
Abstract
As land-use cover types affect wintering birds’ strategies, studying their habitat suitability for species conservation is relevant. Predictive spatial models are considered excellent tools for conservation planning and improving our understanding of species distribution. Here we build models to predict the spatial distribution of raptor species that overwinter in SE Spain. We modelled the wintering raptor species distribution based on the presence-only data obtained between 2017 and 2019 and analysed their habitat preferences based on elevation and land-use cover variables. Our results show that altitude and distance to the coastline are the most important environmental factors to affect most species’ habitat suitability. Habitat type-related factors are other important predictors, and raptors subject of this study prefer lowlands and areas close to the coastline for overwintering by selecting open habitats, which include wetlands and irrigated and rainfed crops. This study highlights the importance of the habitat heterogeneity generated by wetlands and herbaceous crops within a low-altitude range as the optimal environment for wintering raptors. Such information should be considered for raptor conservation planning in human-altered landscapes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
How land-use changes induced by anthropogenic activities affect the requirements of many species is one of the key biodiversity conservation challenges (Chapin et al. 2000; Sala et al. 2000; Newbold et al. 2015; Powers and Jetz 2019). Many land-use changes are the consequence of an increased proportion of land used by agriculture in recent centuries, as well as a growing trend towards agricultural intensification (Meyer and Turner 1992; Foley et al. 2005). Agricultural intensification has detrimental effects on natural systems (Stoate et al. 2001; Foley et al. 2005; Guerrero et al. 2012; Grande et al. 2018), biodiversity and key ecosystem services (Landis 2017; Sirami et al. 2019). Such changes may have, in turn, a detrimental effect on the availability of suitable habitats for many species, which leads to distribution shifts (Jetz et al. 2007; Powers and Jetz 2019) or even local population extinction (Jiménez-Franco et al. 2022).
In the case of avian species, the effects of land-use changes vary by species and the intensity of land-use change (Vickery et al. 1999; Fox 2004; Paprocki et al. 2015). This does not mean that human-made landscapes necessarily pose a threat to birdlife. Indeed, for certain species or, at least during the different periods of their life cycle, change may even provide opportunities for foraging or breeding (Cardador et al. 2011; Donázar et al. 2016; Grande et al. 2018). In line with this, cropland areas are used as a stopover habitat by bird species that can utilise a wide breadth of habitats, as well as by species with preferences for habitats with a similar structure to agricultural areas (Blount et al. 2021).
At temperate latitudes, wintering birds’ survival strategies involve adaptations that range from changes in habitat selection to variations in diet, and they are all conditioned by the migratory displacement of which they are capable to search for favourable areas (Senar and Borras 2004; Blount et al. 2021; Chiatante and Panuccio 2021). Considering the evidence that indicates shortened avian migration distances (e.g. Visser et al. 2009; Paprocki et al. 2014), accompanied by poleward broad-scale shifts in distributions ranges in response to rising global temperatures, there are indications that the Mediterranean Basin’s climatic suitability for wintering Palearctic birds could increase (Tellería et al. 2016). However, in certain Mediterranean regions, such as the SE region of the Iberian Peninsula, the total area of natural sites and land used by extensive farming continues to fall in many landscapes because of agricultural intensification and the consequent increased agricultural landscape homogenisation (Caraveli 2000; Valera et al. 2011). The availability of suitable habitats on the regional scale has been identified as the ultimate factor that, in the short term, determines and threatens locality selection by birds during the non-breeding period (La Sorte et al. 2017). This is a gap in our knowledge. As migratory birds spend up to 85% of their total migration time at stopover sites (Blount et al. 2021), identifying important suitable areas in this stage is a priority for setting up species conservation strategies related to their wintering grounds.
Raptors are often considered good indicators of habitat quality because of their sensitivity to human disturbance and environmental contamination (Newton 1979; Sergio et al. 2005). For this group of birds, habitat transformation (i.e. agricultural intensification) can trigger two opposing effects on habitat-species relationships (Grande et al. 2018): (a) on the negative side, degradation and loss of suitable habitats, along with fewer trophic resources, because of agricultural intensification; (b) on the positive side, the provision of novel habitats and new food sources for some species (Villanúa et al. 2023).
Extensive farming areas represent a key ecosystem for birds wintering in Mediterranean regions (De Juana et al. 1988; Chiatante and Panuccio 2021). More recently, other authors have analysed the factors that determine the distribution and abundance of raptors wintering in different regions of Europe and North America (Kasprzykowski and Rzepala 2002; Jankowiak et al. 2015; Paprocki et al. 2015; Bobola et al. 2018). To date however, very little is known about the raptor communities that winter in the Mediterranean region (Panuccio et al. 2019). In this study, we analyse a raptor community that winters in a semiarid Mediterranean region in which interbasin water transfers have contributed to the proliferation of new intensive irrigation projects (Martínez and Esteve 2002). We relate the raptor wintering distribution to ecological features (i.e. land uses and topography). The aims of this study are to (i) model the spatial distribution of the raptor species that winter in a semiarid Mediterranean region, (ii) describe how environmental factors affect habitat suitability for each species, and (iii) identify the main ecological requirements of the studied species to conserve the wintering areas in the study area. We hypothesise that, although the studied species may have different habitat requirements (food availability) and, consequently, different potential distributions throughout the study area, agricultural areas may be more relevant than natural areas (i.e. forests) due to more food resources. These results will allow wildlife managers to conserve ecosystems that are considered to trophic resources for winter raptor biodiversity.
Materials and methods
Study area
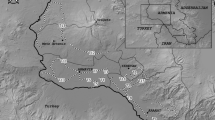
The present study was carried out between November 2017 and February 2019 in the Region of Murcia (Spain), an administrative unit located in the SE region of the Iberian Peninsula. It covers an area of 11,317 km2 (Fig. 1). Due to its geographical location and other physical factors, it is dominated by a semiarid Mediterranean climate. The annual mean temperature for the five-decade 1961–2014 period is 16.7 °C, which tends to increase by 0.135 °C per decade. The annual mean precipitation is 310 mm (Garrido et al. 2015).

Map of the study area (Region of Murcia, SE Spain) with the distribution of road surveys (red transects), including the land-use cover types used in the species distribution models. Coordinates are indicated as UTM 30S (metres)
The average monthly temperature values in winters vary from 12.2 °C in November to 8.5 °C in January. The monthly average precipitation in winter varies from 35 to 24 mm (AEMET 2011). The wide gradient between coastal and inland areas reflects the drop in temperature and the increase in precipitation when moving more inland in winter (AEMET 2011). In the coldest sector of the Region of Murcia (NW), there are relatively few days with snow, which has evolved from around 20 days in 1960 to about 10 days in 2014. This tendency is related to climate change (Garrido et al. 2015). Moreover, this geographical gradient from coastal areas to inland (from SE to NW) also shows a general pattern of a lowering turnover rate in forest bird species associated with climate and land-use change (Jiménez-Franco et al. 2024).
A complex orographic lies system in the study region, in which mountainous units alternate with flatter depressions to generate an altitudinal range from sea level to 2014 m asl. Combined with the wide climatic gradient, this contributes to generate a wide diversity of habitats that can be encompassed in three main ecosystem types: semidesert areas, Mediterranean scrub and coniferous forest (Esteve et al. 2015). From 1965 to 2000, the creation of 50,000 ha of human-modified habitat has significantly reduced the amount of natural area in this region (Calvo et al. 2017), where the agricultural landscape is dominated mainly by irrigated and dry land crops that total > 600,000 ha and represent more than half the land surface (Calvo et al. 2017).
Data collection
Bird occurrence data were collected for two winters from November 1 to February 28 (2017–2018 and 2018–2019) by two complementary methodologies: (a) professional fieldwork, based on road-based vehicle surveys conducted by trained professionals; (b) volunteers’ survey, based on bird record collection by means of volunteer collaborators. The simultaneous use of different methodologies is frequent in avian distribution studies (e.g. Moyer-Horner et al. 2012; Fletcher et al. 2019) because it avoids the detectability problems of certain less conspicuous species and can provide more records of more detailed data of all the species present (Zuberogoitia et al. 2020) that enhance the predictive accuracy of species distribution models (Tehrani et al. 2022). It is for this reason that this complementary methodology can be used successfully to build species distribution models (Broman et al. 2014; Flesch and Belt 2017).
Professional fieldwork. We surveyed five road transects during the 2018–2019 season (Fig. 1), each covering one of the agrarian districts making up the study region (Fernández 2012). Routes had an average length of 163 ± 11 km and an overall length of 817 km in the study area. Sampling journeys were repeated 3 times every ~40 days, and the total covered length was 2451 km. Following the usual methodological recommendations (e.g. Fuller and Mosher 1987; Sunyer and Viñuela 1996; Panuccio et al. 2019), routes were driven at low speed (20–40 km/h) to ensure increased likelihood of bird detection. Whenever a raptor was sighted, the car stopped long enough to allow field optics (binoculars 10×42 and telescope 20–60×) to be used for identification and data collection purposes. Routes were designed to cover the different landscape structures present in the study area, and to include, to the greatest possible extent, raptor activity hours by starting censuses at 2 h after sunrise at the most and finishing 1 h before sunset (Fuller and Mosher 1987; Bibby et al. 2000). Itineraries were not followed when adverse weather conditions (strong winds, fog or precipitation) affected censors’ visibility or bird mobility.
Volunteers’ survey scheme. Volunteers’ data collection was carried out throughout the study period (2017–2018 and 2018–2019 winter periods). A total of 87 cooperating birdwatchers were provided with a survey sheet and an application for smartphones developed specifically for this study (Teacher et al. 2013). Volunteers’ monitoring schemes have been proven suitable for collecting large quantities of data on birds (Sunyer and Viñuela 1990; Bonney et al. 2009; Teacher et al. 2013; Sullivan et al. 2014) at different spatial scales (Hernández-Navarro et al. 2023). We did not consider active citizen monitoring schemes (such as eBird) since the presence data provided was much less accurate for the spatial scale considered in our study. Therefore, to obtain the raptor presence data in the most precise way possible, a given record was considered correct when the collaborator completed all the required fields: date, time of day, location (UTM 30S coordinates), identified species, number of counted individuals and, optionally, any observations that were considered relevant to the sighting.
Environmental variables and processing
Ecological predictors were chosen according to previous descriptions of diurnal raptor communities (e.g. Bustamante and Seoane 2004; Jankowiak et al. 2015; Panuccio et al. 2019). Two different types of ecological variables to describe species-habitat relationships were selected (Seoane et al. 2004):
-
a)
Land uses. Corine Land Cover variables (Büttner and Kosztra 2017) were grouped into eight categories (CORINE Land Cover map 2018) (http://centrodedescargas.cnig.es; Fig. 1): urban environments (including human infrastructures and industrial areas), rainfed crops (extensive herbaceous agriculture), irrigated crops (intensive herbaceous agriculture), permanent crops (almonds, olive groves, vineyards, other arboreous crops), agro-forestry areas, forests, shrublands and wetlands. The frequency of each variable was estimated using a 1000-metre radius mobile-window to include species’ habitat requirements on the landscape scale (Abellán et al. 2011).
-
b)
Topographical. Two continuous quantitative variables were recorded: altitude, obtained from a digital elevation model map (DEM; http://centrodedescargas.cnig.es); the estimated distance to the coastline (km), due to its influence on climatic factors (Davies et al. 2007; Paprocki et al. 2014; Kawamura et al. 2019; Zhang et al. 2019). We took these variables as a proxy of climatic variables given the high correlation between them (Freeman et al. 2018). Moreover, the distance to the coastline in the study area showed a general pattern of a lowering turnover rate in forest bird species associated with climate and land-use change (Jiménez-Franco et al. 2024).
All these layers were used with a grid size of 100×100 m. To ensure a set of independent descriptors, linear correlation tests were performed to eliminate closely correlated variables (r > |0.70|; Zuur et al. 2009; Dormann et al. 2013).
Species distribution models
We modelled species distribution using the statistical approximation of maximum entropy implemented in MaxEnt (version 3.4.1; Phillips et al. 2006). This machine-learning technique is particularly appropriate for working with a presence-only dataset, and its predictive performance has been demonstrated to be more efficient than other presence-only and presence-absence methods, especially for relatively small sample sizes (Elith et al. 2011).
For modelling purposes, the observations obtained by professional fieldwork and volunteers’ monitoring scheme were used after eliminating those referring to the same date in the same 1×1 km cell to prevent possible duplication and oversampling problems in certain locations (Radosavljevic and Anderson 2014). The MaxEnt models were constructed for each species with a sample size composed of more than 15 observations to avoid the uncertainties related to an excessively small sample size (Pearson et al. 2007; Wisz et al. 2008). MaxEnt was run with default settings (regularisation multiplier b = 1; auto-features; convergence threshold = 0.00001) by employing the human footprint layer (HFP) as a bias grid (10,000 background points with a likelihood of presence proportional to HFP; Venter et al. 2016; Fandos and Tellería 2018; Tellería et al. 2019). This bias layer aims to correct the possible effect of areas with greater human presence on the sampling effort (Merow et al. 2013; Sullivan et al. 2014; Tellería et al. 2019). The model was replicated 30 times with 70% of the occurrences used as training data and the remaining 30% for model testing to assess variability in the model (Franklin and Miller 2010; Zeng et al. 2015). The percent contributions of jackknife tests were utilised to analyse the contribution rate and the importance of variables. Model discrimination performance was assessed by the mean area under the curve (AUC) of the receiver operating characteristics (ROC). The AUC is a threshold-independent measure of model evaluation that ranges from 0 to 1. An AUC value between 0.5 and 0.7 indicates poor model performance, while 0.7–0.9 denotes good model performance and > 0.9 very good performance (Swets 1988; Peterson et al. 2011). Only the models with AUC values > 0.75 were retained because these values are considered suitable for conservation planning (Zeng et al. 2015).
Habitat suitability maps were generated with the cloglog output (Phillips et al. 2017), where values range from a minimum probability value of 0 to a maximum value of 1. To facilitate the interpretation of suitability maps, they were transformed and represented using the QGIS software (version 3.4.4; Team QGIS Development 2018) with discrete 5-level categorisation based on the continuous models obtained with MaxEnt: 0–0.2, unsuitable; 0.2–0.4, low suitability; 0.4–0.6, moderate suitability; 0.6–0.8, high suitability; 0.8–1.0, very high suitability (Zhang et al. 2019).
Results
In this study, 2106 observations were recorded by considering both professional fieldwork and volunteers’ survey schemes, recording a total of 15 raptor species (Table 1). The species distribution models were constructed for the eight species (n = 1977 observations) that had a minimum number of records (n > 15) using all the environmental predictors of the starting set given the absence of correlation (Table S1). According to the values extracted from the evaluation of model performance for these eight modelled species (mean AUC, Table 1), prediction performance was generally high, and values were only poor (< 0.75) for the Falco peregrinus, whose predictions were not considered.
The environmental predictors that referred to elevation and distance to the coastline contributed the most to predict habitat suitability for most species by surpassing the land-use descriptors (Fig. S1). Elevation was the best predictor for Buteo buteo, Circus aeruginosus, Falco tinnunculus and Hieraaetus pennatus. In some cases, the high gain obtained in the jackknife test, such as for F. tinnunculus and H. pennatus, indicated that their distribution was strongly influenced by this factor. Distance to the coastline was also one of the main predictors of species distributions, but for none of the species was this factor the main explanatory variable.
Habitat type was the best predictor for Circus cyaneus and Falco columbarius (Fig. S1d, e). Of the anthropic land uses, rainfed crops made a notable contribution for C. cyaneus, irrigated crops for H. pennatus and F. tinnunculus, agro-forestry areas for F. columbarius and urban environments for A. nisus. With natural environments, wetlands were the most relevant predictor and featured among the main variables for C. aeruginosus, C. cyaneus and F. columbarius. Forests made a greater contribution as an explanatory variable for F. columbarius and F. tinnunculus and shrublands did so for C. cyaneus.
Increase in elevation led to a sharp drop in habitat suitability for all the species affected by this factor (Fig. S2). In contrast, distance from the coastline was associated with increased habitat suitability for intermediate distances. The optimum distance in this respect varied from around 40 km for H. pennatus, F. tinnunculus and C. aeruginosus to approximately 80 km for A. nisus and B. buteo.
The areas dominated by tree species, related to agro-forestry landscapes, decreased habitat suitability for F. columbarius and F. tinnunculus. Absence or a low proportion of shrublands, accompanied by a moderate proportion of rainfed crops and wetlands on the landscape scale, was optimal for C. cyaneus. A higher proportion of wetlands was preferred by C. aeruginosus, whose presence substantially increased when such habitats occupied around 60% of the landscape (Fig. S2d).
According to the suitability distribution maps and the proportion of highly suitable habitats (habitat suitability > 0.6) of the modelled raptor species (Fig. 2), it was possible to differentiate between two general distribution patterns for the (a) species that had a wide distribution pattern and a high proportion of suitable habitat, such as B. buteo, A. nisus or F. tinnunculus and (b) species with a more restricted distribution pattern, such as C. aeruginosus, C. cyaneus, F. columbarius or H. pennatus. For some species (B. buteo, F. columbarius, F. tinnunculus and H. pennatus), coastal areas had a high habitat suitability (Fig. 2).

Habitat suitability maps for seven raptor species wintering in the Region of Murcia (SE Spain)
Discussion
Our results showed that altitude and distance to the coastline were the best predictors to affect habitat suitability, probably due to their close association with climatic factors (Freeman et al. 2018). Altitude and climate are two closely related environmental factors that can play a determinant role in species distribution and the composition of local assemblages of birds and other taxa (Storch et al. 2003). Moreover, these variables may drive raptor species distributions on the macro-scale (Zhang et al. 2019). Altitude range does not impose limitations per se to the distribution patterns of wintering raptors but can strongly affect food availability (Zhang et al. 2019), which is often the main constraint for birds in winter in Mediterranean environments (Leal et al. 2019). Previous studies have found that higher altitudes increase environmental thermal stress and adversely affect animal thermoregulation by negatively influencing fitness and survival for wintering contingents. Food resources also decrease as altitude increases (Carrascal et al. 2002; 2012; Senar and Borras 2004). At the same time, the closer to the sea, the milder the winter temperature. This scenario favours winter residence and the survival of bird feeding on open lowlands, an environment in which many prey raptor species are especially abundant in winter (Herbers et al. 2004; Senar and Borras 2004). Therefore, in our case, altitude and distance to the coastline play an indirect role in determining the species distribution of local assemblages of raptors. This pattern has also been observed for forest species richness associated with heterogeneous areas of human-habitat influence, where low areas near the coast have high species richness and species turnover rates associated to the combined effects of climate and land-use changes for the last decades (Jiménez-Franco et al. 2019; 2024).
Altitude and distance to the coastline are also related to land uses in the study area. Forested areas tend to be situated at mid- and high-elevation mountain ranges and are mainly composed of pine (Pinus halepensis, Pinus pinaster and Pinus nigra). Forested landscapes, mostly situated at mid- and high elevation mountain ranges, are avoided by these species of wintering raptors probably due to the scarcity of their favoured foraging habitats (Carrascal and Palomino 2012). Other tree-dominated land cover types that are usually located at lower elevations, such as agro-forestry systems and permanent crops, also failed to contribute significantly towards habitat suitability for any of the modelled species. For this reason, it seems more likely that the presence of a dense tree cover influences foraging behaviour because of reduced prey detectability and accessibility (Bechard 1982; Butler and Gillings 2004; Ontiveros et al. 2005; Bobola et al. 2018).
Despite causing irreversible alterations to landscapes, urban environments can offer new abundant and stable food sources mainly for birds and particularly small- and medium-sized raptors (Donázar et al. 2016; Solaro 2018; Kettel et al. 2019), which can benefit from more hunting success and lower flight activity than those possible in non-urban settings (Rutz 2006). In our case, there was a negative relationship between species distribution and urbanised areas, except for A. nisus. It would seem that this species can successfully exploit urban habitats, a novel environment that may provide a valuable foraging habitat and high food availability for bird-eating raptors (McGrady 1991; Papp 2011; Jankowiak et al. 2015; Schütz and Schulze 2018).
Open habitats, such as wetlands and herbaceous crops, were important in the species distribution models, which suggests the importance of these natural and artificial habitat types in shaping raptor distribution. It is interesting to highlight the effect of wetlands on raptor distribution, which showed similar patterns for C. aeruginosus, C. cyaneus and F. columbarius. It is known that wetlands are critical habitats for the resting and feeding of many avian species in winter (Bobola et al. 2018). In recent decades, a matrix of fields used for intensive agriculture and urbanised areas now surround these wetlands in SE Spain, which generates a high diversity of natural and anthropogenic habitats (Sebastián-González et al. 2010). These represent favourable foraging grounds for certain raptors, such as F. columbarius, C. aeruginosus and C. cyaneus (Carrascal and Palomino 2012; Alves et al. 2014), by offering conditions that lead to the detection and capture of a variety of prey, including passerines, water birds, and small- and medium-sized mammals (Sunyer and Viñuela 1990; Clarke et al. 1993; Bobola et al. 2018).
Our models showed that the two predominant types of herbaceous crops (irrigated and rainfed) had the strongest influence on habitat suitability for raptors. This response is probably influenced by high food availability and the open structure of these environments that generate heterogeneous landscapes (Blount et al. 2021), which may offer increased food accessibility to open-space foragers (Leal et al. 2019). Hence, this pattern would explain why the distribution of the generalist B. buteo, a species that usually occupies open areas of sparse vegetation and scattered trees in winter, is optimal in landscapes made up of 80% irrigated crops (Wuczyński 2005; Baltag et al. 2013; Jankowiak et al. 2015) after having rapidly adapted to habitats that provide natural perches for hunting (Butet et al. 2010; Baltag et al. 2013). The case of H. pennatus is similar because its preference for irrigated crops also suggests that open-field agriculture offers many food resources, particularly when they are close to other habitat types like urban environments, forests, and especially wetlands (Palomino and Molina 2012; Vidal-Mateo and Urios 2017). In contrast to irrigated crops, whose presence seems to increase food availability for more generalist raptors (Cardador et al. 2012; Ferrer-Sánchez 2015; Grande et al. 2018), rainfed crops are used by specialist raptors like C. cyaneus (Carrascal and Palomino 2012; Clarke et al. 1993; Bobola et al. 2018). Therefore, in our case, the greater habitat heterogeneity generated by the closeness of herbaceous crops, wetlands and natural habitats may provide new foraging habitats to open-space raptors (Grande et al. 2018), as suggested by the greater complexity of the raptor communities observed in open spaces in winter (Panuccio et al. 2019). These results corroborate the importance of complex habitats to host biodiversity (Robledano et al. 2010), where it is necessary to enhance and value the multifunctionality of farms and landscapes, and to actively manage these landscapes for production, biodiversity and human well-being (Estrada-Carmona et al. 2022).
Previous studies have shown that traditional agricultural systems ensure a high biodiversity level (Pimentel et al. 1992). Nonetheless, the current model of agricultural intensification has resulted in generally declining bird populations that inhabit crop fields (Rahmann 2011; Bretagnolle et al. 2019). This negative effect of intensive agriculture on bird populations has been linked with loss of ecological heterogeneity and the lower quality of certain habitats (Guerrero et al. 2012). Several studies also suggest a negative impact of this process on raptor communities (Sánchez-Zapata et al. 2003; Carrete et al. 2009). However, as demonstrated herein and by previous studies (Cardador et al. 2011; Bobola et al. 2018), intensive crops and the agricultural infrastructure associated with them can provide new opportunities by, for example, providing new or more abundant trophic resources (Grande et al. 2018), which could be a determining factor for winter survival. Raptors might, therefore, select them to reduce interspecific competition and to facilitate co-existence with species that have similar trophic requirements (Sergio et al. 2007; 2008; Jankowiak et al. 2015; Paprocki et al. 2015; Bobola et al. 2018). In contrast, depending on the differences in each raptor species’ habitat preference, and in line with the conclusions of other studies (see Jankowiak et al. 2015; Bobola et al. 2018; Grande et al. 2018), we suggest that the expansion and intensification of agriculture to the detriment of natural land uses marginalise those species less suited to such changes which need more stable environments. For instance, it may affect the presence and abundance of steppe birds, particularly species of high conservation priority (De Frutos et al. 2015). This means that these habitat-dependent species would be replaced with species that are favoured by such transformations in their wintering grounds to further contribute to biotic homogenisation processes (Le Viol et al. 2012). Therefore, it is relevant to develop field works along years to study the population dynamic and the effects of dynamic landscapes on wintering raptors (Paprocki et al. 2015; McClure et al. 2021). Our results show the importance of agricultural areas as migration stopover sites, for which management actions should be set up to preserve high-quality habitats. Moreover, long-term studies are needed to quantify how land-use changes can affect the occurrence, diversity and abundance of species during the non-breeding period.
Conclusion
Overall, our study compiled information from two data sources (professional fieldwork and volunteers’ survey scheme) by using maximum entropy models to obtain the potential distribution and habitat association of wintering raptor species in a semiarid Mediterranean region. Our approach improves the efficiency of designing biological surveys when time and costs are limited (Robinson et al. 2020) because most survey efforts usually focus on raptors’ breeding period (Jiménez-Franco et al. 2020). Our results underline that some species have a wide distribution pattern, but others are distributed in specific areas that are mostly associated with wetlands where the food availability is high in winter periods. This study highlights the importance of the habitat heterogeneity generated by wetlands and herbaceous crops within a low-altitude range as the optimal environment for wintering raptors. We also consider that these results are useful for improving habitat management in the human-influenced portions of species’ wintering areas (Limiñana et al. 2015).
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Abellán MD, Martínez JE, Palazón JA, Esteve MA, Calvo JF (2011) Efficiency of a protected-area network in a mediterranean region: a multispecies assessment with raptors. Environ Manage 47:983–991. https://doi.org/10.1007/s00267-011-9640-5
AEMET (2011) Iberian Climate Atlas. Air temperature and precipitation (1971-2000). Ministerio de Medio Ambiente y Medio Rural y Marino e Instituto de Meteorologia de Portugal. Spain
Alves M, Ferreira JP, Torres I, Fonseca C, Matos M (2014) Habitat use and selection of the Marsh Harrier Circus aeruginosus in an agricultural-wetland mosaic. Ardeola 61: 351-366. https://doi.org/10.13157/arla.61.2.2014.351
Baltag ES, Pocora V, Sfîcǎ L, Bolboacǎ LE (2013) Common buzzard (Buteo buteo) population during winter season in North-Eastern Romania: the influences of density, habitat selection, and weather. Ornis Fenn 90:186–192
Bechard MJ (1982) Effect of vegetative cover on foraging site selection by Swainson’s hawk. Condor 84:153–159
Bibby CJ, Burgess ND, Hill DA (2000) Bird census techniques, 2nd edn. Academic Press, Oxford
Blount JD, Horns JJ, Kittelberger KD, Neate-Clegg MH, Sekercioğlu ÇH (2021) Avian use of agricultural areas as migration stopover sites: a review of crop management practices and ecological correlates. Front Ecol Evol 9:650641. https://doi.org/10.3389/fevo.2021.650641
Bobola E, Goutner V, Liordos V (2018) Foraging habitat selection and differentiation among coexisting raptors across an estuarine landscape (Evros Delta, northern Greece). Estuar Coast Shelf S 213:108–114. https://doi.org/10.1016/j.ecss.2018.08.009
Bonney R, Cooper CB, Dickinson J, Kelling S, Phillips T, Rosenberg KV, Shirk J (2009) Citizen Science: a developing tool for expanding science knowledge and scientific literacy. BioScience 59:977–984. https://doi.org/10.1525/bio.2009.59.11.9
Bretagnolle V, Siriwardena G, Miguet P, Henckel L, Kleijn D (2019) Local and landscape scale effects of heterogeneity in shaping bird communities and population dynamics: crop-grassland interactions. In: Lemaire G, Carvalho P, Kronberg S, Recous S (Eds.) Agroecosystem Diversity: 231–243. Academic Press, Cambridge, USA
Broman DJA, Litvaitis JA, Ellingwood M, Tate P, Reed GC (2014) Modeling bobcat (Lynx rufus) habitat associations using telemetry locations and citizen-scientist observations: are the results comparable? Wildlife Biol 20:229–237. https://doi.org/10.2981/wlb.00022
Bustamante J, Seoane J (2004) Predicting the distribution of four species of raptors (Aves: Accipitridae) in southern Spain: statistical models work better than existing maps. J Biogeogr 31: 295-306. https://www.jstor.org/stable/3554658
Butet A, Michel N, Rantier Y, Comor V, Hubert-Moy L, Nabucet J, Delettre Y (2010) Responses of common buzzard (Buteo buteo) and Eurasian kestrel (Falco tinnunculus) to land use changes in agricultural landscapes of Western France. Agr Ecosyst Environ 138:152–159. https://doi.org/10.1016/j.agee.2010.04.011
Butler SJ, Gillings S (2004) Quantifying the effects of habitat structure on prey detectability and accessibility to farmland birds. Ibis 146:123–130. https://doi.org/10.1111/j.1474-919X.2004.00352.x
Büttner G, Kosztra B (2017) CLC2018 technical guidelines. European Environment Agency, Wien, Austria
Calvo JF, Hernández-Navarro AJ, Robledano F, Esteve MA, Ballesteros G, Fuentes A et al (2017) Catálogo de las aves de la Región de Murcia (España). Anales Biol 39:7–33. https://doi.org/10.6018/analesbio.39.02
Caraveli H (2000) A comparative analysis on intensification and extensification in Mediterranean agriculture: dilemmas for LFAs policy. J Rural Stud 16:231–242. https://doi.org/10.1016/S0743-0167(99)00050-9
Cardador L, Carrete M, Mañosa S (2011) Can intensive agricultural landscapes favour some raptor species? The Marsh harrier in north-eastern Spain. Anim Conserv 14:382–390. https://doi.org/10.1111/j.1469-1795.2011.00449.x
Cardador L, Planas E, Varea A, Mañosa S (2012) Feeding behaviour and diet composition of marsh harriers Circus aeruginosus in agricultural landscapes. Bird Study 59:228–235. https://doi.org/10.1080/00063657.2011.648165
Carrascal LM, Palomino D (2012) Variación geográfica de la riqueza de especies invernantes en la península Ibérica. Estacionalidad y determinismo ambiental. In: SEO/BirdLife (Ed.) Atlas de las aves en invierno en España 2007-2010: 36-47. Ministerio de Agricultura, Alimentación y Medio Ambiente-SEO/BirdLife. Madrid
Carrascal LM, Palomino D, Lobo JM (2002) Patrones de preferencias de hábitat y de distribución y abundancia invernal de aves en el centro de España. Análisis y predicción del efecto de factores ecológicos. Anim Biodiv Conserv 25: 7-40. https://raco.cat/index.php/ABC/article/view/57559
Carrascal LM, Seoane J, Villén-Pérez S (2012) Temperature and food constraints in wintering birds - an experimental approach in montane Mediterranean oakwoods. Comm Ecol 13: 221-229. https://www.jstor.org/stable/24113700
Carrete M, Tella JL, Blanco G, Bertellotti M (2009) Effects of habitat degradation on the abundance, richness and diversity of raptors across Neotropical biomes. Biol Conserv 142:2002–2011. https://doi.org/10.1016/j.biocon.2009.02.012
Chapin FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL et al (2000) Consequences of changing biodiversity. Nature 405:234–242. https://doi.org/10.1038/35012241
Chiatante G, Panuccio M (2021) Environmental factors affecting the wintering raptor community in Armenia, Southern Caucasus. Comm Ecol 22:79–92. https://doi.org/10.1007/s42974-021-00038-7
Clarke R, Bourgonje A, Castelinjns H (1993) Food niches of sympatric marsh harriers Circus aeruginosus and hen harriers C. cyaneus on the Dutch coast in winter. Ibis 135:424–431. https://doi.org/10.1111/j.1474-919X.1993.tb02115.x
Davies RG, Orme CDL, Storch D, Olson VA, Thomas GH, Ross SG et al (2007) Topography, energy and the global distribution of bird species richness. Proc R Soc Lond B Biol Sci 274:1189–1197. https://doi.org/10.1098/rspb.2006.0061
De Frutos A, Olea PP, Mateo-Tomás P (2015) Responses of medium-and large-sized bird diversity to irrigation in dry cereal agroecosystems across spatial scales. Agr Ecosyst Environ 207:141–152. https://doi.org/10.1016/j.agee.2015.04.009
De Juana E, De Juana F, Calvo S (1988) La invernada de las aves de presa (O. Falconiformes) en la Península Ibérica. In: Tellería JL (Ed.) Invernada de aves en la Península Ibérica: 97-122. Monografías SEO, 1. Sociedad Española de Ornitología. Madrid
Donázar JA, Cortés-Avizanda A, Fargallo JA, Margalida A, Moleón M, Morales-Reyes Z et al (2016) Roles of raptors in a changing world: from flagships to providers of key ecosystem services. Ardeola 63: 181-234. https://doi.org/10.13157/arla.63.1.2016.rp8
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G et al (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:027–046. https://doi.org/10.1111/j.1600-0587.2012.07348.x
Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x
Esteve MA, Martínez J, Hernández I, Robledano F, Pérez MA, Lloret F (2015) Cambio climático y biodiversidad en el contexto de la Región de Murcia. In: Garrido R, Palenzuela JE, Bañón LM, García JA (Ed.) Cambio climático en la Región de Murcia. Evaluación basada en indicadores: 105-132. Consejería de Agua, Agricultura y Medio Ambiente, Comunidad Autónoma de la Región de Murcia
Estrada-Carmona N, Sánchez AC, Remans R, Jones SK (2022) Complex agricultural landscapes host more biodiversity than simple ones: a global meta-analysis. PNAS 119(38):e2203385119. https://doi.org/10.1073/pnas.2203385119
Fandos G, Tellería JL (2018) Range compression of migratory passerines in wintering grounds of the Western Mediterranean: conservation prospects. Bird Conserv Int 28:462–474. https://doi.org/10.1017/S0959270917000120
Fernández J (2012) Caracterización de las Comarcas Agrarias de España. Ministerio de Agricultura, Alimentación y Medio Ambiente
Ferrer-Sánchez Y (2015) Variables que influyen en la distribución y abundancia de rapaces diurnas y en la ubicación de sus sitios de anidación en Cuba. Tesis Doctoral, Centro de Investigaciones Biológicas del Noroeste, S.C. Mexico
Flesch EP, Belt JJ (2017) Comparing citizen science and professional data to evaluate extrapolated mountain goat distribution models. Ecosphere 8(2):e01638. https://doi.org/10.1002/ecs2.1638
Fletcher RJ Jr, Hefley TJ, Robertson EP, Zuckerberg B, McCleery RA, Dorazio RM (2019) A practical guide for combining data to model species distributions. Ecology 100(6):e02710. https://doi.org/10.1002/ecy.2710
Foley JA, Foley JA, DeFries R, Asner GP, Barford C, Bonan G et al (2005) Global consequences of land use. Science 309:570–574. https://doi.org/10.1126/science.1111772
Fox AD (2004) Has Danish agriculture maintained farmland bird populations? J Appl Ecol 41: 427– 439. https://www.jstor.org/stable/3505819
Franklin J, Miller JA (2010) Mapping species distributions: spatial inference and prediction. Cambridge University Press, Cambridge
Freeman BG, Scholer MN, Ruiz-Gutierrez V, Fitzpatrick JW (2018) Climate change causes upslope shifts and mountaintop extirpations in a tropical bird community. PNAS 115(47):11982–11987. https://doi.org/10.1073/pnas.1804224115
Fuller MR, Mosher JA (1987) Raptor survey techniques. In: Geron B, Milleap B, Cline K, Bird D. (Eds.) Raptor management techniques manual: 37-65. National Wildlife Federation, Washington D.C
Garrido R, Palenzuela JE, Bañón LM, García JA (2015) Cambio climático en la Región de Murcia. Evaluación basada en indicadores. Consejería de Agua, Agricultura y Medio Ambiente, Comunidad Autónoma de la Región de Murcia
Grande JM, Orozco-Valor PM, Liébana MS, Sarasola JH (2018) Birds of prey in agricultural landscapes: the role of agriculture expansion and intensification. In: Sarasola JH, Grande JM, Negro JJ (Eds.) Birds of prey: biology and conservation in the XXI century: 197-228. Springer International Publishing, Cham
Guerrero I, Morales MB, Oñate JJ, Geiger F, Berendse F, De Snoo G et al (2012) Response of ground-nesting farmland birds to agricultural intensification across Europe: landscape and field level management factors. Biol Conserv 152:74–80. https://doi.org/10.1016/j.biocon.2012.04.001
Herbers J, Serrouya R, Maxcy K (2004) Effects of elevation and forest cover on winter birds in mature forest ecosystems of southern British Columbia. Can J Zool 82:1720–1730. https://doi.org/10.1139/z04-151
Hernández-Navarro AJ, Robledano F, Jiménez-Franco MV et al (2023) Long-term trends of local bird populations based on monitoring schemes: are they suitable for justifying management measures? J Ornithol. https://doi.org/10.1007/s10336-023-02114-3
Jankowiak L, Antczak M, Kwieciñski Z, Szymañski P, Tobolka M, Tryjanowski P (2015) Diurnal raptor community wintering in an extensively used farmland. Ornis Fenn 92:76–86
Jiménez-Franco MV, Kéry M, León-Ortega M, Ródenas J, Robledano F, Esteve MA, Calvo JF (2024) Evaluating temporal turnover in avian species richness in a Mediterranean semiarid region: different responses to elevation and forest cover. Div Distrib 30(1):106–118. https://doi.org/10.1111/ddi.13791
Jiménez-Franco MV, Kéry M, León-Ortega M, Robledano F, Esteve MA, Calvo JF (2019) Use of classical bird census transects as spatial replicates for hierarchical modeling of an avian community. Ecol Evol 9:825–835. https://doi.org/10.1002/ece3.4829
Jiménez-Franco MV, Martínez JE, Pagán I, Calvo JF (2020) Long-term population monitoring of a territorial forest raptor species. Sci Data 7:166. https://doi.org/10.1038/s41597-020-0503-x
Jiménez-Franco MV, Graciá E, Rodríguez-Caro RC et al (2022) Problems seeded in the past: lagged effects of historical land-use changes can cause an extinction debt in long-lived species due to movement limitation. Landsc Ecol 37:1331–1346. https://doi.org/10.1007/s10980-021-01388-3
Jetz W, Wilcove DS, Dobson AP (2007) Projected impacts of climate and land-use change on the global diversity of birds. PLoS Biol 5:1211–1219. https://doi.org/10.1371/journal.pbio.0050157
Kasprzykowski Z, Rzepala M (2002) Numbers and habitat preferences of raptors Falconiformes wintering in central-eastern Poland. Not Ornitol 43:73–82. https://doi.org/10.3161/068.040.0210
Kawamura K, Yamaura Y, Senzaki M, Ueta M, Nakamura F (2019) Seasonality in spatial distribution: climate and land use have contrasting effects on the species richness of breeding and wintering birds. Ecol Evol 9:7549–7561. https://doi.org/10.5061/dryad.7n404q0
Kettel EF, Gentle LK, Yarnell RW, Quinn JL (2019) Breeding performance of an apex predator, the peregrine falcon, across urban and rural landscapes. Urban Ecosyst 22:117–125. https://doi.org/10.1007/s11252-018-0799-x
Landis DA (2017) Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl Ecol 18:1–12. https://doi.org/10.1016/j.baae.2016.07.005
La Sorte FA, Fink D, Blancher PJ, Rodewald AD, Ruiz-Gutierrez V, Rosenberg KV et al (2017) Global change and the distributional dynamics of migratory bird populations wintering in Central America. Global Change Biol 23:5284–5296. https://doi.org/10.1111/gcb.13794
Leal AI, Acácio M, Meyer CFJ, Rainho A, Palmeirim JM (2019) Grazing improves habitat suitability for many ground foraging birds in Mediterranean wooded grasslands. Agr Ecosyst Environ 270–271:1–8. https://doi.org/10.1016/j.agee.2018.10.012
Limiñana R, Arroyo B, Terraube J, McGrady M, Mougeot F (2015) Using satellite telemetry and environmental niche modelling to inform conservation targets for a long-distance migratory raptor in its wintering grounds. Oryx 49:329–337. https://doi.org/10.1017/S0030605313001075
Le Viol I, Jiguet F, Brotons L, Herrando S, Lindström Å, Pearce-Higgins JW et al (2012) More and more generalists: two decades of changes in the European avifauna. Biol Lett 8:780–782. https://doi.org/10.1098/rsbl.2012.0496
Martínez J, Esteve MA (2002) Agua, regadío y sostenibilidad en el sudeste ibérico. Ed. Bakeaz, Bilbao
McClure CJ, Rolek BW, Grove GW, Katzner TE (2021) Yearly temperature fluctuations and survey speed influence road counts of wintering raptors. Ibis 163(2):593–606. https://doi.org/10.1111/ibi.12914
McGrady MJ (1991) The ecology and breeding behaviours of urban sparrowhawks (Accipiter nisus) in Edinburgh, Scotland. PhD Thesis, University of Edinburgh
Merow C, Smith MJ, Silander JA (2013) A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography 36:1058–1069. https://doi.org/10.1111/j.1600-0587.2013.07872.x
Meyer WB, Turner BL (1992) Human population growth and global land-use/cover change. Annu Rev Ecol Evol S 23: 39-61. https://www.jstor.org/stable/2097281
Moyer-Horner L, Smith MM, Belt J (2012) Citizen science and observer variability during American pika surveys. J Wildlife Manage 76:1472–1479. https://doi.org/10.1002/jwmg.373
Newbold T, Hudson LN, Hill SLL, Contu S, Lysenko I, Senior RA et al (2015) Global effects of land use on local terrestrial biodiversity. Nature 520:45–50. https://doi.org/10.1038/nature14324
Newton I (1979) Population ecology of raptors. T & AD Poyser. London, UK
Ontiveros D, Pleguezuelos JM, Caro J (2005) Prey density, prey detectability and food habits: the case of Bonelli’s eagle and the conservation measures. Biol Conserv 123:19–25. https://doi.org/10.1016/j.biocon.2004.10.004
Palomino D, Molina B (2012) Aguila Calzada (Aquila pennata). In SEO/BirdLife (Ed.) Atlas de las aves en invierno en España 2007-2010: 188-189. Ministerio de Agricultura, Alimentación y Medio Ambiente-SEO/BirdLife. Madrid
Panuccio M, Agostini N, Nelli L, Andreou G, Xirouchachis S (2019) Factors shaping distribution and abundance of raptors wintering in two large Mediterranean islands. Community Ecol 20:93–103. https://doi.org/10.1556/168.2019.20.1.10
Papp S (2011) Breeding of Eurasian sparrowhawks (Accipiter nisus) in two Hungarian towns. Aquila 118:49–54
Paprocki N, Glenn NF, Atkinson EC, Strickler KM, Watson C, Heath JA (2015) Changing habitat use associated with distributional shifts of wintering raptors. J Wildlife Manage 79:402–412. https://doi.org/10.1002/jwmg.848
Paprocki N, Heath JA, Novak SJ (2014) Regional distribution shifts help explain local changes in wintering raptor abundance: implications for interpreting population trends. PLoS ONE 9:e86814. https://doi.org/10.1371/journal.pone.0086814
Pearson RG, Raxworthy CJ, Nakamura M, Townsend Peterson A (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117. https://doi.org/10.1111/j.1365-2699.2006.01594.x
Peterson A, Soberón J, Pearson R, Anderson RP, Martínez-Meyer E, Nakamura M, Araújo MB (2011) Ecological niches and geographic distributions. Princeton University Press, Princeton, NJ
Phillips SJ, Anderson RP, Dudík M, Schapire RE, Blair ME (2017) Opening the black box: an open-source release of Maxent. Ecography 40:887–893. https://doi.org/10.1111/ecog.03049
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Modell 190:231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Pimentel D, Stachow U, Takacs DA, Brubaker HW, Amy R, Meaney JJ et al (1992) Conserving biological diversity in agricultural/forestry systems. BioScience 42:354–362. https://doi.org/10.2307/1311782
Powers RP, Jetz W (2019) Global habitat loss and extinction risk of terrestrial vertebrates under future land-use-change scenarios. Nat Clim Chang 9:323–329. https://doi.org/10.1038/s41558-019-0406-z
Radosavljevic A, Anderson RP (2014) Making better Maxent models of species distributions: complexity, overfitting and evaluation. J Biogeogr 41:629–643. https://doi.org/10.1111/jbi.12227
Rahmann G (2011) Biodiversity and organic farming: what do we know? Landbauforsch Volk 61:189–208
Robinson OJ, Ruiz-Gutierrez V, Reynolds MD, Golet GH, Strimas-Mackey M, Fink D (2020) Integrating citizen science data with expert surveys increases accuracy and spatial extent of species distribution models. Divers Distrib 26:976–986. https://doi.org/10.1111/ddi.13068
Robledano F, Esteve MA, Farinós P, Carreño MF, Martínez-Fernández J (2010) Terrestrial birds as indicators of agricultural-induced changes and associated loss in conservation value of Mediterranean wetlands. Ecol Indic 10:274–286. https://doi.org/10.1016/j.ecolind.2009.05.006
Rutz C (2006) Home range size, habitat use, activity patterns and hunting behaviour of urban-breeding northern goshawks Accipiter gentilis. Ardea 94:185–202
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R et al (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774. https://doi.org/10.1126/science.287.5459.1770
Sánchez-Zapata JA, Carrete M, Gravilov A, Sklyarenko S, Ceballos O, Donázar JA, Hiraldo F (2003) Land use changes and raptor conservation in steppe habitats of Eastern Kazakhstan. Biol Conserv 111:71–77. https://doi.org/10.1016/S0006-3207(02)00251-3
Schütz C, Schulze CH (2018) Park size and prey density limit occurrence of Eurasian sparrowhawks in urban parks during winter. Avian Res 9:30. https://doi.org/10.1186/s40657-018-0122-9
Sebastián-González E, Sánchez-Zapata JA, Botella F (2010) Agricultural ponds as alternative habitat for waterbirds: spatial and temporal patterns of abundance and management strategies. Eur J Wildl Res 56:11–20. https://doi.org/10.1007/s10344-009-0288-x
Senar JC, Borras A (2004) Surviving to winter: strategies of wintering birds in the Iberian Peninsula. Ardeola 51:133–168
Seoane J, Bustamante J, Díaz-Delgado R (2004) Competing roles for landscape, vegetation, topography and climate in predictive models of bird distribution. Ecol Modell 171:209–222. https://doi.org/10.1016/j.ecolmodel.2003.08.006
Sergio F, Caro T, Brown D, Clucas B, Hunter J, Ketchum J et al (2008) Top predators as conservation tools: ecological rationale, assumptions, and efficacy. Annu Rev Ecol Evol S 39:1–19. https://doi.org/10.1146/annurev.ecolsys.39.110707.173545
Sergio F, Marchesi L, Pedrini P, Penteriani V (2007) Coexistence of a generalist owl with its intraguild predator: distance-sensitive or habitat-mediated avoidance? Anim Behav 74:1607–1616. https://doi.org/10.1016/j.anbehav.2006.10.022
Sergio F, Newton I, Marchesi L (2005) Conservation: top predators and biodiversity. Nature 436:192. https://doi.org/10.1038/436192a
Sirami C, Gross N, Baillod AB, Bertrand C, Carrié R, Hass A et al (2019) Increasing crop heterogeneity enhances multitrophic diversity across agricultural regions. Proc Natl Acad Sci 116:16442–16447. https://doi.org/10.1073/pnas.1906419116
Solaro C (2018) Costs and benefits of urban living in raptors. In: Sarasola JH, Grande JM, Negro JJ (Eds.) Birds of prey: biology and conservation in the XXI century: 177-196. Springer International Publishing, Cham
Stoate C, Boatman ND, Borralho RJ, Carvalho CR, De Snoo GR, Eden P (2001) Ecological impacts of arable intensification in Europe. J Environ Manage 63:337–365. https://doi.org/10.1006/jema.2001.0473
Storch D, Konvicka M, Benes J, Martinková J, Kevin J, Gaston KJ (2003) Distribution patterns in butterflies and birds of the Czech Republic: separating effects of habitat and geographical position. J Biogeogr 30:1195–1205. https://doi.org/10.1046/j.1365-2699.2003.00917.x
Sullivan BL, Aycrigg JL, Barry JH, Bonney RE, Bruns N, Cooper CB et al (2014) The eBird enterprise: an integrated approach to development and application of citizen science. Biol Conserv 169:31–40. https://doi.org/10.1016/j.biocon.2013.11.003
Sunyer C, Viñuela J (1990) Migración e invernada del Esmerejón en España. Ardeola 279-290
Sunyer C, Viñuela J (1996) Invernada de rapaces (O. Falconiformes) en España peninsular e Islas Baleares. In: Muntaner J & Mayol J (Eds.) Biología y Conservación de las Rapaces Mediterráneas, 1994: 361-370. Monografías nº 4, SEO. Madrid
Swets JA (1988) Measuring the accuracy of diagnostic systems. Science 240:1285–1293. https://doi.org/10.1126/science.3287615
Teacher AGF, Griffiths DJ, Hodgson DJ, Inger R (2013) Smartphones in ecology and evolution: a guide for the app-rehensive. Ecol Evol 3:5268–5278. https://doi.org/10.1002/ece3.888
Team QGIS Development (2018) QGIS Geographic Information System. Open source geospatial Foundation project. https://qgis.org/es/site/
Tehrani NA, Naimi B, Jaboyedoff M (2022) A data-integration approach to correct sampling bias in species distribution models using multiple datasets of breeding birds in the Swiss Alps. Ecol Inform 69:101501. https://doi.org/10.1016/j.ecoinf.2021.101501
Tellería JL, Fandos G, Tena E, Carbonell R, Onrubia A, Qninba A, Ramírez Á (2019) Constraints on raptor distribution at the southwestern boundary of the Palaearctic: implications for conservation. Biodivers Conserv 28:603–619. https://doi.org/10.1007/s10531-018-1677-9
Tellería JL, Fernández-López J, Fandos G (2016) Effect of climate change on Mediterranean winter ranges of two migratory passerines. PLoS ONE 11:e0146958. https://doi.org/10.1371/journal.pone.0146958
Valera F, Díaz-Paniagua C, Garrido-García JA, Manrique J, Pleguezuelos JM, Suárez F (2011) History and adaptation stories of the vertebrate fauna of southern Spain’s semi-arid habitats. J Arid Environ 75:1342–1351. https://doi.org/10.1016/j.jaridenv.2011.05.004
Venter O, Sanderson EW, Magrach A, Allan JR, Beher J, Jones KR et al (2016) Global terrestrial human footprint maps for 1993 and 2009. Sci Data 3:160067. https://doi.org/10.1038/sdata.2016.67
Vickery PD, Tubaro PL, Da Silva M, Peterjohn B, Herkert J, Cavalcanti RB (1999) Conservation of grassland birds in the western hemisphere. Stud Avian Biol 19:2–26
Vidal-Mateo J, Urios V (2017) Ecología espacial en el período invernal. In: Urios V, Bermejo A, Vidal-Mateo J, De la Puente J (Eds.) Migración y ecología espacial de la población española de águila calzada. Monografía n.o 2 del programa Migra. SEO/BirdLife. Madrid
Villanúa D, Cabodevilla X, Ardaiz J, Lizarraga A, Zufiaurre A (2023) Effect of implementation of irrigation on raptor and corvid populations in a Mediterranean agrosystem. Anim Biod Conserv 46(2): 155-163. https://doi.org/10.32800/abc.2023.46.0155
Visser ME, Perdeck AC, van Balen JH, Both C (2009) Climate change leads to decreasing bird migration distances. Global Change Biol 15:1859–1865. https://doi.org/10.1111/j.1365-2486.2009.01865.x
Wisz MS, Hijmans RJ, Li J, Peterson AT, Graham CH, Guisan A et al (2008) Effects of sample size on the performance of species distribution models. Divers Distrib 14:763–773. https://doi.org/10.1111/j.1472-4642.2008.00482.x
Wuczyński A (2005) Habitat use and hunting behaviour of common buzzards Buteo buteo wintering in south-western Poland. Acta Ornithol 40:147–154. https://doi.org/10.3161/068.040.0210
Zeng Q, Zhang Y, Sun G, Duo H, Wen L, Lei G (2015) Using species distribution model to estimate the wintering population size of the endangered scaly-sided merganser in China. PLoS ONE 10:e0117307. https://doi.org/10.1371/journal.pone.0117307
Zhang J, Jiang F, Li G, Qin W, Li S, Gao H, Cai Z, Lin G, Zhang T (2019) Maxent modeling for predicting the spatial distribution of three raptors in the Sanjiangyuan National Park, China. Ecol Evol 9:6643–6654. https://doi.org/10.1002/ece3.5243
Zuberogoitia I, Martínez JE, González-Oreja JA, de Buitrago CG, Belamendia G, Zabala J et al (2020) Maximizing detection probability for effective large-scale nocturnal bird monitoring. Divers Distrib 26(8):1034–1050. https://doi.org/10.1111/ddi.13075
Zuur AF, Ieno EN, Elphick CS (2009) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14. https://doi.org/10.1111/j.2041-210X.2009.00001.x
Acknowledgements
The authors thank all the participants of the volunteers’ survey scheme who volunteered with their time to collect data, including 87 birdwatchers who contributed to this study. We are also grateful to Francisco Montoya and Eduardo Casabella for providing help with application for smartphones developed specifically for this study.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. M.V.J.-F. is supported by a “Juan de la Cierva-Incorporación” research contract from the Spanish Ministry of Economy and Competitiveness (reference IJC2019-039145-I).
Author information
Authors and Affiliations
Contributions
MA and JFC conceived the ideas and designed the methodology; MA, JEM and MLO collected the data; MA, MVJF and JFC analysed the data; MA and MVJF led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Ethical approval
This is not applicable. This is an observational study.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Álvarez, M., León-Ortega, M., Martínez, J.E. et al. Wintering raptor species distribution in a semiarid Mediterranean region: the relevance of lowlands and open habitats as stopover sites. Eur J Wildl Res 70, 30 (2024). https://doi.org/10.1007/s10344-024-01784-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-024-01784-5