
Abstract
Bryophytes respond to a variety of environmental factors, including tree species composition and geological conditions, which are influenced by both natural and anthropogenic factors. Bryophytes growing on a variety of substrates were identified in ICP Forests monitoring plots in a wide range of close-to-nature managed forests, from lowland to high mountain areas in Slovenia. Based on data on tree layer composition (broadleaves vs. conifers) and on bedrock and soil type (calcareous vs. siliceous bedrock), all study plots were classified into five groups representing forest types. Different ordination analyses and statistical tests were used to represent differences between forest types in bryophyte composition, richness and functional traits. Almost 200 bryophyte species were identified in managed forests, with the most common being Hypnum cupressiforme, Brachytheciastrum velutinum and Polytrichum formosum. Bryophyte species composition varied considerably along the tree species composition and edaphic gradients represented by the selected forest types. Bryophyte species richness was significantly higher on calcareous bedrock than on siliceous bedrock. The higher occurrence of rocks on the ground surface may significantly increase the availability of various microhabitats for bryophytes. Bedrock had a significant influence on the functional diversity and composition of bryophytes, while tree species composition had less influence. Bedrock and tree species composition are important drivers of bryophyte species composition and diversity at taxonomic and functional trait levels. The high diversity of bryophyte species in Slovenian forests can also be attributed to close-to-nature and sustainable forest management practices that preserve favourable microclimatic conditions in forest stands and maintain diverse substrates.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bryophytes are an integral part of forest ecosystems and account for a significant proportion of plant species diversity. They are considered to be good ecological indicators because their specific characteristics make them sensitive to environmental change (Gignac 2001). Mosses and liverworts are also reliable ecological indicators of the condition and naturalness of forest (Sabovljević et al. 2010; Czerepko et al. 2021) and exhibit associations with different forest types (Stefańska-Krzaczek et al. 2022). Bryophytes respond to management intensity and disturbance in temperate forests and therefore have important implications for forest management and conservation (Horvat et al. 2017).
The composition of bryophyte assemblages in forest ecosystems is influenced by climatic factors such as air temperature and precipitation (Sun et al. 2013); historical factors (Fritz and Brunet 2010); forest integrity, which includes natural biodiversity, stand structure and continuity (Frego 2007); silvicultural treatments combining tree felling and burning (Kantvilas et al. 2015) and disturbance from logging (Nelson and Halpern 2005). The elevation gradient, which is closely related to climatic conditions, is an important driver of bryophyte diversity (Grau et al. 2007; Horvat et al. 2017). Moreover, a significant turnover in bryophyte species composition might be expected along a longitudinal gradient (Heilmann-Clausen et al. 2014) as well as along a latitudinal gradient. In addition to latitudinal, longitudinal and altitudinal gradients, topography in terms of microrelief diversity is also an important factor in bryophyte diversity (Bruun et al. 2006). On a small scale, forest management can determine bryophyte diversity and composition through its effects on tree species diversity, vertical structure, canopy closure, microclimate and deadwood availability (Bengtsson et al. 2000; Paillet et al. 2010; Czerepko et al. 2021). Forest stand structure determines light conditions, which strongly affect bryophyte diversity (Márialigeti et al. 2009; Tinya et al. 2009, 2021; Tinya and Ódor 2016), and microclimatic conditions and substrate availability are of great importance for bryophyte composition, cover and diversity (Márialigeti et al. 2009; Király and Ódor 2010; Müller et al. 2019).
In forests, different ecological guilds of bryophytes can be identified based on the substrate they grow on. These include epigeic (terricolous, occurring on soil), epixylic (occurring on deadwood), epiphytic (occurring on the bark of living trees and shrubs) and epilithic (occurring on rocks and stones) species. Some bryophyte species are able to grow ubiquitously, i.e. on a variety of substrates (soil, rocks, bark and deadwood) (Stokland et al. 2012), while others grow exclusively as epixylic or epiphytic species and are thus highly dependent on the quantity of available substrate. Epixylic and epiphytic bryophytes are more abundant and diverse in unmanaged forests than in managed forests (Lesica et al. 1991), and naturally regenerated forests have more bryophyte ground species than plantations (Yan et al. 2013). Because of forest management effects, cryptogamic epiphytes are considered to be a threatened group in temperate forests (Paillet et al. 2010) and are therefore useful for assessing the impact of forest management.
Bryophytes, especially liverworts, are more common in old-growth, close-to-nature and sustainably managed forests, and many of these species are absent in second-growth forests. Many bryophyte species find optimal habitat in old-growth forests, and these species are becoming increasingly rare as old forests are converted to younger forests (Lesica et al. 1991).
Moreover, the diversity of deadwood-inhabiting bryophytes is high in forest reserves along the studied geographical gradient in Europe, especially in pristine forest reserves in Slovenia, suggesting that it is influenced by both climatic and management factors such as forest history and continuity, deadwood availability and habitat fragmentation (Ódor et al. 2006). Sabovljević et al. (2010) confirmed that bryophyte species diversity is higher in primaeval forest stands where a spectrum of deadwood is present at different stages of decay.
Canopy tree species determine bryophyte species richness and cover, and differences have been found between forests with deciduous and coniferous trees (Rola et al. 2021). The ratio between deciduous and coniferous trees can be altered by forest management, and changes in the dominant tree species in the forest stand can significantly affect bryophyte communities (Rola et al. 2021).
The diversity and quality of different growing substrates for bryophytes are significantly affected by forest management. Bryophytes, especially typical woodland species, are therefore suitable indicators of forest management (Nordén et al. 2007; Kriebitzsch et al. 2013; Mölder et al. 2015; Müller et al. 2019) and valuable for estimating the naturalness and integrity of forest stands (Frego 2007). The species richness of typical woodland indicator bryophytes, which strictly depends on forest conditions, is negatively affected by management intensity and is therefore a better indicator of forest integrity than the species richness of all bryophytes (Müller et al. 2019). Therefore, nature conservation efforts should focus on reducing management intensity.
Although the species composition and diversity of bryophytes in Central Europe have been fairly well studied, the patterns associated with their life-history traits are still relatively unknown (Żarnowiec et al. 2021). The concept of bryophyte life forms has been discussed in several studies (Schofield and Hébant 1984; Düll 1991; During 1992; Longton 1997; Bates 1998). The potential of bryophyte life forms as ecological indicators has been tested under different ecological conditions (Żarnowiec 1995, 1996; Vittoz et al. 2010; Pardow et al. 2012; Vieira et al. 2012; Spitale et al. 2020). In addition to life forms, the type of substrate inhabited is another key element for the classification of bryophytes into plant functional groups (Andersson and Hytteborn 1991; Kürschner 1999; Jagodziński et al. 2018; Fojcik et al. 2019; Staniaszek-Kik et al. 2019). The colonization of coarse woody debris has been explained by the functional traits of bryophytes (Żarnowiec et al. 2021). Thus, biological traits explain the distribution of bryophyte species and their responses to climate variability and human impacts.
There are few databases containing bryophyte traits (Düll 1991; Hill et al. 2007; Schmidt et al. 2011; Bernhardt-Römermann et al. 2018). Contemporary bryophyte trait databases include not only different life forms but also their habitat requirements expressed by Ellenberg-type indicator values associated with regeneration, plant size and some regional characteristics related to the phytogeographic conservation status of a species.
In Slovenia, the species diversity of bryophytes has been studied in various forests and neighbouring ecosystems, such as forest reserves (Hočevar et al. 1980a, b, c, 1985; Ódor et al. 2005, 2006), selected managed forests (Kutnar and Martinčič 2008), and mires and surrounding spruce forests (Piskernik and Martinčič 1970, 1985; Kutnar and Martinčič 2001, 2002, 2003).
However, to date there has been no systematic approach to determine the species richness and composition of bryophytes and their functional traits in different Slovenian forests managed according to close-to-nature and sustainable principles. Therefore, the aims of this study were i) to investigate the species composition and diversity of bryophytes in selected managed forest types characterized by different tree species composition and edaphic (e.g. bedrock, soil) conditions and ii) to investigate the functional trait composition and diversity of bryophytes in these managed forest types.
Materials and methods
Study area and sampling of bryophytes
The studied plots were located in very different Slovenian forest vegetation types (Kutnar et al. 2012), which are mainly managed according to close-to-nature, sustainable and multifunctional principles. In lowland areas that are periodically flooded, forest stands occur in narrow strips along rivers and streams and are mainly dominated by willows (Salix sp.), alders (Alnus glutinosa (L.) Gaertn., A. incana (L.) Moench), ashes (Fraxinus excelsior L., F. angustifolia Vahl) and pedunculate oak (Quercus robur L.), with admixtures of European hornbeam (Carpinus betulus L.). In the hilly areas above the floodplains, mixed forests of sessile oak (Quercus petraea (Matt.) Liebl.) and European hornbeam are the predominant forest type. Most of the mid-altitude mountain areas are covered by forests dominated by European beech (Fagus sylvatica L.), with admixtures of other broadleaves (e.g. Acer pseudoplatanus L., Fraxinus excelsior, Ulmus glabra Huds.) and conifers (Abies alba Mill., Picea abies (L.) Karst.). In the Alpine region, various European beech forests mixed with Norway spruce (Picea abies), European silver fir (Abies alba) and European larch (Larix decidua Mill.) reach the timberline up to the belt of the dwarf mountain pine (Pinus mugo Turra). Scots pine (Pinus sylvestris L.) forests occur throughout the country on shallow soils on dolomite and also on acidic, nutrient-poor soils. Minor areas of Austrian pine (Pinus nigra Arnold) forests grow on extreme sites with a warmer microclimate. The sub-Mediterranean region is covered by forests and shrubby vegetation of thermophilous broadleaf species (e.g. Ostrya carpinifolia Scop., Fraxinus ornus L., Sorbus aria (L.) Crantz, Quercus cerris L., Q. pubescens Willd.). A similar type of forest is can also be found all over the country on sun-exposed, south-facing slopes with predominant limestone and dolomite bedrock. The described natural forest types are also interspersed with secondary forest communities dominated by Norway spruce, which have been significantly affected by climate change and various disturbances in recent decades (Kutnar et al. 2021).
Bryophytes (i.e. mosses and liverworts) were sampled in different managed forests in Slovenia between 2004 and 2010. The study area encompasses 47 ICP Forests Level I monitoring plots and 10 ICP Forests Level II monitoring plots (Intensive Monitoring programme; Kutnar and Martinčič 2008), i.e. 57 plots in total. ICP Forests (International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests) is a transnational European network for monitoring the forest status in Europe (de Vries et al. 2003). Level I plots are systematically distributed in a 16 km × 16 km (Urbančič et al. 2009; Kutnar 2011) and 16 km × 8 km grid across Slovenia. Level II plots were established in 10 different locations, which are representative of the heterogeneity of Slovenian forest types (Urbančič et al. 2016). Detailed information on these study plots can be found in various recent studies (Kutnar et al. 2019; Kermavnar and Kutnar 2020; Kermavnar et al. 2021a, b). The Level I and Level II plots are located in forest sites with a wide variety of climatic, geological, edaphic and topographical conditions, as well as a broad range of forest vegetation composition. The altitude of the study plots ranges from 160 to 1490 m, mean annual temperature ranges from 3.2 to 11.7 °C and mean annual precipitation ranges from 791 to 2499 mm. Climatic parameters were obtained from the WorldClim database (WorldClim 2021).
Based on the typology of Slovenian forests (Kutnar et al. 2012) and important stand and site characteristics (e.g. tree species composition, bedrock and soil) in the study plots, all plots were classified into five predefined groups representing different forest types (Fig. 1): i) stands dominated by broadleaves on calcareous bedrocks (B_c: 21 plots; Fig. 2), ii) stands dominated by broadleaves on intermediate bedrocks (B_i: 12 plots; Fig. 3), iii) stands dominated by broadleaves on siliceous bedrocks (B_s: 10 plots; Fig. 4), iv) stands dominated by conifers on calcareous bedrocks (C_c: 6 plots; Fig. 5) and v) stands dominated by conifers on siliceous bedrocks (C_s: 8 plots; Fig. 6).
Among broadleaves, European beech most frequently dominated the forest stands, while Norway spruce was the dominant coniferous species. Among the calcareous bedrocks (Groups B_c and C_c), limestone and dolomite were predominant. In the plots of these groups, the most common soils were Eutric Cambisols, Rendzic Leptosols and Chromic Cambisols (Urbančič et al. 2005, 2009, 2016). In Group B_i with intermediate bedrocks, Pleistocene sediments, alluvium and various mixed rock types were more common, on which Eutric Cambisols, Luvisols, Gleysols, Fluvisols or even Dystric Cambisols were formed. In the groups with siliceous bedrocks (Groups B_s and C_s), various non-calcareous rocks predominated, such as different siliceous metamorphic and igneous rocks, moraines, fluvioglacial gravels and sands, and also siliceous flysch. Among them were very diverse bedrock types such as sandstones, mica schists, dioritoid, tonalite, gneiss and amphibolite. Dystric Cambisols, Dystric Leptosols and Eutric Cambisols were the predominant soil types in the groups with siliceous bedrocks (Urbančič et al. 2005, 2009, 2016).

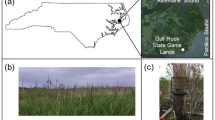
Distribution of 57 study plots across Slovenia, belonging to five groups indicating forest types with similar tree layer composition, bedrock and soil characteristics

Selected representative sites dominated by broadleaves on calcareous bedrocks (Group B_c) (Photograph: L. Kutnar)

Selected representative sites dominated by broadleaves on intermediate bedrocks (Group B_i) (Photograph: L. Kutnar)

Selected representative sites dominated by broadleaves on siliceous bedrocks (Group B_s) (Photograph: L. Kutnar)

Selected representative sites dominated by conifers on calcareous bedrocks (Group C_c) (Photograph: L. Kutnar)

Selected representative sites dominated by conifers on siliceous bedrocks (Group C_s) (Photograph: L. Kutnar)
During the fieldwork, all tree species in the study plots were recorded and the following parameters were estimated: cover of each vegetation layer (tree, shrub and herb) separately (in %), cover of rocks and stones (in %) and cover of woody debris (deadwood) (in %). Descriptive statistics for these parameters are summarized in Table 1.
Bryophytes were sampled in all plots in a survey area of 400 m2. All study plots were systematically sampled for bryophytes on all substrates: (i) forest floor (including bare soils and litter), (ii) deadwood (including stumps, lying logs and branches, standing dead trees), (iii) living wood (including the bark of living trees and shrubs) and (iv) rocks and stones. We estimated the cover of the bryophyte layer (in %). Determination of species identity was finalized in the laboratory using a microscope and different identification keys (e.g. Smith 2004; Frey et al. 2006; Guerra et al. 2006, 2010, 2014, 2018; Guerra and Cros 2007; Brugues and Guerra 2015; Huggonot and Chavoutier 2021). The nomenclature followed Hodgetts et al. (2020). The list of bryophyte species is provided in Supplementary Information 1.
Bryophyte functional traits
We obtained data on different autecological, morphological and reproductive traits of the bryophyte species selected for this study from the BryForTrait database (Bernhardt-Römermann et al. 2018) (Table 2). Indicator values for nutrients were additionally supplemented with information provided by Simmel et al. (2021).
Trait selection was based on our expert decision, previous studies, data completeness and preliminary analysis of response to predictor variables (Supplementary Information 2). The mean data completeness across all selected traits was high (94.6%).
Data analyses
In the statistical analyses, the bryophyte species recorded on the different substrates in each plot were analysed together. Multiple occurrences of a given bryophyte species on different substrates in the selected plot were considered as one entry in the analyses.
The total number of bryophyte species was calculated, and the most frequent species were identified in all study plots. Bryophyte species richness and bryophyte layer cover (%) were calculated for five forest types. The most frequent bryophyte species in these forest types were identified (Supplementary Information 1).
The conservation status of bryophytes was assessed at the European (Bern Convention 1979; EU Habitats Directive 1992; Hodgetts et al. 2019) and national level (Martinčič 2016) (Supplementary Information 1).
All statistical analyses were performed in R version 4.1.1 (R Core Team 2021). The compositional gradient in the presence-absence species × plot matrix was explored by multivariate analyses. Non-metric multidimensional scaling (NMDS) ordination with Bray–Curtis dissimilarity coefficients was used to depict differences in species composition data (metaMDS function in the vegan package; Oksanen et al. 2020). For this analysis, we used two dimensions (k = 2), resulting in a stress level of 0.175, which is below the stress < 0.2 criterion sensu Clarke (1993). We used the ordiellipse function (Oksanen et al. 2020) to plot the 95% confidence intervals of group scores onto the NMDS ordination diagram. To test whether groups (forest types) differed significantly in terms of species composition, we first employed a permutational multivariate analysis of variance (PERMANOVA; Anderson 2017) with 9999 permutations (vegan package, function adonis2). We then extracted NMDS axis 1 and axis 2 scores of samples (study plots) and compared them between groups (forest types) with the non-parametric Kruskal–Wallis test on ranks with Bonferroni correction for multiple comparisons (agricolae R package; de Mandiburu 2021). Differences in species richness (defined as the number of species per plot) between groups were tested using a generalized linear model (GLM) with Poisson error distribution for count data and visualized with boxplots. Significantly different means between groups (forest types) were separated using Tukey's post hoc test.
For functional characterization of bryophyte assemblages, we first calculated community means for each of the 13 traits (Table 2) using the funtcomp function in the FD package (Laliberté et al. 2015). The mean values of traits and trait states (27 in total), which represent the functional composition of assemblages, were then used as the input data for principal component analysis (PCA) to explore the distribution of plots and plot groups in the ordination space using the PCA function with automatic data standardization in the FactoMineR package (Husson et al. 2020). Visualizations were performed with the factoextra R package (Kassambara and Mundt 2020) to plot the 95% confidence intervals of group scores on the two-dimensional PCA ordination diagram. Differences in PCA1 and PCA2 scores between the five groups were tested with the non-parametric Kruskal–Wallis test on ranks with Bonferroni correction for multiple comparisons. Plot-level functional diversity was calculated with the dbFD function in the FD package. Functional dispersion was used as an index of functional diversity (FD) and visualized with boxplots. Functional dispersion is calculated as the mean distance of individual species to the centroid of all species in the community. This frequently used measure of functional diversity is unaffected by community species richness (Laliberté and Legendre 2010). Higher values of the functional dispersion index imply greater differences between species in terms of functional traits (i.e. trait divergence), whereas lower values suggest trait convergence. For each species trait, differences in functional dispersion were tested using a GLM with gamma error distribution. Significantly different means between groups (forest types) were separated using Tukey's post hoc test. Statistical significance was declared at α = 0.05 for all tests.
Results
Species composition and richness
A total of 199 bryophyte species were identified on 57 plots, the most frequent of which were Hypnum cupressiforme Hedw. (present on 91.2% of all plots), Brachytheciastrum velutinum (Hedw.) Ignatov & Huttunen (63.2%) and Polytrichum formosum L. (61.4%) (Supplementary Information 1). Among the more common species were also Ctenidium molluscum (Hedw.) Mitt. (52.6%) and Isothecium alopecuroides (Lam. Ex Dubois) Isov (50.9%).
Bryophyte species composition differed significantly between the five groups (PERMANOVA: p < 0.001, R2 = 0.189; Fig. 7). Forest plots dominated by broadleaves and conifers on calcareous bedrocks (Groups B_c and C_c) had significantly higher NMDS axis 1 scores than plots dominated by broadleaves on siliceous bedrocks (Group B_s). With respect to the NMDS axis 2 scores, plots dominated by broadleaves on intermediate and calcareous bedrocks (Groups B_i and B_c) exhibited significantly higher values compared to plots dominated by conifers on siliceous bedrocks (Group C_s) (Fig. 7).

Non-metric multidimensional scaling (NMDS) ordination according to bryophyte species composition. Ellipsoid hulls represent 95% confidence constructed around the centroid for each of the five forest types: B_c—broadleaves on calcareous bedrocks, B_i—broadleaves on intermediate bedrocks, B_s—broadleaves on siliceous bedrocks, C_c—conifers on calcareous bedrocks and C_s—conifers on siliceous bedrocks
Overall, mean species richness per plot was 19.4. The highest plot-level richness amounted to 36 species, whereas the most species-poor plot contained only five bryophyte species. Forest types differed significantly in bryophyte species richness (Fig. 8; Table 3), with the groups of plots on calcareous bedrocks (mean species richness—B_c: 23.8; C_c: 22.8) exhibiting significantly higher species richness than those on siliceous bedrocks (B_s: 13.8; C_s: 14.1). Group B_i (18.8) significantly differed from Group B_s (Fig. 8).

Comparison of plot-level bryophyte species richness between five forest types (B_c, B_i, B_s, C_c, C_s—see Fig. 7 for explanation). Bold lines indicate medians and whiskers above and below the box denote the 10th and 90th percentiles. Groups not sharing the same letter (beside the boxplot) significantly differ at p < 0.05 (GLM and Tukey's tests)
Functional trait composition and diversity
In the two-dimensional PCA space, the first axis explained 18.9% of trait variation and the second axis explained 12.7% of the variance (Fig. 9). Forest types did not differ significantly in terms of axis 1 scores. However, we found significant differences when PCA axis 2 was tested. Groups B_i and B_c had significantly higher axis 2 scores than Groups B_s and C_s (Fig. 9). Overall, the functional traits (trait states) with the highest contribution were hemeroby (9.1%), the indicator value for soil reaction (7.5%), common fruiting frequency (7.4%), mat as a life form (6.5%) and shoot length (5.8%). In contrast, the traits with the lowest explanatory power were the indicator value for temperature (1.3%), dendroid as a life form (0.8%) and the category expressing the affinity of species to forest habitats—species preferring forest edges and clearings (0.7%). The traits most strongly positively correlated with the first PCA dimension were hemeroby (0.88), species occurring in forests as well as in open land (0.69) and common fruiting frequency (0.63), whereas the following traits showed a strong negative correlation with PCA1: species largely restricted to closed forest (− 0.64), mat as a life form (− 0.61) and the indicator value for soil reaction (− 0.52). The traits most positively correlated with the second PCA dimension were the indicator value for soil reaction (0.61), common fruiting frequency (0.48) and perennial shuttle as a life strategy (0.47), whereas the following traits showed a strong negative correlation with PCA2: shoot length (− 0.68), the indicator value for soil moisture (− 0.52) and perennial stayer as a life strategy (− 0.42) (Fig. 9).

Principial component analysis (PCA) ordination diagrams based on bryophyte functional traits. On the left panel, 95% confidence ellipses indicate five forest types (B_c, B_i, B_s, C_c, C_s—see Fig. 7 for explanation). On the right panel, traits and trait states are coloured according to their contribution (relative importance) and are labelled as follows: indL = indicator value for light, indT = indicator value for temperature, indK = indicator value for continentality, indF = indicator value for moisture, indR = indicator value for soil reaction, indN = indicator value for nutrients; forest1 = M1.1; species largely restricted to closed forests, forest2 = M1.2; species preferring forest edges and in clearings, forest3 = M2.1; species occurring in forests as well as in open land, forest4 = M2.2; species that may occur in forests, but prefer open land; hemeroby = refers to human impact; LF1 = cushion as a life form, LF2 = dendroid as a l. f., LF3 = mat as a l. f., LF4 = turf as a l. f., LF5 = weft as a l. f.; LS1 = colonist as a life strategy, LS2 = perennial shuttle as a l. s., LS3 = perennial stayer as a l. s., LS4 = short-lived shuttle as a l. s.; shoot = shoot length; spore = size of spores; fruit1 = common fruiting frequency, fruit2 = frequent fruiting, fruit3 = occasional fruiting, fruit4 = rare fruiting, fruit5 = very rare fruiting
In the case of functional diversity, statistically significant differences between forest types were detected for the five traits: the indicator value for light, the indicator value for continentality, the indicator value for moisture, shoot length and life form (Fig. 10). Group B_i had significantly higher functional dispersion for light indicator values compared to B_s. Group B_c was characterized by significantly lower dispersion for continentality values than C_s. The latter group also exhibited significantly higher dispersion for moisture indicator values compared to B_i. The greatest differences in functional diversity were observed for shoot length. The highest dispersion for this trait was in Group B_s and was significantly higher than that for B_c and B_i. Plots in Group C_s also differed significantly from Group B_c. Finally, the diversity of life forms was highest in Group C_c and lowest in Group C_s (Fig. 10).

Comparison of functional dispersion between five forest types (B_c, B_i, B_s, C_c, C_s—see Fig. 7 for explanation) for the indicator value for light, the indicator value for continentality, the indicator value for moisture, shoot length and life form. Bold lines indicate medians, and whiskers above and below the box denote the 10th and 90th percentiles. Groups not sharing the same letter (beside the boxplot) significantly differ at p < 0.05 (GLM and Tukey's tests)
Turf (upright shoots with little or no branching, standing close together; Bernhardt-Römermann et al. 2018) was by far the most common life form in the studied dataset. In addition, we observed that the turf life form strongly determined the functional trait dispersion of bryophyte assemblages. Plots with a high representativeness of turfs were characterized by a lower diversity of life forms and vice versa (linear regression between life form community mean value and FD: R2 = 0.95).
Discussion
Species composition and diversity
In this study, a large number of bryophyte species (199) were identified in various Slovenian temperate forests, representing almost a quarter of the total species pool of bryophytes in Slovenia (Martinčič 2016), and the average species richness per plot was 19.4. The number of bryophyte species per plot varied from 5 to 36, and the estimated bryophyte cover ranged from 0.3 to 56.8% of the plot surface. Bryophyte cover is higher in coniferous forests, especially on siliceous bedrock with lower soil pH, which is consistent with Rola et al. (2021), who observed greater bryophyte cover on soils with more acidic pH and lower macronutrient content.
Bryophytes are good indicators of habitat conditions and show associations with different forest types (Stefańska-Krzaczek et al. 2022). The species composition and diversity of bryophytes differed significantly between the forest types in Slovenia with similar stand characteristics, tree species composition and edaphic conditions. The highest bryophyte species richness was observed in Group B_c, and the lowest species richness was observed in Group B_s. In both groups, Fagus sylvatica was the dominant tree species, while Acer pseudoplatanus in the first group and Castanea sativa Mill. in the second group were among the most abundant co-dominant tree species.
Among the most important factors influencing bryophyte species diversity are stand structure and tree species composition (Tinya et al. 2021), which are also significantly influenced by forest management. Forest management is an important factor causing changes in tree species composition and canopy closure, stand structure and also the amount of deadwood (Paillet et al. 2010; Czerepko et al. 2021). Heterogeneous stand structure and tree species composition support various groups of organisms, including bryophytes, in different ways.
In the studied plots in close-to-nature managed forests in Slovenia, tree species composition and bedrock were also important factors for bryophyte species diversity and composition. The species diversity and composition of bryophytes respond to tree species composition (e.g. broadleaves vs. coniferous). However, it appears that bedrock has an even more significant influence on bryophyte species richness, as significant changes in bryophyte species richness were observed along geological and edaphic gradients. In contrast to Rola et al. (2021), who observed significantly higher bryophyte species richness in plots with conifers than in plots with deciduous trees, a higher mean species richness per plot was found on average in broadleaved forests (19.9) than in coniferous forests (17.9) in Slovenia. The mean species richness per plot was significantly higher in forests on calcareous bedrock (23.2) than in forests on siliceous bedrock (13.9), confirming that edaphic conditions have a significant influence on the species richness of bryophytes in close-to-nature managed forests. The mean percentage of rocks and stones per plot area was 17.1% in forests on calcareous bedrock (B_c and C_c) and 2.9% in forests on siliceous bedrock (B_s and C_s). This may also imply variability in the different microsites on rocks/stones. The presence of rocks and stones on the ground surface may significantly increase the availability of different growth substrates and microhabitats for bryophytes, possibly affecting their composition, cover and diversity (Márialigeti et al. 2009; Király and Ódor 2010).
On average, the species richness of bryophytes in Group C_s was low. However, in some Pinus sylvestris forests of this group, we observed a very high cover of ground-dwelling (terricolous) bryophytes with a specific species composition as found in some other similar temperate forests in Central Europe (e.g. Tinya et al. 2021). These are mostly secondary pine forests on siliceous bedrock, which have developed on former sites of beech forests that have been significantly altered in the past for historical land use reasons. In these Pinus sylvestris forests in Slovenia, a dense layer of dwarf shrubs such as Vaccinium myrtillus L. and V. vitis-idaea L. grow in relatively open stands. In such open stands with pines dominating the overstory layer, light is the main determinant of the species composition of ground-dwelling bryophytes (Tinya et al. 2009, 2021; Jagodziński et al. 2018). It has been demonstrated that epiphytic and epixylic bryophytes are much less dependent on light conditions than soil-dwelling bryophytes.
Epiphytic bryophytes are much more sensitive to changes in humidity and temperature, requiring shadier conditions and constant humidity. Therefore, they may suffer from abrupt exposure to sunlight and lower humidity following intensive logging (Friedel et al. 2006). The high number of epiphytic bryophytes in Slovenian temperate forests (Supplementary Information 1) may be due to the high tree layer cover, especially in broadleaved forests, where cover ranges from 91.1 to 97.1%. A previous study demonstrated that the number of bryophyte species and their cover are high in forest types on moist sites (Stefańska-Krzaczek et al. 2022). Presumably, the relatively high humidity in the studied closed broadleaved stands has a positive effect on bryophyte diversity compared to Pinus sylvestris and P. nigra forests with much lower tree layer cover.
In contrast to Müller et al. (2019), bryophyte species richness in temperate Slovenian forests did not increase with an increasing proportion of conifers (19.9 species per plot in broadleaved vs. 17.9 in coniferous forests). In some studies (e.g. Goia and Gafta 2019), no consistent relationship was found with respect to bryophyte preference for beech or spruce deadwood, which are also the two dominant tree species in the forests studied in Slovenia. The pH of living and dead bark is also an important factor for the growth of epiphytic and epixylic bryophytes (Goia and Gafta 2019).
Less intensive forest management and silviculture in Slovenian temperate forests, applying close-to-nature, sustainable and multifunctional principles (ZGS 2022), may also contribute to the diversity of bryophyte species by creating forest stands with favourable microclimatic conditions and providing very diverse microsites for the growth of some rare and protected bryophyte species at the European (Bern Convention 1979; EU Habitats Directive 1992; Hodgetts et al. 2019) and national (Martinčič 2016) level. In the managed forests surveyed in Slovenia, Dicranum viride (Sull. & Lesq.) Lindb was identified in two plots. This species was also reported for some other managed forests and unmanaged forest reserves in Slovenia (Tomović et al. 2022), probably due to its preference for old, large-diameter beech trees (Kaufmann et al. 2018). Among the rare bryophyte species of national importance and conservation interest, our dataset also included Fissidens exilis Hedw. (Tomović et al. 2021). In addition to these two species, Leucobryum glaucum (Hedw.) Aangstr., which is listed in Article 17 and Annex V of the EU Habitats Directive (1992), was also common in the bryophyte material examined. Liverwort Lophozia ascendens (Warnst.) R.M. Schust., which is listed in Annex II of the Habitats Directive (1992), was found in only one investigated plot. This species is a good indicator or signal species of well-preserved and structured forest stands given that it is rarely seen and grows exclusively on decaying logs in advanced stages. Some species, such as the moss Haplocladium angustifolium (Hampe & Müll. Hal.) Broth., are Europe-wide data-deficient species (Hodgetts et al. 2019; Sabovljević et al. 2019) that are rare but not unexpected in Slovenian forest stands.
At the national level, there were some significant detections of rare, threatened or even regionally (nationally) extinct species in the surveyed plots. For example, Campylopus pyriforme (Schultz) Brid., which has not been reported in Slovenia since the nineteenth century (Regionally Extinct in Slovenia according to Martinčič 2016), and Stereodon pratensis (Spruce) Warnst. (Near Threatened in Slovenia according to Martinčič 2016) were reported during surveys of forest plots in Slovenia.
Close-to-nature and sustainable management practices, including selective cutting and small-scale shelterwood logging (ZGS 2022), promote bryophyte diversity by maintaining and increasing substrate and habitat variability, retaining coarse woody debris, increasing stand structural heterogeneity and maintaining favourable microclimatic conditions by retaining clusters of old, mature to over-mature trees in managed forests. Deadwood is also a key habitat in managed forests, determining not only the number of epixylic species, but also the overall richness of bryophyte assemblages (Fojcik et al. 2019). The availability of suitable substrates in forests is an important factor for bryophyte species richness (Müller et al. 2019).
Potential natural vegetation, well supported by close-to-nature and sustainable management, is highly beneficial for the maintaining bryophyte diversity (Rola et al. 2021). In contrast to the close-to-nature and sustainable management practices applied in the studied Slovenian forests, more intensive forest management decreases bryophyte richness. More intensive management may be particularly unfavourable for forest specialists, as they have a high affinity to late-successional developmental stages. Loss of suitable microhabitats, such as large trees, decreases bryophyte richness, especially in highly disturbed forests (Horvat et al. 2017).
Functional trait composition and diversity
In our study, the functional composition of bryophytes based on functional traits and their ecological indicator values (Bernhardt-Römermann et al. 2018; Simmel et al. 2021) was significantly influenced by bedrock and soil, but much less so by tree species composition. In the PCA ordination space, which is based on various bryophyte functional traits, axis 2 was associated with the bedrock characteristics of the plots studied. Thus, Groups B_i and B_c had significantly higher values on axis 2 than the groups of plots on siliceous bedrocks (B_s and C_s). However, no significant difference was found in the distribution of groups with different tree species composition in the PCA ordination, as the groups with different tree species composition were placed together (Groups B_s and C_s, and Groups B_c and C_c overlapped in the ordination space).
However, Żarnowiec et al. (2021) found that the differences in plant traits of bryophytes were related to the host tree, i.e. Fagus sylvatica and Picea abies. These two species are also the dominant tree species in the studied forests in Slovenia. The bryophyte species that preferred a more nutrient-rich substrate and alkaline environment were correlated with beech logs, while the species that preferred higher moisture content were correlated with spruce logs (Żarnowiec et al. 2021).
In terms of the functional trait composition of bryophytes, forests on intermediate and calcareous bedrocks differed from those on siliceous bedrocks. High values of the bryophyte indicator value for soil reaction were associated with forests on calcareous and intermediate bedrocks, and low values were associated with forests on siliceous bedrocks in the PCA ordination space (Fig. 9), so it can be assumed that bryophytes indicate relative differences in soil reaction in these forests.
Longer shoot lengths of bryophytes were associated with different forests on siliceous bedrocks, where tall grasses Calamagrostis arundinacea (L.) Roth and C. villosa (Chaix) J.F. Gmel., Molinia caerulea (l.) Moench subsp. arundinacea (Schrank) K. Richt, ferns Pteridium aquilinum (L.) Kuhn, Blechnum spicant (L.) Roth and berries Vaccinium myrtillus and V. vitis-idaea are common. In some of the study plots, the forest floor was almost completely covered by these species (Kermavnar and Kutnar 2020; Kermavnar et al. 2021a, 2021b). This is consistent with an experimental study of traits in which the bryophyte specific shoot length increased with increasing vegetation height and litter cover, indicating stronger competition from vascular plants (Van Zuijlen et al. 2021). Furthermore, a higher functional dispersion of bryophyte shoot length in forests on siliceous bedrocks was also observed in our study, especially in Group B_s.
On average, it appears that hemeroby, which indicates the intensity of human impact on the ecosystem through bryophytes, is higher in plots dominated by conifers on siliceous bedrocks (C_s) than in plots dominated by broadleaves on calcareous bedrocks (B_c). In contrast, the frequency of bryophyte species which are largely restricted to closed forests (forest1 in Fig. 9) is higher in Group B_c. In Group C_s, there are also some secondary stands of Picea abies and Pinus sylvestris that developed on the primary sites of broadleaved forests. In secondary spruce forests in particular, natural disturbances (e.g. bark beetle outbreaks, windthrows) followed by human interventions have been observed in recent decades (Kutnar et al. 2021), leading to significant changes in tree species composition and site characteristics in the forests studied. Changes in the dominant tree species in the forest stand can have a significant impact on bryophyte communities ( et al. 2021). Therefore, bryophytes can be expected to be sensitive to environmental changes caused by natural and/or human disturbances (Gignac 2001) and to respond to environmental gradients (Stefańska-Krzaczek et al. 2022) and can be considered to be good ecological indicators of forest naturalness (Sabovljević et al. 2010; Czerepko et al. 2021). A greater distance from the potential natural vegetation and tree species composition increases the frequency of bryophyte species that are less bound to closed and preserved forests, which also results in higher hemeroby.
The diversity of bryophyte life forms was found to be lower on siliceous bedrocks. The highest functional dispersion for this trait was found in coniferous forests on calcareous bedrock (C_c), whereas the lowest diversity was characterized by conifers on siliceous bedrock (C_s). It can be assumed that calcareous bedrock with higher cover and different rock microsites supports a higher diversity of bryophyte life forms than siliceous bedrock, and this difference was particularly evident in coniferous forests on different bedrocks, as this difference was significant.
The analysis of community-level traits for different groups revealed that bryophytes may be more affected by future global warming trends than vascular plants and lichens (Van Zuijlen et al. 2021). To a large extent, this could also apply to the Slovenian forests studied, especially those occurring on calcareous bedrock. Among them, forests on limestone bedrock are particularly sensitive to warming because of the development of shallow soils with low water storage capacity. Broadleaf forests on limestone (included in Group B_c), with various available substrates, have high bryophyte species richness and functional trait diversity.
Conclusions
In managed forests in Slovenia, high bryophyte diversity was found at the taxonomic level and at the functional trait level. In addition to the close-to-nature and sustainable forest management in Slovenia, the bedrock and tree species composition were also found to be important drivers of bryophyte species diversity and composition, and the functional diversity and composition of bryophytes were also significantly affected by bedrock and soil, but somewhat less so by tree species composition.
In order to maintain the high species richness and diverse composition of bryophytes in Slovenian forests, including their functional traits, it is recommended to continue close-to-nature and sustainable management practices, to keep forest stands as closed as possible under changing conditions and to avoid intensive management interventions. For conservation of bryophyte diversity, it is recommended to minimize intensive silvicultural practices in more preserved forests with higher bryophyte diversity and in more exposed forests on steep slopes.
References
Anderson MJ (2017) Permutational multivariate analysis of variance (PERMANOVA). In: Wiley StatsRef: statistics reference online, pp 1–15. Wiley
Andersson LI, Hytteborn H (1991) Bryophytes and decaying wood—a comparison between managed and natural forest. Ecography 14(2):121–130. https://doi.org/10.1111/j.1600-0587.1991.tb00642.x
Bates JW (1998) Is ‘life-form’ a useful concept in bryophyte ecology? Oikos 82:223–237. https://doi.org/10.2307/3546962
Bengtsson J, Nilsson SG, Franc A, Menozzi P (2000) Biodiversity, disturbances, ecosystem function and management of European forests. For Ecol Manage 132:39–50. https://doi.org/10.1016/S0378-1127(00)00378-9
Bernhardt-Römermann M, Poschlod P, Hentschel J, Chytrý M (2018) BryForTrait—a life-history trait database of forest bryophytes. J Veg Sci 29(4):798–800. https://doi.org/10.1111/jvs.2018.29.issue-410.1111/jvs.1264
Brugues M, Guerra J (2015) Flora Briofitica Iberica 2. Sociedad Espanola de Briologia/University of Murcia, Murcia
Bruun HH, Moen J, Virtanen R, Grytnes JA, Oksanen L, Angerbjörn A (2006) Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. J Veg Sci 17:37–46. https://doi.org/10.1111/j.1654-1103.2006.tb02421.x
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18:117–143
Bern Convention (1979) Convention on the conservation of European wildlife and natural habitats. Bern, https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A21979A0919%2801%29
Czerepko J, Gawryś R, Szymczyk R, Pisarek W, Janek M, Haidt A, Kowalewska A, Piegdoń A, Stebel A, Kukwa M, Cacciatori C (2021) How sensitive are epiphytic and epixylic cryptogams as indicators of forest naturalness? Testing bryophyte and lichen predictive power in stands under different management regimes in the Białowieża forest. Ecol Indic 125:107532. https://doi.org/10.1016/j.ecolind.2021.107532
de Mandiburu F (2021) Package “agricolae” – Statistical Procedures for Agricultural Research v. 1.3–5. Available on CRAN: https://cran.r-project.org/web/packages/agricolae/agricolae.pdf
de Vries W, Vel EM, Reinds GJ, Deelstra H, Klap JM, Leeters EEJM, Hendriks CMA, Kerkvoorden M, Landmann G, Herkendell J et al (2003) Intensive monitoring of forest ecosystems in Europe. 1. Objectives, set-up and evaluation strategy. For Ecol Manage 174:77–95. https://doi.org/10.1016/S0378-1127(02)00029-4
Düll R (1991) Zeigerwerte von Laub- und Lebermossen. In: Ellenberg H, Weber HE, Düll R, Wirth V, Werner W, Paulissen D (Eds), Zeigerwerte von Pflanzen in Mitteleuropa. Scr Geobot 18:175–214
During HJ (1979) Life strategies of bryophytes: a preliminary review. Lindbergia 5:2–18
During HJ (1992) Ecological classifications of bryophytes and lichens. In: Bates JW, Farmer AM (Eds), Bryophytes and lichens in a changing environment. Clarendon Press, Oxford, pp 1–31
EU Habitats Directive (1992) Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:31992L0043
Fojcik B, Wierzgoń M, Chmura D (2019) Response of bryophytes to disturbances in managed forests. A case study from a Polish forest. Cryptogam Bryol 40(10):105–118. https://doi.org/10.5252/cryptogamie-bryologie2019v40a10
Frego KA (2007) Bryophytes as potential indicators of forest integrity. For Ecol Manage 242:65–75. https://doi.org/10.1016/j.foreco.2007.01.030
Frey W, Frahm JP, Lobin W (2006) Liverworts, mosses and ferns of Europe. Harley, London, UK
Friedel A, von Oheimb G, Dengler J, Härdtle W (2006) Species diversity and species composition of epiphytic bryophytes and lichens—a comparison of managed and unmanaged beech forests in NE Germany. Feddes Repertorium 117:172–185. https://doi.org/10.1002/fedr.200511084
Fritz Ö, Brunet J (2010) Epiphytic bryophytes and lichens in Swedish beech forests –effects of forest history and habitat quality. Ecol Bull 53:95–107. http://www.jstor.org/stable/41442022
Gignac LD (2001) Bryophytes as indicators of climate change. Bryologist 104:410–420
Goia I, Gafta D (2019) Beech versus spruce deadwood as forest microhabitat: Does it make any difference to bryophytes? Plant Biosyst 153(2):187–194. https://doi.org/10.1080/11263504.2018.1448011
Grau O, Grytnes J, Birks H (2007) A comparison of altitudinal species richness patterns of bryophytes with other plant groups in Nepal, Central Himalaya. J Biogeogr 34:1907–1915. https://doi.org/10.1111/j.1365-2699.2007.01745.x
Guerra J, Brugues M, Cano MJ, Cros RM (2010) Flora BriofiticaIberica 4. Sociedad Espanolade Briologia/UniversityofMurcia, Murcia, pp 1–317
Guerra J, Cros RM (2007) Flora Briofitica Iberica 1. Sociedad Espanola de Briologia/University of Murcia, Murcia, pp 1–183
Guerra J, Cano MJ, Cros RM (2006) Flora Briofitica Iberica 3. Sociedad Espanola de Briologia/University of Murcia, Murcia, Spain, pp 1–305
Guerra J, Cano MJ, Brugues M (2014) Flora Briofitica Iberica 5. Sociedad Espanola de Briologia/University of Murcia, Murcia, Spain, pp 1–264
Guerra J, Cano MJ, Brugues M (2018) Flora Briofitica Iberica 6. Sociedad Espanola de Briologia/University of Murcia, Murcia, Spain, pp 1–463
Hill MO, Preston CD, Bosanquet SDS, Roy DB (2007) BRYOATT: attributes of British and Irish mosses, liverworts and hornworts. Centre for Ecology and Hydrology, Huntingdon
Hočevar S, Batič F, Martinčič A, Piskernik M (1980a) Drugotni nižinski pragozd Krakovo v Krakovskem gozdu. Zb Gozd Lesar 18:5–144 (In Slovenian with English Summary)
Hočevar S, Batič F, Martinčič A, Piskernik M (1980b) Mraziščni pragozd Prelesnikova koliševka. Zb Gozd Lesar 18:145–252 (In Slovenian with English Summary)
Hočevar S, Batič F, Martinčič A, Piskernik M (1980c) Panonska pragozdova Donačka gora in Belinovec. Zb Gozd Lesar 18:253–358 (In Slovenian with English Summary)
Hočevar S, Batič F, Martinčič A, Piskernik M (1985) Preddinarski gorski pragozdovi Trdinov vrh in Ravna gora na Gorjancih, Kopa v Kočevskem Rogu in Krokar na hrbtu pogorja Borovška gora – Planina nad Kolpa. Ljubljana, Strokovna in Znanstvena Dela 76:1–267 (In Slovenian with English Summary)
Hodgetts N, Calix M, Englefield E, Fettes N, Garcia Criado M, Patin L, Nieto A, Bergamini A, Bisang I, Baisheva E, Campisi P, Cogoni A, Hallingback T, Konstantinova N, Lockhart N, Sabovljevic M et al (2019) A miniature world in decline: European Red List of Mosses, Liverworts and Hornworts. IUCN, Brussels. https://doi.org/10.2305/IUCN.CH.2019.ERL.2.en
Hodgetts NG, Söderström L, Blockeel L, Caspari S, Ignatov MS, Konstantinova NA, Lockhart N, Papp B, Schröck C, Sim-Sim M, Bell D, Bell NE, Blom HH, Bruggeman-Nannenga MA, Brugués M, Enroth J, Flatberg KI, Garilleti R, Hedenäs L, Holyoak DT, Hugonnot V, Kariyawasam I, Köckinger H, Kučera J, Lara F, Porley RD (2020) An annotated checklist of bryophytes of Europe. Macaronesia and Cyprus J Bryol 42(1):1–116. https://doi.org/10.1080/03736687.2019.1694329
Horvat V, Heras P, García-Mijangos I, Biurrun I (2017) Intensive forest management affects bryophyte diversity in the western Pyrenean silver fir-beech forests. Biol Conserv 215:81–91. https://doi.org/10.1016/j.biocon.2017.09.007
Huggonot V, Chavoutier JL (2021) Les bryophytes de France (vol. 1): Anthocerotes et Hepatiques. MHM, Paris/ Biotope Editions, Meze. p. 1–652.
Husson F, Josse J, Le S, Mazet J (2020) Package ‘FactoMineR’ - Multivariate Exploratory Data Analysis and Data Mining. Available on CRAN: https://cran.r-project.org/web/packages/FactoMineR/FactoMineR.pdf
Jagodziński AM, Wierzcholska S, Dyderski MK, Horodecki P, Rusińska A, Gdula AK, Kasprowicz M (2018) Tree species effects on bryophyte guilds on a reclaimed post-mining site. Ecol Eng 110:117–127. https://doi.org/10.1016/j.ecoleng.2017.10.015
Kantvilas G, Jarman SJ, Minchin PR (2015) Early impacts of disturbance on lichens, mosses and liverworts in Tasmania’s wet eucalypt production forests. Aust Forestry 78:92–107. https://doi.org/10.1080/00049158.2015.1053025
Kassambara A, Mundt F (2020) Package “factoextra”—extract and visualize the results of multivariate data analyses. CRAN: https://cran.r-project.org/web/packages/factoextra/factoextra.pdf
Kaufmann S, Hauck M, Leuschner C (2018) Effects of natural forest dynamics on vascular plant, bryophyte, and lichen diversity in primeval Fagus sylvatica forests and comparison with production forests. J Ecol 106(6):2421–2434. https://doi.org/10.1111/1365-2745.12981
Kermavnar J, Kutnar L (2020) Patterns of understory community assembly and plant trait-environment relationships in temperate SE European forests. Diversity 12:91. https://doi.org/10.3390/d12030091
Kermavnar J, Kutnar L, Marinšek A (2021a) Disentangling the ecological determinants of species and functional trait diversity in herb-layer plant communities in European temperate forests. Forests 12(5):552. https://doi.org/10.3390/f12050552
Kermavnar J, Kutnar L, Marinšek A (2021b) Variation in floristic and trait composition along environmental gradients in the herb layer of temperate forests in the transition zone between Central and SE Europe. Plant Ecol 223:229–242. https://doi.org/10.1007/s11258-021-01203-8
Király I, Ódor P (2010) The effect of stand structure and tree species composition on epiphytic bryophytes in mixed deciduous–coniferous forests of Western Hungary. Biol Conserv 143:2063–2069. https://doi.org/10.1016/j.biocon.2010.05.014
Kriebitzsch W-U, Bültmann H, von Oheimb G, Schmidt M, Thiel H, Ewald J (2013) Forest-specific diversity of vascular plants, bryophytes, and lichens. In: Kraus D, Krumm F (Eds), Integrative approaches as an opportunity for the conservation of forest biodiversity. European Forest Institute, Joensuu, pp. 158–169
Kürschner H (1999) Life strategies of epiphytic bryophytes in Mediterranean Pinus woodlands and Platanus orientalis alluvial forests of Turkey. Cryptogam Bryol 20(1):17–33. https://doi.org/10.1016/S1290-0796(99)80004-2
Kutnar L (2011) Diversity of woody species on forest monitoring plots in Slovenia. GozdVestn 69:271–278 (In Slovenian with English Summary)
Kutnar L, Martinčič A (2001) Vegetacijske značilnosti izbranih pokljuških barij in okoliškega smrekovega gozda. Zb Gozd Lesar 64:57–104 (In Slovenian with English Summary)
Kutnar L, Martinčič A (2003) Ecological relationships between vegetation and soil-related variables along the mire margin-mire expanse gradient in the eastern Julian Alps, Slovenia. Ann Bot Fenn 40:177–189
Kutnar L, Martinčič A (2008) Bryophyte species diversity of forest ecosystems in Slovenia (Intensive Monitoring Programme). Zb Gozd Lesar 85:11–26 (In Slovenian with English Summary)
Kutnar L, Veselič Ž, Dakskobler I, Robič D (2012) Typology of Slovenian forest sites according to ecological and vegetation conditions for the purpose of forest management. GozdVestn 70:195–214 (In Slovenian with English Summary)
Kutnar L, Nagel TA, Kermavnar J (2019) Effects of disturbance on understory vegetation across Slovenian forest ecosystems. Forests 10:1048. https://doi.org/10.3390/f10111048
Kutnar L, Martinčič A (2002) Inicialna oblika barjanskega smrekovja Piceo-Sphagnetum flexuosi ass. nova v Sloveniji. Razpr. IV. razreda SAZU 43 (3):247–266 (In Slovenian with English Summary)
Kutnar L, Kermavnar J, Pintar AM (2021) Climate change and disturbances will shape future temperate forests in the transition zone between Central and SE Europe. Ann For Res 54(2):67–86. https://doi.org/10.15287/afr.2021.2111
Laliberté E, Legendre P, Shipley B (2015) Package “FD” - Measuring functional diversity (FD) from multiple traits, and other tools for functional ecology. Available on CRAN: https://cran.r-project.org/web/packages/FD/FD.pdf
Laliberté E, Legendre P (2010) A distance-based framework for measuring functional diversity from multiple traits. Ecology 91:299–305. https://doi.org/10.1890/08-2244.1
Lesica P, McCune B, Cooper SV, Hong WS (1991) Differences in lichen and bryophyte communities between old-growth and managed second-growth forests in the Swan Valley, Montana. Can J Botany 69:1745–1755. https://doi.org/10.1139/b91-222
Longton RE (1997) Reproductive biology and life-history strategies. Adv Bryol 6:65–101
Mägdefrau K (1968) Die Lebensformen Der Laubmoose Vegetatio 16:285–297
Márialigeti S, Németh B, Tinya F, Ódor P (2009) The effects of stand structure on ground-floor bryophyte assemblages in temperate mixed forests. Biodivers Conserv 18:2223–2241. https://doi.org/10.1007/s10531-009-9586-6
Martinčič A (2016) Updated Red List of bryophytes of Slovenia. Hacquetia 15:107–126
Martinčič A, Piskernik M (1985) Die Hochmoore Sloweniens: floristische, vegetationskundliche und ökologische Untersuchungen, Biološki vestnik, Vol. extraord. 1). Ljubljana: Slovene Biological Society
Mölder A, Schmidt M, Schönfelder E, Engel F, Schulz F (2015) Bryophytes as indicators of ancient woodlands in Schleswig-Holstein (Northern Germany). Ecol Indic 54:12–30. https://doi.org/10.1016/j.ecolind.2015.01.044
Müller J, Boch S, Prati D, Socher SA, Pommer U, Hessenmöller D, Schall P, Schulz ED, Fischer M (2019) Effects of forest management on bryophyte species richness in Central European forests. For Ecol Manage 432:850–859. https://doi.org/10.1016/j.foreco.2018.10.019
Nelson CR, Halpern CB (2005) Short-term effects of timber harvest and forest edges on ground-layer mosses and liverworts. Can J Bot 83:610–620. https://doi.org/10.1139/b05-036
Nordén B, Paltto H, Götmark F, Wallin K (2007) Indicators of biodiversity, what do they indicate? Lessons for conservation of cryptogams in oak-rich forest. Biol Conserv 135:369–379. https://doi.org/10.1016/j.biocon.2006.10.007
Ódor P, van Dort K, Aude E, Heilmann-Clausen J, Christensen M (2005) Diversity and composition of dead wood inhabiting bryophyte communities in European beech forests. Bol Soc Esp Briol 26–27:85–102
Ódor P, Heilmann-Clausen J, Christensen M, Aude E, van Dort KW, Piltaver A, Siller I, Veerkamp MT, Walleyn R, Standovár T, van Hees AFM, Kosec J, Matočec N, Kraigher H, Grebenc T (2006) Diversity of dead wood inhabiting fungi and bryophytes in semi-natural beech forests in Europe. Biol Cons 131(1):58–71. https://doi.org/10.1016/j.biocon.2006.02.004
Oksanen J, Blanchet G, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) Package “vegan” - Community Ecology Package v. 2.5–7. Available on CRAN: https://cran.r-project.org/web/packages/vegan/vegan.pdf
Paillet Y, Bergés L, Hjältén J, Ódor P, Avon C, Bernhardt-Römermann M, Bijlsma RJ, de Bruyn L, Fuhr M, Grandin U, Kanka R, Lundin L, Luque S, Magura T, Matesanz S, Mészáros I, Sebastià M, Schmidt W, Standovár T, Tóthmérész B, Uotila A, Valladares F, Vellak K, Virtanen R (2010) Biodiversity differences between managed and unmanaged forests: meta-analysis of species richness in Europe. Conserv Biol 24:101–112. https://doi.org/10.1111/j.1523-1739.2009.01399.x
Pardow A, Gehrig-Downie C, Gradstein R, Lakatos M (2012) Functional diversity of epiphytes in two tropical lowland rainforests, French Guiana: using bryophyte lifeforms to detect areas of high biodiversity. Biodivers Conserv 21(14):3637–3655. https://doi.org/10.1007/s10531-012-0386-z
Piskernik M, Martinčič A (1970) Vegetacija in ekologija gorskih barij v Sloveniji. Zb Gozd Lesar 8:131–203 (In Slovenian with English Summary)
R Core Team (2021). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rola K, Plášek V, Rożek K, Zubek S (2021) Effect of tree species identity and related habitat parameters on understorey bryophytes – interrelationships between bryophyte, soil and tree factors in a 50-year-old experimental forest. Plant Soil 466:613–630. https://doi.org/10.1007/s11104-021-05074-w
Sabovljević M, Vujičić M, Sabovljević A (2010) Diversity of saproxylic bryophytes in old-growth and managed beech forests in the Central Balkans. Plant Biosyst 144:234–240. https://doi.org/10.1080/11263500903561015
Sabovljević M, Papp B, Blockeel T, Ignatov M, Hallingbäck T, Söderström L. (2019) Haplocladium angustifolium (Europe assessment). The IUCN Red List of Threatened Species 2019: e.T87578048A87784853.
Schmidt M, Kriebitzsch WU, Ewald J (2011) Waldartenlisten der Farn- und Blütenpflanzen, Moose und Flechten Deutschlands. BfN-Skripten 299:1–111
Schofield WB, Hébant C (1984) The morphology and anatomy of the moss gametophore. In: Schuster RM (Ed), New manual of bryology. II. Hattori Botanical Laboratory, Nichinan, Japan, pp. 627–657
Simmel J, Ahrens M, Poschlod P (2021) Ellenberg N values of bryophytes in Central Europe. J Veg Sci 32:e12957. https://doi.org/10.1111/jvs.12957
Smith AJE (2004) Moss flora of Britain and Ireland, 2nd edn. Cambridge University Press, Cambridge, UK
Spitale D, Mair P, Nascimbene J (2020) Patterns of bryophyte life-forms are predicable across land cover types. Ecol Indic 109:105799. https://doi.org/10.1016/j.ecolind.2019.105799
Staniaszek-Kik M, Chmura D, Żarnowiec J (2019) What factors influence colonization of lichens, liverworts, mosses and vascular plants on snags? Biologia 74(4):375–384. https://doi.org/10.2478/s11756-019-00191-5
Stefańska-Krzaczek E, Swacha G, Żarnowiec J, Raduła MW, Kącki Z, Staniaszek-Kik M (2022) Central European forest floor bryophytes: Richness, species composition, coexistence and diagnostic significance across environmental gradients of forest habitats. Ecol Indic 139:108954. https://doi.org/10.1016/j.ecolind.2022.108954
Stokland JN, Siitonen J, Jonsson BG (2012) Biodiversity in Dead Wood. Cambridge University Press, Cambridge, UK
Sun S, Wu Y, Wang G, Zhou J, Yu D, Bing H, Luo J (2013) Bryophyte species richness and composition along an altitudinal gradient in Gongga Mountain. China. Plos One 8:e58131. https://doi.org/10.1371/journal.pone.0058131
Tinya F, Ódor P (2016) Congruence of the spatial pattern of light and understory vegetation in an old-growth, temperate mixed forest. For Ecol Manage 381:84–92. https://doi.org/10.1016/j.foreco.2016.09.027
Tinya F, Márialigeti S, Király I, Németh B, Ódor P (2009) The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in Őrség, Western Hungary. Plant Ecol 204:69–81. https://doi.org/10.1007/s11258-008-9566-z
Tinya F, Kovács B, Bidló A, Dima B, Király I, Kutszegi G, Lakatos F, Mag Z, Márialigeti S, Nascimbene J, Samu F, Siller I, Szél G, Ódor P (2021) Environmental drivers of forest biodiversity in temperate mixed forests – A multi-taxon approach. SCI TOTAL ENVIRON Sci Total Environ 795:148720. https://doi.org/10.1016/j.scitotenv.2021.148720
Tomović G, Sabovljević MS, Denchev TT, Denchev CM, Niketić M, Boycheva P, Ivanov D, Šabanović E, Djordjević V, Kutnar L, Ștefănuţ S, Pantović J, Grdović S, Kuzmanović N, Mašić E, Lazarević P (2021) New records and noteworthy data of plants, algae and fungi in SE Europe and adjacent regions, 4. Botanica Serbica 45(1):129–136. https://doi.org/10.2298/BOTSERB2101129T
Tomović G, Sabovljević MS, Niketić M, Boycheva P, Lazarević P, Kabaš E, Vidaković D, Krizmanić J, Kutnar L, Kermavnar J, Veljković M, Djordjević V, Assyov B, Bozok F, Strgulc Krajšek S, Lobnik Cimerman Ž, Ștefănuţ S, Bîrsan C-C, Hajrudinović-Bogunić A, Begić A (2022) New records and noteworthy data of plants, algae and fungi in SE Europe and adjacent regions, 8. Botanica Serbica 46(1):133–141. https://doi.org/10.2298/BOTSERB2201133T
Urbančič, M., Simončič, P., Prus, T., Kutnar, L. (2005) Forest Soil Atlas of Slovenia. Ljubljana: Zveza gozdarskih društev Slovenije: Gozdarski vestnik: Gozdarski inštitut Slovenije, 100 p. (In Slovenian with English Summary)
Urbančič M, Kutnar L, Kralj T, Kobal M, Simončič P (2009) Site characteristics of permanent plots on the Slovenian 16 km × 16 km net. GozdVestn 67:17–48 (In Slovenian with English Summary)
Urbančič M, Kutnar L, Kobal M, Žlindra D, Marinšek A, Simončič P (2016) Soil and vegetation characteristics on Intensive Monitoring Plots of forest ecosystems. GozdVestn 74:3–27 (In Slovenian with English Summary)
Van Zuijlen K, Klanderud K, Dahle OS, Hasvik Å, Knutsen MS, Olsen SL et al (2021) Community-level functional traits of alpine vascular plants, bryophytes, and lichens after long-term experimental warming. Arctic Science 8(3):843–857. https://doi.org/10.1139/as-2020-0007
Vieira C, Séneca A, Sérgio C, Ferreira MT (2012) Bryophyte taxonomic and functional groups as indicators of fine scale ecological gradients in mountain streams. Ecol Indic 18:98–107. https://doi.org/10.1016/j.ecolind.2011.10.012
Vittoz P, Camenisch M, Mayor R, Miserere L, Vust M, Theurillat J-P (2010) Subalpine-nival gradient of species richness for vascular plants, bryophytes and lichens in the Swiss Inner Alps. Bot Helv 120(2):139–149. https://doi.org/10.1007/s00035-010-0079-8
WorldClim (2021) Global climate and weather data. https://www.worldclim.org/
Yan XL, Bao WK, Pang XY, Zhang NX, Chen J (2013) Regeneration strategies influence ground bryophyte composition and diversity after forest clearcutting. Ann for Sci 70(8):845–861. https://doi.org/10.1007/s13595-013-0323-7
Żarnowiec J (1996) The bryoflora of urban areas – a floristic-ecological case study of Oświęcim town (S Poland). Fragm Flor Geobot 41(1):355–377
Żarnowiec J, Staniaszek-Kik M, Chmura D (2021) Trait-based responses of bryophytes to the decaying logs in Central European mountain forests. Ecol Indic 126:107671. https://doi.org/10.1016/j.ecolind.2021.107671
Żarnowiec J (1995) Bryopsida. In: Faliński JB, Mułenko W (Eds), Cryptogamous plants in the forest communities of Białowieża National Park. General problems and taxonomic groups analysis (Project CRYPTO). Phytocoenosis vol. 7 (N. S.), Archivum Geobotanicum 4:47–61
ZGS (2022) Zavod za gozdove Slovenije (Slovenia Forest Service) http://www.zgs.si/
Acknowledgements
This study was performed as a part of the BioSoil Demonstration project and the Intensive Monitoring Programme in Slovenia (both part of EU Programme ICP Forests). The identification of bryophytes and finalization of the study was supported by the EU Project LIFE Integrated Project for Enhanced Management of Natura 2000 in Slovenia (LIFE17 IPE/SI/000011). We would like to thank Primož Simončič, Daniel Žlindra, Erika Kozamernik, Matej Rupel and many other colleagues from the Slovenian Forestry Institute for their assistance, as well as many experts from the Slovenia Forest Service. We thank Andrej Martinčič for assistance in identification of bryophytes and Philip J. Nagel for proofreading and language editing.
Funding
The research was funded by the Slovenian Ministry of Agriculture, Forestry and Food (JGS task 1/3) and by the Slovenian Research Agency (research core funding No. P4-0107 and grant number Z4-4543). The identification of bryophytes and finalization of the study was funded by the EU Project LIFE Integrated Project for Enhanced Management of Natura 2000 in Slovenia (LIFE17 IPE/SI/000011).
Author information
Authors and Affiliations
Contributions
Conceptualization was done by L.K., J.K. and M.S.S.; the methodology was prepared by L.K., J.K. and M.S.S.; formal analysis was performed by L.K. and J.K.; data curation was done by L.K. and J.K.; writing—original draft preparation was done by L.K., J.K. and M.S.S.; funding acquisition was done by L.K.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Communicated by Gediminas Brazaitis.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kutnar, L., Kermavnar, J. & Sabovljević, M.S. Bryophyte diversity, composition and functional traits in relation to bedrock and tree species composition in close-to-nature managed forests. Eur J Forest Res 142, 865–882 (2023). https://doi.org/10.1007/s10342-023-01560-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-023-01560-1