
Abstract
Lower Devonian (Pragian-Emsian) reefal deposits of Sierra Morena (SW Spain) contain locally abundant calcified cyanobacteria, calcareous algae, and various microfossils including foraminifers. Calcified cyanobacteria are represented by Girvanella spp. A–C, which form crusts and clumps of various shapes. Supposed green algae (?Dasycladales) are represented by a new genus with one new species, Bediaella hispanica gen. et sp. nov. Algospongia include Vasicekia margaritula (Saltovskaya, 1986) n. comb. Microproblematica are represented by Rothpletzella sp. The studied assemblages indicate photic and warm conditions in a shallow and well-agitated environment with normal salinity, and probably mirror episodes of shallowing due to eustatic sea level fluctuations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Among the typical reef-building organisms such as corals and stromatoporoids, various calcified cyanobacteria and calcareous algae were involved in the reef growth during the Palaeozoic (e.g., Chuvashov and Riding 1984; Adachi et al. 2006, 2007). Their role in Palaeozoic reefs has been repeatedly highlighted (e.g., Antoshkina 1996; Wood 2000, 2004; Stephens and Sumner 2003; Antoshkina and Königshof 2008; Liu et al. 2016, 2017, 2021). However, the diversity and palaeoecology of calcified microfossils from Lower Devonian reefal sediments is still poorly understood. Development of Early Devonian reefs was suppressed by various factors, e.g., low sea level and increase in terrigenous influx (e.g., Flügel and Flügel-Kahler 1992; Boulvain and Wood 2007), apparently linked to the closing of the Iapetus ocean, uplift and retreat of large shallow seas, and plate motion reorganizing sea-surface current patterns in the tropics (Copper 2002) with consequent climatic impacts (e.g., Joachimski et al. 2009). Few Lower Devonian reefs have been reported with significant involvement of calcified cyanobacteria and calcareous algae (e.g., Flajs and Hüssner 1996; Antoshkina and Königshof 2008; Hubmann and Suttner 2007; Hubmann and Messner 2013; Koeshidayatullah et al. 2016).
Reefal sediments from the Lower Devonian of Spain were not previously known containing calcified cyanobacteria and/or calcareous algae. Lower to Middle Devonian rocks crop out in the mountains located between the rivers Zújar and Guadámez and to the southeast of them in the Ossa-Morena Zone (SW Spain). These outcrops are part of a long band of Palaeozoic outcrops 200 km long between Badajoz and Córdoba (Fig. 1). The Devonian succession comprises more than 600 m of shales and sandstones that are interbedded with some limestones and marls. The limestones show reefal features of limited lateral and vertical extent. Some of the best outcrops of the reefal facies are, from northwest to southeast, the sections called Guadámez-2, Arroyo del Lobo, Zújar, Peñón Cortado, and Arroyo del Pozo del Rincón (Rodríguez-García 1978). Most of these localities contain Pragian reefal facies and are best exposed in the Peñón Cortado section. Only the Guadámez-2 section contains Emsian reefs. During a field trip in 2007, abundant material from the reefal and peri-reefal facies was collected. These samples were used for description of the rich bryozoan fauna (Ernst and Rodríguez 2023). However, the study of ca 200 thin sections showed the presence of rich calcified microfossils, among them cyanobacteria, green algae, and various microproblematica. The present paper aims to provide an overview and taxonomic description of calcified microfossils from the Peñón Cortado and Guadámez-2 sections at the Ossa-Morena Zone.

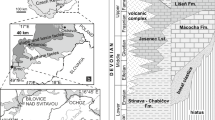
Outcrops of the Lower to Middle Devonian rocks in the Ossa-Morena Zone (SW Spain) (coordinates: Guadámez 2 (38º37′26'' N, 5º48′07''W), Peñón Cortado (38º26′08" N, 5º48′07" W))
Geographical and geological settings
The studied sections reveal the best exposed Lower Devonian sediments in the Sierra Morena. The Peñón Cortado section was named by Herranz (1970). It is located along the railway from Córdoba to Almorchón, in the Angostura valley, 5 km northeast of the village of Valsequillo (Fig. 1; 38º26’08” N, 5º48’07” W). The lower part of the section is composed of shales and marls containing brachiopods, bryozoans, solitary rugosans and tabulate corals. The middle part of the section (units 12–14) contains massive limestones yielding rugose and tabulate corals, stromatoporoids and bryozoans. The upper part of the section comprises again an alternation of limestones and marls containing ramose rugose and tabulate corals, bryozoans, brachiopods, etc. (Fig. 2). The old section was modified to introduce more accurate units. So, the old units 14b to 17 of Rodríguez-García (1978) are changed to units 15–20. The age of the reefal facies is Pragian. The old samples taken in this part of the section are marked in grey and the new samples are marked in black and a B letter in Fig. 3. A fault above the old unit 17 (new 20) repeats a segment of the section in less calcareous facies.

Lithostratigraphic succession at the Peñón Cortado section (thickness of the profile in meters)

Lithostratigraphic succession at the Guadámez-2 section (thickness of the profile in meters)
Two main sections were studied at the bank of the Guadámez River close to the road from Higuera de la Serena to Campillo de Llerena in Badajoz province (Fig. 1) by Rodríguez-García (1978) who named them Guadámez-1 and Guadámez-2 (38º37’26” N, 5º48’07”W). The Guadámez-1 section shows monotonous facies composed of calcareous shales and crinoidal limestones. The Guadámez-2 shows interbedded limestones and marly limestones containing a rich fossil biota including corals, brachiopods, bryozoans, crioconarids, stromatoporoids, ostracods, conodonts, etc. (Fig. 3). The upper part of the section (units 15–19) is composed mainly of stromatoporoids, tabulate corals and bryozoans that built middle-sized (15–20 m wide, 2–3 m thick) bioherms (Rodríguez et al. 2010). The main facies containing bryozoans are tabulate coral rudstones and stromatoporoid–tabulate coral boundstones of Emsian age (Rodríguez et al. 2010).
Several species of foraminifers were found in the studied thin sections including representatives of two unidentified genera, Gen. indet. 1 (Fig. 8k–l) and Gen. indet. 2 (Fig. 8m) from unit 18 of the Guadámez-2 section. Vicinesphaera sp. (Fig. 8n) from unit 18 and Sphaeroporella sp. were identified from unit 20 of the Guadámez-2 section, whereas Parathurammina sp. (Fig. 8p) was found in unit 20 of the Peñón Cortado section.
Materials and methods
The material for the study was collected in autumn 2007 and represents a series of samples taken from the Peñón Cortado and Guadámez-2 sections. From these rock samples, more than 200 thin sections were made (24 × 48 mm, 50 × 50 mm). Some of the thin sections were provided by SR. In 74 thin sections (29 and 46 from Peñón Cortado and Guadámez-2 sections, respectively) calcified cyanobacteria and algae were found. This material is housed at the Senckenberg Museum (Frankfurt am Main, Germany), under the numbers SMF 40580–SMF 40692. The thin sections were studied using an Olympus SZX7 binocular microscope.
In the Emsian Peñón Cortado section, samples collected from units 1, 12, and 17–19 contained various Girvanella species, the calcified microproblematicum Rothpletzella sp., and the green alga Bediaella hispanica gen. et sp. nov. (units 17–19).
The samples from units 1 and 12 comprise floatstones with micritic cements containing bryozoans, tabulate corals, echinoderms, brachiopods and a few gastropods as macrocomponents (Fig. 4b). Unit 14 is a boundstone composed mainly of massive and fasciculate rugose corals, favositids and stromatoporoids. Units 13, 15 and 16 are rudstones composed of the same bioconstructors plus diverse bryozoans, ostracods, echinoderms and brachiopods. The samples from units 17–19 represent rudstones and in part boundstones (Fig. 4a). The matrix contains abundant fragments of bryozoans, ostracod carapaces, echinoderms, brachiopods and microbial mats. Cements are mainly sparitic, locally dolomitized. Larger components are represented by bryozoans, brachiopods and echinoderms. Notable is the presence of large gastropods with calcitic shells and only a few corals. The majority of fragments are enveloped in microbial crusts.

Microfacies of the studied rocks at the Peñón Cortado section. a bryozoan-rich marly limestone, between units 17 and 20, SMF 40580; b bryozoan–crinoid rudstone, unit 12, SMF 40581. Microfacies of the studied rocks at the Guadámez-2 section. c SMF 40582, microbial boundstone with corals, molluscs, and bryozoans, unit G 19, d SMF 40583, bryozoan–crinoid rudstone, unit G 18
The Pragian Guadámez-2 section also contained calcified cyanobacteria (Girvanella) and microproblematica (Rothpletzella). The green alga Bediaella hispanica gen. et sp. nov. was not found in this section. The samples from units G 18–G 20 contain numerous crusts of Girvanella spp. and less abundant Rothpletzella sp. In addition, various other calcified microproblematica and algae were found in these units, among them Vasicekia margaritula (Saltovskaya, 1986) (former Rhabdoporella, see below).
The microfacies of units G 18–G 20 are represented by rudstones with abundant bryozoans, brachiopods, crinoids and a few corals (Fig. 4d). Smaller components include numerous ostracods, smaller bryozoans and diverse skeletal fragments. Cements are predominantly sparitic.
Systematic descriptions
Phylum: Cyanobacteria Stanier, 1974.
Order: Osillatoriales Elenkin, 1949.
Genus: Girvanella Nicholson and Etheridge, 1878.
Type species: Girvanella problematica Nicholson and Etheridge, 1878 emend. Wood, 1957; Upper Ordovician, Scotland.
The studied samples from units 12 and 17–19 of the Peñón Cortado section and units G 18–G 21 of the Guadámez-2 section contain crusts and globular masses, which are formed by tube-shaped remains placed in the genus Girvanella Nicholson and Etheridge, 1878. This fossil is described as calcareous tubular filaments forming nodules and encrusting masses with uniform external diameter. The filaments are long, sinuous to irregularly tangled and have thin micritic walls (e.g., Feng et al. 2010). Girvanella resembles Subtifloria Maslov, 1956 but differs in the disordered arrangement of the filaments. It is regarded as a cyanobacterium (e.g., Riding 1977).
In the studied samples at least three forms can be distinguished. All encrust various components such as bryozoans, brachiopod and gastropod shells, as well as ephemeral objects resulting in the formation of tubular structures after the decay of these soft-bodied organisms (e.g., Fig. 5a, l). Separate sheets and globular masses also occur (e.g., Fig. 5e–h).

Calcified cyanobacteria from the Lower Devonian (Pragian) of SW Spain (all from the unit 20 of the Peñón Cortado section). a–d Girvanella sp. A, tubular encrusting of filaments, SMF 40594; e separate crust of Girvanella sp. A and other microbial mats in the sparitic matrix, SMF 40592; f, g transverse section of the trepostome bryozoan Leptotrypella armata Ernst et al., 2012, encrusted by Girvanella sp. B, SMF 40603; h, i crust formed by Girvanella sp. B, SMF 40595; j crust formed by an assemblage of various Girvanella tubes and microbial crusts, SMF 40604; k crusts of Girvanella sp. A encrusting an ephemeral object (left top) and G. sp. B (right bottom), SMF 40601; l crusts of Girvanella sp. A (left) and G. sp. B (right) in the sparitic matrix, SMF 40606; m, n crust of Girvanella sp. C on bryozoan Leptotrypella armata Ernst et al., 2012, SMF 40591
Girvanella sp. A
Description: Relatively loosely arranged tubes with uniform wall thickness (Fig. 5a–e) forming crusts 0.18–1.00 mm in thickness. Tubes 23–30 µm in diameter with 5–8 µm thick walls.
Remarks: Girvanella sp. A is similar to G. kasakiensis Maslov, 1949.
Girvanella sp. B
Description: Compact masses measuring 0.16–0.81 mm in thickness (Fig. 5f–k), formed by tubes 15–20 µm in diameter with 2–5 µm thick walls.
Remarks: The masses of this species look darker than those of Girvanella sp. A, apparently because of denser packing of the micritic tubes. Alternatively, the presence of fine micrite between the tubes, precipitated within the microbial mat, may be responsible for the dark color of these masses. The difference between the two species of Girvanella is especially evident in Fig. 5l. Girvanella sp. B is similar to G. problematica Nicholson and Etheridge, 1878, as described in Liu et al. (2021).
Girvanella sp. C
Description: Dense masses of small tubes (Fig. 5m–n). Tubes 8–13 µm in diameter with walls of ca 2 µm thickness.
Remarks: Girvanella sp. C differs from G. sp. A and G. sp. B by its smaller tubes (8–13 µm vs. 23–30 µm in G. sp. A and 15–20 µm in G. sp. B).
Calcified microproblematica
Genus: Rothpletzella Wood, 1948
Thin sections from units 12 and 19 of the Peñón Cortado section as well as those from the units G 18–G 20 of the Guadámez-2 section contain fossils which are placed in the genus Rothpletzella Wood, 1948. This microfossil is regarded as a cyanobacterium (Wood 1948), or a microproblematicum (Riding 1991) as no modern analogues are known.
Rothpletzella sp.
Description: Calcareous tubular filaments with micritic walls bifurcating to form sheet-like flat to undulose layers, resembling strings of beads in transverse section. Layers 0.18–0.63 mm in thickness (Fig. 6a–e), rarely concentric in structure (Fig. 6f). Filament internal short dimension (tube height) 20–40 µm, thickness of vertical walls 3–8 µm, and that of horizontal walls (between layers) 10–15 µm.

Calcified microproblematica and Bediaella hispanica gen. et sp. nov., new green alga from the Lower Devonian (Pragian) of SW Spain (a–c, f–n from the unit 20, d, e from the unit 12 of the Peñón Cortado section). a–c crust of Rothpletzella sp. (with intercalations of Girvanella sp. A), SMF 40614; d crust of Rothpletzella sp. on a trepostome bryozoan Anomalotoechus sp., SMF 40620; e globular mass of Rothpletzella sp., SMF 40622; f oncoid of Rothpletzella sp., SMF 40619; g, h Bediaella hispanica gen. et sp. nov., oblique axial section of the thallus; holotype SMF 40643; i Bediaella hispanica gen. et sp. nov., tangential section of the wall with chambers containing apparent cysts, holotype SMF 40643; j Bediaella hispanica gen. et sp. nov., tangential section of the wall with chambers containing apparent cysts, paratype SMF 40645; k, l Bediaella hispanica gen. et sp. nov., transverse section of the thallus, holotype SMF 40643; m, n Bediaella hispanica gen. et sp. nov., longitudinal section, paratype SMF 40648
Calcareous algae
Kingdom: Plantae sensu Copeland, 1956.
Division: Chlorophyta Reichenbach, 1828, emend. Pascher, 1914, emend. Lewis and McCourt, 2004.
Class: Ulvophyceae Stewart and Mattox, 1978.
Order: Dasycladales Pascher, 1931.
Genus: Bediaella gen. nov.
Type species: Bediaella hispanica gen. et sp. nov.
Etymology: The new genus is dedicated to Isabel Méndez Bedia from Oviedo University in Spain.
Occurrence: Peñón Cortado, Ossa-Morena Zone (SW Spain); Lower Devonian (Pragian).
Diagnosis: Subcylindrical thallus not bifurcated with rounded ends. Central cavity relatively broad. Calcareous wall of sparry calcite, probably originally aragonitic, with subspherical, vesiculifer, aspondylous laterals.
Remarks: Bediaella. gen. nov. differs from Nigrella Mamet and Préat, 1994 by the presence of elongate vesiculiform (barrel-shaped) laterals. Bediaella differs from Amicus Maslov, 1956, which has only one narrow central siphon. Due to its apparent uniaxial construction, Amicus was attributed to the primitive Dasycladales. Other Devonian taxa have similarities with Dasycladales: Zeapora Penecke, 1894 emend. Hubmann, 2000 (= Litanaella; see also Hubmann and Reuter 2013, 2017; Hubmann and Messner 2013); Coelotrochium Schlüter, 1879; Scribroporella Spriesterbach, 1935; and Hoegenites Nitecki and Spjeldnæs, 1989 (see discussion about this latter genus in Nitecki and Spjeldnæs 1989; Mamet and Préat 2005; Khodjanyazova and Mamet 2003; Mamet and Khodjanyazova 2006; Vachard and Cózar 2010; Rhebergen 1997, 2009; van Keulen and Rhebergen 2017).
Furthermore, Bediaella differs from Wagonella Mamet and Préat, 1994, a very poorly known genus, by the more irregular calcification of the interpores. Bediaella differs from Gyroporella Gümbel, 1872, by its more spherical laterals (not drum-shaped) and more aspondylous arrangement (this of Gyroporella is sometimes denominated mesospondylous, because it is almost euspondylous). Moreover, Gyroporella appears in the Moscovian (Racz 1966; Kochansky-Devidé 1970). In addition, two contiguous specimens in our material display a growth entirely unknown in Gyroporella (Fig. 7).
The new genus differs also from Uragiellopsis Vachard in Vachard and Montenat, 1981 by the absence of intercalated acrophorous laterals, and from Megaporella Deloffre and Beun, 1986 by the presence of a narrow extremity in the laterals, inconspicuous in our material.
Bediaella hispanica gen. et sp. nov.

Bediaella hispanica gen. et sp. nov., a–c longitudinal section, paratype SMF 40650
Etymology: The new species is named after Spain where it was found.
Holotype: Thin section SMF 40643 (Fig. 6g–i).
Paratypes: Seven thin sections (SMF 40644–SMF 40650).
Repository of the types: Senckenberg Museum (Frankfurt am Main, Germany).
Type locality: Peñón Cortado, Ossa-Morena Zone (SW Spain).
Type horizon: Lower Devonian (Pragian).
Material: More than ten specimens identified in eight thin sections, in longitudinal, oblique and transverse orientations.
Diagnosis: As for the genus, by monotypy.
Description: Seven thin sections from unit 20 contain remnants of an apparent green alga (Dasycladales). This fossil comprises a tubular, probably tapering body with a recrystallized wall 0.08–0.11 mm in thickness. The tubes are 0.54–0.97 mm in diameter. The wall contains densely spaced oval laterals, 0.05–0.09 mm in width (Fig. 6g–n; Table 1). The laterals were not observed with openings either internally or externally. Majority of chambers contain mostly spherical, sometimes collapsed, thin-walled bodies, which may represent cysts (Fig. 6i, j). This fossil is interpreted as possibly belonging to the green algae (Dasycladales) because of its general shape and recrystallized walls that suggest an original aragonitic mineralization. Two contiguous specimens display a growth rarely observed among the Dasycladales (Fig. 7a–c).
Occurrence: As for the genus.
Algae incertae sedis
Class: Algospongia Termier et al., 1977 ex Vachard and Cózar, 2010.
Order: Moravamminales Pokorny, 1951 ex Vachard in Termier et al., 1975.
Family: Issinellaceae Deloffre, 1987 emend. Vachard and Cózar, 2010.
Genus: Vasicekia Pokorny, 1951.
Type species: Vasicekia moravica Pokorny, 1951.
Diagnosis: Thallus cylindrical to claviform, simple or once or several times bifurcated. Numerous thin radiate, aspondylous, cylindrical laterals. Wall calcitic, yellowish.
Description: The morphological diversity may be described following the work of Saltovskaya (1986) in which the genus is misinterpreted as Rhabdoporella.
Included species: Vasicekia moravica Pokorny, 1951; Vasicekia? sp. of Mamet et al. (1999, pl. 4, Figs. 1–3); Parmacaulis hulgensis Shuysky and Shirshova in Chuvashov et al., 1987; Rhabdoporella perfecta Saltovskaya, 1986; Rhabdoporella fragile Saltovskaya, 1986; Rhabdoporella elongata Saltovskaya, 1986; Rhabdoporella clavaeformis Saltovskaya, 1986; Rhabdoporella margaritula Saltovskaya, 1986.
Remarks: These taxa differ from true Rhabdoporella by their cylindrical and not acrophore laterals, yellowish and probably originally calcitic wall, and not aragonitic as in the true dasycladales. Hence, they are comparable with Issinella, Serrisinella, Issinellina, etc. rather than Rhabdoporella.
Rhabdoporella sensu Vachard and Gélard 1980 is a primitive dasycladale Seletonellaceae relatively similar to the Dasyporelleae. Rhabdoporella sensu Pia 1920, and perhaps Poncet (1986), is another taxon with an aspondylous arrangement of acrophore laterals. Vasicekia differs by its typical issinellacean laterals (cylindrical, very thin, aspondylous) and the yellowish, calcitic wall (never recrystallized microsparite). Issinella, Issinellina and Serrisinella are cylindrical thalli; Vasicekia being claviform.
Moreover, Rhabdoporella is bifurcated and segmented (Serrisinella is also weakly segmented). Vasicekia has a thin wall and a proportionally large axial cavity. Even if Vasicekia was initially described as a foraminifer by Pokorny (1951). Subsequently, Mamet et al. (1999), Vachard and Cózar (2010), and Mamet and Préat (2013) considered Vasicekia as an alga.
Occurrence: Upper Ordovician–Lower Silurian of Tajikistan, Caucasus, England, Gotland, Prebaltica (Saltovskaya 1986). It is noteworthy that this dating is disputable in Russia and has been revised as Lower Devonian (Vachard et al. 2010; Nestell et al. 2011). Lower Devonian of Spain (present publication). Lower to Middle Devonian of Northern and Prepolar Urals. Eifelian of Morocco. Givetian of Moravia. Givetian of western France (Vachard and Cózar 2010). Middle Devonian of Poland (Halamski et al. 2022; Gajewska 2022).
Vasicekia margaritula (Saltovskaya, 1986) n. comb.
Figures 8a–j

Vasicekia margaritula (Saltovskaya, 1986) n. comb., from the Lower Devonian (Emsian) of SW Spain, Guadámez-2 section, unit G 18. a–f transverse sections showing the very fine aspondylous, acrophore, radiate perforations in the wall (8a—SMF 40659, 8b—SMF 40656, 8c—SMF 40661, 8d—SMF 40664, 8e–SMF 40664, 8f—SMF 40677); g–j, oblique axial sections showing the claviform thallus and the very fine aspondylous, acrophore, radiate perforations in the wall (8g—SMF 40658, 8h—SMF 40653, 8i—SMF 40664, 8j—SMF 40655). Foraminifers found in the studied profiles of the Lower Devonian of SW Spain: k–l Gen. indet. 1 (8k—SMF 40687, 8l—SMF 40688); m Gen. indet. 2 (SMF 40689); n Vicinesphaera sp. (SMF 40690); o Parathurammina sp. (SMF 40691); p Sphaeroporella sp. (SMF 40692)
Material: More than 50 specimens identified in 36 thin sections, in longitudinal, oblique and transverse sections (SMF 40651–SMF 40686).
Diagnosis: Thallus claviform with well-preserved wall.
Description: Thirty-six thin sections in the sample from unit G 18 of the Guadámez-2 section contain remnants of an apparent issinellacean (incertae sedis algae). This fossil is tubular, probably claviform, with well-preserved walls, and 0.35–0.43 mm in length. The tubes are 0.20–0.29 mm in diameter. The wall, 0.025–0.050 mm thick, contains densely spaced cylindrical laterals, 0.05–0.12 mm in width (Fig. 8g–j). The laterals are all open either internally or externally (Fig. 9)

.
Occurrence: Upper Ordovician of Tajikistan (Saltovskaya 1986). Discovered in the Lower Devonian of Spain (unit G 18 of the Guadámez-2 section).
Discussion
Calcified cyanobacteria and algae were found only within narrow intervals in two localities of the Lower Devonian in SW Spain: Peñón Cortado (Pragian) and Guadámez-2 (Emsian). From the total sequence of these two localities, more than 200 thin sections were made for study of bryozoans (Ernst and Rodríguez 2023).
Distribution within studied sections
Girvanella spp. A–C occur in units 1, 12 and 17–19 of the Peñón Cortado section and units G 18–G 21 of the Guadámez-2 section, whereas Rothpletzella sp. was found in units 12 and 19 of the Peñón Cortado section, and in units G 18–G 20 of the Guadámez-2 section (Table 2). Units G 18–G 21 represent a succession deposited on a protected shallow platform with patch reefs, normal salinity and moderate water energy (Rodríguez et al. 2010).
Three types of Girvanella sp. were identified within the studied units. They differ in size and arrangement of filaments. Girvanella sp. A produced larger relatively loosely arranged filaments, whereas those of G. sp. B and C are smaller and more densely packed (see description above). All three varieties appear as masses and crusts, Girvanella sp. A and B were observed in tubular congregations, apparently around ephemeral objects (e.g., algae).
Rothpletzella sp. produces crusts on bryozoans and corals, as well free-lying globular masses which can reveal a concentric arrangement (oncoid, Fig. 6f). Rothpletzella sp. is often intercalated with Girvanella sp. A. Similar co-occurrences of Girvanella and Rothpletzella are quite common in reefal sediments (e.g., Tsien 1979; Liu et al. 2016, 2017, 2021).
The green alga Bediaella hispanica gen. et sp. nov. is restricted to units 18–20 of the Peñón Cortado section. Vasicekia margaritula (Saltovskaya, 1986) was found only in unit G 18 of the Guadámez-2 section. This unit (and the succeeding G 19) represents patch reefs built mainly by thamnoporoids and stromatoporoids (Rodríguez et al. 2010).
Furthermore, various laminated micritic structures of apparently microbial origin are abundant in unit 20 of the Peñón Cortado section (e.g., Figs. 4a, 5e, j, 6b). and in unit G 19 of the Guadámez-2 section (Fig. 4c). They often show indistinct relicts of tubular structures.
Palaeoecology and environmental interpretation
The crusts produced by Girvanella spp. A–C and Rothpletzella sp. often cover bryozoans sealing their apertures (e.g., Figs. 5m–n, 6a–b, d–e). The temporal succession of these overgrowths is indistinct, however. The encrusted bryozoans were probably already dead before the apertures were sealed by crusts of calcareous microorganisms. However, the sealing of living biotic substrates causing death cannot be completely excluded as no unequivocal evidence for either alternative was found in the studied material.
Abundant tubular aggregations (e.g., Fig. 5a, k–l) of Girvanella spp. A–B indicate the presence of ephemeral organisms, probably cyanobacteria. Such bioclaustrations are especially abundant in unit 18 of the Peñón Cortado section, and in unit G 21 of the Guadámez-2 section. These tubes are unbranching and may have represented uncalcified cyanobacteria.
The abundance and diversity of calcified microbial structures observed in the studied units suggests a relatively shallow and agitated environment within the photic zone, as confirmed by the presence of green algae (Bediaella) and algae of uncertain affinity (Vasicekia). The high level of fragmentation of most components in many beds indicates a considerable hydrodynamic energy. The accompanying fauna, including abundant rugose corals, brachiopods and echinoderms, indicates a normal marine salinity and well-oxygenated waters.
Cyanobacterial calcification is believed to require high overall seawater carbonate saturation (e.g., Riding 1992, 2011). Such conditions normally occur during evaporation in restricted lagoons or soda lakes. Although abundant and diverse in marine environments (Hoffman 1999), modern calcimicrobes experience intensive calcification mostly in fresh water and rarely in subtidal marine environments (e.g., Flügel 2010). In contrast, the fossil representatives of this group are found in tidal and subtidal biotopes, often in reefs, as, for example, shown for some Ordovician (Liu et al. 2021) and Devonian occurrences (Stephens and Sumner 2003). Rothpletzella is suggested to occur in shadowed, protected environments like lagoons, but also in reefs and open platform habitats (e.g., Chuvashov and Riding 1984; Antoshkina 1996; Wood 2000, 2004).
Calcareous microfossils and Lower Devonian depositional history
Studied assemblages of calcareous microfossils from the Lower Devonian of the Sierra Morena might mirror regional and global sea level changes. The two main occurrences in the Pragian Peñón Cortado section (units 17–20) and in the Emsian Guadámez-2 section (units G 18–G 20) probably mark peaks of regressive cycles observed during the Pragian and Emsian, which were separated by the transgression near the Pragian/Emsian transition (e.g., Johnson et al. 1985, 1996; Chlupáč and Kukal 1986; Flajs and Hüssner 1996; Keller 1997). Reefal sediments in the Lower Devonian are relatively rare being mainly restricted to lower latitudes and deposition in normal marine conditions (e.g., Copper 2002; Kiessling 2002; Vennin et al. 2007). There are various explanations for this reef reduction including such abiotic factors as high water temperature as well as certain developments in reef-forming biotas (e.g., Joachimski et al. 2009; May 2022). The Lower Devonian was postulated to be an episode of reduced cyanobacterial calcification (Riding 1992). Nevertheless, cyanobacteria and calcareous algae have been reported from different reefal constructions, e.g., in the Lower Devonian of Belgium (Tsien 1979), Austria (Hubmann and Suttner 2007), Prague Basin (e.g., Flajs and Hüssner 1996), Russia (Antoshkina and Königshof 2008), and Saudi Arabia (Koeshidayatullah et al. 2016). Such occurrences of calcified cyanobacteria, alongside other calcified microorganisms and algae, may indicate short intervals of increased carbonate saturation, probably because of basin shallowing and increased temperature.
Conclusion
Certain horizons of the reefal deposits of two Lower Devonian sections in Sierra Morena (SW Spain) contain abundant calcified cyanobacteria, calcareous algae, and various microfossils including the problematicum Rothpletzella and foraminifers. Calcified cyanobacteria (Girvanella spp. A–C) form crusts and clumps of various shapes, notably tubular structures.
A new genus with one new species, Bediaella hispanica gen. et sp. nov., apparently belonging to Dasycladales (green algae), is described from the Lower Devonian (Pragian) sediments of the Peñón Cortado section.
A new combination Vasicekia margaritula (Saltovskaya, 1986) n. comb. (Algospongia) is described from the Lower Devonian (Emsian) deposits of the Guadámez-2 section.
Microproblematica are represented by Rothpletzella sp., which often occurs intercalated with Girvanella sp. A.
Horizons with calcified cyanobacteria and calcareous algae were apparently deposited during brief episodes with higher carbonate saturation as supposed for the Lower Devonian (e.g., Riding 1992). These temporal environmental changes might be linked to regressions due to eustatic sea level fluctuations (e.g., Johnson et al. 1985, 1996). The studied assemblages contain predominantly autotrophic organisms and indicate photic and warm conditions in a shallow and well-agitated environment with normal salinity.
Data availability
Studied material is housed at the Senckenberg Museum (Frankfurt am Main, Germany).
Code availability
Not applicable.
References
Adachi N, Ezaki Y, Pickett JW (2006) Marked accumulation patterns characteristic of Lower Devonian stromatoporoid bindstone: palaeoecological interactions between skeletal organisms and microbes. Palaeogeogr Palaeoclimatol Palaeoecol 231:331–346. https://doi.org/10.1016/j.palaeo.2005.09.002
Adachi N, Ezaki Y, Pickett JW (2007) Interrelations between framework-building and encrusting skeletal organisms and microbes: more refined growth history of Lower Devonian bindstones. Sedimentology 54:89–105. https://doi.org/10.1111/j.1365-3091.2006.00831.x
Antoshkina A (1996) Ordovician reefs of the Ural Mountains, Russia: a review. Facies 35:1–8
Antoshkina A, Königshof P (2008) Lower Devonian reef structures in Russia: an example from the Urals. Facies 54:233–251. https://doi.org/10.1007/s10347-008-0135-7
Boulvain F, Wood R (2007) Devonian. In: Vennin E, Aretz M, Boulvain F, Munnecke A (eds.) Facies from Palaeozoic reefs and bioaccumulations. Muséum national d'Histoire naturelle, Paris, pp. 171–223 (Mémoires du Muséum national d'Histoire naturelle 195)
Chlupáč I, Kukal Z (1986) Reflection of possible Global Devonian events in the Barrandian area, C.S.S.R. In: Walliser OH (ed) Global Bio-Events. Lecture Notes in Earth History 8. Springer, Berlin, pp. 169–179
Chuvashov B, Riding R (1984) Principal floras of Palaeozoic marine calcareous algae. Palaeont 27(3):487–500
Chuvashov BI, Lyuchinina VA, Shuysky VP, Chaikin IM, Berchenko OI, Ishchenko AA, Saltovskaya VD, Shirshova DI (1987) Iskopaemye izvestkovye vodorosli morfologiya, sistematika, metody izucheniya (Fossil calcareous algae, morphology, systematics, methods of study). Akademiya Nauk SSSR, Sibirskoe Otdelenie, Trudy Instituta Geologii i Geofiziki 674:52–24 (in Russian)
Copeland HF (1956) Classification of the lower organisms. Pacific Books, Palo Alto. https://doi.org/10.5962/bhl.title.4474
Copper P (2002) Silurian and Devonian reefs: 80 million years of global greenhouse between two ice ages. SEPM Spec Publ 72:181–138
Deloffre R (1987) Nouvelle classification des algues Dasycladales fossile. Comptes Rendus de l'Académie des Sciences, Serie II 305:1017–1020
Deloffre R, Beun N (1986) Megaporella boulangeri, nouvelle algue dasycladale du Kimméridgien inférieur marocain. Rev Micropal 28(4):233–242
Elenkin AA (1949) Monographia algarum Cyanophycearum aquidulcium et terrestrium in finibus U.R.S.S. inventarum. Fasc. II Moskau u. Leningrad (in Russian)
Ernst A, Fernández LP, Fernández-Martínez E, Vera C (2012) Description of a bryozoan fauna from mud mounds of the Lebanza Formation (Lower Devonian) in the Arauz area (Pisuerga–Carrión Province, Cantabrian Zone, NW Spain). Geodiversitas 34(4):693–738. https://doi.org/10.5252/g2012n4a1
Ernst A, Rodríguez S (2023) Palaeoecology and palaeobiogeographic relationships of Lower Devonian bryozoans from the Guadámez and Peñón Cortado Sections of Sierra Morena (SW Spain). Span Jour Palaeontol 38(2):173–220
Feng Q, Gong Y-M, Riding R (2010) Mid-Late Devonian calcified marine algae and cyanobacteria. South China J Paleont 84(4):569–587. https://doi.org/10.1666/09-108.1
Flajs G, Hüssner H (1996) Lower Devonian coral/stromatoporoid reefs in the Koneprusy area (Czech Republic)–A preliminary report. In: Reitner J, Neuweiler F, Gunkel F (eds.) Global and Regional Controls on Biogenic Sedimentation. I. Reef Evolution. Research Reports. Göttinger Arbeiten zur Geologie und Paläontologie, 2:355–357
Flügel E (2010) Microfacies of carbonate rocks: analysis, interpretation and application. Springer, Berlin Heidelberg. https://doi.org/10.1007/978-3-642-03796-2
Flügel E, Flügel-Kahler E (1992) Phanerozoic reef evolution: basic questions and database. Facies 26:167–278
Gajewska M (2022) Middle Devonian Foraminifera from the Holy Cross Mountains. Ann Soc Geol Pol 92:411–424
Gümbel CW (1872) Die sogenannten Nulliporen (Lithothamnium und Dactylopora) und ihre Betheiligung an der Zusammensetzung der Kalkgesteine. Zweiter Theil: Die Nulliporen des Thierreichs (Dactyloporideae) nebst Nachtrag zum ersten Theile. Abh. d. II. Cl. d. k. Ak. d. Wiss, Math. phys. Cl., München, Band XI, Abh. I, pp. 231–290, 4 pl. h.-t. (D.I-IV)
Halamski AT, Baliński A, Racki G, Amler MRW, Basse M, Denayer J, Dubicka Z, Filipiak P, Kondas M, Krawczyński W, Mieszkowski R, Narkiewicz K, Olempska E, Wrzołek T, Wyse Jackson PN, Zapalski MK, Zatoń M, Kozłowski W (2022) The pre-Taghanic (Givetian, Middle Devonian) ecosystems of Miłoszów (Holy Cross Mts, Poland). Ann Soc Geol Pol 92:323–379
Herranz P (1970) Nota preliminar sobre el estudio geológico de las sierras paleozoicas entre Oliva de Mérida y Hornachos (Badajoz). Seminarios De Estratigrafía 6:1–16
Hubmann B (2000) Zeapora Penecke 1894 redefined: a halimedacean alga. Mitt Österr Geol Ges 91:31–37
Hubmann B, Messner F (2013) Field Trip B3: Lower to Middle Devonian algal limestones of the Graz Palaeozoic (Austria). 11th Workshop on Alpine Geological Studies and 7th IFAA. Berichte Geologische Bundesanstalt 99:271–285
Hubmann B, Reuter M (2013) Zeapora – an endemic Devonian ‘praecodiacean’ of Graz or a common tropical cosmopolitan? 11th Workshop on Alpine Geological Studies and 7th IFAA. Berichte Geologische Bundesanstalt 99:110
Hubmann B, Reuter M (2017) Synonymy and circum-tropical biogeography of the Devonian calcareous alga Zeapora Penecke 1894 (Chlorophyta, Bryopsidales). Isl Arc 26:e12187. https://doi.org/10.1111/iar.12187
Hubmann B, Suttner T (2007) Siluro-Devonian Alpine reefs and pavements. Geol Soc Spec Publ Lond 275:95–107. https://doi.org/10.1144/GSL.SP.2007.275.01.07
Joachimski MM, Breisig S, Buggisch W, Talent J, Mawson R, Gereke M, Morrow JR, Day J, Weddige K (2009) Devonian climate and reef evolution: insights from oxygen isotopes in apatite. Earth Planet Sci Lett 284:599–609. https://doi.org/10.1016/j.epsl.2009.05.028
Johnson JG, Klapper G, Sandberg CA (1985) Devonian eustatic fluctuations in Euramerica. Geol Soc Am Bull 96:567–587
Johnson JG, Klapper G, Elrick M (1996) Devonian transgressive-regressive cycles and biostratigraphy, northern Antelope range, Nevada: establishment of reference horizons for global cycles. Palaios 11:3–14. https://doi.org/10.2307/3515112
Keller M (1997) Evolution and sequence stratigraphy of an Early Devonian carbonate ramp, Cantabrian Mountains, northern Spain. J Sediment Res 67(4):638–652. https://doi.org/10.1306/D4268603-2B26-11D7-8648000102C1865D
Khodjanyazova R, Mamet B (2003) Paleozoic calcareous algae from southern Tien Shan, Uzbekistan, Central Asia. Geol Belg 6:97–117
Kiessling W (2002) Secular variations in the Phanerozoic reef ecosystem. Spec Publ Soc Econ Paleontol Mineral 72:625–690
Kochansky-Devidé V (1970) Die Kalkalgen des Karbons vom Velebit-Gebirge (Moskovien und Kasimovien). Palaeontol Jugoslav 10:1–32
Koeshidayatullah A, Al-Ramadan K, Hughes GW (2016) Facies mosaic and diagenetic patterns of the early Devonian (Late Pragian-Early Emsian) microbialite-dominated carbonate sequences, Qasr Member, Jauf Formation. Saudi Arabia Geol J 51(5):704–721. https://doi.org/10.1002/gj.2678
Lewis LA, McCourt RM (2004) Green algae and the origin of land plants. Am J Bot 91(10):1535–1556. https://doi.org/10.3732/ajb.91.10.1535
Liu LJ, Wu YS, Yang HJ, Riding R (2016) Ordovician calcified cyanobacteria and associated microfossils from the Tarim Basin, Northwest China: systematics and significance. J Syst Palaeont 14:183–210. https://doi.org/10.1080/14772019.2015.1030128
Liu LJ, Wu YS, Jiang HX, Wu NQ, Jia LQ (2017) Paleoenvironmental distribution of Ordovician calcimicrobial associations in the Tarim Basin, Northwest China. Palaios 32:462–489. https://doi.org/10.2110/palo.2016.054
Liu L, Wu Y, Bao H, Jiang H, Zheng L, Chen Y (2021) Diversity and systematics of Middle-Late Ordovician calcified cyanobacteria and associated microfossils from Ordos Basin, North China. J Paleont 95(1):1–23. https://doi.org/10.1017/jpa.2020.82
Mamet B, Khodjanyazova R (2006) Taxonomic note on some Paleozoic marine algae (Uzbekistan, Central Asia). Geobios 39:507–512. https://doi.org/10.1016/j.geobios.2005.04.003
Mamet B, Préat A (1992) Algues du Dévonien moyen de Wellin (Synclinorium de Dinant, Belgique). Rev Micropal 35:53–75
Mamet B, Préat A (1994) Algues calcaires de l’Eifélien inférieur (Calcaire de Couvin, Synclinorium de Dinant). Rev Micropal 37:135–159
Mamet B, Préat A (2005) Microfaciès d’une lentille biohermale à la limite Eifélien-Givétien (Wellin, bord sud du Synclinorium de Dinant). Geol Belg 8:85–111
Mamet B, Préat A (2013) Essai de description d’algues nouvelles paléozoïques. Geol Belg 16(1–2):35–48
Mamet B, Préat A, Lehmami M (1999) Algues calcaires marines du Dévonien marocain (Meseta). Rev Micropal 42(4):301–314
Maslov VP (1956) Iskopaemye izvestkovye vodorosli SSSR. Tr Inst Geol Nauk Akad Nauk SSSR 160:1–302 (in Russian)
Maslov VP (1949) The alga Girvanella, its ecology and stratigraphic significance. Bulletin Moskovskogo Obshchestva Isoytatelei Prirody 54:89–100 (in Russian)
May A (2022) Why were reefs and stromatoporoids so rare in the Lower Devonian? Sciences of Europe 104:24–32. https://doi.org/10.5281/zenodo.7298618
Nestell G, Heredia S, Mestre A, Beresi M, González M (2011) The oldest Ordovician foraminifers (Oepikodus evae conodont Zone, Floian) from South America (Les plus vieux foraminifères (Zone à conodontes Oepikodus evae, Floien) d’Amérique du Sud). Geobios 44:601–608. https://doi.org/10.1016/j.geobios.2011.02.007
Nicholson HA, Etheridge R Jr. (1878) A monograph of the Silurian fossils of the Girvan District in Ayrshire with special reference to those contained in the “Gray Collection.”, Fasciculus 1 (Rhizopoda, Actinozoa, Trilobita). W. Blackwood and Sons, Edinburgh 1 135
Nitecki MH, Spjeldnæs N (1989) A new Ordovician tubular ‘‘alga” from Norway. Nor Geol Tidsskr 69:95–102
Pascher A (1914) Über Flagellaten und Algen. Ber Dtsch Bot Ges 32:136–160
Pascher A (1931) Systematische Übersicht über die mit Flagellaten in Zusammenhang stehenden Algenreihen und Versuch einer Einreihung dieser Algenstämme in die Stämme des Pflanzenreiches. Beihefte Zum Botanischen Centralblatt 48:317–332
Penecke AK (1894) Das Grazer Devon. Jahrbuch Der Kaiserlich-Koniglichen Geologischen Reichansantstalt 48:567–616
Pokorny V (1951) The middle Devonian Foraminifera of Celechovice, Czechoslovakia. Vestnik Kraslovske Ceske Spolecnosti Nauk, Trida Matematicko-Prirodovedecka 9:1–29
Poncet J (1986) Les algues calcaires du Paléozoïque inférieur de la Baie d’Hudson et de l’Archipel arctique canadien. Bull Centres Rech Explor-Prod Elf-Aquitaine 10(2):259–282
Rácz L (1966) Late Paleozoic calcareous algae in the Pisuerga basin (North Palencia, Spain). Leidse Geol Meded 1964(31):241–260
Reichenbach HGL (1828) Conspectus Regni Vegetabilis, p. 23
Rhebergen F (1997) Ordovicische algen II: een vergaartbak. Grondboor and Hamer 51:1–10
Rhebergen F (2009) Ordovician sponges (Porifera) and other silicifications from Baltica in Neogene and Pleistocene fluvial deposits of the Netherlands and northern Germany. Estonian J Earth Sci 58:24–37
Riding R (1977) Calcified Plectonema (blue-green algae), a Recent example of Girvanella from Aldabra Atoll. Palaeont 20:33–46
Riding R (1991) Calcified cyanobacteria. In: Riding R (ed) Calcareous algae and stromatolites. Springer-Verlag, Berlin, pp 55–87
Riding R (1992) Temporal variation in calcification in marine cyanobacteria. J Geol Soc (Lond) 149:979–989
Riding R (2011) Calcified cyanobacteria. In: Reitner J, Thiels V (eds) Encyclopedia of geobiology. Encyclopedia of Earth Science series. Springer, Heidelberg, pp 211–223
Rodríguez S, Fernández-Martínez E, Cózar P, Valenzuela-Ríos JI, Pardo Alonso MV, Liao J-C, May A (2010) Stratigraphic succession, facies and depositional environment of Emsian reefal carbonates in the Ossa-Morena Zone (SW Spain). Neues Jahrb Geol Palaontol Abh 257(1):69–83. https://doi.org/10.1127/0077-7749/2010/0063
Rodríguez-García S (1978) Corales rugosos del Devónico de la Sierra del Pedroso. Estud Geol 34:331–350
Saltovskaya VD (1986) O formakh rosta vodoroslei Rhabdoporella Stolley, 1893 (Dasycladaceae) [On the shapes of the alga Rhabdoporella Stolley, 1893 (Dasycladaceae)]. Vopr Mikropaleontol 28:145–151 (in Russian)
Schlüter C (1879) Coelotrochium decheni, eine Foraminifere aus dem Mitteldevon. Z Dtsch Geol Ges 31:668–675
Spriesterbach J (1935) Beitrag zur Kenntnis der Fauna des rheinischen Devon. Jahrb Preuss Geol Landesanst 55:475–525
Stanier RY (1974) Division I. The Cyanobacteria. In: Buchanan RE, Gibbons NE (eds) Bergey’s Manual of Determinative Bacteriology. William & Wilkins Co, Baltimore, p 22
Stephens NP, Sumner DY (2003) Famennian microbial reef facies, Napier and Oscar Ranges, Canning Basin, western Australia. Sediment 50:1283–1302. https://doi.org/10.1046/j.1365-3091.2003.00605.x
Stewart KD, Mattox KR (1978) Structural evolution in the flagellated cells of green algae and land plants. BioSystems 10(1–2):145–152. https://doi.org/10.1016/0303-2647(78)90036-9
Stolley E (1893) Ueber Silurische Siphoneen. Neues Jahrb Mineral Geol Palaontol, II Band: 35–146
Termier G, Termier H, Vachard D (1977) Etude comparative de quelques Ischyrosponges. Geol Mediterr 4(2):139–180
Tsien HH (1979) Paleoecology of algal-bearing facies in the Devonian (Couvinian to Frasnian) reef complexes of Belgium. Palaeogeogr Palaeoclimatol Palaeoecol 27:103–127. https://doi.org/10.1016/0031-0182(79)90096-8
Vachard D, Cózar P (2010) An attempt of classification of the Palaeozoic incertae sedis Algospongia. Rev Esp Micropaleontol 42:129–241
Vachard D, Montenat C (1981) Biostratigraphie, micropaléontologie et paléogéographie du Permien de la région de Tezak (Montagnes Centrales d’Afghanistan). Palaeontogr Abt B 178:1–88
Vachard D, Pille L, Gaillot J (2010) Paleozoic Foraminifera: systematics, palaeoecology and responses to the global changes (Les foraminifères paléozoïques: systématique, paléoécologie et réponses aux changements globaux). Rev Micropal 53:209–254. https://doi.org/10.1016/j.revmic.2010.10.001
Vachard D, Gélard J-P (1980) Rhabdoporella pachyderma (Algue Dasycladale) dans le Dévonien du massif de Chellata (Grande Kabylie, Algérie). Bulletin mensuel de la Société linnéenne de Lyon, 49 année, n°7, septembre 1980. pp 401–409. http://www.persee.fr/doc/linly_0366-1326_1980_num_49_7_10437.
van Keulen P, Rhebergen F (2017) Typology and fossil assemblage of Sandbian (Ordovician) ‘baksteenkalk’: an erratic silicified limestone of Baltic origin from the northeastern Netherlands and adjacent areas of Germany. Estonian J Earth Sci 66:198–219
Vennin E, Aretz M, Boulvain F, Munnecke A (2007) Facies from Palaeozoic reefs and bio-accumulations. Mémoires du Muséum national d´Histoire naturelle 195:1–341
von Pia J (1920) Die Siphoneae verticillatae vom Karbon bis zur Kreide. Abhandlungen Der Zoologisch-Botanischen Gesellschaft in Wien 11(2):1–263
Wood A (1948) “Sphaerocodium”, a misinterpreted fossil from the Wenlock Limestone. Proc Geol Assoc 59:9–22
Wood R (2000) Palaeoecology of a Late Devonian back-reef: Canning Basin, Western Australia. Palaeontol 43:671–703
Wood R (2004) Palaeoecology of a post-extinction reef: Famennian (Late Devonian) of the Canning Basin, Western Australia. Palaeontol 47:415–445
Acknowledgements
Andrej Ernst is grateful to the Deutsche Forschungsgemeinschaft for financial support (DFG project ER 278/4.1 and 2). Bernhard Hubmann, Graz, and an anonymous reviewer are thanked for their helpful comments and suggestions to the manuscript. Paul D. Taylor, London, is greatly appreciated for his critical reading of the manuscript and helpful corrections.
Funding
Open Access funding enabled and organized by Projekt DEAL. See Acknowledgements. Deutsche Forschungsgemeinschaft, ER 278/4.1 and 2, Andrej Ernst.
Author information
Authors and Affiliations
Contributions
AE designed the research, made the thin sections (in part), measured the material, and described the cyanobacteria and calcified microproblematica. DV provided descriptions of the algae and IDs of foraminifers. SR wrote introduction parts and provided thin sections (in part). AE, DV, and SR contributed to discussion and conclusions.
Corresponding author
Ethics declarations
Conflict of interest
None.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ernst, A., Vachard, D. & Rodríguez, S. Palaeoecology of calcified microfossils from the Lower Devonian (Pragian-Emsian) of Sierra Morena (SW Spain). Facies 70, 6 (2024). https://doi.org/10.1007/s10347-024-00680-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10347-024-00680-3