroforestry in Europe: a review of the disappearance of traditional systems and development of modern agroforestry practices, with emphasis on experiences in Germany. Agroforestry Syst 87:475–492
Putman RJ (1997) Deer and road traffic accidents: options for management. J Environ Manage 51(1):43–57
Reidsma P, Tekelenburg T, Van den Berg M, Alkemade R (2006) Impacts of land-use change on biodiversity: an assessment of agricultural biodiversity in the European Union. Agr Ecosyst Environ 114(1):86–102
Reisner Y, De Filippi R, Herzog F, Palma J (2007) Target regions for silvoarable agroforestry in Europe. Ecol Eng 29(4):401–418
Ringler A, Roßmann D, Steidl L (1997) Hecken und Feldgehölze Landschaftspflegekonzept Bayern, Band II.12. Alpeninstitut GmbH, Bremen. Bayerisches Staatsministerium für Landesentwicklung und Umweltfragen (StMLU) und Bayerische Akademie für Naturschutz und Landschaftspflege, Munich, Germany, 523 pp
Šálek M, Václav R, Sedláček F (2020) Uncropped habitats under power pylons are overlooked refuges for small mammals in agricultural landscapes. Agr Ecosyst Environ 290:106777
Seibold S, Gossner MM, Simons NK, Blüthgen N, Müller J, Ambarli D et al (2019) Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 574(7780):671–674
Seidel D, Ehbrecht M, Puettmann K (2016) Assessing different components of three-dimensional forest structure with single-scan terrestrial laser scanning: a case study. For Ecol Manage 381:196–208
Seidel D, Ehbrecht M, Annighöfer P, Ammer C (2019) From tree to stand-level structural complexity- a case study from a temperate broad-leaved forest. Agr Forest Meteorol 278:107699
Seidel D, Annighöfer P, Ehbrecht M, Magdon P, Wöllauer S, Ammer C (2020) Deriving stand structural complexity from airborne laser scanning data—what does it tell us about a forest? Remote Sens 12(11):1854
Sheppard JP, Bohn Reckziegel R, Borrass L, Chirwa PW, Cuaranhua CJ, Hassler SK, Hoffmeister S, Kestel F, Maier R, Mälicke M, Morhart C, Ndlovu NP, Veste M, Funk R, Lang F, Seifert T, du Toit B, Kahle HP (2020) Agroforestry: an appropriate and sustainable response to a changing climate in Southern Africa? Sustainability 12(17):6796
Somarriba E (1992) Revisiting the past: an essay on agroforestrydefinition. Agroforestry Syst 19:233–240
Stiers M, Willim K, Seidel D, Ehbrecht M, Kabal M, Ammer C, Annighöfer P (2018) A quantitative comparison of the structural complexity of managed, lately unmanaged and primary European beech (Fagus sylvatica L.) forests. For Ecol Manage 430:357–365
Tscharntke T, Clough Y, Bhagwat SA, Buchori D, Faust H, Hertel D et al (2011) Multifunctional shade-tree management in tropical agroforestry landscapes—a review. J Appl Ecol 48(3):619–629
Verheyen K, Buggenhout M, Vangansbeke P, De Dobbelaere A, Verdonckt P, Bonte D (2014) Potential of short rotation coppice plantations to reinforce functional biodiversity in agricultural landscapes. Biomass Bioenerg 67:435–442
Willim K, Stiers M, Annighöfer P, Ehbrecht M, Kabal M, Ammer C, Seidel D (2019) Assessing understory complexity in beech-dominated forests (Fagus sylvatica L.) in Central Europe—from managed to primary forests. Sensors 19(7):1684
Woodgate W, Jones SD, Suarez L, Hill MJ, Armston JD, Wilkes P et al (2015) Understanding the variability in ground-based methods for retrieving canopy openness, gap fraction, and leaf area index in diverse forest systems. Agri For Met 205:83–95
Zemp DC, Ehbrecht M, Seidel D, Ammer C, Craven D, Erkelenz J et al (2019) Mixed-species tree plantings enhance structural complexity in oil palm plantations. Agr Ecosyst Environ 283:106564
Zheng G, Moskal LM, Kim SH (2013) Retrieval of effective leaf area index in heterogeneous forests with terrestrial laser scanning. IEEE Trans Geosci Remote Sens 51(2):777–786
Acknowledgements
The German Research Foundation (DFG) is acknowledged for funding this research through grant SE2383/7-1 provided to Dominik Seidel. The help of Maria Hollmann and Anna-Jurina Finkenstaedt during planning and scanning is much appreciated. We thank the managers of the three Exploratories, Kirsten Reichel-Jung, Swen Renner, Katrin Hartwich, Sonja Gockel, Kerstin Wiesner, and Martin Gorke for their work in maintaining the plot and project infrastructure; Christiane Fischer and Simone Pfeiffer for giving support through the central office, Michael Owonibi for managing the central data base, and Markus Fischer, Eduard Linsenmair, Dominik Hessenmöller, Jens Nieschulze, Daniel Prati, Ingo Schöning, François Buscot, Ernst-Detlef Schulze, Wolfgang W. Weisser and the late Elisabeth Kalko for their role in setting up the Biodiversity Exploratories project.
Funding
Open Access funding enabled and organized by Projekt DEAL. Funding was provided to Dominik Seidel through the German Research Foundation, Grant SE2383/7–1.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Consent for publication
All authors agreed to the publication of the submitted manuscript.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Seidel, D., Stiers, M., Ehbrecht, M. et al. On the structural complexity of central European agroforestry systems: a quantitative assessment using terrestrial laser scanning in single-scan mode. Agroforest Syst 95, 669–685 (2021). https://doi.org/10.1007/s10457-021-00620-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-021-00620-y