
Abstract
Avena sterilis ssp. ludoviciana (Durieu) is a problematic weed in the wheat crop of Australia. Pinoxaden is an effective herbicide for the control of this weed. However, late cohorts of A. ludoviciana escape from the early application of pinoxaden and produce seeds. This research investigated the integrated effect of tillage systems (no-tillage and conventional tillage), seeding rates (100 and 200 seeds m−2), and weed control treatments (nontreated control and pinoxaden application at Z12 and Z33 stages of wheat) on Avena ludoviciana control and wheat yield. The wheat yield remained similar in no-tillage and conventional tillage systems; however, the no-tillage system helped in reducing A. ludoviciana seed production by 28% compared with the conventional tillage system. In the nontreated control, the increased seeding rate of wheat reduced A. ludoviciana biomass and seed production by 33 and 66%, respectively, compared with the low seeding rate. These results suggest that a high seeding rate could be useful in the organic production of wheat. Application of pinoxaden at Z12 and Z33 stages of wheat resulted in an improvement in grain yield by 170 and 150%, respectively, compared with the nontreated control. At both seeding rates, the application of pinoxaden at the Z33 stage of wheat reduced weed seed production by 99% compared with the nontreated control. These results implied that the delayed application of pinoxaden at the Z33 stage of wheat effectively reduced weed biomass and seed production of A. ludoviciana without compromising grain yield as the yield in this treatment was similar to the pinoxaden application at the Z12 stage.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Weeds in crops are an issue for improved crop productivity as they compete with crops for water and nutrients, causing biotic stress. In Australia, two Avena species namely, A. fatua L. (wild oats) and A. sterilis ssp. ludoviciana (Durieu), (hereafter, A. ludoviciana, sterile oats) are the dominant weed species that interfere in winter season crops. These two species, along with A. barbata Pott ex Link (slender oats), collectively cause a loss of about AU$ 28 million per annum to the Australian grain industry when estimated in terms of yield loss and control measures (Llewellyn et al. 2016). Avena ludoviciana is a dominant species in the winter crops of eastern Australia, while A. fatua is dominant in cropping regions of southern and western Australia (Nugent et al. 1999).
The infestation of A. ludoviciana in wheat (Triticum aestivum L.) fields, if left uncontrolled, could cause a yield loss of up to 80% (Martin et al. 1987; Storrie 2007). A recent study conducted in Australia predicted a 50% wheat yield loss at A. ludoviciana density of 16 plants m−2, suggesting that this weed has a strong competitive ability in wheat (Mahajan and Chauhan 2021a). Therefore, control of A. ludoviciana is important in improving wheat productivity in Australia.
In eastern Australia, the no-till wheat production system is common, especially in dryland regions, and has been adopted at a large scale (Llewellyn et al. 2012). The no-till production system helps in residual soil moisture utilization and improves nutrient uptake, as well as improve the soil structure through crop residue retention (Llewellyn et al. 2012). However, weeds remain a problem in no-till production systems due to weed emergence prior to crop emergence, and the low efficacy of pre-emergence herbicides as a result of residue crop cover (Chauhan et al. 2006; Walsh and Powles 2007; Triplett and Dick 2008, Nord et al. 2012). The expansion of the cotton (Gossypium hirsutum L.) industry in eastern Australia has increased the area of conventional tilled irrigated wheat in some pockets as it helps in breaking disease and pest cycles in cotton-based cropping systems (Sykes 2012). Weeds behave differently in response to varied tillage systems (Chauhan et al. 2006, 2012).
Avena ludoviciana could behave differently in no-till and conventional-till production systems due to varied emergence patterns in response to burial depth. It has been found that A. ludoviciana emergence was greater from 2 and 5‑cm soil depths compared with that on the surface (Mahajan and Chauhan 2021b). These results suggest that the emergence of A. ludoviciana could be greater in conventional systems compared with the no-till system. Information on the growth behavior and seed reproduction potential of A. ludoviciana in response to the tillage system is limited in Australia. The ability of A. ludoviciana to produce seeds in a water-stress environment could increase the persistence of this weed in response to varied tillage systems (Sahil et al. 2020). The plants of A. ludoviciana are prolific in seed production. It was estimated that early cohorts (May) of A. ludoviciana could produce up to 4000 seeds plant−1 (Mahajan and Chauhan 2021c). The peak emergence time of A. ludoviciana coincides with the optimum sowing time of wheat (first fortnight of May) in Australia (Mahajan and Chauhan 2021b). These observations suggest that it is essential to control A. ludoviciana before it produces seeds to reduce the weed seed bank in the soil and for improving wheat yield.
In Australia, A. ludoviciana biotypes have evolved resistance to Group 1 and 2 herbicides (Heap 2023). Herbicides belonging to Groups 1 and 2, in general, are applied at a young seedling stage (2–3 leaf stage) for high efficacy and effective control (Beckie et al. 2002). However, growers wait for a greater number of weeds to emerge to obtain maximum weed control in the field. However, by that time weeds become large and difficult to control (Chauhan et al. 2021). This type of situation is very frequent when weeds have multiple cohorts and late emerging weeds escape from timely spray applications (personnel, communication). In Australia, multiple cohorts of A. ludoviciana appeared from March to October (Mahajan and Chauhan 2021b). The staggered emergence pattern of A. ludoviciana could lead to a decision-making process difficult for spraying post-emergent herbicide application.
Pinoxaden is a selective post-emergence herbicide used for controlling annual weeds in winter cereals (Anonymous 2022). It kills weeds by inhibiting acetyl-CoA carboxylase (ACCase) (Hofer 2006, Bitarafan and Andreasen 2020). Many weeds have evolved resistance to pinoxaden in more than 15 countries (Heap 2023). Large weed plants may build up resistance to herbicides with time, even when applied at an optimal rate, due to poor weed control (Botterman and Leemans 1988). Therefore, the stage of the plant is very critical for effective control during the spray application of herbicides. We hypothesized that late application of pinoxaden in wheat (Z33 stage: third node formation) may provide complete control of early and late cohorts of A. ludoviciana.
Exploring crop competition for weed suppression is found to be an effective cultural strategy for weed control in Australian cropping regions (Lemerle et al. 1996; Scott et al. 2013). Crop competitiveness against weeds can be increased by using a high seeding rate (Mahajan and Chauhan 2022). We hypothesized that the competitiveness of a crop in response to a tillage system can be increased by using a high seeding rate and that competitiveness can also be exploited for reducing the seed production of A. ludoviciana without using any herbicides. Increased crop competition using a high seeding rate in wheat may suppress weeds without using any herbicides and promote organic wheat.
This research aimed to answer two questions: (i) can an increase in seed rate of wheat in different tillage systems suppress A. ludoviciana, reduce its seed number, and improve wheat productivity; and (ii) can a late application of pinoxaden provide effective control of A. ludoviciana without compromising grain yield.
Materials and Methods
A 2-yr field study was conducted at the Research Farm of the University of Queensland, Gatton (27.5514° S and 152.3428° E), Australia. In each year (2020 and 2021), the experiment was conducted in the winter season (May-October). The experimental location belongs to the subtropical climatic region of Australia and has an average annual rainfall of 728 mm. The soil in the experimental field was clay with a pH of 6.9 and 1.4% organic matter. The experimental location was the same in both years. Experimental plots in conventional tillage treatments were tilled (12–15 cm deep) with a disc-harrow (two times) followed by a rotovator operation. The field was kept fallow after the first wheat crop.
Experiment Design and Herbicide Treatments
The experiment was arranged in a split-split plot design with two tillage regimes (no and conventional tillage) in main plots, two seeding rates (100 and 200 seeds m−2) in sub-plots, and three weed control treatments (nontreated control, pinoxaden application at Z12 (two-leaf stage), and Z33 stage (third node formation) of wheat) in sub-sub plots. Wheat (variety Spitfire) was sown using a cone planter (six rows) at a row spacing of 35 cm in both years. The crop was sown on 7th May in both years.
All treatments were replicated thrice each year, and the size of each plot was 4.0 m (length) × 1.4 m (width). Avena ludoviciana seeds at a rate of 40 kg ha−1 were mixed with sand and broadcasted before wheat planting and tillage operation for ensuring uniform weed infestation across the field each year. Pinoxaden spray was done with a CO2 backpack sprayer equipped with four flat-fan nozzles (Airmix 110015 TeeJet nozzles, Model 25611) spaced at 50 cm and using a volume of 100 L of water ha−1 at 200 kPa. Pinoxaden 100 EC (Axial = active constituent; 100 g L−1 pinoxaden +25 g L−1 cloquintocet-mexyl; Syngenta, Australia) was applied at a rate of 20 g ai ha−1. To create the herbicide solution, pinoxaden was mixed with Adigor adjuvant at 500 ml per 100 L water.
The crop was harvested on 14th and 7th October in 2020 and 2021, respectively. At crop harvest, A. ludoviciana density, biomass, and seed number m−2 were determined using a quadrat (50 cm × 50 cm) placed randomly in each plot at two places. Weeds from the quadrat area in each plot were counted and converted into plants m−2. For biomass sampling, weeds were removed from the base level in each plot, placed in paper bags, and then dried in an oven at 70 ℃ for 72 h. After oven drying, weed samples were weighed for biomass. For weed seed count, florets (empty and nonempty) of each A. ludoviciana’s panicle were counted that occurred in the quadrat area. In both years, the crop was harvested when it attained maturity using a combine harvester. For effective tillers, tillers bearing earheads were counted from two places in each plot by placing a ruler of 1 m length randomly and converted into plants m−2. Grain yield was recorded from a net area of 4.2 m2 (3.0 m × 1.4 m) per plot and converted to t ha−1 at a 12% moisture content.
Statistical Analyses
The results of the analysis of variance (ANOVA) for each parameter presented an overall picture of the relative effects of years, tillage, seeding rates, weed control treatments, and all possible interactions on A. ludoviciana and wheat (Table 1). In a combined analysis of data, parameters where the interactions of years × treatments were nonsignificant, data were pooled over the years.
Data were analyzed using the software CPCS1 developed by Punjab Agricultural University (www.pau.edu), Ludhiana, India (Mahajan and Chauhan 2022) and verified with GenStat 21st edition (VSN International, Hemel Hempstead, UK). Before ANOVA, data were also validated for meeting the assumptions of normality and equal variance. Treatment means were separated with the use of Fisher’s Protected LSD test. Unless indicated otherwise, after ANOVA, means were separated using LSD at P = 0.05.
Results
Weather Conditions
The crop received a higher amount of rainfall in May and July of 2021 (May: 90 mm and June: 82.6 mm) compared with 2020 (Fig. 1; Mahajan and Chauhan 2022,). In both years, July had the lowest mean monthly maximum temperature (21.7 ℃ for 2020 and 20.5 ℃ for 2021). The mean monthly minimum temperature for June and July 2021 was lower than in 2020. In 2020, the mean monthly minimum temperature was lowest in August (6.8 ℃); however, in 2021, it was lowest in July (7.1 ℃). These observations suggest that the early season of 2021 (July) was more congenial for the emergence of A. ludoviciana than in 2020 due to the occurrence of higher rainfall and more favorable temperatures, as supported by a recent study conducted in Australia revealed that those conditions in the winter season proved to be a catalyst for the emergence of A. ludoviciana (Mahajan and Chauhan 2021b).

Weather parameters (mean monthly maximum and minimum temperature, and rainfall) recorded at Gatton, Queensland, in 2020 (a) and 2021 (b) during the growth duration of wheat crops (Mahajan and Chauhan 2022)
Weed Biomass and Weed Seed Production
For weed (A. ludoviciana) biomass, the interaction between the seeding rate and the weed control treatment was found to be significant and all other interactions were found to be nonsignificant (Table 1). In the nontreated control, the high seeding rate reduced the biomass of A. ludoviciana by 33% compared with the low seeding rate (Table 2). In herbicide-treated plots, the seeding rate did not influence weed biomass. Averaged over tillage treatments, at the low seeding rate, weed biomass in the nontreated plot was 508 g m−2, and this biomass was reduced by 89 and 99% with pinoxaden application at Z12 and Z33 stages, respectively (Table 2). Averaged over tillage treatments, at the high seeding rate, weed biomass in the nontreated plot was 340 g m−2, and this weed biomass was reduced by 95 and 99% with pinoxaden application at Z12 and Z33 stages, respectively (Table 2).
Seed production of A. ludoviciana followed a similar trend to biomass as the interaction between seeding rate and weed control treatment was significant (Table 2). In the nontreated control, the use of the high seeding rate reduced weed seed production by 66% compared with the low seeding rate (Table 2). Application of pinoxaden at Z12 and Z33 stages of wheat grown at the low seeding rate reduced weed seed production by 89 and 99%, respectively, compared with nontreated control (3500 seeds m−2) (Table 2). Similarly, at the high seeding rate, application of pinoxaden at Z12 and Z33 stages of wheat reduced weed seed production by 88 and 98%, respectively, compared with the nontreated control (1200 seeds m−2) (Table 2).
Averaged over seeding rate and weed control treatments, weed biomass was similar in both tillage systems. However, seed production of A. ludoviciana was reduced by 28% in the no-tillage system compared with the conventional tillage system (Fig. 2).

Avena ludoviciana seed production (number m−2) in response to tillage treatments (Averaged over seeding rate and weed control treatments). Error bars indicate the least significant differences (LSD) at a 5% level of significance
Grain Yield
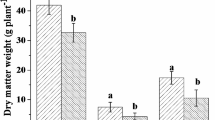
All interactions (two-way and three-way) between tillage, seeding rate, and weed control treatments for grain yield were found to be non-significant. Averaged over seeding rate and weed control treatments, grain yield was similar (4.7–5.1 t ha−1) in both tillage systems (data not shown). Averaged over tillage and weed control treatments, the grain yield at both seeding rates was also similar (4.7–5.1 t ha−1, data not shown). Averaged over tillage and seeding rate, wheat yield in the nontreated control plot was 2.1 t ha−1 and it increased by 171 and 148% with pinoxaden application at Z12 and Z33 stages, respectively, compared with nontreated control (Fig. 3).

a Grain yield (t ha−1) and b effective tillers (number m−2) and of wheat in response to weed control treatments (Averaged over tillage treatments). Error bars indicate the least significant (LSD) differences at a 5% level of significance. Z12 (two-leaf stage), and Z33 stage (third node formation) of wheat
A similar trend was also observed for effective tillers m−2 (Fig. 3). Averaged over tillage and seeding rate, effective tillers m−2 in the nontreated control plot were found to be 131 and it increased by 232 and 227% with pinoxaden application at Z12 and Z33 stages, respectively, compared with nontreated control (Fig. 3).
Discussion
This research demonstrated that wheat yield did not increase with an increased seeding rate, irrespective of tillage systems. However, an increased seeding rate helped in reducing weed biomass and weed seed production, especially in the nontreated control treatment. This information could be useful in making strategies for weed control in the organic production of wheat. Effective weed control through increasing seeding rate is a proven cultural weed management practice and is exploited in many crops (Gill and Holmes 1997; Lemerle et al. 2004). The concept behind the increased seeding rate in different crops is to increase the competitive ability of crops against weeds by attaining an early canopy closure (Lemerle et al. 1995; Mahajan and Chauhan 2022). The manipulation of this cultural practice is very useful where weeds look phenotypically similar (Lemerle et al. 2001, Cousens 1996). Thickening of a crop canopy through increased seeding rates was traditionally practiced to control weeds when high tillering varieties and herbicides were not available (Downing 1921).
Previous studies in Australia also reported the effect of increased wheat planting density on the suppression of A. ludoviciana (Radford et al. 1980). The authors found a negative correlation between wheat density and A. ludoviciana biomass (Radford et al. 1980). Reduced biomass of Lolium rigidum Gaudin (rigid ryegrass) was observed when the density of lupin was increased (Lupinus spp.), which resulted in an increase in yield (Allen 1977; Medd et al. 1985). Similarly, the effect of increased wheat density on L. rigidum suppression and improved wheat yield was also observed in Western Australia (Hashem et al. 1998). In the current study, weed seed production decreased with an increased seeding rate. Likewise, previous workers also reported that increasing the seeding rate of wheat from 50 to 200 kg ha−1 helped in reducing the seed production of L. rigidum (Fee and Anderson 1997).
The current study also demonstrated that yield remained similar when pinoxaden was applied at the Z12 and Z33 stages of wheat. However, the reduction in magnitude for weed biomass and weed seed production was higher when pinoxaden was applied at the Z33 stage compared with the nontreated control. These observations suggest that pinoxaden may be applied at a late stage (Z33) of the crop and could effectively reduce weed seed reduction without compromising grain yield. Late application of pinoxaden could help in controlling early and late cohorts of weeds as A. ludoviciana emerges in multiple cohorts. In addition to this, a delayed application of pinoxaden in wheat (ZCK31-32) caused a biomass reduction (60%) of Alopecurus myosuroides Huds (Pintar et al. 2021).
It is well-known fact that younger weeds are more sensitive to herbicides than those in more advanced stages (Kudsk 2017). However, this phenomenon depends on many factors, such as weed growth rate, type of weeds, herbicides, and environmental conditions. Pinoxaden causes inhibition of ACCase enzyme and results in chlorosis of leaves within a week after application followed by necrosis and death of rapidly growing meristematic tissue. Pinoxaden kills the susceptible plants (Avena spp., Phalaris spp., Lolium spp., etc.) completely within two to three weeks after application (Hofer 2006; Bitarafan and Andreasen 2020; Anonymous 2022).
Weed control strategies that prevent flowering and reduce the seed production potential of weeds may help in reducing the soil weed seed bank and enable fewer weed problems in the future. Avena ludoviciana plants have high seed-shattering ability (Mahajan and Chauhan 2021a), therefore, late application of pinoxaden may provide effective control of early and late cohorts before producing seeds. The current study also suggested that the no-till system helps in reducing weed seeds per unit area. A previous study revealed that A. ludoviciana seeds decayed fast on the soil surface and its emergence was lower on the soil surface (Mahajan and Chauhan 2021b). This could be the reason for reduced weed infestation in no-till plots compared with the conventional tilled plots, and ultimately lower weed seed production per unit area. Several studies reported a faster decline in the seed bank of Avena spp. in no-till compared with the conventional till system (McGillion and Storrie 2006; Nugent et al. 1999; Osten et al. 2007). It was postulated that the germination of Avena spp. was promoted with tillage operations (Chancellor 1976).
Conclusions
An increased seeding rate of wheat under weedy situations helped in reducing A. ludoviciana biomass and weed seed production, which resulted in a significant improvement in the yield. This information could be useful in the organic production of wheat. Wheat yield under no-till and conventional tilled systems remained similar; however, the no-till production system helped in reducing weed seed production. This information suggests that the no-till system had an added advantage of weed control. The application of pinoxaden at the Z33 stage (third node formation) helped in controlling early and late cohorts of A. ludoviciana without compromising the grain yield.
References
Allen JM (1977) Weeds in grain lupins.1. The effect of weeds on grain lupin yields. Aust J Exp Agric 17:112–116
Anonymous (2022) Axial 50 EC herbicide label. https://www.syngenta.co.zm/product/crop-protection/axial-050-ec. Accessed 25 Mar 2022
Beckie HJ, Thomas AG, Stevenson FC (2002) Survey of herbicide-resistant wild oat (Avena fatua) in two townships in Saskatchewan. Can J Plant Sci 82:463–471
Bitarafan Z, Andreasen C (2020) Seed production and retention at maturity of blackgrass (Alopecurus myosuroides) and silky windgrass (Apera spica-venti) at wheat harvest. Weed Sci 68:1–25
Botterman J, Leemans J (1988) Engineering of herbicide resistance in plants. Biotechnol Genet Eng Rev 6:321–340
Chancellor RJ (1976) Seed behaviour. In: Jones DP (ed) Wild oats in world agriculture. Agric. Res. Counc., London, pp 65–87
Chauhan BS, Gill GS, Preston C (2006) Tillage system effects on weed ecology, herbicide activity and persistence: a review. Aust J Exp Agric 46:1557–1570
Chauhan BS, Singh RG, Mahajan G (2012) Ecology and management of weeds under conservation agriculture: a review. Crop Prot 38:57–65
Chauhan BS, Congreve M, Mahajan G (2021) Management options for large plants of glyphosate-resistant feather fingergrass (Chloris virgata) in Australian fallow conditions. PLoS ONE 16(12):e261788
Cousens RD (1996) Comparative growth of wheat, barley, and annual ryegrass (Lolium rigidum) in monoculture and mixture. Aust J Agric Res 47:449–464
Downing RG (1921) Thick or thin seeding for wheat? Agricultural Gazette of N.S.W., p 205
Fee C, Anderson WK (1997) Crop management systems to Improve the Competitiveness of wheat against weeds and reduce the reliance on herbicides. GRDC Project DAW363W Report (Agriculture Western Australia)
Gill GS, Holmes JE (1997) Efficacy of cultural control methods for combating herbicide-resistant Lolium rigidum. Pestic Sci 51:352–358
Hashem A, Radosevich SR, Roush ML (1998) Effect of proximity factors on competition between winter wheat (Triticum aestivum) and Italian ryegrass (Lolium multiflorum). Weed Sci 46:181–190
Heap IM (2023) International survey of herbicide resistant weeds. https://www.weedscience.org/Home.aspx. Accessed 10 Jan 2023
Hofer U (2006) Pinoxaden: For broad spectrum grass weed management in cereal crops. J Plant Dis Prot 20:989–995
Kudsk P (2017) Optimising herbicide performance. In: Hatcher PE, Froud-Williams RJ (eds) Weed research: expanding horizons, 1st edn. Wiley, Oxford, pp 149–179
Lemerle D, Verbeek B, Coombes N (1995) Losses in grain yield of winter crops from Lolium rigidum competition depend on crop species, cultivar and season. Weed Res 35:503–509
Lemerle D, Verbeek B, Cousens RD, Coombes NE (1996) The potential for selecting wheat varieties strongly competitive against weeds. Weed Res 36:505–513
Lemerle D, Gill GS, Murphy CE, Walker SR, Cousens RD, Mokhtari S, Peltzer SJ, Coleman R, Luckett DJ (2001) Genetic improvement and agronomy for enhanced wheat competitiveness with weeds. Crop Past Sci 52:527–548
Lemerle D, Cousens RD, Gill GS, Peltzer SJ, Moerkerk M, Murphy CE, Collins D, Cullis BR (2004) Reliability of higher seeding rates of wheat for increased competitiveness with weeds in low rainfall environments. J Agric Sci 142:395–409
Llewellyn RS, D’Emden FH, Kuehne G (2012) Extensive use of no-tillage in grain growing regions of Australia. Field Crop Res 132:204–212
Llewellyn R, Ronning D, Ouzman J, Walker S, Mayfield A, Clarke M (2016) Impact of weeds on Australian grain production: the cost of weeds to Australian grain growers and the adoption of weed management and tillage practices. Report for GRDC, CSIRO Australia.
Mahajan G, Chauhan BS (2021a) Interference of wild oats (Avena fatua L.) and sterile oats (Avena ludoviciana Durieu) in wheat (Triticum aestivum L.). Weed Sci 69:485–491
Mahajan G, Chauhan BS (2021b) Seed longevity and seedling emergence behavior of wild oat (Avena fatua) and sterile oat (Avena sterilis ssp. ludoviciana) in response to burial depth in eastern Australia. Weed Sci 69:362–371
Mahajan G, Chauhan BS (2021c) Biological traits of six sterile oat biotypes in response to planting time. Agron J 113:42–51
Mahajan G, Chauhan BS (2022) Improving wheat productivity through integrated management of Avena ludoviciana. Agron J. https://doi.org/10.1002/agj2.21251
Martin RJ, Cullis BR, McNamara DW (1987) Prediction of wheat yield loss due to competition by wild oats (Avena spp.). Aust J Agric Res 38:487–499
McGillion T, Storrie AE (2006) Integrated weed management in Australian cropping systems—a training resource for farm advisors. CRC for Australian Weed Management, Adelaide
Medd RW, Auld BA, Kemp DR, Murison RD (1985) The influence of wheat density and spatial arrangement on annual ryegrass, Lolium rigidum Gaudin, competition. Aust J Agric Res 36:61–371
Nord EA, Ryan MR, Curran WS, Mortensen DA, Mirsky SB (2012) Effects of management type and timing on weed suppression in soybean no-till planted into rolled-crimped cereal rye. Weed Sci 60:624–633
Nugent T, Storrie A, Medd R (1999) Managing wild oats. CRC for Weed Management Systems and Grains Research and Development Corporation. https://archive.lls.nsw.gov.au/__data/assets/pdf_file/0004/495346/archive-wild_oats.pdf. Accessed 10 Jan 2023
Osten VA, Walker SR, Storrie A, Widderick M, Moylan P, Robinson GR, Galea K (2007) Survey of weed flora and management relative to cropping practices in the north-eastern grain region of Australia. Aust J Exp Agric 47:57–70
Pintar A, Svečnjak Z, Šoštarčić V, Lakić J, Barić K, Brzoja D, Šćepanović M (2021) Growth stage of Alopecurus myosuroides huds. determines the efficacy of pinoxaden. Plants 10:732
Radford BJ, Wilson BJ, Cartledge O, Watkins FB (1980) Effect of wheat seeding rate on wild oat competition. Aust J Exp Agric 20:77–81
Sahil, Mahajan G, Loura D, Raymont K, Chauhan BS (2020) Influence of soil moisture levels on the growth and reproductive behaviour of Avena fatua and Avena ludoviciana. PLoS ONE 15(7):e234648
Scott BJ, Martin P, Riethmuller GP (2013) Row spacing of winter crops in broad scale agriculture in Southern Australia. Graham Centre Monograph, vol 3. NSW Department of Primary Industries, Orange
Storrie A (2007) ‘Wild oat resistance options’, Grains Research Update—Northern region, Grains Research & Development Corporation. http://users.tpg.com.au/icanadsl/newsletters/NL37V4.pdf. Accessed 10 Jan 2023
Sykes J (2012) Irrigated wheat—Best practice guidelines in cotton farming systems. Cotton Catchment Communities CRC and Grains Research & Development Corporation
Triplett GB Jr, Dick WA (2008) No-tillage crop production: A revolution in agriculture! Agron J 100:153
Walsh MJ, Powles SB (2007) Management strategies for herbicide-resistant weed populations in Australian dryland crop production systems. Weed Technol 21:332–338
Funding
This research was funded by GRDC with grant number US00084.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions
Author information
Authors and Affiliations
Contributions
Conceptualization: B.S. Chauhan, G. Mahajan; Formal analysis: G. Mahajan; Funding acquisition: B.S. Chauhan; Investigation: G. Mahajan and B.S. Chauhan and; Methodology: G. Mahajan and B.S. Chauhan; Project administration: B.S. Chauhan; Writing—original draft: G. Mahajan; Review & editing: B.S. Chauhan. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
G. Mahajan and B.S. Chauhan declare that they have no competing interests.
Additional information
Data Availability Statement
All relevant data are within the manuscript.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mahajan, G., Chauhan, B.S. Avena sterilis ssp. ludoviciana (Durieu) Control in Wheat Through Integration of Tillage, Seeding Rate, and Herbicide Application. Gesunde Pflanzen 75, 2337–2344 (2023). https://doi.org/10.1007/s10343-023-00909-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10343-023-00909-1