
Abstract
Increasing abundance of large ungulates is raising human-wildlife impacts and the effectiveness of recreational hunting to reduce their population growth is increasingly questioned. We report on long-term trends (> 15 years) in wildlife-vehicle collisions (WVC) and hunting bags, and on associations between the annual growth rate of WVC and that of hunting bags for three ungulates – the wild boar, the red deer, and the roe deer – and the grey wolf in northwest Spain to evaluate the regulating capacity of recreational hunting at large spatial scale. Wildlife-vehicle collisions increased by 332% in 16 years and 91% of all traffic accidents were caused by collisions with these three ungulates. All ungulate species showed significant positive trends in WVC and hunting bags, but we did not observe a negative association between annual growth rate of hunting bags and that of WVC except for the wild boar. Results suggest that recreational hunting was unable to reduce ungulate population growth at the regional scale. There was no upward trend of vehicle collisions with wolves over the study period, possibly reflecting stable wolf populations. Natural mortality due to predation could be promoted through the protection of apex predators, but the lethal management of apex predators, often based on sociopolitical pressures rather than damage levels, can conflict with the strategy for mitigating ungulate impacts. Ungulate management needs to be reconsidered from an ecological perspective that integrates human management measures, including recreational hunting, based on the population dynamics and the recovery of predator–prey interactions by favoring the expansion of apex predators.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Humans are responsible for the massive loss of Earth’s biodiversity (Dirzo et al. 2014; Johnson et al. 2017). However, human actions also underlie the population growth and geographical expansion of many species, which are thriving due to multiple factors such as land-use changes, release from ecological constraints, or legislative changes on wildlife management (Martin et al. 2020; Pyšek et al. 2020). Increasing abundance of these species can result in conflicts when they cause socio-economic and ecosystem impacts (i.e., overabundant species sensu Caughley 1981). Mitigation of damages caused by such overabundant species involves human interventions to reduce population numbers or nuisances, including lethal and nonlethal methods such as fertility control, barriers to movements, habitat management or compensatory payments among others (Nyhus 2016; Pyšek et al. 2020).
Populations of large ungulates are considered overabundant in many regions of Europe and North America due to the rapid growth experienced in recent years (Apollonio et al. 2010; Côté et al. 2004; Valente et al. 2020). Although large ungulates were driven almost to the brink of extinction in many regions of Europe and North America by the nineteenth century, their recovery was favoured since the mid-twentieth century by a suite of conditions, including large-scale processes of rural–urban migration, natural afforestation, mild winters, lack of natural predators, and legislative changes on management (Deinet et al. 2013; Martin et al. 2020). Large ungulates, when overabundant, provoke important damages to crop and forestry production, raise the risk of disease transmission to humans and livestock and of traffic accidents, as well as nuisance to humans when animals wander into urban areas (Côté et al. 2004; Linnell et al. 2020; Martin et al. 2020; Valente et al. 2020). Ungulate-induced damages can generate large economic costs when populations are at high densities: deer-vehicle collision costs can exceed $1 billion annually in the U.S.A. (Conover et al. 1995); in Spain, the annual economic cost of wildlife-vehicle collisions, mostly due to ungulate game species, is estimated in €105 million (Sáenz-de-Santa-María and Tellería, 2015); annual damage to agricultural crops by wild boars (Sus scrofa) costs €80 million in Europe (Apollonio et al. 2010) and $800 million in the U.S.A. (Pimentel et al. 2005). Management of overabundant populations of large ungulates is therefore a major challenge for national and regional administrations (Apollonio et al. 2017).
In Europe, recreational hunting (i.e., hunting for self-amusement or trophies) constitutes the main source of mortality and the principal management tool to control large ungulate populations (Apollonio et al. 2017; Quirós-Fernández et al. 2017). Yet, despite increasing hunting pressure, populations of large ungulates have steadily grown in many regions (Milner et al. 2006). Red deer (Cervus elaphus) harvest increased by 400–700% in several European countries (Milner et al. 2006) whereas population size estimates increased from 1.1 million to 1.7 million individuals in Europe around the last quarter of the twentieth century (Burbaite and Csányi 2010). Thus, evidence calls into question the role of recreational hunting as an effective management tool to halt the widespread population increments of large ungulates and to reduce associated damages (Massei et al. 2015; Melis et al. 2009; Simard et al. 2013; but see Hothorn and Müller 2010). Recreational hunting of large ungulates acts mostly on adult male mortality but less often on the reproductive potential of populations via female or juvenile killing, productivity being a key demographic parameter to suppress population growth (Ueno et al. 2010; Eacker et al. 2017). Moreover, the capacity of recreational hunting to reduce population size of overabundant ungulates is expected to further decrease when hunter numbers or the social acceptance of lethal management are declining (Liordos et al. 2017; Massei et al. 2015).
Restoring predator–prey interactions is another complementary yet controversial measure to reduce the impacts of overabundant ungulates (Apollonio et al. 2017; Marrotte et al. 2022; Nilsen et al. 2007). Large carnivores predate on large ungulates (Jedrzejewska and Jedrzejewski 1998; Jedrzejewski et al. 2002; Ripple and Beschta 2012; van Beeck Calkoen et al. 2023) and modulate prey behaviour through landscapes of fear (Epperly et al. 2021; Gaynor et al. 2019). The grey wolf (Canis lupus) – the apex predator with the largest distribution in Europe (Chapron et al. 2014) – can reduce the risk of disease transmission from wild boars to livestock by removing diseased individuals (Tanner et al. 2019) and the incidence of deer-vehicle collisions by keeping ungulates away from roads (Raynor et al. 2021). Recent studies estimate that wolf predation leads to annual reductions in the cost of ungulate-vehicle collisions of about $157,000 – $377,000 and €4,500 – €14,700 per wolf in the U.S.A. and France, respectively (Raynor 2017; Sèbe et al. 2022). The annual compensation cost for wolves was €7.9 million in Europe (Bautista et al. 2019), far below the costs of wild boar’s damages to agriculture (€80 million; Apollonio et al. 2010). Despite the benefits of large carnivores to human societies through their complex effects on biological networks (Gilbert et al. 2017; Kuijper 2011; Ripple et al. 2014), there is an enormous social pressure to undertake lethal management (e.g., to prevent livestock attacks), which has been or is permitted in many regions (e.g., Sweden, Norway, Finland, Estonia, Romania, Spain, USA) despite poor knowledge on their population dynamics or bad conservation status (Creel et al. 2015; Laikre et al. 2022; Quevedo et al. 2019).
We report here on temporal trends in recreational hunting in northwest Spain, and on the effectiveness of this wildlife management tool to reduce problems associated with overabundant ungulates, specifically wildlife-vehicle collisions (WVC). We used traffic accident records and hunting bags of the three most abundant and widely distributed ungulates across Europe: the wild boar, the roe deer (Capreolus capreolus), and the red deer (Deinet et al. 2013). Traffic accidents constitute a major impact of overabundant ungulates and can be also used as a proxy of population abundance in these species (e.g., Fernández-López et al. 2022; Neumann et al. 2020; Saint-Andrieux et al. 2020). The study region, Castilla y León, is of particular interest to investigate this issue due to its socioecological context. Castilla y León presents the highest rate of WVC in Spain (Sáenz-de-Santa-María and Tellería, 2015) and is one of the largest most unpopulated regions of Europe (https://ec.europa.eu/eurostat), where the sociological factors that favour the growth of ungulate populations such as rural emigration, afforestation, or agricultural intensification will likely continue over the next years. In addition, it is one of the few European regions where wolves have persisted despite massive persecution in 1850–1970 and farmers have maintained traditional measures to prevent livestock attacks (Deinet et al. 2013). Because the study region is well suited to consider opportunities for an integrative management rooted in the functioning of natural systems, which also takes the role of apex predators into account, we also evaluated the temporal trends of vehicle collisions and recreational hunting for the wolf.
To explore the role of recreational hunting as a management tool for ungulate and wolf populations, we first evaluated long-term changes (more than 15-year data) by determining the temporal trend in WVC and in hunting bags for each species. We considered that recreational hunting would be an effective management tool if it caused negative or no population growth in the long term, which should be observed through a declining or asymptotic pattern of WVC, respectively. We also explored the potential short-term effects of recreational hunting by analysing relationships between the annual growth rate of hunting bags and of WVC of the same and the following year. We used two time-lags because the effects of harvesting on the number of traffic accidents could be immediate due to reduced population size or delayed if harvest causes lower recruitment rate. If hunting was effective at limiting population growth, we expected a negative relationship between the annual growth rate of WVC and that of hunting bags because an increased harvest rate may reduce population growth and thereby the number of WVCs.
Methods
Study area
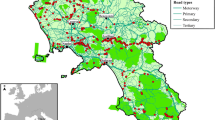
Castilla y León (94,226 km2; Spain, southwestern Europe, Fig. 1a) is characterized by continental-Mediterranean climate and presents two major landscapes. The plateau, with a flat or slightly undulated topography, occupies the centre of the region and is dominated by agricultural land (ca. 3.7 million ha). Several mountainous formations surround the plateau where forestry production and bovine livestock constitute the two main economic activities. Human population density is low, 25.29 inhabitants/km2 (www.ine.es), less than one third of the Spanish and EU average (https://ec.europa.eu/eurostat), and depopulation levels have increased over the last decade (6.8% decline in 2012–2022; https://estadistica.jcyl.es). The network of roads is not dense (35.09 km/100 km2; www.ine.es; Fig. 1b), even if this Spanish region has the greatest length of road network per inhabitant. The region is occupied by Iberian wolves (C. lupus signatus), a subspecies of the grey wolf. In this region, wolf hunting was only permitted in lands at the north side of the Duero River (which crosses the region east to west; Fig. 1b) after the implementation of the Habitats Directive (42/2007 law) in Spain in December 2007, which assigned different protection regimes at the north and south of the Duero River. In December 2021, the Spanish Parliament approved the inclusion of the wolf into the List of Wild Species in Regimen of Special Protection (LESRPE), banning wolf killing for the whole Spanish territory, which immediately raised great social and political controversy.

Location of the study region (in grey), Castilla y León (Spain), in southern Europe (a). Map showing hotspots of wildlife-vehicle collisions (b) for the wild boar, the roe deer, the red deer, and the wolf during 2006–2021. Hotspots were drawn using kernel density estimators (built with the ‘ks’ package in R; Duong 2022) where red, orange, and green areas indicate the 25%, 50%, and 75% contour level respectively. Dotted lines show the road network and solid lines show the administrative boundaries for the 9 provinces of Castilla y León. After the implementation of the Habitats Directive in December 2007, wolf hunting was only allowed in lands at the north of the Duero River, represented by the blue line, until December 2021 when hunting was completely banned in the Spanish territory
Data on wildlife-vehicle collisions and hunting bags
We obtained data on WVC events for the period 2006–2021 from official records of traffic accidents registered by traffic authorities of Castilla y León. For each accident, the dataset included information on date, location, type of accident (whether it caused human victims or not) and animal species involved. We compiled the available data on hunting bags for the three ungulates and the wolf from 2004–2005 to 2021–2022 hunting seasons from the annual hunting-season reports of Junta de Medio Ambiente de Castilla y León (https://medioambiente.jcyl.es/). To compare the temporal trends of WVC and hunting bags, we aggregated the number of WVC events per hunting season – which begins in September and lasts until August of the next year – for each study species. Because human activity can affect the probability of animal-vehicle collisions (Fernández-López et al. 2022), we corrected absolute numbers of WVC dividing by annual values of traffic intensity in the region (measured as vehicles/km and downloaded from https://www.transportes.gob.es/) to control for yearly differences in human movements (Figure S1 in supplementary material). We also tried correcting for citizen population density dividing yearly WVC data by human population density (https://estadistica.jcyl.es), obtaining similar results and conclusions.
Statistical analysis
First, we analysed the long-term temporal change of WVC – as a proxy of population abundance – and hunting bags to compare trends and evaluate the potential of recreational hunting to limit population growth of ungulates and wolves over the years. For each study species, we built two generalized linear regression models with Gaussian error distribution and using hunting season as a continuous explanatory variable. One model used the number of WVC – corrected for traffic intensity – as the response variable and data between the hunting seasons 2006–2007 and 2020–2021 (N = 15). The second model used the number of hunted animals as the response variable with data between the hunting seasons 2004–2005 and 2021–2022 (N = 18).
To evaluate whether recreational hunting affected short-term population growth, we calculated the annual growth rate of WVC and hunting bags using the following equations:
Where t in Eqs. 1 and 2 indicates the hunting season. Values of 0 indicate no population growth, values over 0 indicate population increments, and values below 0 indicate population declines. Because wolf’s recreational hunting was banned in the hunting seasons 2018–2019 and 2021–2022, we assigned a value of 1 killed animal to these hunting seasons to obtain finite values of growth rate. For each species, we built generalized linear regression models (with Gaussian error distribution) using the annual growth rate of WVC as response variable and the annual growth rate of hunting bags as the explanatory variable. We built two models for each species: (i) one model regressing the annual growth rate of hunting bags with that of WVC during the same hunting season (data between hunting seasons 2007/2008 to 2020/2021; N = 14), (ii) another model regressing the annual growth rate of hunting bags (hunting seasons 2006/2007 to 2019/2020) with that of WVC during the subsequent hunting season (2007/2008 to 2020/2021) (N = 14). This would allow us to detect immediate effects of hunting on population size or delayed effects when hunting reduces population growth, respectively. We checked model assumptions by inspection of model residuals and model performance using adjusted-R2. All analyses were done with the R software (R Core Team 2023).
Results
Wildlife-vehicle collisions increased from 2,654 to 11,474 accidents in the period 2006–2021, which represents a 332% rise in 16 years. From the 100,806 WVC registered in that period, 91% of accidents were caused by collisions with the three ungulates: 52.6% with roe deer, 32.8% with wild boars, and 5.4% with red deer. Wolves were involved in 192 collisions (0.2%). Hotspots of collisions with these species were concentrated in the east, northeast, and northwest of Castilla y León (Fig. 1b). Of all reported WVC, 1.1% had injured or deceased victims (N = 1,080; increased by 225% in 2006–2021, from 28 accidents to 91), among which 47.2% were caused by collisions with the wild boar, 30.9% with the roe deer, and 7.2% with the red deer.
The number of WVC increased significantly for the three ungulate species (supplementary material Table S1, Fig. 2), with the roe deer showing the steepest increase (426%; Fig. 2c). Traffic accidents caused by collisions with the wild boar and the red deer experienced a lower but steady increase of about 137% and 150%, respectively (Fig. 2a, b). The number of traffic accidents involving wolves showed no significant temporal trend (Table S1; Fig. 3a). Regarding hunting bags, all ungulate species showed significant positive trends (Table S1). The roe deer experienced the highest rate of increase (507%), followed by the wild boar (276%) and the red deer (133%; Fig. 2). The temporal pattern of wolf’s hunting showed no consistent change over time (Table S1). Wolf hunting declined with the implementation of the Habitats Directive in 2007 that allowed wolf hunting only at the north of the Duero River (Fig. 3b). Afterwards, hunting steadily increased, reaching similar numbers to those of the period when the wolf was considered as a game species in the entire region of Castilla y León, until the regional court declared invalid the wolf’s 2016–2019 hunting plan in 2018 (Fig. 3b). Wolf hunting was banned in the Spanish territory in December 2021.

Temporal trends in the number of animals killed by collisions with vehicles (WVC) and by recreational hunting for the wild boar (a, d), the red deer (b, e), and the roe deer (c, f). Numbers are given in thousands for each hunting season. The red solid line shows the slope for the regression against hunting season, which was significant in all models. For clarity, y-axis of WVC graphs provide absolute numbers instead of values corrected for traffic intensity. Note that y-axis scales differ between graphs, and therefore, the steepness of the different slopes are not directly comparable among the figures

Temporal trends in the number of wolves killed by collisions with vehicles (WVC, a) and by recreational hunting (b). The red dashed line shows the non-significant slope for the regression against hunting season. For clarity, y-axis of the WVC graph provide absolute numbers instead of values corrected for traffic intensity
For the roe deer, the red deer, and the wolf we found no effect of the annual growth rate of hunting bags on that of WVC of the same hunting season (roe deer: estimate ± SE = – 0.10 ± 0.63, P = 0.874, adjusted R2 = – 0.08; red deer: estimate ± SE = – 0.41 ± 0.35, P = 0.274, adjusted R2 = 0.02; wolf: estimate ± SE = 0.07 ± 0.10, P = 0.478, adjusted R2 = – 0.04) nor of the next hunting season (roe deer: estimate ± SE = 0.65 ± 0.72, P = 0.385, adjusted R2 = -0.02; red deer: estimate ± SE = 0.14 ± 0.36, P = 0.695, adjusted R2 = – 0.07; wolf: estimate ± SE = – 0.07 ± 0.10, P = 0.487, adjusted R2 = – 0.04) (supplementary material Figs. S2-S7). For the wild boar, however, we found a positive relationship between the annual growth rates of hunting bags and WVC of the same hunting season (estimate ± SE = 1.16 ± 0.34, P = 0.006, adjusted R2 = 0.45; Fig. S8) and a negative association between the annual growth rates of hunting bags and WVC of the following hunting season (estimate ± SE = – 0.90 ± 0.38, P = 0.034, adjusted R2 = 0.27; Fig. S9).
Discussion
Recreational hunting is an important socioeconomic activity and the principal factor of ungulate mortality in Europe (Apollonio et al. 2017; Middleton 2014), which underlies the narrative about its crucial role for regulating ungulate populations and mitigating nuisance. There are studies showing that recreational hunting can indeed reduce ungulate numbers and their impacts (e.g., Hothorn and Müller 2010; Quirós-Fernández et al. 2017). However, our long- and short-term analyses of hunting bags and WVC add to previous evidence manifesting the inability of current models of recreational hunting to prevent or significantly reduce population growth of ungulate species and associated nuisance such as traffic accidents at a large spatial scale (Massei et al. 2015; Melis et al. 2009; Milner et al. 2006).
Despite the increase in hunting bags, the positive temporal trends of vehicle collisions with the wild boar and both deer species indicate that their populations have steadily grown as in many other European regions (Apollonio et al. 2010; Deinet et al. 2013; Massei et al. 2015). Moreover, there is no evidence of a recent slowing down in these temporal trends. Similarly, we did not observe a limiting effect of recreational hunting on the annual growth rate of vehicle collisions with the roe deer and the red deer. However, we found a negative association between the annual growth rate of vehicle collisions in a given year (t) and that of hunting bags in the previous hunting season (t-1) for the wild boar. This result would support a role of recreational hunting in limiting ungulate population growth. However, the positive association observed when using vehicle collisions of the same hunting season indicates that hunting bags increased concomitantly with the population size of this species. Neumann et al. (2020) found that the association between WVC and hunting bag data varied with the spatial scale and the ungulate species (Neumann et al. 2020). Therefore, additional studies should explore the effects of the spatial scale on the relationship of these variables with population density to allow for their reliable use as proxies of species abundance.
Although we cannot rule out that recreational hunting had reduced population growth over the study period (Quirós-Fernández et al. 2017), game management, which is the main source of ungulate mortality (Apollonio et al. 2017), has been insufficient to hold back (no growth) or revert (negative growth) the population increase of these ungulates at regional scale in our study region. This steady increase in traffic accidents caused by collisions with ungulates can raise the risk of road fatality, thus colliding with the United Nations resolution of reducing to half road victims in 2030 (UN General Assembly 2020). However, the resolution of the United Nations does not contemplate the role of wildlife-vehicle collisions in road safety (Ascensão et al. 2021). The socio-economic impacts of overabundant ungulates on road safety, agriculture and timber production, or disease transmission to livestock or humans will likely aggravate as ungulate populations keep growing and human interventions to mitigate damages will become more complex to implement from a monetary, logistic, and societal perspective.
Adapting current models of recreational hunting to achieve management goals could be an option as lethal management is still the main source of ungulate mortality in Europe (Apollonio et al. 2017). However, this strategy encounters several problems. For suppressing population growth, hunters should extract 50–70% of the population (Croft et al. 2020; Keuling et al. 2013) but detailed demographic data are most often lacking (Neumann et al. 2020). Although hunting pressure has intensified in several regions, hunter numbers have been declining (Hagen et al. 2014; Massei et al. 2015; Milner et al. 2006). This process is also happening in our study region, where people with hunting licenses decreased from 160,265 to 117,163 between 2006 and 2022 (27%, https://datosabiertos.jcyl.es/). Further increments in the number of harvested animals per hunter or promotion of selective hunting of females and juveniles – the key groups to effectively reduce population growth (Bieber and Ruf 2005; González-Crespo et al. 2018; Ueno et al. 2010) – might encounter little support from certain hunter profiles due to operational or motivational reasons (Keuling et al. 2016; Vercauteren et al. 2011). On the other hand, lethal management faces increasing societal opposition (Liordos et al. 2017) and the use of lethal control might appear acceptable only under situations of severe ungulate damages (Martínez-Jauregui et al. 2020). When lethal management is approved by authorities, management objectives should be achieved through the application of ecological knowledge.
Additional management actions have been proposed to reduce overabundant populations or alleviate their negative effects. Culling programs can aid in population control by increasing mortality rate and reducing reproductive success when females and juveniles are the targets (Bieber and Ruf 2005; Carpio et al. 2021; Côté et al. 2004; González-Crespo et al. 2018); fertility control together with culling could cause important population reductions in closed populations of wild boar (Croft et al. 2020); supplementary feeding and fencing might be used to mitigate agroforestry damages or traffic accidents (Côté et al. 2004; Milner et al. 2014). However, these actions might convey unintended deleterious effects – enhanced disease transmission or compensatory survival that enhance population growth (Bengsen and Sparkes 2016; Milner et al. 2014) –, and locally limited effectiveness due to complex and costly logistics for their implementation (Bengsen and Sparkes 2016; Côté et al. 2004; Massei and Cowan 2014; McCann and Garcelon 2008).
Management strategies for overabundant ungulates based on recreational hunting seem insufficient under the widespread increments in ungulate abundance, declines in hunter numbers, and declining social acceptance of lethal control of wildlife (Liordos et al. 2017; Massei et al. 2015). The integration of community dynamics into adaptive management, in particular restoring predator–prey interactions, can aid recreational hunting in controlling ungulate numbers and reducing associated impacts (Apollonio et al. 2017; Gilbert et al.2017; Marrotte et al. 2022; Nilsen et al. 2007). Although conditions are nowadays favourable for the recovery of large carnivores in human-dominated landscapes (Chapron et al. 2014), there is still an enormous sociopolitical pressure for the lethal management of wolves, including recreational hunting to limit the increase of population numbers and livestock attacks. In our study region, we did not find a relationship between the annual growth rates of wolf-vehicle collisions and of hunting bags, suggesting that recreational hunting had no effect on the population growth of wolves in the short-term. However, there was no upward trend in wolf-vehicle collisions as observed for ungulates, and wolf's hunting reflected legislative changes in the wolf protection status. With no studies on the relationship between vehicle collision data and wolf abundance, we acknowledge limitations of relying on vehicle collisions as a proxy of wolf population growth due to the high dispersal capacity of this species (Morales-González et al. 2022). Nonetheless, wolves are heavier animals than roe deer and vehicle collisions with a wolf should provoke material damages at least of similar consideration to those caused by roe deer, thus generating police reports for claiming insurance compensations. Therefore, our results should warn about implementing management strategies to control an apparent increment of wolf populations not supported from annual monitoring and under rough population estimates (Prieto et al. 2022; Quevedo et al. 2019), because the use of hunting bags as proxies of population density might not be valid for all species (e.g., Neumann et al. 2020).
Despite the lack of scientific consensus on the relative extent to which predation limits large herbivore abundance (Kuijper 2011; Melis et al. 2009; Ripple and Beschta 2012), lethal management of predators might collide with the goal of controlling overabundant ungulates by limiting predation impact. Wolf harvest can reduce the mortality rate of wild herbivores due to lower wolf numbers, destabilized wolf packs (Fernández-Gil et al. 2016), or lower interactions with ungulates due to altered activity patterns of wolves (Frey et al. 2022). A low density of predators can even result in a negligible impact of predation on ungulate numbers (Gervasi et al. 2012; Sæther 1997). Any lethal management of apex predators requires a good knowledge of population demography, which was not the case for the wolf in Spain and most probably in other countries where the species is present (Creel et al. 2015; Laikre et al. 2022). The role of wolves and other apex predators should be leveraged in mitigating the multifaceted issue of overabundant ungulates because most of the environmental changes that facilitate the increase of large ungulate numbers – mild winters, natural afforestation due to rural abandonment, or the intensification of agriculture – will not revert in the short term and are complex to manage at a large spatial scale.
Availability of data and materials
Data on hunting bags can be accessed by any citizen on the website cited in the manuscript. Data on traffic accidents were provided by Junta de Castilla y León, and hence, we are not allowed to upload it in a repository as we are not the data owners. However, any researcher can contact Spanish traffic authorities to make use of this information.
References
Apollonio M, Andersen R, Putman R (2010) European ungulates and their management in the 21st Century. Cambridge University Press, New-York
Apollonio M, Belkin VV, Borkowski J et al (2017) Challenges and science-based implications for modern management and conservation of European ungulate populations. Mammal Res 62:209–217
Ascensão F, Barrientos R, D’Amico M (2021) Wildlife collisions put a dent in road safety. Science 374:1208
Bautista C, Revilla E, Naves J et al (2019) Large carnivore damage in Europe: analysis of compensation and prevention programs. Biol Conserv 235:308–316
Bengsen AJ, Sparkes J (2016) Can recreational hunting contribute to pest mammal control on public land in Australia? Mammal Rev 46:297–310
Bieber C, Ruf T (2005) Population dynamics in wild boar Sus scrofa: ecology, elasticity of growth rate and implications for the management of pulsed resource consumers. J Appl Ecol 42:1203–1213
Burbaite L, Csányi S (2010) Red deer population and harvest changes in Europe. Acta Zool Lituanica 20:179–188
Carpio AJ, Apollonio M, Acevedo P (2021) Wild ungulate overabundance in Europe: contexts, causes, monitoring and management recommendations. Mammal Rev 51:95–108
Caughley G (1981) Overpopulation. In: Jewell PA, Holst S, Hart D (eds) Problems in Management of Locally Abundant Wild Mammals. Academic Press, New York, USA, pp 7–19
Chapron G, Kaczensky P, Linnell JDC et al (2014) Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 346:1517–1519
Conover MR, Pitt WC, Kessler KK, DuBow TJ, Sanborn WA (1995) Review of human injuries, illnesses, and economic losses caused by wildlife in the United States. Wildlife Soc B 23:407–414
Côté S, Rooney D, Tremblay TP, Dussault C, Waller DM (2004) Ecological impacts of deer overabundance. Annu Rev Ecol, Evol S 35:113–147
Creel S, Becker M, Christianson D et al (2015) Questionable policy for large carnivore hunting. Science 350:1473–1475
Croft S, Franzetti B, Gill R, Massei G (2020) Too many wild boar? Modelling fertility control and culling to reduce wild boar numbers in isolated populations. PLoS One 15:e0238429
Deinet S, Ieronymidou C, McRae L, Burfield IJ, Foppen RP, Collen B, Böhm M (2013) Wildlife comeback in Europe: the recovery of selected mammal and bird species. Final report to Rewilding Europe by ZSL, Birdlife International and the European Bird Census Council. London, UK: ZSL.
Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJB, Collen B (2014) Defaunation in the Anthropocene. Science 345:401–406
Duong T (2022). Ks: kernel smoothing. R package version 1.14.0. https://CRAN.R-project.org/package=k
Eacker DR, Lukacs PM, Proffitt KM, Hebblewhite M (2017) Assessing the importance of demographic parameters for population dynamics using Bayesian integrated population modeling. Ecol Appl 27:1280–1293
Epperly HK, Clinchy M, Zanette LY, McCleery RA (2021) Fear of large carnivores is tied to ungulate habitat use: evidence from a bifactorial experiment. Sci Rep 11:12979
Fernández-Gil A, Naves J, Ordiz A, Quevedo M, Revilla E, Delibes M (2016) Conflict misleads large carnivore management and conservation: brown Bears and Wolves in Spain. PLoS ONE 11(3):e0151541
Fernández-López J, Blanco-Aguiar JA, Vicente J, Acevedo P (2022) Can we model distribution of population abundance from wildlife-vehicle collision data? Ecography 5:e06113
Frey S, Tejero D, Baillie-David K, Burton AC, Fisher JT (2022) Predator control alters wolf interactions with prey and competitor species over the diel cycle. Ecology 8:e08821
Gaynor KM, Brown JS, Middleton AD, Power ME, Brashares JS (2019) Landscapes of fear: spatial patterns of risk perception and response. Trends Ecol Evol 34:355–368
Gervasi V, Nilsen EB, Sand H et al (2012) Predicting the potential demographic impact of predators on their prey: a comparative analysis of two carnivore-ungulate systems in Scandinavia. J Anim Ecol 81:443–454
Gilbert SL, Sivy KJ, Pozzanghera CB, DuBour A, Overduijn K, Smith MM, Zhou J, Little JM, Prugh LR (2017) Socioeconomic benefits of large carnivore recolonization through reduce wildlife-vehicle collisions. Conserv Lett 10:431–439
González-Crespo C, Serrano E, Cahill S, Castillo-Contreras R, Cabañeros L, López-Martín JM, Roldán J, Lavín S, López-Olvera JR (2018) Stochastic assessment of management strategies for a Mediterranean peri-urban wild boar population. PLoS One 13(8):e0202289
Hagen R, Heurich M, Kröschel M, Herdtfelder M (2014) Synchrony in hunting bags: reaction on climatic and human induced changes? Sci Tot Environ 468–469(140):146
Hothorn T, Müller J (2010) Large-scale reduction of ungulate browsing by managed sport hunting. Forest Ecol Manag 260:1416–1423
Jedrzejewska B, Jedrzejewski W (1998) Predation in vertebrate communities. The Białowieza Primeval Forest as a case study. Springer Verlag, Berlin, Germany. https://doi.org/10.1007/978-3-662-35364-6
Jedrzejewski W, Schmidt K, Theuerkauf J, Jedrzejewska B, Selva N, Zub K, Szymura L (2002) Kill rates and predation by wolves on ungulate populations in Bialowieza Primeval Forest (Poland). Ecology 83:1341–1356
Johnson CN, Balmford A, Brook BW, Buettel JC, Galetti M, Guangchun L, Wilmshurst JM (2017) Biodiversity losses and conservation responses in the Anthropocene. Science 356:270–275
Keuling O, Baubet E, Duscher A, Ebert C, Fischer C, Monaco A, Podgórski T, Prevot C, Ronnenberg K, Sodeikat G, Stier N, Thurfjell H (2013) Mortality rates of wild boar Sus scrofa L. in central Europe. Eur J Wildlife Res 59(6):805–814
Keuling O, Strauß E, Siebert U (2016) Regulating wild boar populations is “somebody else’s problem”! – Human dimension in wild boar management. Sci Tot Environ 554–555:311–319
Kuijper DPJ (2011) Lack of natural control mechanisms increases wildlife-forestry conflict in managed temperate European forest systems. Eur J Forest Res 130:895
Laikre L, Allendorf FW, Aspi J et al (2022) Planned cull endangers Swedish wolf population. Science 377:162
Linnell JDC, Cretois B, Nilsen EB, Rolandsen CM, Solberg EJ, Veiberg V, Kaczensky P, Van Moorter B, Panzacchi M, Rauset GR, Kaltenborn B (2020) The challenges and opportunities of coexisting with wild ungulates in the human-dominated landscape of Europe’s Anthropocene. Biol Conserv 244:108500
Liordos V, Kontsiotis VJ, Georgari M, Baltzi K, Baltzi I (2017) Public acceptance of management methods under different human-wildlife scenarios. Sci Tot Environ 579:685–693
Marrotte RR, Patterson BR, Northrup JM (2022) Harvest and density-dependent predation drive long-term population decline in a northern ungulate. Ecol Appl 32:e2629
Martin JL, Chamaillé-Jammes S, Waller DM (2020) Deer, wolves, and people: costs, benefits and challenges of living together. Biol Rev 95:782–801
Martínez-Jauregui M, Delibes-Mateos M, Arroyo B (2020) Addressing social attitudes toward lethal control of wildlife in national parks. Conserv Biol 34:868–878
Massei G, Cowan D (2014) Fertility control to mitigate human-wildlife conflicts: a review. Wildlife Res 41:1–21
Massei G, Kindberg J, Licoppe A et al (2015) Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag Sci 71:492–500
McCann BE, Garcelon DK (2008) Eradication of feral pigs from Pinnacles National Monument. J Wildlife Manag 72:1287–1295
Melis C, Jedrzejewska B, Apollonio M et al (2009) Predation has a greater impact in less productive environments: variation in roe deer, Capreolus capreolus, population density across Europe. Global Ecol Biogeogr 18:724–734
Middleton A (2014) The economics of hunting in europe. Towards a conceptual framework. FACE final report. p 14. https://face.eu/sites/default/files/attachments/framework_for_assessing_the_economics_of_hunting_final_.en_.pdf
Milner JM, Bonenfant C, Mysterud A, Gaillard JM, Csányi S, Stenseth NC (2006) Temporal and spatial development of red deer harvesting in Europe: biological and cultural factors. J Appl Ecol 43:721–734
Milner JM, van Beest FM, Schmidt KT, Brook RK, Storaas T (2014) To feed or not to feed? Evidence of the intended and unintended effects of feeding wild ungulates. J Wildlife Manag 78:1322–1334
Morales-González A, Fernández-Gil A, Quevedo M, Revilla E (2022) Patterns and determinants of dispersal in grey wolves (Canis lupus). Biol Rev 97:466–480
Neumann W, Widemo F, Singh NJ, Seiler A, Cromsigt JPGM (2020) Strength of correlation between wildlife collision data and hunting bags varies among ungulate species and with management scale. Eur J Wildlife Res 66:86
Nilsen EB, Milner-Gulland EJ, Schofield L, Mysterud A, Stenseth NC, Coulson T (2007) Wolf reintroduction to Scotland: public attitudes and consequences for red deer management. P Roy Soc B-Biol Sci 274:995–1002
Nyhus PJ (2016) Human-wildlife conflict and coexistence. Annu Rev Env Res 41:143–171
Pimentel D, Zuniga R, Morrison D (2005) Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol Econ 52:273–288
Prieto A, González V, Barrios L, Palacios F (2022) Field work effort to evaluate biological parameters of interest for decision-making on the wolf (Canis lupus). Hystrix 33:65–72
Pyšek P, Hulme PE, Simberloff D et al (2020) Scientists’ warning on invasive alien species. Biol Rev 95:1511–1534
Quevedo M, Echegaray J, Fernández-Gil A (2019) Lethal management may hinder population recovery in Iberian wolves. Biodivers Conserv 28:415–432
Quirós-Fernández F, Marcos J, Acevedo P, Cortázar C (2017) Hunters serving the ecosystem: the contribution of recreational hunting to wild boar population control. Eur J Wildlife Res 63:57
R Core Team (2023) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Raynor JL (2017) Wolves as a biological control for deer: measuring the indirect economic impacts of predators. Essays on measuring the economic impacts of keystonre species. University of Wisconsin-Madison, pp 1–157
Raynor JL, Grainger CA, Parker DP (2021) Wolves make roadways safer, generating large economic returns to predator conservation. P Natl Acad Sci USA 118:e2023251118
Ripple WJ, Beschta RL (2012) Large predators limit herbivore densities in northern forest ecosystems. Eur J Wildlife Res 58:733–742
Ripple WJ, Estes JA, Beschta RL et al (2014) Status and ecological effects of the world’s largest carnivores. Science 343:1241484
Sáenz-de-Santa-María A, Tellería JL (2015) Wildlife-vehicle collisions in Spain. Eur J Wildlife Res 61:399–406
Sæther BE (1997) Environmental stochasticity and population dynamics of large herbivores: a search for mechanisms. Trends Ecol Evol 12:143–149
Saint-Andrieux C, Calenge C, Bonenfant C (2020) Comparison of environmental, biological and anthropogenic causes of wildlife-vehicle collisions among three large herbivore species. Popul Ecol 62:64–79
Sèbe M, Brion F, Kinds A (2022) Does predation by wolves reduce collisions between ungulates and vehicles in France? Hum Dimens Wildl. https://doi.org/10.1080/10871209.2022.2036391
Simard MA, Dussault C, Huot J, Côté SD (2013) Is hunting an effective tool to control overabundant deer? A test using an experimental approach. J Wildlife Manag 77:254–269
Tanner E, White A, Acevedo P, Balseiro A, Marcos J, Cortázar C (2019) Wolves contributes to disease control in a multi-host system. Sci Rep 9:7940
Ueno M, Kaji K, Saitoh T (2010) Culling versus density effects in management of a deer population. J Wildlife Manag 74:1472–1483
UN General Assembly (2020) Improving global road safety. https://undocs.org/en/A/RES/74/299
Valente AM, Acevedo P, Figueiredo AM, Fonseca C, Torres RT (2020) Overabundant wild ungulate populations in Europe: management with consideration of socio-ecological consequences. Mammal Rev 50:353–366
van Beeck Calkoen STS, Kuijper DPJ, Apollonio M, Blonder L, Dormann CF, Storch I, Heurich M (2023) Numerical top-down effects on red deer (Cervus elaphus) are mainly shaped by humans rather than large carnivores across Europe. J Appl Ecol. https://doi.org/10.1111/1365-2664.14526
Vercauteren KC, Anderson CW, van Deelen T, Drake D, Walter WD, Vantassel SM, Hygnstrom SE (2011) Regulated comercial harvest to manage overabundant white-tailed deer: an idea to consider? Wildlife Soc B 35:185–194
Acknowledgements
We are grateful to B. Arroyo, M. Delibes, and J. Rodriguez for their valuable comments on the manuscript and J. López for his help with databases. We thank Junta de Castilla y León for providing access to data on traffic records and hunting bags. We are grateful to two anonymous reviewers for their constructive comments to the manuscript.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. R.T. is supported by the project ‘CLU-2019–01 – iuFOR Institute Unit of Excellence’ of the University of Valladolid, funded by the Junta de Castilla y León and co-financed by the European Union (ERDF ‘Europe drives our growth’). We declare no conflict of interest.
Author information
Authors and Affiliations
Contributions
FM, JJLL and RT conceived the ideas and designed the methodology; FM and JJLL collected the data; RT analyzed the data and led manuscript writing; All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Ethical approval
Not applicable.
Conflict of interest
We declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tarjuelo, R., Luque-Larena, J.J. & Mougeot, F. Moving towards an ecological management of overabundant ungulates: insights from wildlife-vehicle collisions and hunting bag data. Eur J Wildl Res 70, 51 (2024). https://doi.org/10.1007/s10344-024-01801-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-024-01801-7