
Abstract
The frequent outbreaks of European spruce bark beetle Ips typographus (L.) have destroyed huge amounts of Norway spruce Picea abies (L.) forests in central and Northern Europe. Identifying the risk factors and estimating the damage level is important for strategic damage control. The risk factors of forest damage by spruce bark beetles have mostly been analyzed on the landscape scale, while the in-stand risk factors have been less investigated. This study aims at exploring the local-scale risk factors in a flat area with spruce-dominated forest in southern Sweden. The investigated factors include four abiotic factors, i.e., soil wetness, solar radiation, slope gradient, and aspect, and three biotic factors, i.e., the number of deciduous trees and trees that died from attacks in previous years that remained (TreesLeft) and removed (TreesRemoved) from the forest stand. We put up 24 pheromone bags in six stands attacked by bark beetle in the previous years, resulting in different numbers of infested trees in each plot. We explored in which microenvironment a pheromone bag resulted in more colonization, the impact radius of each factor, and the necessary factors for a risk model. The environmental factors were obtained from remote sensing-based products and images. A generalized linear model (GLM) was used with the environmental factors as the explanatory variables and the damage levels as the response variables, i.e., the number of attacked trees for the plot scale, and healthy/infested for the single-tree scale. Using 50 m and 15 m radius of the environmental factors resulted in the best fit for the model at plot and individual tree scales, respectively. At those radii, the damage risk increased both at plot and individual tree level when spruce were surrounded by more deciduous trees, surrounded by dead trees that had been removed from the forest, and spruces located at the north and east slopes (315°–135° of aspect, > 2° slope). Soil wetness, solar radiation, and remaining standing dead trees in the surrounding did not significantly impact the damage level in the microenvironment of the study area. The GLM risk model yielded an overall accuracy of 0.69 in predicting individual trees being infested or healthy. Our efforts to investigate the risk factors provide a context for wall-to-wall mapping in-stand infestation risks, using remote sensing-based data.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The European spruce bark beetle Ips typographus (L.) is a native species to Europe and is important for the natural dynamics of the forest, with a positive impact on biodiversity. However, driven by global warming, the bark beetle population has been increasing with increasing air temperatures, and the forest has been weakened by the shifting precipitation patterns and frequent windstorms (Senf et al. 2021; Schuldt et al. 2020), triggering bark beetle outbreaks at an unprecedented intensity across Europe. It causes mortality of Norway spruce Picea abies (L.) trees, a major resource for the European timber industry (Hlásny et al. 2021), and causing severe ecological and economic damage. In recent years, it has been the most devastating and aggressive insect species to the forest across Eurasia. During outbreaks in 2018–2022, Sweden reported a 32 million m3 timber loss, while in the Czech Republic, it has been estimated that 260 million EUR of state interventions were needed to compensate for decreased timber prices, excessive workloads, and other cascading effects (Hlásny et al. 2021). It is projected that, by 2030, climate change in Europe will have exacerbated bark beetle disturbances sevenfold compared to 1971–1980 (Seidl et al., 2014). Identifying forests with a high risk of bark beetle damage is important for prevention measures of financial loss, as the forest owners can harvest the forest before infestations to save the timber value. Understanding the risk factors is also crucial for damage control, with a management adapted to the new conditions under climate change.
Several climate conditions at a large-scale have been shown to aggravate forest damage caused by bark beetles. The warmer temperature is one of the major causes. Temperature conditions under the bark determine the developmental time from egg to imago, and higher temperatures generally shorten the time and increase the beetle population (Wermelinger 2004). The bark beetle has a one-year life cycle and usually develops 1–2 generations in Nordic counties and 2–3 generations in Central Europe. With a limited population, the bark beetle attacks newly dead or dying spruce and unbarked timber, while with a larger population, the bark beetle also kills growing trees. Based on the temperature sum, forecasts of beetle swarming can be made on yearly basis, and predictions of the damage trends can be made for the coming years (Mezei et al. 2017b). Precipitation also impacts the forest damage level by influencing the vitality of the forest (Kautz et al. 2022). Spruce trees can usually fight back, during the beginning of colonization, e.g., by releasing resin and defensive chemicals (Wermelinger 2004). The defense can be weakened and lead to successful colonization and tree mortality, due to low tree vitality under drought stress. Therefore, low precipitation can also lead to severe forest damage, e.g., the extremely dry summer in 2018 weakened the spruce forest and resulted in huge beetle damage in the following year in Sweden. In addition, other extreme weather events such as windstorms and ice storms also have a strong correlation with the bark beetle outbreaks (Groot et al. 2018; Okland and Berryman, 2004).
Such climate data can be used to forecast a general trend for the coming years and at a large-scale (Groot and Ogris 2019; Ogris and Jurc 2010). However, for local damage management aiming at in-time sanitary cutting and timber-value saving, a small scale risk analysis is also urgently needed, to precisely locate forests with high risk. For such purposes, climate data such as temperature and precipitation contribute less to the risk model, since evaluated forests are often located in a similar climate environment. Therefore, surrogate data with more variation at a small scale can be used, and are often correlated with temperature and precipitation. For example, solar radiation is highly related to temperature conditions, but has more variation at a small scale caused by topography. Studies have used solar radiation as an explanatory variable to predict the beetle damages, and found that stronger solar radiation increases the damage level (Mezei et al. 2014b, 2019). Soil condition and topography have also been used to predict beetle damage (Ogris and Jurc 2010; Mezei et al. 2014b; Blomqvist et al. 2018), considering their correlation with the water regime in the forest.
Besides abiotic factors, biotic factors such as the forest characteristics, structure, and diversity also influence the damage risk. Bark beetles prefer mature spruce with a certain range of diameters at the breast height (DBH) than young spruce (Wermelinger 2004). Some studies, showed that forest stands with older trees (Overbeck and Schmidt 2012) and larger DBH of spruce were more exposed to beetle attacks (Mezei et al. 2014b), while other studies (Blomqvist et al. 2018; Sproull et al. 2015) also exhibited the largest trees, e.g., DBH > 60 cm, which were less prone to be attacked. At the landscape level, forests located in heterogeneous landscapes and with high tree species diversity were observed with fewer beetle damages (Kärvemo et al. 2016). Once attacked by the beetles, a forest tends to be attacked in the following years by the beetles overwintering in the soil or under the bark, while this process is also influenced by management measures. For example, one study showed that salvage logging lowered beetle infestations two years after the execution, while sanitation felling only marginally reduced the beetle population (Havašová et al. 2017). Another study showed that sanitation felling and salvage logging reduce the emergence of new infestation spots (Stadelmann et al. 2013). However, studies have also shown that sudden exposure of the stand after harvesting predisposed forest edges to bark beetle infestations (Marešová et al. 2020; Kautz et al. 2013). Therefore, the impact of previous infestations removed or left in forests needs to be further explored.
So far, studies analyzing the damage risks of beetle attacks have mainly focused on a large-scale, and the smallest scale of the risk analysis, so far is at the stand level (Blomqvist et al. 2018). Therefore, in-stand scale factors can be explored to improve the resolution of early warning of beetle damages. We used existing data from a field experiment, where pheromone bags were used to initiate the bark beetle attacks. The assumption was that the environmental factors influenced the amount of infestations, i.e., influenced the damage risk. This study investigated the risk factors at the within-stand level using pheromone bags to attract bark beetles, addressing the following scientific questions.
-
(1)
Which environmental factors result in more colonization?
-
(2)
What is the impact radius of each factor?
-
(3)
Which factors are needed for modeling the damage risk?
This study mainly focused on a limited number of factors, although many other factors may also influence the risk. The investigated factors were A: forest conditions in the surrounding, including the number of deciduous trees, the number of previously attacked trees left in the forest, and the number of previously attacked trees removed from the forest, and B: abiotic environmental factors including soil wetness, solar radiation, slope gradient, terrain aspect. These factors were selected based on previous studies mentioned, above showing their potential influence on the beetle population and tree vulnerability. These factors are also chosen, because the data can be obtained from remote sensing data and wall-to-wall risk maps can potentially be made using the influencing factors.
Materials and methods
Study area and field observations
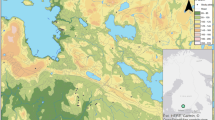
The study area is in Remningstorp (58°27′18″N, 13°39′8″E, Fig. 1a), Sweden. It is a hemi-boreal forest estate covering 1602 ha, dominant by Norway spruce and Scotch pine Pinus sylvestris (L.) managed for wood production. In the study area, bark beetle attack has occurred in some stands.

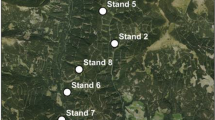
The study area in Remningstorp, Sweden (a), stands and plots (b), a pheromone bag (c) and a trap for bark beetles (d) on the central tree of a plot
In this study, we attached pheromone bags in the forest to attract beetles, and analyzed the environmental factors that influence the damage levels resulting from the pheromone bags. In the spring, six mature stands were selected based on having spruce > 80% of the total volume, and having attacks from previous years (Fig. 1b). These six stands were at least 600 m away from each other and can be considered to be spatially uncorrelated. Four plots with 15 m radius were set up in each stand, with 71 m average distance to the nearest neighbor plot. The locations of the plots were selected to have about 20–50 m distance to the previous attacks, and avoiding other tree species and dead trees in the plots as much as possible. The DBH and species were registered in the field, and the locations of the single trees were measured using a PosTex system, an instrument using ultrasound to position the objects.
A pheromone bag was attached to the central tree of a plot to attract the bark beetles (Fig. 1c). The pheromone bags contained Phero-X-Lure IT simulating the aggregation of pheromones emitted by bark beetles. They were attached to the tree stem 2 m above the ground, on the south side of the trunk. The pheromone bags remained on the trees from week 16 (April 5, before the first swarm) to week 22 (May 31). Below each pheromone bag, a flight interception trap was mounted. The traps were made from a vertical piece of transparent plastic with a container below with a mix of glycol, water, and some detergent (we used windshield washer fluid) to lower the surface tension (Fig. 1d). The flight interception traps helped to monitor the swarm and identify the attacks, and they were mounted in the same way in every plot. The first swarm occurred in week 20 (May 17) and the pheromone bags were removed two weeks after the swarming so the parental beetles could attack other trees. The second swarm occurred at week 30 (July 27). The field monitoring of the health status was conducted weekly from the middle of April to the middle of August and one time in the middle of October. The recorded symptoms included resin flow, holes in the bark, bark damage, discoloration and defoliation. Trees with these symptoms were defined as damaged trees. Additional information about the experiment and field data collection are available in (Huo et al. 2023). In the following sections, the center trees with pheromone bags attached were excluded from the analysis, as they were infested regardless of the predisposition (Table 1).
Environmental factors and remote sensing data
In this study, two types of factors were considered. We aimed to examine the influences from trees in the surroundings, so-called biotic factors, including deciduous trees, and previously attacked trees left or removed from the forest. We also aimed to examine the impact of soil wetness, solar radiation, and terrain condition, so-called abiotic factors. The factors were:
-
The number of deciduous trees in the surrounding.
-
The number of previously attacked trees left in the forest (denoted as TreesLeft) in the surrounding.
-
The number of previously attacked trees removed from the forest (denoted as TreesRemoved) in the surrounding.
-
Soil wetness (1-dry, 2-moist, 3-wet)
-
Solar radiation (kWh/m2)
-
Slope gradient (degree)
-
Aspect (north 315°–45°, east 45°–135°, south 135°–225°, west 225°–315°)
The locations of deciduous trees and previously attacked trees were visually interpreted from drone images in May, June, August, and October 2021 and Pleiades satellite images in 2020-09-18 and 2019-07-17. The drone images had 0.06 m resolution and the Pleiades images had 0.5 m resolution. We used the true color, i.e., red, green, blue bands, for interpreting both images. We identified deciduous trees that were gray in May, green in June and August, and yellow in October. We identified TreesLeft that were green in the Pleiades images and gray in all drone images. We identified TreesRemoved that were green in the Pleiades in 2019, gray in the Pleiades in 2020, and missing in drone images in 2021. Some of the TreesRemoved were removed before May 2020, and others were removed from May to June 2021. The number of deciduous trees, TreesLeft and TreesRemoved surrounding each plot and each tree were then calculated with different radii. The used radii were denoted as Rbiotic.
The soil wetness map was provided by the Swedish University of Agricultural Sciences (SLU) with 2 m resolution. The soil wetness was classified as dry, moist, and wet. The digital terrain model (DTM) was derived from the national laser scanning with 1 m resolution (GSD-Höjddata grid 1 + . The potential incoming solar radiation was calculated in QGIS (QGIS Development Team, 2009) Terrain Analysis, and the total insolation was calculated using DEM and calculated from 2021-04-01 to 2021-10-31. The slope gradient and terrain aspect products were calculated at 10 m resolution using QGIS.
The soil wetness, solar radiation, slope gradient, and terrain aspect of each plot and each tree were calculated from the rasters with different radii. The used radii were denoted as Rabiotic. The distance-weighted mean values were calculated for the soil wetness, solar radiation, and slope gradient. The distance-weighted median values were calculated for the terrain aspect.
Damage modeling
This study aimed to analyze the environmental factors of the damage level. The environmental factors were calculated from remotely sensed data and used as independent variables, and the damage levels were obtained from the field inventory and used as dependent variables. At the plot level, we used the number of trees attacked in 2021 in the plot, i.e., infestation patch size, to represent the damage levels. At the tree level, we used binomial values, i.e., healthy or attacked in 2021, to represent the damage levels of individual trees in the plot.
To visualize in which environment a pheromone bag resulted in more colonization, we first displayed the distribution of the environmental factors among different damage levels. At the plot level, three damage levels were analyzed, i.e., mild, moderate, and severe damages, with < 5, 5–9, and ≥ 10 attacked trees in the plots, respectively. At the tree level, two damage levels, i.e., healthy and attacked, were used to define the damage levels. At this step, Rbiotic = 50 m and Rabiotic = 15 m were used for the plot level, corresponding to the plot size, while Rbiotic = 50 m and Rabiotic = 5 m were used for the tree level, corresponding to a common crown size. This step is for a qualitative description of the distribution of environmental factors among damages with different levels.
A generalized linear model (GLM, Eq. 1) was used to quantify the impact of the factors on damage risks. The damage risks were set as the dependent variable, and the seven environmental factors were the independent variables. To reduce, the number of variables and avoid possible overfitting, aspect was classified into three classes: northeast slope (315°–135°), southwest slope (135°–315°), and flat terrain with slope gradient ≤ 2°). A GLM with a Poisson distribution was used at the plot level and a binomial distribution was used at the tree level. At the tree level, we also used the direction and distance to the pheromone bag as independent variables, to consider the potential impact on the damage level. We tested the significance of the independent variables using p-values, and selected better models according to the log-likelihood. In this step, we tested the impact radius of each factor. At the plot level, we set Raduisf and Rterrain as 20 m, 30 m, 40 m, and 50 m, and tested the changes in the log-likelihood of the models. At the tree level, we set Rbiotic and Rabiotic as 10 m, 15 m, 20 m, and 50 m. A model with a higher log-likelihood value implied a better fit with a certain radius, and the radius was used for further analysis. Then, we changed the radius of each factor one at a time, while keeping all the other factors with fixed radii. We answered, which factors significantly impact the damage level and the impact radius of each factor.
where the response\({y}_{i}\), \(i=1,\dots , n\) is modeled by a linear function of explanatory variables\({x}_{j}\), \(j=1,\dots , p\) plus an error term.
To answer, which factors are needed for predicting the damage risk, we used the significant factors with the radii decided from previous steps as the independent variables of GLM. At the individual tree level, the direction and distance to the pheromone bag were not used in the model, since they would not be available when predicting the damage risk of other areas. We compared, the Akaike information criterion (AIC) of models by removing factors one at a time. If removing one factor resulted in a significant AIC increase, we determined the factors necessary for the prediction model. At the tree level, we separated training and test dataset and evaluated the overall accuracy for predicting healthy/attacked. We randomly sampled healthy trees to have a balanced number of samples with the attacked tree, and then randomly sampled 70% of healthy and attacked trees as training samples, and the rest as test samples. We ran the sampling and estimation model 50 times and calculated the average Kappa coefficient and the overall accuracy. The model was not evaluated with separate training and testing datasets, due to the limited number of plots. The predicted and actual number of attacks were presented in a confusion matrix.
Results
Results from the controlled experiment
In the controlled experiment, 24 plots were set up with 15 m radius in 6 stands, including a total of 977 trees. The pheromone bags attracted bark beetles attacking the trees in the plots, resulting in 208 attacked trees. Large variations in the number of attacked trees, i.e., infestation patch size, were observed, even within the same stand (Fig. 2a). When having 30–64 trees in the plots, 38% of the plots had less than 5 attacked trees (mild damage), and 38% of the plots had 5–9 attacked trees (moderate damage). 25% of the plots had more than ten attacked trees (severe damage), including three plots with more than twenty attacked trees.

The distribution of healthy and attacked trees in each plot (a) and the DBH distribution in each plot (b) and stand (c). The boxes in (b) mark the median with red lines, 25th and 75th percentiles with box edges. The whiskers mark the most extreme data except for the outliers which are marked with red + symbol. c The number of healthy and attacked trees were marked with green and light red bars, and the overlapping numbers, e.g., in Stand No. 5, were marked with dark red
Only four out of 90 trees with DBH < 20 cm were attacked. For trees with DBH > 20 cm, the severity of the damages did not show an obvious trend with the DBH (Fig. 2b and c). The plots with larger mean DBH had more attacked trees only in 3 out of 6 stands (No. 1, 2, and 6). We tested, whether there was a linear correlation between the number of attacked trees and the mean DBH. The p-value of the linear correlation was 0.64, i.e., the mean DBH did not significantly influence the damage levels. At the individual tree level, the attacked trees had significantly larger DBH than the healthy ones only in 2 out of 6 stands at 5% significance level, although the DBH of healthy and attacked in all stands showed significant differences (Table 2).
In the controlled experiment, we used pheromone bags to attract bark beetles to attack the tree in the plots. Therefore, theoretically, the distances to the pheromone bags and the wind direction should have an impact on the spread of the infestation. We compared the proportion of attacked trees at different distances and directions to the pheromone bags. Trees closer to the pheromone bags and trees located south of the pheromone bags showed higher probabilities of being attacked (Fig. 3). The proportion of attacked trees significantly decreased with larger distances to the bags (p < 0.01 when testing the correlation, Fig. 3b). Among all trees located at the southeast and southwest to the center trees, 26% and 23% were attacked, while the proportions were only 12% and 16% for the trees located at the northeast and northwest to the center. This pattern could be attributed to the tendency of infestations being more prone spreading to the south border of the stands, where trees experienced higher solar radiation and direct radiation. This might also be related to the fact that we attached the pheromone bags at the south side of the trunks of the center trees before and during the first two weeks of swarming.

Distribution of the attacked trees. a Locations of healthy (green) and attacked (red) trees in all plots. b The proportion of attacked trees with different distances to the pheromone bags. c The proportion of attacked trees with different directions to the phenomenon bags. The blue bars are the number of all trees, including the number of attacked trees marked with red bars and the percentages
Distribution of the environmental factors
In this section, we showed the distribution of the environmental factors among plots with different attack levels and among healthy and attacked individual trees. The quantitative analysis of the factors is present in 3.3.
Deciduous trees and previous infestations
Compared to the mildly and moderately attacked plots, the severely attacked plots had significantly more deciduous trees and fewer TreesLeft, in the 50 m surroundings, tested with p value < 0.05 by a two-sided Wilcoxon rank-sum test. The severely attacked plots also had more TreesRemoved on average (Fig. 4), although the p value was 0.12. At the tree level, attacked trees had significantly more deciduous trees, fewer TreesLeft, and more TreesRemoved (Fig. 5). Note, that the attacks in this study were attracted by pheromone bags instead of natural spreads of infestations. The median distances to the nearest previous infestations were 28.9 m for plots and 30.1 m for individual trees. Therefore, this study does not present how infestations even closer (e.g., within 5 m or 10 m) influence the damage risks.

Distribution of the deciduous trees (a) and previous infestations (b) and (c) for plots with infestations < 5, 5–9, and ≥ 10 (green, yellow, and red dots). The black lines mark the average values

Violin plots of the deciduous trees (a) and previous infestations (b) and (c) around healthy and infested individual trees. The distribution curves were calculated using kernel density estimation and normalized to the same width shown in the figures. The black lines mark the medians
Soil wetness
In total, 88% of the plots and 88% of the trees were located in dry areas, and only three plots and 113 trees were in moist/wet areas. In the whole forest estate, 70% of the land was dry, and 26% and 4% were moist and wet, respectively. The samples selected from the forest estate were biased towards being located in dry areas. We set up the plots close to previous attacks, and we interpreted the results as the previous attacks occurred more in dry areas than in moist or wet areas. The proportions of severely attacked plots and attacked individual trees were much higher in dry areas than in moist or wet areas (Fig. 6).

The distribution of plots (a) and individual trees (b) in wet, moist and dry areas. The percentages in the figures mean that 29% of the plots located in dry areas were severely damaged, and 23% of the single trees in dry areas were attacked
Solar radiation
The severely attacked plots and attacked individual trees were distributed in areas with significantly less solar radiation compared to the mildly attacked plots and healthy trees (Fig. 7), tested with p value < 0.01 by a two-sided Wilcoxon rank-sum test.

The distribution of solar radiation of plots (a) and trees (b). The distribution curves on the tree level were calculated using kernel density estimation and normalized to the same width shown in the figures. The black lines mark the medians
Slope gradient
The proportions of severely attacked plots and attacked individual trees were higher on slopes than in flat areas (Fig. 8a). The occurrence of attacks increased on slopes with larger gradients (Fig. 8b).

The distribution of plots (a) and individual trees (b) on different slope gradients. The percentages in the figures referred to the plots with severe damage in proportion to the total number of plots (a), and the number of attacked trees in proportion to the total number of trees (b)
Terrain aspect
In our dataset, there were more plots and individual trees distributed on south slopes than the other directions, but the proportions of severe damage or attacked trees were lower. Infestations occurred more on the north and east slopes than on the south and west slopes (Fig. 9). Among all plots, 100% and 50% plots on the north and east slopes were severely damaged, while none on the south and west slope. No plot on the North Slope was mildly or moderately damaged, while 33%, 66%, and 50% of plots on the east, south, and west slopes were moderately damaged. Among individual trees, the proportions of trees being infested were 42%, 36%, 10%, and 10% on north, east, south, and west slopes, respectively.

Distribution of damages on different slope aspects. a The aspects of plots with different damage levels. b The aspects of healthy and attacked trees. The orientations of the points are accurate while random distances to the center were given to avoid points overlapping too much
Impact radius of the environmental factors
We used generalized linear models to test the significance of each factor on the risks, and test the impact radius of the factors. We first tested the correlation between variables by using the variance inflation factor (VIF). The VIF of the variables ranged from 1.04 to 1.60, indicating moderately correlated variables but insignificant multicollinearity that needed to be corrected (VIF < 10). We calculated log-likelihood to evaluate the model with factors of different radii. At the plot level, we set Rbiotic and Rabiotic to 20, 30, 40, and 50 m, respectively. The model showed better agreement with larger Rbiotic, while the models did not show a clear trend with increasing Rabiotic (Fig. 10a and b). We decided to use 50 m for both Rbiotic and Rabiotic for the later modeling. At the individual tree level, we set Rbiotic and Rabiotic to 10, 15, 20, …, 50 m, respectively. The model showed the best fit with Rbiotic = 15, and the log-likelihood decreased with increasing Rbiotic larger than 15 m (Fig. 10c). Similar to the plot level, the models at the tree level did not have constant changes by increasing Rabiotic (Fig. 10d). We set Rabiotic and Rbiotic both to 15 m for the tree level modeling.

Log-likelihood of models with different Rbiotic and Rabiotic at the plot level (a and b) and tree level (c and d)
We also tested the significance of the factors with different radii (Table 3). Among all factors, flat terrain (with a slope gradient no larger than 2 degrees) was the most significant factor, decreasing the damage risk, among all radii. Compared to northern slopes, southern slopes significantly decreased the damage risk. The effect was stronger on the local terrain aspect with smaller radii and weaker influences on the surrounding terrain with larger radii. The slope gradient (continuous degrees) was significant for larger radii but not smaller radii. At the plot level, solar radiation was not significant for any radius, and the soil wetness was only significant occasionally. At the tree level, solar radiation and soil wetness were not significant for 10–30 m radii, and showed stronger impacts for larger radii from 35 to 50 m.
More deciduous trees and removing attacked trees from the forest significantly increased the damage risks. Leaving attacked trees in the forest hardly influenced the damage risks at the plot level and the tree level when using small radii. With larger radii at the tree level, leaving attacked trees in the forest decreased the damage risk. Note that at the tree level, the model with larger radii did not fit as good as with smaller radii, although more factors were significant.
Modeling the damage risks
At the plot level, we used a 50 m radius to calculate all variables. The significant variables (Table 4) were the number of deciduous trees and TreesRemoved, slope gradient, and aspect (including northern slope, southern slope, and no slope). When only using these four variables, the AIC changed insignificantly compared to using all factors. However, removing any of these four variables made a significant decrease in AIC. At the tree level, the significant variables were the number of deciduous trees, the number of TreesRemoved, and the aspect (including the northern slope, southern slope, and no slope). When only using the number of deciduous trees and the aspect, the model fitted similarly compared to using all variables. The number of TreesRemoved did not improve the model, although it was significant. The model predicted number of attacked trees with R2 = 0.6 and RMSE = 3.9 (Fig. 11).

The predicted and actual number of attacked trees per plot
We evaluated the risk model by separating training and test data and used the overall accuracy and kappa coefficient on the test data to show the performance. We randomly sampled healthy trees to have a balanced number of samples with the attacked tree. When training on three plots in each stand and testing on the remaining plots in each stand, the average overall accuracy and kappa coefficient were 0.69 and 0.38. When training on trees in five stands and testing on trees in the remaining stand, the average overall accuracy and kappa coefficient were 0.56 and 0.13. The results illustrated that the model can be used to produce wall-to-wall maps with sampling data from the same stand, but the model cannot be transferred for risk estimation for other stands without sampling data. The low transferability to other stands may be due to the limited data size, i.e., only six stands. When training using trees in three plots in each stand and testing on trees in the remaining plot in each stand, an example of a confusion matrix from a single run is shown in Table 5:
Discussion
In this study, we observed the damage levels induced by pheromone bags, and investigated the environmental factors causing the different damage levels. We first presented the distribution of the environmental factors and observed that the damage occurred more in the areas with more deciduous trees, fewer previous infestations left in the stand, more removed previous infestations, dry soil, less solar radiation, undulating terrain, and north-east slope. However, not all these factors influenced the damage risk significantly, evaluated using a generalized linear model, and not all factors were necessary to predict the damage risk.
In this study, soil wetness did not impact the risk significantly when it was tested in the GLM model, although the proportions of severely attacked plots and attacked individual trees were much higher in dry areas than in moist and wet areas. The plots were selected in the forest stands with infestations from previous years, resulting in sample plots and trees being distributed more in dry areas. Insufficient number of samples located in moist and wet areas might lead to the insignificant impact of soil wetness on the damage risk. Similarly, solar radiation did not influence the risk, which might be due to flat terrain and small radiation differences. All samples were selected from mature spruce forest, and for trees with DBH > 20 cm, DBH did not impact the damage risk. The insignificant factors were consistent with some studies, while different from other studies. For example, in the study of Kautz et al. (2022), the standardized precipitation-evapotranspiration-index was the most important factor in explaining symptom occurrence of individual trees, while the study did not present the influence as positive or negative. The height and DBH did not explain the symptom occurrence, with data from field inventory on 85 hectares of spruce-dominated unmanaged forest. In the study of Mezei et al. (2014a, b), solar radiation was significant in the epidemic phase instead of the incipient epidemic and post-epidemic phase.
In this study, flat terrain (with slope gradient ≤ 2°) significantly decreased the damage risk. Similar results were shown by Blomqvist et al. (2018), where the lowest risk was associated with gentle slopes, and the highest risk was associated with moderately steep slopes. In the study of Kautz et al., (2022), slope gradient (°) showed high predictor importance for symptom occurrence tested by boosted regression trees and random forest (without presenting the effects as positive or negative), with data from field inventory. However, in the study of Groot and Ogris (2019) in Slovenia, the slope was considered to decrease the risk, examined by the amount of sanitary felling because of bark beetles on a large-scale. The suggested explanation was that areas with lower altitudes and flatter slopes were warmer and drier compared to mountainous areas, decreasing the tree vitality. The study by Nardi et al., (2022) on the landscape level (1 km grid on 100000 km2 areas) in north-eastern France also showed higher risks in flat areas, and similar results were also found in Alpine forests in Austria (Pasztor et al. 2014). Compared to the large-scale studies (Groot and Ogris 2019; Nardi et al. 2022; Pasztor et al. 2014), both this study and the study by Blomqvist et al. (2018) were conducted in flat areas in northern Europe, without a large-scale comparison with mountainous areas.
Besides slope steepness, the aspect also influenced the risk significantly and was one of the most important factors to predict the risk. In this study, damage risk was lower for the south and west aspect. Similar results were found in a study in Finland (Blomqvist et al. 2018), where the risk was lower at the southern and southwestern aspects. This phenomenon can be explained by the excessive direct solar radiation on the southern and western slopes that induce desiccation of the phloem and high thermal conditions, restricting infestations (Blomqvist et al. 2018; Wermelinger 2004; Jakuš, 1998). However, many studies have also presented higher risks at the south-facing slopes on different scales and regions (Pasztor et al. 2014; Overbeck and Schmidt 2012). This may be related to the triggering factors of the outbreaks.
Besides natural microclimate conditions, forest species composition also impacted the risk significantly. In this study, the infestation risk was higher with more deciduous trees in the surrounding. A possible explanation could be that the deciduous trees created canopy gaps in the winter and spring without leaves. Canopy gaps allowed direct radiation that caused physiological stress on spruce trees, and increased the temperature in the winter and spring, raising the beetle overwintering population (Pasztor et al. 2014; Netherer and Pennerstorfer 2001; Wermelinger 2004). A study in the Arkhangelsk region, Russia, showed that longer sunshine duration increased tree loss (Trubin et al. 2022). The deciduous trees could be strong water competitors that made the spruce more stressed and vulnerable. Nevertheless, a higher risk with a larger amount of other tree species, including deciduous trees, was not common in previous studies (Kärvemo et al. 2016; Groot et al. 2018; Overbeck and Schmidt 2012). Many large-scale studies have shown an increasing infestation risk with a higher amount of spruce, which was not surprising since the spruce bark beetle only colonizes spruce, and the infestation risk was modeled using the amount of damaged spruce divided by the total amount of trees (or per pixel) instead of the total amount of spruce trees. Deciduous trees might decrease the damage level of a large area, e.g., a stand, as suggested by other studies (Kärvemo et al. 2016), while deciduous trees might increase the risk of each individual spruce in the surrounding, as suggested by our study. In this study, the selected forest was mature with volumes of spruce larger than 80%. In such spruce-dominated forest, spruce volume might reach the threshold that deciduous trees started to increase the risk.
Removing standing dead trees in the 10–50 m range also significantly increased the damage risk and was one of the important factors in predicting the risk in this study. Ideally, the infested trees should be removed from the forest with the larvae under the bark to prevent the spread by the next generation. However, in practice, the attacked trees were often not found until they were already dead. For example, in this study, the trees attacked from the previous years were removed during the winter and early summer. Such a belated sanitation felling has been shown to be insufficient for population control, proven by a comparison between managed and unmanaged forests in conservation areas (Fora and Balog 2021). On the contrary, removing standing dead trees created canopy gaps and enabled direct solar radiation on trees inside the forest, and caused the physiological stress of spruce, similar to the effect of the deciduous trees in winter and spring. In addition, the standing dead trees can provide breeding materials for the competing species, and keeping them in the forest benefits insect predators and can lower the attack risk by bark beetles and maintain biodiversity (Wermelinger 2004).
The analyses and conclusions were based on our dataset with previous infestations within the 10–50 m range, but no closer than that. We would expect that the trees surrounding previous infestations, e.g., < 10 m range, had high risks of being attacked in the following years, but we did not have data in this study to quantify the risk. Similarly, we did not have data for the analysis with larger radius than 50 m. Therefore, there might be influence from longer distances than 50 m that we did not consider in this study. Besides, this study investigated the effect of removing individual standing dead trees instead of large tree groups, without other damaging events such as wind storms. Therefore, the investigated effect might be different from the sanitation felling of large infested areas and damaged areas by storms (Stadelmann et al. 2013; Mezei et al. 2017a; Overbeck and Schmidt 2012).
This study also illustrated the effective radius of the environmental factors on influencing the damage level. The effective radius was considered from two aspects: (1) whether the environmental factors were significant at a certain radius, and (2) whether they explained the model with a certain radius better, i.e., higher log-likelihood. When testing 10–50 m radii, the factors explained the model better with larger radii of the biotic factors for plots with 15 m radius, while the opposite was shown at the tree level. The radius of the biotic factors with the highest log-likelihood was 50 m to explain the infestation patch size and 15 m to explain a tree being infested or not. Changing the radius of the abiotic factors did not show a clear pattern in explaining the models, although some of them were more significant with larger radii. For example, soil wetness and solar radiation were insignificant to the tree level damage with a 15 m radius, but were more significant with larger radii, such as 50 m. Nevertheless, they did not contribute to predicting the infestation. This may be explained by the spatial autocorrelation, when using too large radius. When using the radius with the highest log-likelihood, i.e., 50 m for plots and 15 m for trees, the significance of environmental factors was consistent among plot and tree levels, while inconsistent with other radii. The results illustrated the importance of using a suitable radius of the environmental factors in analyzing the damage risks.
In general, the effects of environmental factors on the damage risks are complicated and can be case-specific. Conclusions can be very different from studies on a landscape or in-stand scales (Groot and Ogris 2019; Kärvemo et al. 2016), in a mountainous or flat region (Blomqvist et al. 2018; Overbeck and Schmidt 2012; Pasztor et al. 2014), in different climate zones (Blomqvist et al. 2018), short- or long-time effects (Hlásny and Turčáni 2013; Mezei et al. 2017a, 2014a), in different phases of the outbreak (incipient epidemic phase, epidemic phase, post-epidemic phase) (Mezei et al. 2014b), and influenced by previous disturbances such as wind storms (Groot et al. 2018; Stadelmann et al. 2013; Kärvemo et al. 2023), ice storms (Groot et al. 2018), drought (Nardi et al. 2022; Kärvemo et al. 2023), and air pollution (Jakuš, 1998).
In this study, the selected stands experienced wind-damage- and drought stress-triggered infestations before the data collection, but the initial attack during the study year was triggered by pheromone bags instead of occurring entirely naturally. We put up pheromone bags on a center tree before the first swarm, so the tree experienced mass colonization with limited time of defense. The pheromone bags were removed two weeks after the first swarm, so the parental beetles could select and attack other trees, and the defense and colonization process were more similar to the natural conditions. Similarly, the second swarm occurred in the late summer, leading to many trees being infested in some plots, while very few trees were infested in other plots. We assumed the pheromone bags to have a similar effect to the plots, and therefore, the damage level was determined by the predisposition influenced by the environmental factors. Nevertheless, using pheromone bags to trigger the initial attacks was still different from the natural case, and this may be the underlying reason for some of the results in the study.
This study also proposed risk models to predict the infestation size and predict individual trees being infested, based on the analysis of the environmental factors. At the plot level, the used factors were slope gradient, number of deciduous trees, the number of TreesRemoved, and the location on a south aspect or flat terrain. At the tree level, the used factors were the number of deciduous trees, located on a south aspect or flat terrain. TreesRemoved was significant in the model, but it was excluded without influencing the AIC. The model predicted the infestation size with an RMSE of 3.9 trees and overall accuracy of 0.7 in predicting individual trees being infested. We also stressed, that the tree risk model needed sampling data from the same stand, while having low transferability to stands without training. Compared to other risk models developed at the landscape level (Koreň et al. 2021; Mezei et al. 2014b), the models in this study were developed based on a controlled experiment and, thus limited by the sample size.
In the past, lots of studies have been analyzing the environmental factors of the beetle population (Marini et al. 2013; Fora and Balog 2021; Wermelinger 2004), which is an important aspect of the damage risks, while the forest vitality and resistance to the attacks also need to be considered. Remote sensing techniques can provide wall-to-wall forest damage maps as well as forest vulnerability estimates (Koreň et al. 2021; Huo et al. 2021), and thus should be further developed and applied in risk analysis in the future.
Conclusions
This study analyzed the environmental factors that influence the beetle damage level induced by pheromone bags. The investigated factors included soil wetness, solar radiation, slope gradient, terrain aspect, and the number of deciduous trees, the number of dead trees attacked in previous years that were removed or remained in the forest. When using GLM, factors within radii of 15 m for the individual tree level and 50 m for the plot level showed the best fit for modeling the damage level. Locating at the northeast slopes (315°–135° of aspect, > 2° slope), surrounded by more deciduous trees, and removing dead trees from the stand significantly increased the damage risk at both plot and individual tree level. Soil wetness, solar radiation, and the number of standing dead trees attacked from previous years did not impact the damage level significantly. The risk model on individual trees yielded 0.69 overall accuracy in predicting trees being infested or healthy, using the significant factors and trained on three plots and tested on the remaining plot in each stand. We concluded, the significant factors in the context of the study area, i.e., a plain area in southern Sweden. The proposed method can potentially contribute to wall-to-wall maps of in-stand damage risks using remote sensing data.
References
Blomqvist M, Kosunen M, Starr M, Kantola T, Holopainen M, Lyytikäinen-Saarenmaa P (2018) Modelling the predisposition of Norway spruce to Ips typographus L. infestation by means of environmental factors in southern Finland. Eur J for Res 137:675–691
de Groot M, Ogris N (2019) Short-term forecasting of bark beetle outbreaks on two economically important conifer tree species. For Ecol Manage 450:117495
de Groot M, Ogris N, Kobler A (2018) The effects of a large-scale ice storm event on the drivers of bark beetle outbreaks and associated management practices. For Ecol Manage 408:195–201
Fora CG, Balog A (2021) The effects of the management strategies on spruce bark beetles populations (Ips typographus and Pityogenes chalcographus), in Apuseni Natural Park. Romania for 12:760
Havašová M, Ferenčík J, Jakuš R (2017) Interactions between windthrow, bark beetles and forest management in the Tatra national parks. For Ecol Manage 391:349–361
Hlásny T, Turčáni M (2013) Persisting bark beetle outbreak indicates the unsustainability of secondary Norway spruce forests: case study from Central Europe. Ann for Sci 70:481–491
Hlásny T, Zimová S, Merganičová K, Štěpánek P, Modlinger R, Turčáni M (2021) Devastating outbreak of bark beetles in the Czech Republic: drivers, impacts, and management implications. For Ecol Manage 490:119075
Huo L, Persson HJ, Lindberg E (2021) Early detection of forest stress from European spruce bark beetle attack, and a new vegetation index: normalized distance red & SWIR (NDRS). Remote Sens Environ 255:112240
Huo L, Lindberg E, Bohlin J, Persson HJ (2023) Assessing the detectability of European spruce bark beetle green attack in multispectral drone images with high spatial- and temporal resolutions. Remote Sens Environ 287:113484
Jakuš R (1998) Types of bark beetle (Coleoptera: Scolytidae) infestation in spruce forest stands affected by air pollution, bark beetle outbreak and honey fungus (Armillaria mellea). J Pest Sci 71:41–49
Kärvemo S, Johansson V, Schroeder M, Ranius T (2016) Local colonization-extinction dynamics of a tree-killing bark beetle during a large-scale outbreak. Ecosphere 7:161
Kärvemo S, Huo L, Öhrn P, Lindberg E, Persson HJ (2023) Different triggers, different stories: bark-beetle infestation patterns after storm and drought-induced outbreaks. For Ecol Manage 545:121255
Kautz M, Schopf R, Ohser J (2013) The “sun-effect”: microclimatic alterations predispose forest edges to bark beetle infestations. Europ J for Res. https://doi.org/10.1007/s10342-013-0685-2
Kautz M, Peter FJ, Harms L, Kammen S, Delb H (2022) Patterns, drivers and detectability of infestation symptoms following attacks by the European spruce bark beetle. J Pest Sci 5:87
Koreň M, Jakuš R, Zápotocký M, Barka I, Holuša J, Ďuračiová R, Blaženec M (2021) Assessment of machine learning algorithms for modeling the spatial distribution of bark beetle infestation. Forests 12:395
Marešová J, Majdák A, Jakuš R, Hradecký J, Kalinová B, Blaženec M (2020) The short-term effect of sudden gap creation on tree temperature and volatile composition profiles in a Norway spruce stand. Trees 34:1397–1409
Marini L, Lindelöw Å, Jönsson AM, Wulff S, Schroeder LM (2013) Population dynamics of the spruce bark beetle: a long-term study. Oikos 122:1768–1776
Mezei P, Grodzki W, Blaženec M, Jakuš R (2014a) Factors influencing the wind–bark beetles’ disturbance system in the course of an Ips typographus outbreak in the Tatra Mountains. For Ecol Manage 312:67–77
Mezei P, Grodzki W, Blaženec M, Škvarenina J, Brandýsová V, Jakuš R (2014b) Host and site factors affecting tree mortality caused by the spruce bark beetle (Ips typographus ) in mountainous conditions. For Ecol Manage 331:196–207
Mezei P, Blaženec M, Grodzki W, Škvarenina J, Jakuš R (2017a) Influence of different forest protection strategies on spruce tree mortality during a bark beetle outbreak. Ann for Sci 74:660
Mezei P, Jakuš R, Pennerstorfer J, Havašová M, Škvarenina J, Ferenčík J, Slivinský J, Bičárová S, Bilčík D, Blaženec M, Netherer S (2017b) Storms, temperature maxima and the Eurasian spruce bark beetle Ips typographus—An infernal trio in Norway spruce forests of the Central European High Tatra Mountains. Agric for Meteorol 242:85–95
Mezei P, Potterf M, Škvarenina J, Rasmussen JG, Jakuš R (2019) Potential solar radiation as a driver for bark beetle infestation on a landscape scale. Forests 10:604
Nardi D, Jactel H, Pagot E, Samalens J-C, Marini L (2022) Drought and stand susceptibility to attacks by the European spruce bark beetle: a remote sensing approach. Agric for Entomol 6:161
Netherer S, Pennerstorfer J (2001) Parameters relevant for modelling the potential development of Ips typographus (Coleoptera: Scolytidae). Integr Pest Manag Rev 6:177–184
Ogris N, Jurc M (2010) Sanitary felling of Norway spruce due to spruce bark beetles in Slovenia: a model and projections for various climate change scenarios. Ecol Model 221:290–302
Okland Bjorn, Berryman Alan (2004) Resource dynamic plays a key role in regional fluctuations of the spruce bark beetles Ips typographus. In Agric Forest Ent 6(2):141–146. https://doi.org/10.1111/j.1461-9555.2004.00214.x
Overbeck M, Schmidt M (2012) Modelling infestation risk of Norway spruce by Ips typographus (L.) in the Lower Saxon Harz Mountains (Germany). For Ecol Manage 266:115–125
Pasztor F, Matulla C, Rammer W, Lexer MJ (2014) Drivers of the bark beetle disturbance regime in Alpine forests in Austria. For Ecol Manage 318:349–358
Schuldt B, Buras A, Arend M, Vitasse Y, Beierkuhnlein C, Damm A, Gharun M, Grams TEE, Hauck M, Hajek P, Hartmann H, Hiltbrunner E, Hoch G, Holloway-Phillips M, Körner C, Larysch E, Lübbe T, Nelson DB, Rammig A, Rigling A, Rose L, Ruehr NK, Schumann K, Weiser F, Werner C, Wohlgemuth T, Zang CS, Kahmen A (2020) A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl Ecol 45:86–103
Seidl R, Schelhaas M-J, Rammer W, Verkerk PJ (2014) Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 4:806–810
Senf C, Sebald J, Seidl R (2021) Increasing canopy mortality affects the future demographic structure of Europe’s forests. One Earth 4:749–755
Sproull GJ, Adamus M, Bukowski M, Krzyżanowski T, Szewczyk J, Statwick J, Szwagrzyk J (2015) Tree and stand-level patterns and predictors of Norway spruce mortality caused by bark beetle infestation in the Tatra Mountains. For Ecol Manage 354:261–271
Stadelmann G, Bugmann H, Meier F, Wermelinger B, Bigler C (2013) Effects of salvage logging and sanitation felling on bark beetle (Ips typographus L.) infestations. For Ecol Manage 305:273–281
Trubin A, Mezei P, Zabihi K, Surový P, Jakuš R (2022) Northernmost European spruce bark beetle Ips typographus outbreak: Modelling tree mortality using remote sensing and climate data. For Ecol Manage 505:119829
Wermelinger B (2004) Ecology and management of the spruce bark beetle Ips typographus—a review of recent research. For Ecol Manage 202:67–82
Acknowledgements
This project was funded by the Stiftelsen Nils och Dorthi Troëdssons Foundation (1042-21) and co-funded by the European Union (Horizon project RESDINET), with data collection co-funded by Hildur & Sven Wingquist’s Foundation for Forest Science Research (21/22-5 / 123-7), the SLU Forest Damage Center, the Stiftelsen Seydlitz MP bolagen Foundation (No. 83). We thank Magnus Ekström for providing consultation in the statistical modeling of the study. We thank Ronja Risberg for the field data collection.
Funding
Open access funding provided by Swedish University of Agricultural Sciences.
Author information
Authors and Affiliations
Contributions
LH: Conceptualization, Methodology, Software, Validation, Formal analysis, Investigation, Resources, Data curation, Writing—original draft, Writing—review & editing, Visualization, Project administration, Funding acquisition. HP: Conceptualization, Methodology, Validation, Formal analysis, Writing—Original Draft, Writing—Review & Editing, Funding acquisition. EL: Conceptualization, Investigation, Methodology, Writing—Original Draft, Writing—Review & Editing, Funding acquisition.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Communicated by Kathrin Blumenstein.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Huo, L., Persson, H.J. & Lindberg, E. Analyzing the environmental risk factors of European spruce bark beetle damage at the local scale. Eur J Forest Res 143, 985–1000 (2024). https://doi.org/10.1007/s10342-024-01662-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-024-01662-4