
Abstract
The intensification of agricultural land-use, abandonment and afforestation have caused severe loss and degradation of nutrient-poor, semi-natural grasslands across Europe. Calcareous grasslands have an outstanding value for nature conservation due to their highly diverse flora and fauna, including birds. However, knowledge of environmental factors driving the composition of bird communities in calcareous grasslands is still scarce. The aim of our study was to compare the breeding-bird assemblages of calcareous grasslands and the second most common semi-natural grassland type in the study area (Diemel Valley, Central Germany), mesic grasslands, along an elevation (climate) gradient. For each grassland type, we randomly selected 27 square-shaped plots with a size of 5 ha each and a cover of the focal grassland type of at least 50% within the plot. Our study revealed that both the number and density of threatened species of breeding birds were higher in plots of calcareous grasslands than in those of mesic grasslands. Based on multivariable generalised linear mixed-effects models, the most important predictors of overall species richness and density of breeding birds in plots of calcareous grasslands were the area of juniper-rich calcareous grassland and Shannon habitat diversity. By contrast, the number and density of threatened species were in both grassland types mainly driven by breeding-season temperature. Heterogeneous, juniper-rich calcareous grasslands seem to be a key habitat for bird assemblages with high species richness and density. These grasslands usually consisted of small-scale habitat mosaics providing suitable nesting sites and foraging grounds in close proximity. By contrast, mesic grasslands had rather homogeneous swards and shrubs or trees were normally restricted to the edges of the grassland patches. Based on our study, increasing habitat heterogeneity at the landscape scale and within grasslands is the key to enhancing species richness and density of breeding birds.
Zusammenfassung
Bedeutung des Lokalklimas und der Habitatheterogenität für Brutvogelgemeinschaften des Magergraslandes
Die Intensivierung und Aufgabe der Landnutzung sowie Aufforstungen haben zu einem starken Flächenrückgang und einer Abnahme der Habitatqualität des Magergraslands in Europa geführt. Kalkmagerrasen haben eine herausragende Bedeutung für den Naturschutz aufgrund ihrer artenreichen Flora und Fauna, inklusive der Vogelwelt. Unser Wissen über die Faktoren, die die Zusammensetzung der Vogelgemeinschaften in Kalkmagerrasen bestimmen ist aber immer noch gering. Das Ziel unserer Studie war es, die Zusammensetzung der Brutvogelgemeinschaften in Kalkmagerrasen und dem zweithäufigsten, nährstoffarmen Graslandtyp im Untersuchungsgebiet (dem Diemeltal, Deutschland) – frischem Magergrasland – entlang eines Höhen- bzw. Klimagradienten zu untersuchen. Für jeden Magergraslandtyp wurden 27 quadratische Probeflächen (PF) mit einer Größe von 5 ha und einem Anteil des namengebenden Graslandtyps von mindestens 50% zufällig ausgewählt. Unsere Studie zeigte, dass sowohl die Artenzahl als auch die Dichte an gefährdeten Brutvogelarten in PF der Kalkmagerrasen höher war als in denen des frischen Magergraslands. In multivariablen, generalisierten linearen gemischten Modellen der PF der Kalkmagerrasen waren die Fläche an wacholderreichen Kalkmagerrasen und der Shannon-Habitatdiversitätsindex die wichtigsten Prädiktoren für die Artenzahl und Dichte der Brutvogelarten insgesamt. Im Gegensatz dazu hing die Artenzahl und Dichte gefährdeter Brutvogelarten in beiden Magergraslandtypen vor allem von der Temperatur zur Brutzeit ab. Heterogene, wacholderreiche Kalkmagerrasen scheinen Schlüsselhabitate für arten- und individuenreiche Brutvogelgemeinschaften zu sein. Sie wiesen normalerweise kleinräumige Habitatmosaike auf, die aus geeigneten Brutplätzen und Nahrungshabitaten in unmittelbarer Nachbarschaft bestanden. Im Gegensatz dazu war das frische Magergrasland eher durch eine homogene Grasnarbe gekennzeichnet und Sträucher oder Bäume waren vor allem am Rand der Parzellen vorhanden. Basierend auf unserer Studie ist die Erhöhung der Habitatheterogenität auf der Landschaftsebene und innerhalb des Magergraslandes der Schlüssel zur Förderung arten- und individuenreicher Brutvogelgemeinschaften.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Most of Europe’s biodiversity is associated with agricultural land (Donald et al. 2006; Henle et al. 2008; Kleijn et al. 2009). For centuries, traditional farmland-management practices have contributed to the creation of semi-natural ecosystems hosting an outstanding biodiversity (Plieninger et al. 2006, 2015; Löffler and Fartmann 2017; Poschlod 2017). However, since the beginning of the industrial era, land-use change has led to a dramatic loss of wild biota in agricultural habitats (Donald et al. 2006; Bengtsson et al. 2019). Consequently, maintaining farmland biodiversity has become increasingly difficult throughout Europe (Donald et al. 2006; Flohre et al. 2011).
The intensification of agricultural land-use, abandonment and afforestation have caused severe loss and degradation of nutrient-poor, semi-natural grasslands across Europe (Wallis De Vries et al. 2002; Baur et al. 2006; Löffler et al. 2020). As a result, large-scale grasslands of conservation concern, such as calcareous grasslands, are now mainly restricted to mountain ranges, that are usually characterised by a low land-use intensity (MacDonald et al. 2000; Plieninger et al. 2006). Calcareous grasslands have an outstanding value for nature conservation due to their highly diverse flora and insect fauna (Poschlod and Wallis De Vries 2002; van Swaay 2002). Therefore, calcareous grasslands in general and those with occurrence of juniper (Juniperus communis) shrubs are now protected under the EU Habitats Directive (EC 2007). Previous studies on the biodiversity of calcareous grasslands have primarily concentrated on plants and insects (e.g., Poschlod and Wallis De Vries 2002; Poniatowski and Fartmann 2008; Krämer et al. 2012; Helbing et al. 2021). Recent work revealed that these grasslands may also play a vital role as refuges for species-rich bird assemblages in Central European landscapes (Köhler et al. 2016; Ernst et al. 2017). However, knowledge of environmental factors driving the composition of bird assemblages in calcareous grasslands is still scarce and sometimes findings have even been contradictory. While Köhler et al. (2016) showed that year-round grazing favoured species richness in calcareous grasslands, Ernst et al. (2017) detected a higher richness and abundance of farmland birds in abandoned calcareous grasslands. In comparison to calcareous grasslands, our knowledge on the biodiversity of mesic grasslands is even worse since they suffered more severely from habitat loss due agricultural intensification (Poschlod 2017; Fartmann et al. 2021).
Birds are excellent indicators of farmland biodiversity and human-driven habitat alterations in semi-natural grasslands (Fuller 2012; Newton 2017; Reif and Hanzelka 2020). In particular, specialised species are highly sensitive to land-use change and, as a consequence, have frequently become threatened in recent decades (Reif et al. 2008; Correll et al. 2019; Keller et al. 2020). They are mainly affected by changes in food supply and breeding habitats caused by altered farmland-management practices (Benton et al. 2002; Newton 2004). Additionally, temperature is another important driver of bird species richness and density (Pearce-Higgins and Green 2014; Keller et al. 2020; Fumy and Fartmann 2021).
Our study area, the Diemel valley in Central Germany (Fig. 1), is rich in nutrient-poor semi-natural grasslands with low land-use intensity (Fartmann 2004). For calcareous grasslands, it is even the most important stronghold in the northern half of Germany. The aim of our study was to compare the breeding-bird assemblages of calcareous grasslands and the second most common semi-natural grassland type in the study area, mesic grasslands (Fig. 2). We compared the species richness and density of breeding birds as well as environmental conditions between the two grassland types along the elevation (climate) gradient of the study area. Moreover, we assessed indicator species for each grassland type by their relative abundance and relative frequency using an indicator-species analysis. Key drivers of bird diversity and density were identified by generalised linear mixed-effects models to derive evidence-based measures for future habitat management in calcareous and mesic grasslands.

Location of the Diemel Valley and plots in Central Germany

Schematic drawing of typical stands of the two focal grassland types, calcareous (a) and mesic grasslands (foreground) (b), and juniper shrub (Juniperus communis) (c) as a characteristic keystone structure in calcareous grasslands of the study area. Drawing: M. Freienstein
Materials and methods
Study area
The study was carried out in the Diemel Valley (100–600 m a.s.l.; Central Germany), which has an area of about 460 km2 along the border between the federal states of North Rhine-Westphalia and Hesse (51° 22′ N/8° 38′ E and 51° 38′ N/9° 25′ E; Fig. 1). The study area covers ~ 750 ha of calcareous and ~ 250 ha of mesic grasslands (Fartmann 2004). The climate is suboceanic (Müller-Wille 1981). The surrounding matrix of these semi-natural grasslands is dominated by a patchwork of woodland, improved grassland and arable fields (Poniatowski and Fartmann 2010). In accordance with the elevation gradient in the study area, annual precipitation increases with elevation from 732 to 948 mm, while annual temperature decreases from 8.8 to 7.9 °C (long-term mean: 1981–2010; weather station Warburg and Brilon, respectively; German Meteorological Service 2021). To account for possible spatial autocorrelation, the study area was divided into seven subareas according to the landscape configuration (Fig. 1).
Sampling design
Plots
We studied two types of semi-natural grasslands with low land-use intensity: (i) calcareous and (ii) mesic grasslands (Fig. 2). For each grassland type, we randomly selected 27 square-shaped plots with a size of 5 ha and a cover of the focal grassland type of at least 50% within the plot (cf. Fartmann et al. 2018) across the elevation (climate) gradient of the study area (N = 54).
Environmental conditions
We used digital elevation models with a spatial resolution of 4 ha to calculate the mean elevation of the plots (Geobasis 2021). Climate data were derived from grid maps with a spatial resolution of 1 km2 containing information on long-term averages of temperature and precipitation (period 1981–2010, German Meteorological Service 2021). For further analyses, we considered both mean annual values and mean values for the breeding season (March–August).
For each plot, we recorded the habitat composition in the field according to the German habitat classification scheme (scale: 1:1000; 15 major habitat types (Table 1); Finck et al., 2017). Later we digitised the habitat maps using ArcGIS 10.5. Additionally, we used these data to calculate the Shannon habitat diversity (H') of each plot (Fartmann et al. 2018; Schwarz et al. 2018):
with pi = ni/N and where N is the area of all habitat types per plot and ni is the area of each habitat type in the plot.
Breeding-bird surveys
We mapped breeding-bird territories in all plots from the beginning of April to July 2019 (Fischer et al. 2005). Territory mapping is among the most accurate methods for estimating species richness and density of breeding birds (Bibby et al. 2000). Altogether, we performed five surveys early in the morning with an interval of at least 10 days between each visit (Fartmann et al. 2018; Schwarz et al. 2018). During every survey, we followed a non-linear route covering the whole plot and noted all signs of territorial behaviour, such as singing, on a map (scale 1:1000) (Bibby et al. 2000). Breeding was assumed if a bird showed territorial behaviour twice within a period of 10 days between each visit (Fischer et al. 2005). We counted single observations as a territory only if breeding (e.g., nestlings in a nest) was documented (Schmidt et al. 2022). Prior to further analyses, we classified breeding-bird species as threatened (including near-threatened species) according to the red-data books of breeding birds in North Rhine-Westphalia (Grüneberg et al. 2017) and Hesse (Wolf and Widdig 2016).
Statistical analysis
We performed all statistical analyses using R 3.6.1 (R Core Team 2021). For detecting significant differences in environmental parameters (Table 1) as well as in species richness and territory density between the grassland types (Fig. 3), we applied univariable generalised linear mixed-effects models (GLMM) (R packages lme4, Bates et al. 2020) with ‘subarea’ as a random factor (Crawley 2007). Grassland type served as a nominal fixed factor, and we used the analysed parameters as dependent variables. We conducted negative binomial GLMMs for reducing overdispersion. We analysed the overall effect of grassland type on the environmental parameters by comparing the full models with reduced models without ‘grassland type’ as the fixed factor (intercept-only models) and applying likelihood-ratio tests.

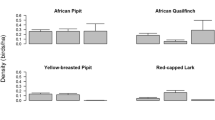
Mean values (± SE) of species richness and density of all (a, b) and threatened (c, d) breeding-bird species in plots of calcareous and mesic grasslands (N = 54). Differences between the two grassland types were analysed using GLMM with ‘subarea’ as a random factor. a Z = − 1.891, P < 0.05, b Z = − 2.013, P < 0.05, c Z = − 1.951, P < 0.05, d Z = − 2.506, P < 0.01. *P < 0.05; **P < 0.01
We calculated multivariable GLMMs (negative binomial error structure) to detect environmental parameters that explain the species richness and territory density of breeding-bird assemblages, separately for all and threatened species, in the two grassland types. To avoid model over-fitting, we excluded intercorrelated (|rs|> 0.5) variables and we used only the ecologically most meaningful variable in GLMMs (Tables A1 and A2) (cf. Löffler and Fartmann 2017). Therefore, for the plots of calcareous grasslands, we excluded elevation, annual precipitation and temperature, open and blackthorn-rich calcareous grasslands, shrubberies and broad-leaved forests from the GLMM analyses. For the plots of mesic grasslands, we eliminated elevation, precipitation (annual and breeding season), annual temperature, arable land, improved meadows and hedges/copses. For all GLMMs, the variable ‘subarea’ was used as a random factor (Crawley 2007). To increase model robustness and identify the most important environmental parameters, we conducted model averaging based on an information-theoretic approach (Burnham and Anderson 2002; Grueber et al. 2011). Model averaging was conducted using the dredge function (R package MuMIn, Bartón 2020) and included only top-ranked models within ΔAICC < 3 (Grueber et al. 2011). The model-averaged regression coefficients were calculated from a range of top-ranked submodels within ΔAICC < 3. This means that the averaged coefficients represent a ‘weighted average’ (based on AICc weights) across all submodels. For more details on the calculation see Burnham and Anderson (2002; Eq. 4.7). Non-significant predictors were also included in some of the top-ranked submodels as they can increase model accuracy. Therefore, they were also retained in the averaged models.
To identify indicator species for each grassland type an indicator-species analysis (ISA) (Dufrêne and Legendre 1997; Cáceres and Jansen 2016) was carried out. ISA uses the relative abundance and relative frequency of a species to estimate the strength of its association with the two grassland types. The more the indicator value (IV; scaled from 0 to 1) of a species increases, the more abundant is a species and the more frequently it occurs compared with other species in the focal grassland type. The statistical significance of this relationship is tested using a permutation test. This method is a common statistical tool in community ecology and is frequently used to identify characteristic species of certain habitat types (e.g. Thorn et al. 2016; Fartmann et al. 2018).
Results
Environmental conditions
In contrast to climatic conditions, habitat composition differed between plots of calcareous and mesic grasslands (Table 1). Each grassland type was dominated by its respective eponymous grassland (open, blackthorn-rich and juniper-rich calcareous grasslands vs. mesic meadows and pastures). Additionally, plots of calcareous grasslands had a larger area of coniferous forests than plots of mesic grasslands. By contrast, the area of hedges/copses and broad-leaved forests were larger in plots of mesic grasslands. All other habitat parameters did not differ.
Breeding-bird assemblages
Altogether, we detected 67 breeding bird species on the 54 plots (Table A3). Among them were 23 threatened species. The most common species (i.e., > 100 territories on all plots), in decreasing order, were Yellowhammer (Emberiza citrinella), Eurasian Blackcap (Sylvia atricapilla), Chiffchaff (Phylloscopus collybita), Great Tit (Parus major), Blackbird (Turdus merula), Whitethroat (Sylvia communis), Chaffinch (Fringilla coelebs) and Robin (Erithacus rubecula). The most common threatened species (i.e., > 30 territories on all plots) were Yellowhammer, Willow Warbler (Phylloscopus trochilus), Lesser Whitethroat (Sylvia curruca), Linnet (Carduelis cannabina), Red-backed Shrike (Lanius collurio) and Tree Pipit (Anthus trivialis).
Relationship between breeding-bird assemblages and environmental conditions
Both the number and density of all and threatened species were higher in plots of calcareous grasslands than in those of mesic grasslands (Fig. 3). Altogether, we identified eight indicator species of calcareous grasslands; three of them, Linnet, Tree Pipit and Willow Warbler, were considered threatened (Table 2). A least four of the indicator species regularly used juniper for breeding: Bullfinch (Pyrrhula pyrrhula), Linnet, Long-tailed Tit (Aegithalos caudatus) and Song Thrush (Turdus philomelos) (own observation). By contrast, mesic grasslands had no indicator species.
In the multivariable GLMM analyses, the overall number and density of breeding bird species increased in plots of calcareous grasslands with the area of juniper-rich calcareous grasslands and Shannon habitat diversity (Table 3, Fig. 4). By contrast, the number and density of threatened species increased in calcareous and mesic grasslands with breeding-season temperature (Tables 3 and 4, Figs. 4 and 5). Additionally, in plots of mesic grasslands, the overall density increased with breeding-season temperature and overall species richness increased with the area of coniferous forest (Table 4, Fig. 5).

Relationship between species richness and density of all and threatened breeding bird species, respectively, and the significant environmental parameters of averaged models for plots of calcareous grasslands (N = 27) (see Table 3). The regression slopes (including 95% confidence intervals) were fitted using a single predictor GLMM (negative binomial error structure)

Relationship between species richness and density of all and threatened breeding-bird species, respectively, and the significant environmental parameters of averaged models for plots of mesic grasslands (N = 27) (see Table 4). The regression slopes (including 95% confidence intervals) were fitted using a single predictor GLMM (negative binomial error structure)
Discussion
Our study revealed that both the number and the density of all and threatened species of breeding birds were higher in plots of calcareous grasslands than in those of mesic grasslands. Based on the multivariable GLMMs, the most important predictors of overall species richness and density of breeding birds in plots of calcareous grasslands were the area of juniper-rich calcareous grassland and Shannon habitat diversity. By contrast, the number and density of threatened species were in both grassland types mainly driven by breeding-season temperature.
Since diverse habitats offer more available niches, habitat heterogeneity is known to be an important driver of species richness and density in breeding-bird assemblages (Benton et al. 2003; Vickery and Arlettaz 2012; Farwell et al. 2020). Shannon habitat diversity based on habitat types is a rather coarse proxy for habitat heterogeneity, usually not reflecting the sometimes high three-dimensional structural diversity within a certain habitat type (cf. Cooper et al. 2020). Nevertheless, we detected a positive effect of Shannon habitat diversity on both overall species richness and density of breeding birds in plots of calcareous grasslands. Additionally, the area of juniper-rich calcareous grasslands was another important predictor of species richness and density of breeding birds in plots of this grassland type. While many plant and insect taxa of conservation concern prefer open habitat structures rich in bare ground (Bourn and Thomas 2002; Fartmann et al. 2012; Poniatowski et al. 2020), juniper-rich calcareous grasslands seem to be key habitats for bird assemblages with high species richness and density. In general, birds depend on (i) suitable foraging habitats with high prey accessibility and (ii) breeding habitats that provide shelter from potential nest predators (Benton et al. 2003; Moorcroft et al. 2002; Fartmann et al. 2018). Prey accessibility in grassland birds is granted by patches of low-growing vegetation and bare ground (Tagmann-Ioset et al. 2012; Kämpfer and Fartmann 2019; Fumy and Fartmann 2021). Taller vegetation is thought to reduce the predation risk of eggs and chicks in ground nests (Baines 1990; Schwarz et al. 2018). Dense and especially evergreen shrubs, such as the juniper (Fig. 2), offer sheltered nesting sites for shrub-breeding birds, exposed song posts and hunting perches (Gatter 2000; Schwarz et al. 2018). The calcareous grasslands in the study area are characterised by an extraordinarily high three-dimensional structural diversity (Poniatowski and Fartmann 2008; Poniatowski et al. 2018) (cf. Fig. 2). This is especially true for the juniper-rich ones. They consist of small-scale mosaics of sparsely-vegetated swards, taller vegetation and interspersed juniper shrubs (Fartmann 2004) (Fig. 2) that provide all the vital microhabitats for birds mentioned before (cf. Berg and Part 1994; Kujawa and Tryjanowski 2000).
By contrast, mesic grasslands were usually characterised by rather homogeneous swards (especially in meadows; own observation), although land-use intensity was low. Plots of mesic grasslands included larger areas of hedges and copses (cf. also Fig. 2), which also offer potential nesting sites, song posts or hunting perches. However, these habitat elements were mostly restricted to the edges of the grassland patches limiting the number of territories of breeding birds.
In particular, specialised bird species depend on habitat heterogeneity (Fuhlendorf et al. 2006; Reif et al. 2008; Correll et al. 2019). Hence, the high number of indicator species in plots of calcareous grasslands can likely also be attributed to the higher habitat heterogeneity of calcareous, especially juniper-rich calcareous grasslands compared with mesic grasslands.
Apart from habitat heterogeneity, temperature is another important driver of bird species richness and density (Pearce-Higgins and Green 2014; Keller et al. 2020). In the rather homogeneous mesic grasslands of the study area it was even the most important one. Overall species richness and the number and density of threatened species increased in plots of mesic grasslands with breeding-season temperature (= lower elevations, cf. Table A2). The same was true for the number and density of threatened species in plots of calcareous grasslands. Several thermophilic bird species detected in our study reach their regional upper distribution limit in the Diemel Valley. For example, the main distribution of Cuckoo (Cuculus canorus), Eurasian Turtle Dove (Streptopelia turtur), Green Woodpecker (Picus viridis), Nightingale (Luscinia megarhynchos) and Wryneck (Jynx torquilla) are largely restricted to lower elevations in the study area (cf. Grüneberg et al. 2013). Except Green Woodpecker and Nightingale all are considered threatened (Table A3). Moreover, in plots of mesic grasslands breeding-season temperature was intercorrelated with the area of hedges and copses (Table A2). As a result, the observed relationship also reflects a higher availability of potential nesting sites, song posts or hunting perches due to more hedges and copses in plots of mesic grasslands characterised by higher temperatures and lower elevations.
The area of coniferous forests was a further predictor of overall species richness of breeding birds in plots of mesic grasslands. Among the detected breeding-bird species there were several species that strongly depend on coniferous forests, such as Coal Tit (Periparus ater), Crested Tit (Lophophanes cristatus), Firecrest (Regulus ignicapilla), Goldcrest (Regulus regulus), or Treecreeper (Certhia familiaris), explaining this pattern (Table A3; Gatter 2000; Fartmann et al. 2018). To exclude potential effects of other habitat types such as coniferous forest on bird species composition, further studies should use plots that only consist of one of the two considered grassland types.
In conclusion, (i) habitat heterogeneity and (ii) warmer climatic conditions fostered species richness and density of breeding-bird species in semi-natural grasslands along an elevation (climate) gradient. Plots of calcareous grasslands had a high species richness and density. Especially the heterogeneous, juniper-rich calcareous grasslands seem to be a key habitat for bird assemblages with high species richness and density. These grasslands usually consisted of small-scale habitat mosaics providing suitable nesting sites and foraging grounds in close proximity. By contrast, mesic grasslands had rather homogeneous swards and shrubs or trees were normally restricted to the edges of the grassland patches.
Implications for conservation
Based on our study, increasing small-scale habitat heterogeneity within calcareous and mesic grasslands is the key to promoting richness and density of bird species. Calcareous grasslands are hotspots of plant and insect diversity (Wallis De Vries et al. 2002; Poniatowski and Fartmann 2008; Krämer et al. 2012). As our study showed, they are also an important habitat for birds. Much of the plant and insect diversity, especially of threatened species, is associated with early and mid-successional stages of calcareous grasslands (Bourn and Thomas 2002; Fartmann et al. 2012; Poniatowski et al. 2020). Consequently, conservation management in calcareous grasslands aims to stop succession and to retain open swards in favour of specialised plant and insect species (Dekoninck et al. 2007; Poniatowski et al. 2018). For future management, habitat preferences of threatened species of breeding birds should be considered as well. Especially in areas where a continuous management (usually grazing) cannot be sustained, the successional development towards semi-open juniper-rich calcareous grasslands should be supported in favour of threatened breeding birds. Such heterogeneous habitats have generally become scarce throughout Central European landscapes with negative effects on biodiversity at the landscape scale (Diacon-Bolli et al. 2012; Bonari et al. 2017; Poschlod and Braun-Reichert 2017). Additionally, juniper-rich calcareous grasslands are even legally protected under the EU Habitats Directive (EC 2007).
In mesic grasslands, habitat heterogeneity should be increased (Bonari et al. 2017). In particular, management should aim to promote stands consisting of mosaics with varying sward heights, interspersed shrubs and some solitary trees (cf. Hartel and Plieninger 2014; Plieninger et al. 2015; Jakobsson et al. 2020; Tschumi et al. 2020). Both low-intensity rough grazing and year-round grazing systems with low stocking rates seem to be suitable tools to increase the heterogeneity in mesic grasslands and, thus, should generally be preferred against regular mowing (Olff et al. 1999; Fraser et al. 2014; Köhler et al. 2016; Kmecl and Denac 2018).
References
Baines D (1990) The roles of predation, food and agricultural practice in determining the breeding success of the lapwing (Vanellus vanellus) on upland grasslands. J Anim Ecol 59:915–929. https://doi.org/10.2307/5022
Bartón K (2020) Multi-Model Inference (Package MuMIn: version 1.43.17). <https://cran.r-project.org/package=MuMIn>. Accessed 18 Oct 2020
Bates D, Maechler M, Bolker B, Walker S (2020) Linear mixed-effects models using ‘Eigen’ and S4 (Package lme4: version 1.1-23). <http://cran.r-project.org/web/packages/lme4>. Accessed 24 Sept 2020
Baur B, Cremene C, Groza G, Rakosy L, Schileyko AA, Baur A, Stoll P, Erhardt A (2006) Effects of abandonment of subalpine hay meadows on plant and invertebrate diversity in Transylvania, Romania. Biol Conserv 132:261–273. https://doi.org/10.1016/j.biocon.2006.04.018
Bengtsson J, Bullock JM, Egoh B, Everson C, Everson T, O’Connor T, O’Farrell PJ, Smith HG, Lindborg R (2019) Grasslands—more important for ecosystem services than you might think. Ecosphere 10:e02582. https://doi.org/10.1002/ecs2.2582
Benton TG, Bryant DM, Cole L, Crick HQP (2002) Linking agricultural practice to insect and bird populations: a historical study over three decades. J Appl Ecol 39:673–687. https://doi.org/10.1046/j.1365-2664.2002.00745.x
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188. https://doi.org/10.1016/S0169-5347(03)00011-9
Berg A, Part T (1994) Abundance of breeding farmland birds on arable and set-aside fields at forest edges. Ecography 17:147–152. https://doi.org/10.1111/J.1600-0587.1994.TB00087.X
Bibby CJ, Burgess ND, Hill DA, Mustoe SH (2000) Bird census techniques, 2nd edn. Academic Press, London
Bonari G, Fajmon K, Malenovský I, Zelený D, Holuša J, Jongepierová I, Kočárek P, Konvička O, Uřičář J, Chytrý M (2017) Management of semi-natural grasslands benefiting both plant and insect diversity: the importance of heterogeneity and tradition. Agric Ecosyst Environ 246:243–252. https://doi.org/10.1016/j.agee.2017.06.010
Bourn NAD, Thomas JA (2002) The challenge of conserving grassland insects at the margins of their range in Europe. Biol Conserv 104:285–292. https://doi.org/10.1016/S0006-3207(01)00193-8
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
de Cáceres M, Jansen F (2016) Package indicspecies. https://cran.r-project.org. Accessed 20 Aug 2021
Cooper WJ, McShea WJ, Forrester T, Luther DA (2020) The value of local habitat heterogeneity and productivity when estimating avian species richness and species of concern. Ecosphere 11:e03107. https://doi.org/10.1002/ecs2.3107
Correll MD, Strasser EH, Green AW, Panjabi AO (2019) Quantifying specialist avifaunal decline in grassland birds of the Northern Great Plains. Ecosphere 10:e02523. https://doi.org/10.1002/ecs2.2523
Crawley MJ (2007) The R Book. Wiley, Chichester
Dekoninck W, De Koninck H, Baugnée JY, Maelfait JP (2007) Ant biodiversity conservation in Belgian calcareous grasslands: active management is vital. Belg J Zool 137:137–146
Diacon-Bolli J, Dalang T, Holderegger R, Bürgi M (2012) Heterogeneity fosters biodiversity: linking history and ecology of dry calcareous grasslands. Basic Appl Ecol 13:641–653. https://doi.org/10.1016/j.baae.2012.10.004
Donald PF, Sanderson FJ, Burfield IJ, Van Bommel FPJ (2006) Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agric Ecosyst Environ 116:189–196. https://doi.org/10.1016/j.agee.2006.02.007
Dufrêne M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr 67:345–366. https://doi.org/10.1890/0012-9615(1997)067[0345:SAAIST]2.0.CO;2
Ernst LM, Tscharntke T, Batary P (2017) Grassland management in agricultural vs. forested landscapes drives butterfly and bird diversity. Biol Conserv 216:51–59. https://doi.org/10.1016/j.biocon.2017.09.027
European Commission (EC) (2007) The Habitats Directive 92/43/EEC. Brussels, European Community
Fartmann T (2004) Die Schmetterlingsgemeinschaften der Halbtrockenrasen-Komplexe des Diemeltales: Biozönologie von Tagfaltern und Widderchen in einer alten Hudelandschaft. Abhandlungen Aus Dem Westfälischen Museum Für Naturkunde 66:1–269
Fartmann T, Krämer B, Stelzner F, Poniatowski D (2012) Orthoptera as ecological indicators for succession in steppe grassland. Ecol Indic 20:337–344. https://doi.org/10.1016/j.ecolind.2012.03.002
Fartmann T, Kämpfer S, Brüggeshemke J, Juchem M, Klauer F, Weking S, Löffler F (2018) Landscape-scale effects of Christmas-tree plantations in an intensively used low-mountain landscape—applying breeding bird assemblages as indicators. Ecol Indic 94:409–419. https://doi.org/10.1016/j.ecolind.2018.07.006
Fartmann T, Jedicke E, Streitberger M, Stuhldreher G (2021) Insektensterben in Mitteleuropa. Ursachen und Gegenmaßnahmen. Eugen Ulmer, Stuttgart
Farwell LS, Elsen PR, Razenkova E, Pidgeon AM, Radeloff VC (2020) Habitat heterogeneity captured by 30-m resolution satellite image texture predicts bird richness across the United States. Ecol Appl 30:e02157. https://doi.org/10.1002/eap.2157
Finck P, Heinze S, Raths U, Riecken U, Ssymank A (2017) Rote Liste der gefährdeten Biotoptypen Deutschlands: third edition. Naturschutz Und Biologische Vielfalt 156:1–642
Fischer S, Flade M, Schwarz J (2005) Revierkartierung. In: Südbeck P, Andretzke H, Fischer S, Gedeon K, Schikore T, Schröder K, Sudfeldt C (eds) Methodenstandards zur Erfassung der Brutvögel Deutschlands. Mugler, Radolfzell, pp 47–58
Flohre A, Fischer C, Aavik T, Bengtsson J, Berendse F, Bommarco R, Ceryngier P, Clement L, Dennis C, Eggers S, Emmerson MC, Geiger F, Guerrero I, Hawro V, Inchausti P, Liira J, Morales MB, Onate JJ, Part T, Weisser W, Winqvist C, Thies C, Tscharntke T (2011) Agricultural intensification and biodiversity partitioning in European landscapes comparing plants, carabids, and birds. Ecol Appl 21:1772–1781. https://doi.org/10.2307/23023116
Fraser MD, Moorby JM, Vale JE, Evans DM (2014) Mixed grazing systems benefit both upland biodiversity and livestock production. PLoS ONE 9:e89054. https://doi.org/10.1371/journal.pone.0089054
Fuhlendorf SD, Harrell WC, Engle DM, Hamilton RG, Davis CA, Leslie DM (2006) Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol Appl 16:1706–1716. https://doi.org/10.1890/1051-0761(2006)016[1706:shbtbf]2.0.co;2
Fuller RJ (ed) (2012) Birds and habitat: relationships in changing landscapes. Cambridge University Press, Cambridge
Fumy F, Fartmann T (2021) Climate and land-use change drive habitat loss in a mountain bird species. Ibis 163:1189–1206. https://doi.org/10.1111/ibi.12954
Gatter W (2000) Vogelzug und Vogelbestände in Mitteleuropa. AULA-Verlag, Wiebelsheim
German Meteorological Service (2021) Climate Data Center. URL: https://www.dwd.de/DE/leistungen/cdcftp/cdcftp.html. Accessed 5 July 2021
Geobasis NRW (2021) District government of Cologne. Digital elevation model. https://www.bezreg-koeln.nrw.de. Accessed 8 June 2021
Grueber CE, Nakagawa S, Laws RJ, Jamieson IG (2011) Multimodel inference in ecology and evolution: challenges and solutions. J Evol Biol 24:699–711. https://doi.org/10.1111/j.1420-9101.2010.02210.x
Grüneberg C, Sudmann CR, Weiss J, Jöbges M, König H, Laske V, Schmitz M, Skibbe A (2013) Die Brutvögel Nordrhein-Westfalens. LWL-Museum für Naturkunde, Münster
Grüneberg C, Sudmann SR, Herhaus F, Herkenrath P, Jöbges M, König H, Nottmeyer K, Schidelko K, Schmitz M, Schubert W, Stiels D, Weiss J (2017) Rote Liste der Brutvogelarten Nordrhein-Westfalens: 6. Fassung, Stand: Juni 2016. Charadrius 52:1–66
Hartel T, Plieninger T (eds) (2014) European wood-pastures in transition. A socio-ecological approach. Routledge, London
Helbing F, Fartmann T, Poniatowski D (2021) Restoration measures foster biodiversity of important primary consumers within calcareous grasslands. Biol Conserv 256:109058. https://doi.org/10.1016/j.biocon.2021.109058
Henle K, Alard D, Clitherow J, Corb P, Firbank L, Kull T, McCracken D, Moritz RFA, Niemela J, Rebane M, Wascher D, Watt A, Young J (2008) Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe—a review. Agric Ecosyst Environ 124:60–71. https://doi.org/10.1016/j.agee.2007.09.005
Jakobsson S, Wood H, Ekroos J, Lindborg R (2020) Contrasting multi-taxa functional diversity patterns along vegetation structure gradients of woody pastures. Biodivers Conserv 29:3551–3572. https://doi.org/10.1007/s10531-020-02037-y
Kämpfer S, Fartmann T (2019) Breeding populations of a declining farmland bird are dependent on a burrowing, herbivorous ecosystem engineer. Ecol Eng 140:105–592. https://doi.org/10.1016/j.ecoleng.2019.105592
Keller V, Herrando S, Voříšek P, Franch M, Kipson M, Milanesi P, Martí D, Anton M, Klvaňová A, Kalyakin MV, Bauer HG, Foppen RPB (2020) European Breeding Bird Atlas 2: Distribution, Abundance and Change. European Bird Census Council & Lynx Edicions, Barcelona
Kleijn D, Kohler F, Baldi A, Batary P, Concepcion E, Clough Y, Diaz M, Gabriel D, Holzschuh A, Knop E, Kovacs A, Marshall E, Tscharntke T, Verhulst J (2009) On the relationship between farmland biodiversity and land-use intensity in Europe. Proc R Soc B: Biol Sci 276:903–909. https://doi.org/10.1098/rspb.2008.1509
Kmecl P, Denac K (2018) The effects of forest succession and grazing intensity on bird diversity and the conservation value of a Northern Adriatic karstic landscape. Biodivers Conserv 27:2003–2020. https://doi.org/10.1007/s10531-018-1521-2
Köhler M, Hiller G, Tischew S (2016) Year-round horse grazing supports typical vascular plant species, orchids and rare bird communities in a dry calcareous grassland. Agric Ecosyst Environ 234:48–57. https://doi.org/10.1016/j.agee.2016.03.020
Krämer B, Poniatowski D, Fartmann T (2012) Effects of landscape and habitat quality on butterfly communities in pre-alpine calcareous grasslands. Biol Conserv 152:253–261. https://doi.org/10.1016/j.biocon.2012.03.038
Kujawa K, Tryjanowski P (2000) Relationships between the abundance of breeding birds in Western Poland and the structure of agricultural landscape. Acta Zool Acad Sci Hungaricae 46:103–114
Löffler F, Fartmann T (2017) Effects of landscape and habitat quality on Orthoptera assemblages of pre-alpine calcareous grasslands. Agric Ecosyst Environ 248:71–81. https://doi.org/10.1016/j.agee.2017.07.029
Löffler F, Poniatowski D, Fartmann T (2020) Extinction debt across three taxa in well-connected calcareous grasslands. Biol Conserv 246:108588
MacDonald D, Crabtree JR, Wiesinger G, Dax T, Stamou N, Fleury P, Gutíerrez Lazpita J, Gibon A (2000) Agricultural abandonment in mountain areas of Europe: environmental consequences and policy response. J Environ Manage 59:47–69. https://doi.org/10.1006/jema.1999.0335
Moorcroft D, Whittingham MJ, Bradbury RB, Wilson JD (2002) The selection of stubble fields by wintering granivorous birds reflects vegetation cover and food abundance. J Appl Ecol 39:535–547. https://doi.org/10.1046/j.1365-2664.2002.00730.x
Müller-Wille W (1981) Westfalen: Landschaftliche Ordnung und Bindung eines Landes, 2nd edn. Aschendorffsche Verlagsbuchhandlung, Münster
Newton I (2004) The recent declines of farmland bird populations in Britain: an appraisal of causal factors and conservation actions. Ibis 146:579–600. https://doi.org/10.1111/j.1474-919X.2004.00375.x
Newton I (2017) Farming and birds. William Collins, London
Olff H, Vera FWM, Bokdam J, Bakker ES, Gleichman JM, Maeyer K, Smit R (1999) Shifting mosaics in grazed woodlands driven by the alternation of plant facilitation and competition. Plant Biol 1:127–137. https://doi.org/10.1111/j.1438-8677.1999.tb00236.x
Pearce-Higgins JW, Green RE (2014) Birds and climate change. Impacts and conservation responses. Cambridge University Press, Cambridge
Plieninger T, Höchtl F, Spek T (2006) Traditional land-use and nature conservation in European rural landscapes. Environ Sci Policy 9:317–321. https://doi.org/10.1016/j.envsci.2006.03.001
Plieninger T, Hartel T, Martín-López B (2015) Wood-pastures of Europe: Geographic cover-age, social-ecological values, conservation management, and policy implications. Biol Conserv 190:70–79. https://doi.org/10.1016/j.biocon.2015.05.014
Poniatowski D, Fartmann T (2008) The classification of insect communities: lessons from orthopteran assemblages of semi-dry calcareous grasslands in central Germany. Eur J Entomol 105:659–671. https://doi.org/10.14411/eje.2008.090
Poniatowski D, Fartmann T (2010) What determines the distribution of a flightless bush-cricket (Metrioptera brachyptera) in a fragmented landscape? J Insect Conserv 14:637–645. https://doi.org/10.1007/s10841-010-9293-3
Poniatowski D, Hertenstein F, Raude N, Gottbehüt K, Nickel H, Fartmann T (2018) The invasion of Bromus erectus alters species diversity of vascular plants and leafhoppers in calcareous grasslands. Insect Conserv Divers 11:578–586. https://doi.org/10.1111/icad.12302
Poniatowski D, Stuhldreher G, Helbing F, Hamer U, Fartmann T (2020) Restoration of calcareous grasslands: the early successional stage promotes biodiversity. Ecol Eng 151:105858. https://doi.org/10.1016/j.ecoleng.2020.105858
Poschlod P (2017) Geschichte der Kulturlandschaft, 2nd edn. Eugen Ulmer, Stuttgart
Poschlod P, Braun-Reichert R (2017) Small natural features with large ecological roles in ancient agricultural landscapes of Central Europa—history, value, status, and conservation. Biol Conserv 211:60–68. https://doi.org/10.1016/j.biocon.2016.12.016
Poschlod P, Wallis De Vries MF (2002) The historical and socioeconomic perspective of calcareous grasslands—lessons from the distant and recent past. Biol Conserv 104:361–376. https://doi.org/10.1016/S0006-3207(01)00201-4
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Reif J, Hanzelka J (2020) Continent-wide gradients in open-habitat insectivorous bird declines track spatial patterns in agricultural intensity across Europe. Glob Ecol Biogeogr 29:1988–2013. https://doi.org/10.1111/geb.13170
Reif J, Voříšek P, Šťastný K, Bejcek V, Petr J (2008) Agricultural intensification and farm-land birds: new insights from a central European country. Ibis 150:596–605. https://doi.org/10.1111/j.1474-919X.2008.00829.x
Schmidt A, Fartmann T, Kiehl K, Kirmer A, Tischew S (2022) Effects of perennial wildflower strips and landscape structure on birds in intensively-farmed agricultural landscapes. Basic Appl Ecol 58:15–25. https://doi.org/10.1016/j.baae.2021.10.005
Schwarz C, Trautner J, Fartmann T (2018) Common pastures are important refuges for a declining passerine bird in a pre-alpine agricultural landscape. J Ornithol 159:945–954. https://doi.org/10.1007/s10336-018-1561-0
Tagmann-Ioset A, Schaub M, Reichlin TS, Weisshaupt N, Arlettaz R (2012) Bare ground as a crucial habitat feature for a rare terrestrially foraging farmland bird of Central Europe. Acta Oecol 39:25–32. https://doi.org/10.1016/j.actao.2011.11.003
Thorn S, Werner SAB, Wohlfahrt J, Bässler C, Seibold S, Quillfeld P, Müller J (2016) Response of bird assemblages to windstorm and salvage logging: Insights from analyses of functional guild and indicator species. Ecol Indic 65:142–148. https://doi.org/10.1016/j.ecolind.2015.06.033
Tschumi M, Birkhofer K, Blasiusson S, Jörgensen M, Smith HG, Ekroos J (2020) Woody elements benefit bird diversity to a larger extent than semi-natural grasslands in cereal-dominated landscapes. Basic Appl Ecol 46:15–23. https://doi.org/10.1016/j.baae.2020.03.005
Van Swaay CAM (2002) The importance of calcareous grasslands for butterflies in Europe. Biol Conserv 104:315–318. https://doi.org/10.1016/S0006-3207(01)00196-3
Vickery J, Arlettaz R (2012) The importance of habitat heterogeneity at multiple scales for birds in European agricultural landscapes. In: Fuller RJ (ed) Birds and habitat. Cambridge University Press, Cambridge, pp 177–204
Wallis De Vries MF, Poschlod P, Willems JH (2002) Challenges for the conservation of calcareous grasslands in northwestern Europe: integrating the requirements of flora and fauna. Biol Conserv 104:265–273. https://doi.org/10.1016/S0006-3207(01)00191-4
Wolf B, Widdig T (eds) (2016) Rote Liste der bestandsgefährdeten Brutvogelarten Hessens, 10. Fassung, Stand Mai 2014. Hessisches Ministerium für Umwelt, Klimaschutz, Landwirtschaft und Verbraucherschutz, Wiesbaden, Germany
Acknowledgements
We are grateful to Matthias Tschumi and three anonymous reviewers for valuable comments on an earlier version of the manuscript. The study was funded by the Stöckmann Foundation for Environment and Nature Conservation (Grant: S0393/10017/2019).
Funding
Open Access funding enabled and organized by Projekt DEAL. This work was funded by Stöckmann Foundation for Environment and Nature Conservation (Grant no: S0393/10017/2019).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Communicated by T. Gottschalk.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brüggeshemke, J., Drung, M., Löffler, F. et al. Effects of local climate and habitat heterogeneity on breeding-bird assemblages of semi-natural grasslands. J Ornithol 163, 695–707 (2022). https://doi.org/10.1007/s10336-022-01972-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-022-01972-7