
Abstract
Most European ungulate species are increasing in numbers and expanding their range. For the management and monitoring of these species, 64% of European countries rely on indirect proxies of abundance (e.g., hunting bag statistics). With increasing ungulate numbers, data on ungulate-vehicle collisions (UVC) may provide an important and inexpensive, complementary data source. Currently, it is unclear how bag statistics compare with UVC. A direct comparison of these two indices is important because both are used in ungulate management. We evaluated the relationship between UVC and ungulate hunting bags across bioclimatic, regional, and local scales, using five time lags (t−3 to t+1) for the five most common wild ungulate species in Sweden. For all species, hunting bags and UVC correlated positively, but correlation strength and time lags varied across scales and among species. The two indices correlated most strongly at the local management scale. Correlation between both indices was strong for the smaller deer species and wild boar, in particular, but much weaker for moose where we found the best fit using a 2-year time lag. For the other species, indices from the same year correlated best. We argue that the reason for moose data behaving differently is that, in Sweden, moose are formally managed using a 3-year time plan, while the other species are not. Accordingly, moose hunting bags are influenced more strongly by density-independent processes than bags of the other species. Consequently, the mismatch between the two indices may generate conflicting conclusions for management depending on the method applied.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Europe is experiencing strong increases in the number of ungulates, and several species are expanding their ranges (Apollonio et al. 2010). These changes are leading to new challenges for European ungulate management (Putman et al. 2011; Linnell et al. 2020). Monitoring these changes at different management scales is essential for adaptive ungulate management. Different monitoring methods have different costs and benefits (Månsson et al. 2011; Putman et al. 2011; Brazeal et al. 2017). Characteristics such as biology, movements, and social behavior that may vary among species can also affect the performance of different methods (Jarnemo et al. 2017; Massei et al. 2018). While specific methods can successfully monitor populations of established species (e.g., the system of moose (Alces alces) observations by hunters in Sweden, Månsson et al. 2011), it may be challenging to apply the same methods to accurately monitor expanding species (e.g., wild boar (Sus scrofa), red deer (Cervus elaphus), Pfeffer et al. 2017). To monitor the ongoing changes in Europe’s ungulate communities (Linnell et al. 2020), we therefore need to find methods that are easily accessible, inexpensive, and applicable to multiple species and from local to national scales.
Today, in addition to direct observations, the majority of European countries (64 %) uses hunting bag statistics as a proxy for assessing relative changes in the distribution and abundance of ungulates at national scales (Apollonio et al. 2010; Putman et al. 2011). Following the need of cross-border management (Linnell et al. 2020), recent initiatives develop abundance models using hunting statistics at the European scale (ENETWILD-consortium et al. 2019). Hunting bags, however, may not be reliable if the hunting effort of a species is not related to its density and when this relationship changes over time (Imperio et al. 2010; Willebrand et al. 2011; Ferretti et al. 2016; Eriksen et al. 2018). This may be particularly true for expanding species that occur at low densities or species that are not (yet) part of the mainstream monitoring and ungulate management. For those species, ungulate-vehicle collisions (UVC) data might provide a useful complementary monitoring method. UVC are increasing in many countries (Apollonio et al. 2010; Seiler et al. 2016; Linnell et al. 2020), and accident data is typically stored in national traffic safety databases or as hunter reports (e.g., Germany: Hothorn et al. 2015; Spain: Rosell et al. 2013; Sweden: Seiler et al. 2019). Yet, a growing number of countries also involve special reporting software and websites for a more efficient and accurate registration of animal-vehicle accidents (Heigl et al. 2016; Bíl et al. 2017; Waetjen and Shilling 2017). The timestamp and geographic coordinates of UVC provide detailed information on where and when accidents occur and thus animal distribution in time and space. While countries collect this data for managing UVC (i.e., increasing traffic safety), this data can serve as a standardized, low-cost ungulate population monitoring method, similar to bag statistics.
Previous work suggests that UVC closely correlate with other proxies of population abundance (e.g., observations, hunting bags) for some species in certain systems (e.g., moose in Fennoscandia, Seiler 2004; Rolandsen et al. 2011; Ueno et al. 2014; Niemi et al. 2017; roe deer in Germany (Capreolus capreolus), Hothorn et al. 2012; and wild boar in France (Saint-Andrieux et al. 2020)). However, these studies also highlight that density-independent factors and not necessarily ungulate densities drive the dynamics inherent in UVC (Seiler and Helldin 2006; Hothorn et al. 2015). More specifically, density-independent factors that influence UVC dynamics include annual and seasonal variation in ungulate activity patterns (see for roe deer, Hothorn et al. 2015, Neumann et al. 2019; for red deer, Mysterud 2004; for moose, Neumann et al. 2012, 2019) and anthropogenic drivers such as road density and type, human density, wildlife fences, and landscape fragmentation (Gunson et al. 2009; Rolandsen et al. 2011; Saint-Andrieux et al. 2020). As such, the spatiotemporal distribution of UVC may not relate linearly to ungulate numbers, and this may vary among ungulate species and environmental regions. Finally, most available studies have focused on national to county scale, or case studies (e.g., Seiler et al. 2016), and on single ungulate species. We lack studies that looked at UVC dynamics at the relevant local scale of ungulate management and studies that assessed UVC dynamics for multiple co-occurring species simultaneously (although see Saint-Andrieux et al. (2020), who compare UVC dynamics among roe deer, red deer, and wild boar in France).
Although previous work suggests that changes in bag statistics and UVC, respectively, may reflect changes in population densities, we lack assessments of how these two indices relate to each other for the diverse European ungulate species (but see Saint-Andrieux et al. 2020). Such a direct comparison is highly relevant, because different interest groups in ungulate management may use either hunting bags or UVC to argue for given management measures (Neumann et al., personal observations). If these indices do not align and provide different proxies for ungulate abundance, this may generate conflicts among the different groups within ungulate management. A better understanding of the relationship between these two data sets for different ungulate species is thus vital. Strong correlations across time and space would suggest that both methods reveal similar trends in ungulate population abundance, whereas a weak correlation would suggest the opposite. Here, we deliberately use the term trends, since even a strong correlation between the two methods may not necessarily mean that the methods reflect absolute population density very well. Ungulate management, however, rarely has access to data on absolute density and heavily depends on indirect proxies such as from bag statistics and UVC.
Here, we investigated three key questions: (1) Do hunting bags and UVC data correlate positively and significantly for different co-occurring ungulate species across time and space? (2) Does the strength of the correlation between hunting bags and UVC differ across different spatial scales that are relevant for management i.e., bioclimatic, regional (county), and local (management) scale? and (3) Does the strength of the correlation between hunting bags and UVC differ among ungulate species? We investigated these questions for five European ungulate species (i.e., moose, roe deer, red deer, fallow deer (Dama dama), and wild boar) using nationwide data from Sweden. We also tested how relationships varied for different time lags (t−3 to t+1) between UVC data and hunting statistics.
Materials and methods
Study area
Our study area encompasses the whole of Sweden, except the island of Gotland, and thus covers 14 degrees of latitude (55 to 69 ͦN), and six different bioclimatic zones, from the alpine to the nemoral zone (439,243 km2; Fig. 1). The distributions of the ungulate species (e.g., wild boar, and fallow deer) as well as human activity show a distinct change between the boreal and boreo-nemoral zone (Fig. 1, Appendix 1). The alpine to southern boreal zone have an average of 125–225 days of snow cover and an average vegetation growing period between 100 and 180 days (SMHI), depending on latitude. Tundra vegetation characterizes the alpine zone, while boreal forests dominated by coniferous trees with smaller patches of deciduous forest (mainly birch (Betula sp.) and aspen (Populus tremula)) characterize the boreal zone. Intensive forestry is the dominant land use in the boreal zone. Densities of humans and main roads are considerably lower in the alpine and boreal zones compared with boreo-nemoral and nemoral zones (7 inhabitants per km2 versus 72 inhabitants per km2 in 2017; www.scb.se; 98 m main road per km2 versus 339 m per km2; Fig. 1). Southern Sweden has a milder boreal climate with 25–125 days of snow cover and a vegetation growing period of 180–220 days. Main vegetation consists of a mixture of coniferous and broad-leafed forest with deciduous species, with species such as elm (Ulmus glabra), oak (Quercus robur), maple (Acer platanoides), and beech (Fagus sylvatica) becoming more common. Both forestry and agriculture are the dominant land uses.

Distribution of vegetation zones and major roads across latitudes in Sweden
While moose and roe deer occur throughout Sweden in all bioclimatic zones and regions, the distribution of red deer and particularly fallow deer and wild boar is uneven and mostly limited to southern Sweden (Appendix 1). Harvests of all ungulate species have strongly increased since the 1940s, although moose and roe deer harvests have been more or less stable during the last decades after peaking and then decreasing in the 1980s and 1990s, respectively (Bergström and Danell 2009; www.viltdata.se). In addition to rising harvest numbers, the distribution of red deer, fallow deer, wild boar, and to a lesser extent roe deer has greatly expanded since the 1980s (Apollonio et al. 2010). This process has been facilitated by local release of new individuals and increased food availability through altered land use (including supplementary feeding). Moose are being managed in so-called moose management areas (MMA), which include several hunting parishes where landowners and hunters jointly decide on offtake based on the status of the population and levels of damage to forestry. These management goals are set in 3-year plans that have to be approved by the corresponding County Administrative Board (CAB, www.lansstyrelsen.se). MMA vary in size to broadly match moose biology (particularly migration behavior) in a given area (mean 2,960 km2, minimum 225 km2, maximum 30,173 km2). Red deer, where abundant, are also managed using formal management plans, but often at a smaller spatial scale than moose (mean 175 km2, Swedish EPA 2015). The other ungulate species are hunted without formal management plans; thus, hunting is more opportunistic and oriented toward local conditions and objectives, often in dialogue with the landowners. In Sweden, hunting rights come with owning land.
Hunting is a major source of mortality in all five ungulate species. Impact of large carnivores (wolf (Canis lupus), brown bear (Ursus arctos), lynx (Lynx lynx), wolverine (Gulo gulo)) varies for the different ungulate species and across Sweden (Swedish EPA 2018). Except for roe deer, carnivores mostly affect the survival of calves and sub-adults where carnivores and a given ungulate species co-occur (Swenson et al. 2007; Andren and Liberg 2015; Wikenros et al. 2015; Sand et al. 2016; Swedish EPA 2018). Road traffic may account for up to 10–15% of known mortality for roe deer (Seiler et al. 2004; Seiler and Helldin 2006). Hunters may adapt their harvest of deer to the presence of competing predators, sometimes overcompensating for the effects of predation (e.g. Wikenros et al. 2015).
Data preparation and analyses
We carried out our analyses at three spatial scales for all five species: bioclimatic mean 73,207 ± 51,047 km2 standard deviation (SD), regional (i.e., county, mean 22,146 ± 25,172 km2 SD), and local hunting management area (i.e., the moose management units or MMA, mean 2,960 km2 ± 4,474 km2 SD; Appendix 2). To test for time lags between hunting bags and UVC data for a given species, we evaluated the two data sets covering different periods. Hunting bag statistics covered two hunting seasons (i.e., hunting year 2014/2015 and 2015/2016), which started on July 01 and ended June 30 in the next year. Hunting times are restricted between August and February for all species, except wild boar that can be hunted year around (Appendix 3). We assigned the number of UVC that fell into the same period to a given hunting season.
Management may affect the relationship between hunting bags and UVC data, which may not necessarily show the same pattern in a given year but may have a lagged relationship (e.g., moose, Rolandsen et al. 2011; Ueno et al. 2014; Niemi et al. 2017). To test for the best temporal relationship for each species, we linked the 2014/2015 and 2015/2016 hunting bags of a given species to their UVC statistics from the years 2011 to 2017, generating a series of time lags (t−3, t−2, t−1, t0, and t+1; Appendix 4). We built one model for each time lag. This allowed us to test for five time lags from t−3 to t+1. We corrected the number of UVC for traffic volume and road length for each MMA by dividing the number of UVC by the average year-round number of vehicles per day and kilometers of state roads per MMA (Seiler 2005; NVDB 2014). Traffic volume is recorded for state roads only. More than 80% of all UVC, however, are reported from state roads, and only few are reported from municipality and private roads (Jägerbrand et al. 2018). Hunting bags were collected as heads shot in a given MMA. We calculated and analyzed hunting bags and UVC, as number of animals per 1000 hectare (ha), because this is the unit commonly applied within the ungulate management in Sweden.
We did our analyses separately per species. To aggregate the hunting bag statistics and the UVC data at a given spatial scale, we counted the heads shot and the number of UVC of a given species within a given spatial unit, divided it by the unit’s area in ha, and multiplied it by 1000. In this way, we determined the total number of heads shot and UVC per 1000 ha at the bioclimatic, the county, and the management scale. We used the bioclimatic scale, because in Sweden, the distribution of the different ungulate species strongly varies among the bioclimatic zones (Appendices 1 and 2). More specifically, the main distribution of wild boar covers the nemoral and boreal-nemoral zone, whereas roe deer and moose occur throughout the country. The distribution of red deer and fallow deer is patchier than for the other three species, and they mostly occur in the nemoral and boreal-nemoral zone. The county scale was used because the county is the overarching management scale for moose as ultimately each CAB approves the moose plans of each MMA. Finally, we used the MMA as the main local management scale for moose. Management plans for moose are made at this scale, and moose bag statistics are also given at this scale. Bag statistics for the other ungulates, however, are provided at even a lower scale (i.e., hunting parishes). For the local management scale analysis, we thus need to aggregate bag statistics of all species at the MMA scale. Therefore, we extrapolated the hunting bags of roe deer, wild boar, fallow deer, and red deer from the hunting parishes to the coarser scale of MMA using the percentage share of a given parish within a given MMA. We accessed hunting bag statistics by the County Administrative Boards and the Swedish Association for Hunting and Wildlife Management (www.viltdata.se, www.algdata.se, www.jagareforbundet.se) and UVC data by the National Council for Wildlife-Vehicle Collisions (www.viltolycksradet.se).
For species that did not occur throughout the country (i.e., red deer, fallow deer, and wild boar), we logically did not have hunting bags from all local management units (MMA). Therefore, we only included MMA with hunting bags larger than zero (red deer: 119/148; fallow deer: 99/148; wild boar: 116/148). To account for the hierarchical structure and unbalance in the data, and the repeated measures, we applied linear mixed models. Depending on the scale we tested, we applied the minimum unit as random effect (e.g., county for the regional scale analysis, and MMA for the locale-scale analysis). We applied a nested design to account for the hierarchical structure in the data (i.e., regional scale: county nested in bioclimatic zone; local management scale: MMA nested in county and bioclimatic zone). To meet the assumption of parametric statistics, we transformed our response variable (i.e., hunting bags per 1000 ha) when necessary using log-transformation (roe deer), cube root (red deer), and Tukey’s ladder of powers (fallow deer and wild boar; R package rcompanion, Mangiafico 2019). All results presented are back-transformed. We used model selection based on the AICc to rank the models and to obtain Akaike weights of the model and averaged models considering those with ΔAICc < 2 (R package MuMIn; Bartoń 2019). For each model within this subset, we calculated the likelihood-ratio based pseudo-R-squared value (R2) that represents the variance explained by the fixed factors (R package MuMIn; Bartoń 2019).
We used the software ArcGIS 10.4.1 for all GIS analyses (ESRI, Redlands, CA, USA). We carried out all statistical analyses in the R environment for statistical computing (R 3.6.0). We used a statistical significance of p < 0.05.
Results
Bioclimatic scale
Nationally, 198,396 roe deer (31 % of all ungulates shot), 170,007 moose (27 %), 69,235 fallow deer (11 %), 15,939 red deer (2 %), and 187,107 wild boar (29 %) were harvested during the hunting seasons 2014/2015 and 2015/2016. During the period 2011–2017 (covering the hunting periods 2011/2012–2016/2017), the police registered 202,915 vehicle collisions with roe deer (76% of all UVC), 30,827 with moose (12%), 8809 with fallow deer (3%), 1828 red deer (1%), and 22,071 with wild boar (8%). On average and across species, the number of UVC was 14 % of the total ungulate harvest, with considerable variation among species.
For all ungulates, except moose, several alternative models explain the relationship between hunting bags and UVC equally well (Table 1). The bioclimatic scale did not show any significant relationship between hunting bags and UVC, except for moose and wild boar where the most parsimonious models suggested a positive relationship for t0 in moose and three time lags (t−3, t−1, and t0) in wild boar (Table 2).
Regional (county) scale
At the county scale, UVC data correlated positively to hunting bags for all species, with more UVC where more animals were shot (Table 2). The models explain a high proportion of the variance (≥ 0.85, Table 1). In contrast to the bioclimatic scale, we found distinct temporal relationships for each species with one time lag being the most relevant, except for roe deer and fallow deer where the time lags t−3 and t0 explained the relationship between bag statistics and UVC equally well (Table 1).
Local (management) scale
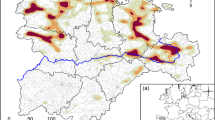
The number of animals shot was positively related to the number of UVC during the same year for all ungulate species, except for moose where a time lag of two years (t−2) fit best (moose: F1,147 = 2.2, p = 0.03; roe deer: F1,147 = 1.9, p = 0.049; red deer: F1,101 = 4.8, p < 0.0001; wild boar: F1,114 = 4.2, p = 0.0001; Fig. 2a). For fallow deer, the number of animals shot was positively related to the number of UVC from the same year and from 3 years back (t−3: F1,92 = 3.8, p = 0.0004; t0: F1,92 = 3.8, p = 0.0004). For all species, the fitted estimates differ among management units (Fig. 2a). For fallow deer, the fitted estimates of the two time lags showed similar patterns among management units, but their size varied largely between the two suggested time lags. This suggests that the number of fallow deer shot increased more rapidly with UVC from 3 years ago than with UVC of the same year (Fig. 2b). For moose, we found the largest estimates along the coast and in the inland in southern Sweden, meaning that relatively more moose were shot per UVC in these MMA (Fig. 2a). However, estimates vary considerably among neighboring management units (Fig. 2a). For roe deer, estimates are high in southern Sweden with the highest in the southern-most units and at the coast, suggesting a relative higher number of harvested roe deer in relation to those involved in collisions (Fig. 2 A). For wild boar, we find high estimates throughout southern Sweden with higher values in the eastern half (Fig. 2a). For red deer, higher estimates are patchily distributed with higher values around and south of Stockholm and in the most southern unit (Fig. 2a).

Average estimates given by the most parsimonious linear mixed model to test the relationship between hunting bags per 1000 ha and ungulate-vehicle collisions (UVC) per 1000 ha in a management area by moose, roe deer, fallow deer, red deer, and wild boar (a), and fallow deer (b) in Sweden. Darker colors indicate higher values
The proportion of the explained variation varies among species with the largest amount explained in roe deer (R2 = 0.91) and the lowest in moose (R2 = 0.47; Table 1). This suggests a strong relationship between the number of roe deer killed by vehicles and by hunters, but a poor one for moose. The proportion of the explained variance is also relatively high for the other three ungulate species (R2: 0.72 (fallow deer), 0.78 (red deer), 0.83 (wild boar); Table 1).
Discussion
Existing studies that link ungulate hunting bags to other sources of monitoring data generally focused on one or two species, and did not consider hunting bags from multiple co-occurring species (but see Saint-Andrieux et al. 2020). Moreover, only few studies evaluated the relationship on the scale of the actual management unit (but see, Seiler 2004). We tested the correlation between hunting bag statistics and UVC data, both used for monitoring in ungulate management, for the five main European sympatric ungulate species (i.e., moose, roe deer, red deer, fallow deer, and wild boar). For all study species, our results highlight that UVC correlated positively with hunting bags at the regional, county, and the local management scale. For moose and wild boar, both indices also correlated positively at the largest, bioclimatic, scale. The time lags, at which the two indices correlated best, varied among ungulate species at the county scale. This means that models have to be adjusted per species, if managers want to use both indices for management decisions at this scale. On the other hand, with the local management scale (i.e., the moose management area or MMA), the indices correlated best without time lags for all species, except moose. For fallow deer, a time lag of 3 years explained the variation equally well. Except for moose, the correlation between UVC and bag statistics at the local management scale was also rather strong (R2 > 0.7).
For moose, we found a time lag of 2 years between UVC and bag statistics to result in the best fit, being in line with previous research (Rolandsen et al. 2011; Ueno et al. 2014; Niemi et al. 2017). Yet, our results also highlight a low correlation between collisions with moose and number of shot moose (R2 0.47), indicating a poor relationship between these two methods. Thus, other factors may affect each of these two statistics (Seiler 2004; Rolandsen et al. 2011; Ueno et al. 2014). In contrast to previous research, we analyzed the relation between UVC and bag statistics at the actual scale of moose management, the MMA. In Sweden, moose harvest quotas are controlled by 3-year management plans, and these plans therefore generally consider several long-term management goals (e.g., percentage of adult males and calves in the population, browsing damage), which are not necessarily correlated to moose density in a given year and the occurrence of moose-vehicle collisions (www.naturvardsverket.se). This is an important aspect to keep in mind, and it likely contributes to explaining the 2-year time lag, but also to the poor correlation between hunting bags and UVC that we observed. For all other ungulate species, we found that UVC data per 1000 ha correlated best with hunting bags during the same year on the local management scale. Those species, except red deer in areas with dense populations, are not formally managed, and harvest quotas and management goals are thus not set in long-term management plans in the same way as for moose. Consequently, management is most strictly organized for moose, followed by red deer, and then the other smaller ungulate species. Thus, due to the lack of a long-term management plan, harvests of smaller ungulates are controlled less strictly over time compared with moose. As a result, we suggest that harvest quotas are more proportional to the density of these species than for moose, because individual landowners and hunters have more flexibility to adjust their harvest rates to varying deer densities on a yearly basis and on their respective hunting ground. This may explain the more straightforward correlation between hunting bags and UVC data for the smaller ungulates due to a more bottom-up, informal, management approach compared with the more formal management approach in moose, which involves more stakeholders and is in that sense more top-down.
During our study period, roe deer stand for 31 % of the total ungulate harvest, whereas roe deer accounted for 76% of all UVC. In contrast, moose harvest and UVC accounted for 27% and 12%, respectively, of the total. Thus, we see a considerable variation among species in their relationship between UVC and hunting bag statistics, which may have implications for the applicability of either method in a given species. Differences in the life histories are crucial to consider while explaining the relationships between these kinds of data sets for the species in consideration. The important life cycle events such as calving, migration, and rut of these species occur at different times during the year, and the number of young produced per reproductive event as well as sex ratio of calves may be different (Gaillard et al. 2000). This may reflect in their birth and death rates as well as population dynamics and possibly influencing the relationships between such indices of density and abundance as well as the time lag relationships. The hunting methods applied in a given area may influence the relationship between bag and UVC numbers of a given ungulate species. Hunting disturbances (as targeted and non-targeted species) affect animal movement activity and distribution temporally (e.g., roe deer Grignolio et al. 2010, red deer Sunde et al. 2009, moose Ericsson et al. 2015, wild boar Thurfjell et al. 2013), which may result in more animals crossing roads, possibly increasing the UVC-risk. Different hunting methods affect animal response differently with driven hunts or hunting with hounds generally having a stronger impact than stalking or still hunt (e.g. roe deer Grignolio et al. 2011, wild boar Thurfjell et al. 2013). Yet, other factors than hunting disturbances may influence animal movement and UVC-risk more strongly (Neumann et al. 2012; Steiner et al. 2014; Neumann and Ericsson 2018; Saint-Andrieux et al. 2020), and most of the wild boar in Sweden are shot over bait (Swedish Government Official Report 2014).
Given their high contribution in total UVC, the reduction in UVC numbers of particularly roe deer in a given management unit may be of high interest, compared with other management interests and other ungulate species. Collisions with moose produce six times larger societal costs than the cost of collisions with roe deer (Gren and Jägerbrand 2017). Yet, UVC with roe deer occur 8 times more often (Neumann et al. 2011), thereby also becoming a considerable socio-economical cost (Apollonio et al. 2010; Gren et al. 2018). In central Europe, UVC with roe deer correlate positively with both browsing intensity and harvest numbers (Hothorn et al. 2012), suggesting that UVC data can represent densities and distribution in roe deer. Compared with the other ungulates species, we found that in roe deer, the number of UVC explained the highest percentage of the variance in hunting bags, suggesting a good fit between the two data sets. Given its biology (i.e., activity pattern, proximity to human settlements; Torres Tinoco et al. 2011; Neumann et al. 2019), roe deer might be more exposed to traffic compared with other ungulate species and in relation to their abundance (Seiler 2004).
Management interests vary among species and stakeholders (Linnell et al. 2020). For wild boar, next to recreational hunting, damage on agricultural fields and UVC are the driving management interests in many places (Apollonio et al. 2010; Linnell et al. 2020). In management areas with well-established wild boar populations, the supply of wild meat is an additional important reason for hunting wild boar. Like roe deer, the number of UVC and hunting bags also showed a strong fit (high R2) for wild boar. In Sweden, the wild boar population is currently increasing quickly, which may support the close relationship between UVC and hunting bags. A similar pattern was observed in moose in the late 1970s prior to the peak-years in moose harvest (Lavsund et al. 2003) and in roe deer during the population rise in the mid-1980s (Apollonio et al. 2010). Hunting adult wild boar is allowed between April and February, and yearlings can be shot year-around, thus covering a considerably longer part of the hunting year compared with all other ungulate species we evaluated. In addition, there are no hunting quotas that restrict the number of wild boar shot. Most collisions with wild boar occurred between September and March (www.viltolycka.se). Because hunting of wild boar is allowed during the entire year, the timing of hunting and UVC overlap by necessity. Most of the wild boar are shot over bait in Sweden, and only 13% are shot during driven hunts (Swedish Government Official Report 2014), which potentially may increase UVC through increased movement. Given the lack of set hunting quotas, landowners thus may adjust their harvest more directly in response to higher densities and where damage rates are higher (e.g., more UVC).
Our results suggest a good correlation between red deer UVC and the number of shot red deer, which was unexpected since red deer are managed using formal management plans similar to moose management (Swedish EPA 2015). We have no definite answer to why the correlation for red deer is much stronger than for moose, but one hypothesis we have is that red deer quotas are treated more flexibly than moose quotas and that it is more likely that hunters are allowed to shoot more red deer after they have filled their quotas (Widemo, personal experience). An underlying reason might be that the formal red deer management was only initiated very recently, and red deer hunters may thus still be adjusting to this new way of management. In Sweden, red deer is closely connected to the agriculture-forest landscape that also holds high densities of roads and humans. Landscape composition affects both movement patterns and frequency of vehicle UVC with red deer (Allen et al. 2014; Hedlund and Hamre 2018; Saint-Andrieux et al. 2020), possibly increasing the frequency of vehicle UVC with red deer where regular movement across different habitat types and many major roads with more traffic coincide. Fallow deer are also closely connected to the agriculture-forest landscape, but we found two time lags that explain the correlation between UVC and hunting bags equally well, yet with similar overarching spatial pattern. In contrast to red deer, fallow deer had a heavily skewed ratio of harvested deer (11 % of all ungulate harvested) to number of UVC (1 % of all UVC). Fallow deer’s movement behavior is limited, which maintains their heavily fragmented distribution across the country (Apollonio et al. 2010), as well as it may explain the observed low percentage of UVC compared with percentage of harvested fallow deer. In summary, it is thus important to realize the different approaches to manage a given ungulate species, as this can influence the relation between hunting bag size and the actual density of the species. This relation might be indeed weaker for species such as moose, which are managed for diverse reasons, than for the other species, which would aid to explain the low correlation between hunting bags and UVC data for moose.
It is important to note the possible limitation of UVC data to reflect population distribution adequately in areas where human population is low and/or the road network is sparse due to lower likelihood of UVC in spite of the presence of the ungulate species (Colino-Rabanal and Peris 2016). Similarly, low ungulate population densities will result in fewer UVC for an area of a given size, and the number of accidents will affect the ability to use UVC as a relative abundance proxy (Rolandsen et al. 2011; Ueno et al. 2014). Even though hunting bags present a more active sampling of the population, low population densities of course also affect their value as a relative abundance proxy. For species with very high UVC numbers, UVC data are likely more suitable for estimating population distribution and density at lower spatial resolution compared with species with lower numbers of UVC. Similarly, hunting bag data experience flaws. Even though remote parts of Sweden, and areas in more populated areas but with few roads, are visited for hunting, which may generate a stronger correlation between ungulate abundance and harvest compared with abundance and UVC data, we still assume some deficits of hunting bags in less accessible areas where hunting pressure might be lower. In summary, we expect the spatial correlations between hunting bag statistics and UVC to be higher in areas where sampling of the population by traffic and hunting occurs in a similar way, which likely is in areas with more roads.
Areas may also differ in their capacity or willingness to reduce ungulate populations in relation to number of UVC. In many countries, the number of hunters is decreasing as well as aging, whereas ungulate numbers increase, bearing risk that the remaining hunter cohort has difficulties to control ungulate numbers and thus to reduce damages caused by wildlife (Riley et al. 2003; Massei et al. 2015; Gren et al. 2018). Lastly, in order to consider UVC data as a proxy for relative ungulate abundance and distribution, an important precondition is of course that UVC are reported regularly and with a large national coverage. Misclassification or underreporting of UVC can affect the data quality, particularly if it varies among species and occurs in non-random manner (Snow et al. 2015; Niemi et al. 2017). In Sweden, although reporting of UVC is mandatory, 10–15% of incidences are not reported, and further 5–10% may not be coded as UVC or provide insufficient or erroneous information (Seiler and Jägerbrand 2016). Hunting bag statistics may also be suffering from similar problems. Moose and red deer shot must be reported, but it is voluntary to report the harvest of fallow deer, roe deer, and wild boar. Thus, results from voluntary reports are extrapolated across hunting parishes to yield estimated total bag statistics (SAHWM 2019). To improve applicability of UVC data within wildlife management, we recommend future research to study magnitude and pattern of missed UVC and to investigate non-linear effects of traffic volume, speed, and road parameters (e.g., fencing, lighting, and guard rails) as well as vehicle technology on UVC occurrence.
Management implications
Our results emphasize different patterns in the relationship between UVC data and hunting bag statistics among the ungulate species studied. More specifically, we found a close correlation between UVC and hunting bags for species where management was least formal and done by individual hunters or hunting teams (roe deer and wild boar) rather than the more formal management by groups of stakeholders for moose and to some extent red deer. For these latter two species, quotas are set on a longer time frame and are the result of discussions by stakeholders that may have varying goals (e.g., forestry that aims at minimizing damage, and thus perhaps ungulate densities, versus hunters that may want to maintain higher densities for future hunting success). As a result, harvest quota may not necessarily correspond to actual current density but may be compensatory responses with a lag in time (Ueno et al. 2014). As a consequence, particularly for moose as the most formally managed species with a very poor correlation between the two indices, this can generate conflicting conclusions for management depending on the method used. For the least formally managed species, roe deer and wild boar, both indices correlated strongly in this Swedish context and thus likely lead to similar support for management decisions. To test whether the type and intensity of management indeed influences the relation between UVC and bag statistics in the way we described, we need more studies in other European countries where these indices are compared for multiple species simultaneously.
Change history
22 October 2020
For the first author, the given and family name are mixed up.
References
Allen AM, Månsson J, Jarnemo A, Bunnefeld N (2014) The impacts of landscape structure on the winter movements and habitat selection of female red deer. Eur J Wildl Res 60:411–421
Andren H, Liberg O (2015) Large impact of Eurasian lynx predation on roe deer population dynamics. PLOS One 10:e0120570
Apollonio M, Andersen R, Putman R (2010) European ungulates and their management in the 21st century. Cambridge University Press
Bartoń K (2019) R package MuMIn with tools for performing model selection and model averaging. https://cran.r-project.org/web/packages/MuMIn/index.html
Bergström R, Danell K (2009) Mer vilt i dag än för 50 år sedan. Vilt och fisk fakta 4. Vilt & Fisk Adaptiv Förvalting. Swedish University of Agricultural Science [In Swedish]
Bíl M, Kubeček J, Sedoník J, Andrášik R (2017) Srazenazver.cz: A system for evidence of animal-vehicle collisions along transportation networks. Biological Conservation 213:167–174
Brazeal JL, Weist T, Sacks BN (2017) Noninvasive genetic spatial capture-recapture for estimating deer population abundance. J Wildl Manage 81:629–640. https://doi.org/10.1002/jwmg.21236
Colino-Rabanal VJ, Peris SJ (2016) Wildlife roadkills: improving knowledge about ungulate distributions ? Hystrix-Italian Journal of Mammalogy 27:2. https://doi.org/10.4404/hystrix-27.2-11279
ENETWILD-consortium, Acevedo P, Croft S, Smith GC, Blanco-Aguiar JA, Fernandez-Lopez J, Scandura M, Apollonio M, Ferroglio E, Keuling O, Sange M, Zanet S, Brivio F, Podgórski T, Petrović K, Body G, Cohen A, Soriguer R, Vicente J (2019) ENETwild modelling of wild boar distribution and abundance: update of occurrence and hunting data-based models. European Food Safety Authority. EFSA Supporting publication 2019:EN-1674. https://doi.org/10.2903/sp.efsa.2019.EN-1674
Eriksen LF, Moa PF, Nilsen EB (2018) Quantifying risk of overharvest when implementation is uncertain. J Appl Ecol 55:482–493. https://doi.org/10.1111/1365-2664.12992
Ferretti F, Fattorini L, Sforzi A, Pisani C (2016) The use of faeces counts to estimate relative densities of wild boar in a Mediterranean area. Population Ecol 58:329–334. https://doi.org/10.1007/s10144-016-0536-3
Gaillard JM, Festa-Bianchet M, Yoccoz NG, Loison A, Toigo C (2000) Temporal variation in fitness components and population dynamics of large herbivores. Annual Review of ecology and Systematics 31:367–393
Gren I-M, Jägerbrand A (2017) Costs of animal-vehicle collisions with ungulates in Sweden. Working Paper. Swedish University of Agricultural Sciences 2017:03
Gren I-M, Haggmark-Svensson T, Elofsson K, Engelmann M (2018) Economics of wildlife management-an overview. Eur J Wildl Res 64:22. https://doi.org/10.1007/s10344-018-1180-3
Grignolio S, Merli E, Bongi P, Ciuti S, Apollonio M (2011) Effects of hunting with hounds on a non-target species living on the edge of a protected area. Biol Conserv 144:641–649
Gunson KE, Clevenger AP, Ford AT, Bissonette JA, Hardy A (2009) A comparison of data sets varying in spatial accuracy used to predict the occurrence of wildlife-vehicle collisions. Environ Manage 44:268–277
Hedlund SJ, Hamre LN (2018) Scale-dependent effects of landscape composition and configuration on deer vehicle collision and their relevance to mitigation and planning options. Landsc Urban Plan 169:178–184
Heigl F, Stretz C, Steiner W, Suppan F, Bauer T, Laaha G, Zaller J (2016) Comparing Road-Kill Datasets from Hunters and Citizen Scientists in a Landscape Context. Remote Sensing 8:832
Hothorn T, Brandl R, Muller J (2012) Large-Scale Model-Based Assessment of Deer-Vehicle Collision Risk. Plos One 7:e29510
Hothorn T, Mueller J, Held L, Most L, Mysterud A (2015) Temporal patterns of deer-vehicle collisions consistent with deer activity pattern and density increase but not general accident risk. Accident Analysis and Prevention 81:143–152
Imperio S, Ferrante M, Grignetti A, Santini G, Focardi S (2010) Investigating population dynamics in ungulates: do hunting statistics make up a good index of population abundance? Wildl Biol. 16:205–214. https://doi.org/10.2981/08-051
Jägerbrand AK, Gren I-M, Seiler A, Johansson Ö (2018) Uppdatering och nya effektsamband i effektmodellen för viltolyckor. Calluna AB, Linköping. https://doi.org/10.13140/RG.2.2.15211.23849
Jarnemo A, Jansson G, Månsson J (2017) Temporal variations in activity patterns during rut - implications for survey techniques of red deer, Cervus elaphus. Wildl Res 44:106–113. https://doi.org/10.1071/WR16156
Lavsund S, Nygrén T, Solberg EJ (2003) Status of moose populations and challenges to moose management in Fennoscandia. Alces 39:109–130
Linnell JDC, Cretois B, Nilsen EB, Rolandsen CM, Solberg EJ, Veiberg V, Kaczensky P, Van Moorter B, Panzacchi M, Rauset GR, Kaltenborn B (2020) The challenges and opportunities of coexisting with wild ungulates in the human-dominated landscapes of Europe’s Anthropocene. Biol Conserv 244:108500. https://doi.org/10.31219/osf.io/2heqx
Mangiafico S (2019) R package rcompanion with statistical tools. https://cran.r-project.org/web/packages/rcompanion/index.html
Månsson J, Hauser CE, Andren H, Possingham HP (2011) Survey method choice for wildlife management: the case of moose Alces alces in Sweden. Wildl Biol 17:176–190
Massei G, Coats J, Lambert MS, Pietravalle S, Gill R, Cowan D (2018) Camera traps and activity signs to estimate wild boar density and derive abundance indices. Pest Manage Sci 74:853–860. https://doi.org/10.1002/ps.4763
Mysterud A (2004) Temporal variation in the number of car-killed red deer Cervus elaphus in Norway. Wildl Biol 10:203–211
Neumann W, Ericsson G (2018) Influence of hunting on movements of moose near roads. J Wildl Manage 82:918–928
Neumann W, Ericsson G, Sävberger L, Seiler A (2011) Adaptiv älgförvaltning nr 12: Älg och trafik. Faktablad Fakta Skog 21, p 4. Updated June 2019.
Neumann W, Ericsson G, Dettki H, Bunnefeld N, Keuler N, Helmers D, Radeloff V (2012) Difference in spatiotemporal patterns of wildlife road-crossings and wildlife-vehicle collisions. Biol Conserv 145:70–78
Neumann W, Stenbacka F, Malmsten J, Nilsson K, Dettki H, Singh N, Ericsson G (2019) Slutrapport: Älg och rådjur i stormanas spar – GPS-märkta älgar och rådjur I Växjö 2015-2019. Sveriges lantbruksuniversitet, 28 pp
Niemi M, Rolandsen CM, Neumann W, Kukko T, Tiilikainen R, Pusenius J, Solberg EJ, Ericsson G (2017) Temporal patterns of moose-vehicle collisions with and without personal injuries. Accident Analysis and Prevention 98:167–173
NVDB (2014) Swedish Transport Administration. Digital maps on the road network with meta data on road features. www.trafikverket.se
Pfeffer SE, Spitzer R, Allen AM, Hofmeester TR, Ericsson G, Widemo F, Singh NJ, Cromsigt JPGM (2017) Pictures or pellets? Comparing camera trapping and dung counts as methods for estimating population densities of ungulates. Remote Sens Ecol Conserv 4:173–183
Putman R, Watson P, Langbein J (2011) Assessing deer densities and impacts at the appropriate level for management: a review of methodologies for use beyond the site scale. Mammal Review 41:197–219. https://doi.org/10.1111/j.1365-2907.2010.00172.x
Riley SJ, Decker DJ, Enck JW, Curtis PD, Lauber TB, Brown TL (2003) Deer populations up, hunter populations down: Implications of interdependence of deer and hunter population dynamics on management. Ecoscience 10:455–461. https://doi.org/10.1080/11956860.2003.11682793
Rolandsen CM, Solberg EJ, Herfindal I, Van Moorter B, Sæther B-E (2011) Large-scale spatiotemporal variation in road mortality of moose: Is it all about population density? Ecosphere 2:113
Rosell C, Fernández-Bou M, Camps F, Boronat C, Navàs F, Martínez M, Sorolla A (2013) Animal vehicle collisions: A new cooperative strategy is needed to reduce the conflict. Proceedings ICOET 2013 International Conference on Ecology and Transportation. Scottsdale, Arizona, USA. www.icoet.net
Saint-Andrieux C, Calenge C, Bonenfant C (2020) Comparison of environmental, biological and anthropogenuic cuases of wildlife-vehicle collisions among three large herbivore species. Population Ecology 62:64–79. https://doi.org/10.1002/1438-390X.12029
Sand H, Eklund A, Zimmermann B, Wikenros C (2016) Prey selection of Scandinavian wolves: single large or several small? Plos One 11:e0168062
Seiler A (2004) Trends and spatial patterns in ungulate-vehicle collisions in Sweden. Wildl Biol 10:301–313
Seiler A, Helldin JO (2006) Mortality in wildlife due to transportation. The ecology of transportation: managing mobility for the environment. J. Davenport and J. L. Davenport. Amsterdam, Kluwer: 165–-190
Seiler A, Jägerbrand AK (2016) Mörkertal i viltolycksstatistik – tolkning av viltolycksenkät och databasanalyser (Hidden statistics in wildlife-vehicle accident statistics - interpretation of drivers surveys and database comparisons). Borlänge, Trafikverket Publikation 2016:055
Seiler A, Helldin JO, Seiler C (2004) Road mortality in Swedish mammals: results of a drivers’ questionnaire. Wildl Biol 10:225–233
Seiler A, Willebrand S, Olsson M, Wahlman H (2019) Viltolyckskartor - Teknisk beskrivning för datahantering och produktion. (Wildlife-vehicle accident maps - technical description for data management and production. In Swedish only.) - Swedish Transport Administration, Publication 2019:179, TVD-52097. http://trafikverket.diva-portal.org/smash/record.jsf?pid=diva2%3A1366843&dswid=-3646
Seiler A, Rosell C, Torellas M, Sjölund M, Rolandsen CM, Solberg EJ, Van Moorter B, Lindstrøm IM (2016) Case studies on the effect of local road and verge features on ungulate-vehicle collisions. SAFEROAD Technical report 8, Conference of European Directors of Roads (CEDR).
Snow NP, Porter WF, Williams DM (2015) Underreporting of wildlife-vehicle collisions does not hinder predictive models for large ungulates. Biolog Conserv 181:44–53
Steiner W, Leisch F, Hacklaender K (2014) A review on the temporal pattern of deer-vehicle accidents: impact of seasonal, diurnal and lunar effects in cervids. Accidents Analysis and Prevention 66:168–181
Sunde P, Olesen CR, Madsen TL, Haugaard L (2009) Behavioural responses of GPS-collared female red deer Cervus elaphus to driven hunts. Wildl Biol 15:454–460
Swedish Government Official Report (2014) Vildsvin och viltskador - om utfodring, kameraövervakning och arrendatorers jakträtt, p. 75. SOU 2014:54 https://www.regeringen.se/contentassets/183d45f8bbb64fdb8477581e370a5110/vildsvin-och-viltskador%2D%2D-om-utfodring-kameraovervakning-och-arrendatorers-jaktratt-sou-201454
Swedish EPA (2015) An evaluation of Swedish deer management. Swedish Environmental Protection Agency, Report 6673. In Swedish.
Swedish EPA (2018) Information about distribution of the large carnivores given by the Swedish Environmental Protection Agency. www.naturvardsverket.se. Accessed 23 June 2018
Swedish Association for Hunting and Wildlife Management (2019) National hunting bag statistics on all game species in Sweden. www.jagareforbundet.se
Swenson JE, Dahle B, Busk H, Opseth O, Johansen T, Söderberg A, Wallin K, Cederlund G (2007) Predation on moose calves by European brown bears. J Wildl Manage 71:1993–1997
Thurfjell H, Spong G, Ericsson G (2013) Effects of hunting on wild boar Sus scrofa behavior. Wildl Biol 19:87–93
Torres Tinoco R, Carvalho JC, Panzacchi M, Linnell JDC, Fonseca C (2011) Comparative use of forest habitats by roe deer and moose in a human-modified landscape i southeastern Norway during winter. Ecol Res 26:781–789. https://doi.org/10.1007/s11284-011-0837-0
Ueno M, Solberg EJ, Iijima H, Rolandsen CM, Gangsei LE (2014) Performance of hunting statistics as spatiotemporal density indices of moose (Alces alces) in Norway. Ecosphere 5:2
Waetjen DP, Shilling FM (2017) Large Extent Volunteer Roadkill and Wildlife Observation Systems as Sources of Reliable Data. Frontiers in Ecology and Evolution 5. Dot: https://doi.org/10.3389/fevo.2017.00089
Wikenros C, Sand H, Bergstrom R, Liberg O, Chapron G (2015) Response of moose hunters to predation following wolf return in Sweden. Plos One 10:e0119957
Willebrand T, Hornell-Willebrand M, Asmyhr L (2011) Willow grouse bag size is more sensitive to variation in hunter effort than to variation in willow grouse density. Oikos 120:1667–1673. https://doi.org/10.1111/j.1600-0706.2011.19204.x
Acknowledgments
Open access funding provided by Swedish University of Agricultural Sciences. We are grateful to the Swedish Association for Hunting and Wildlife Management for supplying us with data from the Swedish Game Monitoring and to the Swedish National Wildlife accident Council for providing UVC statistics. We thank S. Juthberg for help with editing the English in this paper.
Funding
This study formed part of the research program “Beyond Moose–ecology and management of multispecies ungulate systems” and was financially supported by the Swedish Environmental Protection Agency (Beyond Moose NV-01337-15/NV-03047-16/NV-08503-18).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interest
The authors declare that they have no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Road Ecology
Guest Editor: Marcello D’Amico
Electronic supplementary material
ESM 1
(DOCX 2.52 mb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Neumann, W., Widemo, F., Singh, N.J. et al. Strength of correlation between wildlife collision data and hunting bags varies among ungulate species and with management scale. Eur J Wildl Res 66, 86 (2020). https://doi.org/10.1007/s10344-020-01421-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-020-01421-x