
Abstract
During migration, birds need to optimize their time and/or energy management, especially during stop-overs. Previous studies with caged birds under controlled condition support the notion that departure decisions are condition-dependent, but they did not take into account the availability or the actual intake of food. In the study reported here we investigated whether food intake interacts with condition in influencing migratory disposition in temporarily caged wild migrants. We conducted the study on Garden Warblers (Sylvia borin) and European Robins (Erithacus rubecula) at a spring stop-over site that is reached after a long non-stop flight over the Mediterranean Sea. The birds were held in cages during the day and the first night following capture, and their locomotor activity, food intake, and body mass change were recorded. In both species, food intake was positively correlated with body mass change. Body condition was positively correlated with migratory restlessness (Zugunruhe) in both species and was also negatively related to diurnal activity in Garden Warblers. Food intake was negatively correlated with diurnal activity in Garden Warblers. When only birds with low fat scores were considered, we found a positive effect of food intake on Zugunruhe in Garden Warblers. In general, European Robins consumed less food than Garden Warblers and no significant effects on Zugunruhe were found. Our results show that food intake interacts with condition in affecting migratory behavior during stop-overs. In particular, food intake may induce Garden Warblers with low fat stores at arrival to leave a stop-over site earlier. The large individual variability in food consumption supports the notion that birds vary in their capacity to refuel soon after arrival at a stop-over site. Overall, our work suggests that the decision to stay at the stop-over site or to resume migration depends on the interaction of condition, refueling capacity and rate, and migratory strategy.
Zusammenfassung
Einfluss von Körperkondition und Nahrungsaufnahme auf Abflugentscheidungen während der Rast beim Frühjahrszug von Gartengrasmücken und Rotkehlchen
Zugvögel müssen insbesondere auf Rastplätzen ihre Zeit und/oder Energie bestmöglich nutzen. Experimentelle Studien mit Vögeln unter kontrollierten Bedingungen in Gefangenschaft haben gezeigt, dass die Entscheidung, wann von einem Rastplatz wieder weitergeflogen wird, konditionsabhängig ist. Nahrungsangebot und -aufnahme wurden bei diesen Versuchen allerdings nicht berücksichtigt. Für diese Studie haben wir daher die Interaktion zwischen Nahrungsaufnahme und Körperkondition bei Zugvögeln in temporärer Gefangenschaft untersucht. Wir erforschten Gartengrasmücken (Sylvia borin) und Rotkehlchen (Erithacus rubecula) an einem Frühjahrs-Rastplatz, den sie nach einem langen Nonstop-Flug über das Mittelmeer nutzen. Nach dem Fang wurden die Vögel für einen ganzen Tag und eine ganze Nacht in Käfigen gehalten, um Aktivität, Nahrungsaufnahmerate und Veränderungen des Körpergewichts zu messen. In beiden Arten war die Nahrungsaufnahme positiv mit der Zunahme des Körpergewichts korreliert. Die Körperkondition beider Arten korrelierte positiv mit Zugunruhe, und in der Gartengrasmücke negativ mit Tagesaktivität. In der Gartengrasmücke war außerdem die Nahrungsaufnahme negativ mit der Tagesaktivität korreliert. Wenn wir nur Vögel mit geringen Fettreserven betrachteten, fanden wir bei der Gartengrasmücke einen positiven Effekt von Nahrungsaufnahme auf die Zugunruhe. Im Allgemeinen nahmen Rotkehlchen weniger Nahrung als Gartengrasmücken auf und die Nahrungsaufnahme hatte keinen Einfluss auf deren Zugunruhe. Diese Ergebnisse zeigen, dass Nahrungsaufnahme im Zusammenwirken mit Körperkondition Abflugentscheidungen vom Rastplatz beeinflussen kann. Insbesondere Gartengrasmücken mit niedrigen Fettereserven bei der Ankunft konnten den Rastplatz schneller verlassen, sofern sie Nahrung aufnahmen. Die Ergebnisse zeigten große interindividuelle Variabilität in der Nahrungsaufnahme der beiden untersuchten Arten. Dies weist darauf hin, dass Vögel sich in ihrer Fähigkeit, Nahrung gleich nach der Landung auf einem Rastplatz aufzunehmen, unterscheiden. Unsere Studie zeigt zum ersten Mal, dass die Entscheidung, an einem Rastplatz zu verweilen oder weiterzufliegen, vom Wechselspiel zwischen Körperkondition, Nahrungsaufnahmerate und -kapazität sowie der Zugstrategie abhängig ist.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Every year, millions of small passerine birds migrate between Northern Europe and sub-Saharan Africa (Hahn et al. 2009), covering large distances and facing substantial ecological barriers, such as seas and deserts. Most birds do not complete migration in a single flight but instead frequently stop-over to accumulate energy resources, recover from fatigue, or wait for favorable weather conditions (Carmi et al. 1992; Klaassen 1996; Schmaljohann et al. 2007). In the spring and autumn migratory birds are under time pressure to reach their breeding grounds (Kokko 1999) and their wintering grounds (Delingat et al. 2006), respectively, and hence they are expected to optimize the pace of migration. In terms of time, most of the migration period is spent at stop-over sites (normally >80%; Hedenström and Alerstam 1997), and thus the number and duration of stop-overs determine the overall pace of migration. Optimal migration theory predicts that stop-over time should be reduced when birds experience high fuel deposition rates (Alerstam and Hedenström 1998) and that they should then leave a stop-over site as they reach the appropriate optimal fuel load (Alerstam and Lindström 1990). Therefore, birds arriving at a stop-over site in poor condition should require longer stop-overs or should be under stronger pressure to increase their fuel deposition rate to keep their stop-over as short as possible.
The dependency of stop-over duration on individual condition has not been shown consistently in field studies that assessed minimum stop-over duration in passerines. Condition dependence was shown in several studies conducted in the autumn (Biebach 1985; Biebach et al. 1986; Arizaga et al. 2008; Smolinsky et al. 2013), but there are some exceptions (Tsvey et al. 2007; Schmaljohann et al. 2013). One study found that stop-over duration depended mostly on condition in long-distance migrants, but not in short-distance migrants (Bairlein 1985). Another study found condition dependence only at one of three stop-over sites in three species studied during the autumn migration (Schaub et al. 2008). During the spring migration, the patterns are more variable depending on species and location, with many studies showing condition-dependent stop-over durations (Yong and Moore 1993; Gannes 2002; Fusani et al. 2009; Goymann et al. 2010; Lupi et al. 2016) and others not (Salewski and Schaub 2007; Maggini and Bairlein 2011). The large number of studies with conflicting results implies that the condition dependency of stop-over duration may vary among species, migratory season, and location. Most of these studies, however, were observational and based on recaptures of ringed birds. The few studies in which migratory behavior was recorded under controlled laboratory conditions consistently found that lean birds showed less migratory restlessness than their fatter counterparts (Fusani et al. 2009; Eikenaar and Schläfke 2013; Lupi et al. 2016). These studies took advantage of a well-established relationship between the disposition to migrate and the intensity of nocturnal or migratory restlessness, also called Zugunruhe, in caged birds (Berthold et al. 1972; Eikenaar et al. 2014). However, the latter studies either did not provide food and thus refueling opportunities to the birds (Lupi et al. 2016) or they did not measure food intake (Fusani et al. 2009; Eikenaar and Schläfke 2013). It has recently been shown that fuel loss during fasting is a better predictor for migratory restlessness (hereafter Zugunruhe) than initial condition in birds that are temporarily caged during migration (Eikenaar and Bairlein 2014). In the only study in which fuel deposition rates were manipulated in birds that were successively released, food intake (and thus fuel deposition rates) was the major determinant of departure from a stop-over site (Smith and McWilliams 2014). These data indicate that stop-over decisions may depend on whether birds are given the opportunity to refuel during the day or not.
In the study reported here, we investigated the influence of food intake and individual body condition on departure decisions from a spring stop-over site, the island of Ponza. We tested these factors in two species that differ strongly in migration patterns, namely, Garden Warblers (Sylvia borin) and European Robins (Erithacus rubecula, hereafter Robins). Garden Warblers are long-distance migrants, and the populations that come through Ponza need to fly over the Sahara Desert and the Mediterranean Sea before reaching their breeding grounds in Northern Europe. In contrast, Robins are short-distance migrants, and the populations that come through Ponza migrate from Northern Africa to Italy or Central Europe (Spina and Volponi 2008). Both species show similar patterns of condition-dependent stop-over behavior in Ponza, i.e., lean birds stop-over longer or show less migratory restlessness than fat birds (Fusani et al. 2009; Goymann et al. 2010; Lupi et al. 2016). We captured Garden Warblers and Robins and held them temporarily in captivity. However, in contrast to previous studies we offered them food to obtain a measure of their fuel deposition rate and body mass change. At the same time, we measured diurnal locomotor activity and Zugunruhe to assess migratory disposition. If the amount of food consumed affected departure timing, we would expect birds to resume migration soon (as indicated by elevated amounts of Zugunruhe) after refueling, and their propensity to migrate should increase with increasing refueling. The alternative hypothesis is that condition alone determines departure. In that case, food intake should not influence the amount of Zugunruhe shown by the birds, which would be higher in fat birds than in lean birds.
Methods
Experimental design
The study was carried out between 19 March and 19 May 2012 at the Ponza Research Station (http://www.inanellamentoponza.it), located on the island of Ponza (40°50′ N, 12°58′ E; surface area 9.87 km2), about 50 km from the Tyrrhenian coast of Italy. Garden Warblers and Robins were trapped with mist nets, with continuous monitoring between sunrise and sunset. Upon capture they were ringed and their body condition assessed based on body mass, fat stores, and pectoral muscle size (see below). Fat was scored on a scale from 0 (no visible fat) to 8 (fat completely covering the pectoral muscle), and pectoral muscle was scored on a scale from 0 (muscle concave) to 3 (muscle bulging, sternum no longer visible) following Bairlein (1995).
We used 63 Garden Warblers and 88 Robins for this experiment. Only birds that were captured in the morning hours (before 1300 hours) were used so that the birds had sufficient time to accustom to housing conditions. After ringing, all individuals were weighed and immediately transferred to individual custom-built fabric cages (50 × 30 × 30 cm) inside a room close to the capture site. These cages have been used in a number of previous studies (Bauchinger et al. 2008; Fusani et al. 2009, 2011, 2013; Lupi et al. 2016) and have been optimized for short-term housing of migratory passerines. In each cage an infrared sensor attached to the wall detected bird movements, and a custom-made software summarized the movement counts in 2-min intervals. Depending on capture success, up to 24 birds per day were tested simultaneously.
The birds were provided with water ad libitum and 8 ± 1 g of a mixture of dry insect food (Aleckwa Tiernahrung Thomas Mayer, Altrip, Germany), boiled egg, and banana (1 egg and a one-half banana per 200 g dry insect food) and were exposed to a natural photoperiod. The food was weighed before the birds were placed in the cages, and the remaining food (including what had been scattered around the cage by the birds) was weighed after the birds were released the next morning. To account for evaporative loss of food weight, a reference bowl of food was placed into an empty cage and weighed at the same times. Food intake was then calculated by subtracting the weight of the food bowl at the end of the experiment from the weight of the bowl at the beginning of the experiment, followed by correction for evaporation using the difference in weight of the control food bowl.
Zugunruhe during the first part of the night was calculated as the mean number of movements per 2-min interval between civil twilight (Central European Time) and midnight (0000 hours). We decided to interrupt the measurements at this time because some of the birds were used for another experiment after midnight. However, departure from a stop-over site is likely to happen during the first part of the night in the majority of nocturnal migrants (Gwinner 1967; Goymann et al. 2010). Diurnal activity was calculated as the mean number of movements per 2-min interval between 1300 hours and civil twilight. All birds were weighed again prior to their release the following morning at 0600 hours. Body mass change was then calculated by subtracting body mass at the time of caging from body mass at release.
A total of 87 birds (38 Garden Warblers, 49 Robins) were equipped with a logger (1.5 g) for a separate experiment not reported here. We accounted for possible effects of the logger in the statistical analysis. In addition, activity data recorded during the period in which a researcher was present in the room until 1 h after the end of the disturbance were excluded from the analysis in order to remove possible effects of this disturbance from the results. We calculated the time elapsed from the introduction of the birds into the cage and the beginning of the data recording (1300 hours) and introduced this variable in the analysis to account for differences in acclimation to captivity.
Data analysis
We extracted a body condition index called CONDITION by conducting a principal component analysis on body mass, fat score, and muscle score at capture for each species (as described in Fusani et al. 2009, 2011). In Garden Warblers, CONDITION (eigenvalue 2.34) explained 78% of the total variance, and the individual variable loadings were: body mass 0.90; fat 0.89; muscle 0.86. In Robins, CONDITION (eigenvalue 1.76) explained 59% of the total variance, and the individual variable loadings were: body mass 0.88; fat 0.88; muscle 0.47. We performed linear regression models to test for the effect of CONDITION, food intake, logger, elapsed time from caging to the beginning of the experiment, and the interaction between CONDITION and food intake on Zugunruhe and diurnal activity, separately for Garden Warblers and Robins. We calculated variance inflation factors (VIFs) for all models to control for collinearity of variables. VIFs were never higher than 2. These models were compared to generalized additive models (GAMs) performed using the same explanatory variables to identify possible non-linear relationships. The results of the GAMs suggested that linear relationships were a good approximation of the data. Adjusted R 2 values were comparable to the linear regression models, and Akaike information criteria of the GAMs were always higher than those of the linear regression models. For this reason, we presented only the results of the linear regression models in this report. We calculated Pearson’s correlation coefficients to assess the relationship between food intake and body mass change. Student’s t test was used to check for differences in food consumption between Garden Warblers and Robins, and a Mann–Whitney non-parametric test was used to compare fat scores between species. Since birds that had large fat stores ate little or nothing (see section "Results"), linear regression models were run on two different datasets for each species: (1) the full dataset, including all individuals, (2) a low fat scores dataset, including only the individuals with fat scores of 0–3. As no Robins with fat score 0 were used in the experiments, the low fat scores dataset for Robins included only birds with fat scores of 1–3. The models were checked for normality of residuals and heteroscedasticity by plotting residuals (Crawley 2007). All statistical analyses were performed with R version 3.0.2 (R Core Team 2014). Due to missing values for food intake, 62 of 63 Garden Warblers and 85 of 88 Robins were included in the linear models with the full dataset, and 51 of 52 Garden Warblers and 75 of 80 Robins were included in the linear models with the low fat scores dataset.
Results
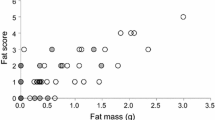
Food intake and body mass change were positively correlated in both Garden Warblers (Fig. 1a) and Robins (Fig. 1b), with birds having higher food intake showing a higher increase in body mass. Birds in a relatively better condition showed lower food intake in both Garden Warblers (Pearson’s correlation r = −0.398, p = 0.002) and Robins (r = −0.299, p = 0.006). Overall, Garden Warblers ate significantly more [mean ± 95% confidence interval (CI) 1.686 ± 0.233 g] than Robins (mean ± 95% CI 0.634 ± 0.107 g; t test: t = 4.102, p < 0.001; Fig. 2). The fat score of Garden Warblers was on average lower than that of Robins (Mann–Whitney test: W = 2629, p < 0.001).

Relationships between food intake and body mass change between placement into the cage and release the next morning in Garden Warblers (Sylvia borin) (a) and European Robins (Erithacus rubecula) (b). There was a significant positive correlation between food intake and body mass change in both Garden Warblers (r = 0.724, p < 0.001) and Robins (r = 0.805, p < 0.001). Star beside the panel letter indicates a significant correlation, lines indicate the linear regression between food intake and body mass change

Box plot of food intake by fat score in Garden Warblers (gray) and Robins (white). Numbers below the boxes indicate the sample sizes
In Garden Warblers, birds with higher CONDITION values at capture showed more Zugunruhe and less diurnal activity in both the full and low fat scores datasets (Tables 1, 2; Fig. 3). Food intake affected Zugunruhe and diurnal activity in Garden Warblers, such that higher food intake was associated with higher intensity of Zugunruhe when the low fat scores dataset was considered and with lower diurnal activity in the full dataset (Tables 1, 2). Garden Warblers with loggers showed a lower intensity of Zugunruhe than birds without loggers in the full dataset (Table 1).

Relationships between the body condition index (CONDITION) and diurnal activity or migratory restlessness (Zugunruhe) in Garden Warblers (a, c) and Robins (b, d). There was a significant negative correlation between CONDITION and diurnal activity (r p = −0.490, p < 0.001; a) and a positive correlation between CONDITION and Zugunruhe (r p = 0.328, p = 0.009; c) in Garden Warblers. In Robins, CONDITION was significantly positively correlated with Zugunruhe (r p = 0.213, p = 0.046; d), but not with diurnal activity (r p = −0.189, p = 0.077; b). Star beside the panel letter indicates a significant correlation, lines indicate the linear regression between CONDITION and diurnal activity or Zugunruhe
In Robins, birds with higher CONDITION values at capture showed more Zugunruhe when the full dataset was considered and less diurnal activity when the low fat scores dataset was considered (Tables 1, 2; Fig. 3). Food intake did not affect Zugunruhe or diurnal activity (Tables 1, 2). Elapsed time from caging affected both Zugunruhe and diurnal activity: birds that spent a longer time in the cage before the beginning of the experiments showed more Zugunruhe and less diurnal activity in both the full and low fat scores datasets (Tables 1, 2).
Discussion
The results of this study support previous findings that birds with sufficient energy stores are more likely to leave a stop-over site than birds with insufficient energy stores (Fusani et al. 2009; Goymann et al. 2010; Eikenaar and Schläfke 2013; Lupi et al. 2016; Goymann et al. 2017). These studies also reported that higher Zugunruhe in fatter birds was coupled to lower activity during the day. The large individual variability in food intake in the low fat score classes prompted us to investigate how fuel deposition rate affected the departure decisions of these birds (as predicted by Zugunruhe; Eikenaar et al. 2014). Our results show that higher food intake resulted in more Zugunruhe in Garden Warblers but not in Robins.
Body mass was found to change according to food intake in both Garden Warblers and Robins. In both species, birds that did not consume food lost body mass, while birds that ate substantial amounts of food were able to increase their body mass. Body mass change was obviously also influenced by rehydration, although it was not possible to quantify this variable, which may explain the positive body mass change in birds that ate little (Fig. 1a). Hume and Biebach (1996) subjected birds to a prolonged fast and found that there was a reduction of the digestive tract, which limited the immediate capacity of the birds to eat in the first day of refeeding and prevented an increase in body mass. Unfortunately, in our study it was not possible to assess whether the day of capture coincided with the day of arrival at the stop-over site or whether the birds had more time to rebuild their digestive tract. Long-distance migrants commonly tend to migrate faster in the spring than in the autumn due to the advantages of early arrival at the breeding sites (Dierschke et al. 2005; Maggini and Bairlein 2010; Karlsson et al. 2012). Garden Warblers, as strict long-distance migrants, fit this scenario well. The strong endogenous regulation of migration in this species (Gwinner 1996) causes a very consistent response to intrinsic and extrinsic factors, such as body condition and food availability, for example, which is not masked by experimental conditions such as temporary captivity. In our study, Garden Warblers at time of capture had a lower average fat score than Robins at time of capture. We suggest that Garden Warblers may have skipped refueling at the Northern African coast to reduce overall migration time (as suggested by Pilastro and Spina 1997). On the contrary, food intake was rather low for all fat classes in Robins. This species could be particularly sensitive to the reduction of the digestive tract that is often observed in migrants after long-distance flights, such as when they are crossing the Mediterranean Sea (Hume and Biebach 1996; Bauchinger et al. 2005; McWilliams and Karasov 2005). Alternatively, a low fuel deposition rate could reflect capture stress (Carr 2002). In Robins that did refuel, food intake had no significant effect on Zugunruhe. This observation could be linked to the fact that Robins have a shorter expected migration distance left to cover. A study by Dierschke and Delingat (2001) showed that Northern Wheatears (Oenanthe oenanthe) left a stop-over site close to their breeding area quickly and independently of refueling rate and weather conditions. In contrast, birds that faced a large ecological barrier finely adjusted their departure depending on body condition. Similarly, Robins in our study might have shown the observed patterns in food intake and Zugunruhe because they were close to their breeding grounds. Robins caught on Ponza are likely to breed in nearby locations on mainland Italy (Spina and Volponi 2008). It is also possible that birds switch to an energy-saving strategy when approaching the breeding grounds in view of the breeding season. For example, arriving at the breeding grounds with sufficient energy stores has been shown to increase fitness in some migrants (e.g., Sandberg and Moore 1996). Robins in our study responded differently to the amount of time spent in the cages. Birds that entered the cage earlier in the day were less active during the diurnal measurement period (which started at 1300 hours, while caging usually occurred at any time between 0800 and 1300 hours) but more active during the night. This observation, along with the overall low refueling rates of Robins, indicates that these birds might concentrate their active period in the first few hours of captivity and use the remaining daylight hours for resting prior to starting Zugunruhe the next night. Accordingly, Robins that were caged later in the day just had less time to refuel, and the active period was prolonged into the diurnal measurement period.
The combined effects of food availability and/or intake and condition on migratory behavior have been described in several studies (e.g., Biebach 1985; Fusani and Gwinner 2004; Fusani et al. 2011). When Blackcaps and Garden Warblers with substantial fat stores are subjected to a prolonged fast (1–2 days), they typically respond to subsequent food availability with a dramatic reduction or even suppression of migratory restlessness (Biebach 1985; Gwinner et al. 1992; Fusani and Gwinner 2004; Fusani et al. 2013). The extent of the response depends on condition, particularly on residual mass after fast (Biebach 1985) and on seasonal factors, i.e., the response is stronger during the autumn migration than during the spring migration (Fusani and Gwinner 2004). In previous work, we showed that food availability affects Zugunruhe (Fusani et al. 2011), but we did not measure food intake. The present study shows that the actual food intake rather than the simple presence of food increases Zugunruhe, at least in Garden Warblers. Our findings might appear to be in contrast with the prediction that food availability after a prolonged fasting would inhibit Zugunruhe, as found in the above-mentioned studies (Biebach 1985; Gwinner et al. 1992; Fusani and Gwinner 2004; Fusani et al. 2013). This discrepancy can be explained in view of the different situation at our study site in Ponza. In this study, we took a snapshot of migratory disposition of individuals whose refueling history is unknown. In addition, the motivation for longer stop-overs on the island may be reduced on Ponza since the Italian mainland is usually visible to migrants landing in Ponza during the daytime.
Birds of comparable, low condition that had access to the same amount of food showed different levels of Zugunruhe depending on whether they ate or not (with Zugunruhe increasing with increasing food intake). This observation shows that there is a large individual variability in food consumption that is not explained by body condition as determined by body mass, fat stores, and muscle size. This variability may reflect a different status of the digestive tract, which can be greatly reduced during long migratory flights to support the energetic needs and/or the intermediates of lipid metabolism (McWilliams et al. 2004). Alternatively, personality traits, such as neophobia (when presented a new environment, e.g., Greenberg 1984; Mettke-Hofmann et al. 2009) and differential responsiveness to stressors, or endogenous stressors, such as parasite burden (van Gils et al. 2007; Klaassen et al. 2012), could be partly responsible for the (large) unexplained variance in the response of Zugunruhe to condition and food intake. In a recent study, we found that the appetite-regulating hormone, ghrelin, reduces food intake and increase migratory restlessness in Garden Warblers (Goymann et al. 2017). The concentration of hormone in the blood reflects well the condition of the animals and correlates with the extent of the subcutaneous fat stores (Goymann et al. 2017). These findings allow us to draw a physiological interpretation of the results of the present study: the motivation to eat in birds with a low condition might depend on both ghrelin levels, which are condition dependent, and on time spent at the stop-over site and thus the birds’ foraging history. Birds in similar conditions might be either migrants that have just arrived to the stop-over site or birds that have been on the island for a few days and have already eaten, although at a rate that did not lead to body mass increase. Further studies on the regulation of food intake by ghrelin should help us to test this hypothesis.
This study revealed an interplay of body condition and food intake on the decision whether to depart from a stop-over site or not. The relative importance of one or the other may vary among and within species, probably in relation to migration strategy and the minimization of either time or energy. Our results reveal the important role of food availability and food intake on stop-over decisions of birds, which deserve further attention in future studies.
References
Alerstam T, Hedenström A (1998) The development of bird migration theory. J Avian Biol 29:343–369. doi:10.2307/3677155
Alerstam T, Lindström A (1990) Optimal bird migration: the relative importance of time, energy, and safety. In: Gwinner E (ed) Bird migration: physiology and ecophysiology. Springer, Berlin, pp 331–351
Arizaga J, Barba E, Belda EJ (2008) Fuel management and stopover duration of Blackcaps Sylvia atricapilla stopping over in northern Spain during autumn migration period. Bird Study 55:124–134. doi:10.1080/00063650809461513
Bairlein F (1985) Body weights and fat deposition of Palaearctic passerine migrants in the central Sahara. Oecologia 66:141–146. doi:10.1007/Bf00378566
Bairlein F (1995) Manual of field methods. European-African Songbird Migration Network, Wilhelmshaven
Bauchinger U, Wohlmann A, Biebach H (2005) Flexible remodeling of organ size during spring migration of the garden warbler (Sylvia borin). Zoology 108:97–106. doi:10.1016/j.zool.2005.03.003
Bauchinger U, Van’t Hof T, Biebach H (2008) Migratory stopover conditions affect the developmental state of male gonads in Garden Warblers (Sylvia borin). Horm Behav 54:312–318. doi:10.1016/j.yhbeh.2008.03.007
Berthold P, Gwinner E, Klein H (1972) Circannuale Periodik bei Grasmücken. I. Periodik des Körpergewichtes, der Mauser und der Nachtunruhe bei Sylvia atricapilla und S. borin unter verschiedenen konstanten Bedingungen. J Ornithol 113:170–190. doi:10.1007/BF01640500
Biebach H (1985) Sahara stopover in migratory flycatchers: fat and food affect the time program. Experientia 41:695–697. doi:10.1007/Bf02007727
Biebach H, Friedrich W, Heine G (1986) Interaction of bodymass, fat, foraging and stopover period in trans-Sahara migrating passerine birds. Oecologia 69:370–379. doi:10.1007/Bf00377059
Carmi N, Pinshow B, Porter WP, Jaeger J (1992) Water and energy limitations on flight duration in small migrating birds. Auk 109:268–276. doi:10.2307/4088195
Carr JA (2002) Stress, neuropeptides, and feeding behavior: a comparative perspective. Integr Comp Biol 42:582–590. doi:10.1093/icb/42.3.582
Crawley MJ (2007) The R book. Wiley & Sons, Chichester UK, Hoboken NJ
Delingat J, Dierschke V, Schmaljohann H, Mendel B, Bairlein F (2006) Daily stopovers as optimal migration strategy in a long-distance migrating passerine: the Northern Wheatear Oenanthe oenanthe. Ardea 94:593–605. http://ardea.nou.nu/ardea_show_abstract.php?lang=uk&nr=621
Dierschke V, Delingat J (2001) Stopover behaviour and departure decision of Northern Wheatears, Oenanthe oenanthe, facing different onward non-stop flight distances. Behav Ecol Sociobiol 50:535–545. doi:10.1007/s002650100397
Dierschke V, Mendel B, Schmaljohann H (2005) Differential timing of spring migration in Northern Wheatears Oenanthe oenanthe: hurried males or weak females? Behav Ecol Sociobiol 57:470–480. doi:10.1007/s00265-004-0872-8
Eikenaar C, Schläfke JL (2013) Size and accumulation of fuel reserves at stopover predict nocturnal restlessness in a migratory bird. Biol Lett 9:20130712. doi:10.1098/rsbl.2013.0712
Eikenaar C, Bairlein F (2014) Food availability and fuel loss predict Zugunruhe. J Ornithol 155:65–70. doi:10.1007/s10336-013-0987-7
Eikenaar C, Klinner T, Szostek KL, Bairlein F (2014) Migratory restlessness in captive individuals predicts actual departure in the wild. Biol Lett 10:20140154. doi:10.1098/rsbl.2014.0154
Fusani L, Cardinale M, Carere C, Goymann W (2009) Stopover decision during migration: physiological conditions predict nocturnal restlessness in wild passerines. Biol Lett 5:302–305. doi:10.1098/rsbl.2008.0755
Fusani L, Cardinale M, Schwabl I, Goymann W (2011) Food availability but not melatonin affects nocturnal restlessness in a wild migrating passerine. Horm Behav 59:187–192. doi:10.1016/j.yhbeh.2010.11.013
Fusani L, Coccon F, Rojas Mora A, Goymann W (2013) Melatonin reduces migratory restlessness in Sylvia warblers during autumnal migration. Front Zool 10:79. doi:10.1186/1742-9994-10-79
Fusani L, Gwinner E (2004) Simulation of migratory flight and stopover affects night levels of melatonin in a nocturnal migrant. Proc Biol Sci 271:205–211. doi:10.1098/rspb.2003.2561
Gannes LZ (2002) Mass change pattern of Blackcaps refueling during spring migration: evidence for physiological limitations to food assimilation. Condor 104:231–239. doi:10.1650/0010-5422(2002)104[0231:Mcpobr]2.0.Co;2
Goymann W, Spina F, Ferri A, Fusani L (2010) Body fat influences departure from stopover sites in migratory birds: evidence from whole-island telemetry. Biol Lett 6:478–481. doi:10.1098/rsbl.2009.1028
Goymann W, Lupi S, Kaiya H, Cardinale M, Fusani L (2017) Ghrelin affects stopover decisions and food intake in a long-distance migrant. Proc Natl Acad Sci USA. doi:10.1073/pnas.1619565114
Greenberg R (1984) Neophobia in the foraging-site selection of a neotropical migrant bird: an experimental study. Proc Natl Acad Sci USA 81:3778–3780. doi:10.1073/pnas.81.12.3778
Gwinner E (1967) Circannuale Periodik der Mauser und der Zugunruhe bei einem Vogel. Naturwissenschaften 54:447. doi:10.1007/BF00603157
Gwinner E (1996) Circannual clocks in avian reproduction and migration. Ibis 138:47–63. doi:10.1111/j.1474-919X.1996.tb04312.x
Gwinner E, Schwabl H, Schwabl-Benzinger I (1992) The migratory time program of the garden warbler: is there compensation for interruptions? Ornis Scand 23:264–270
Hahn S, Bauer S, Liechti F (2009) The natural link between Europe and Africa—2.1 billion birds on migration. Oikos 118:624–626. doi:10.1111/j.1600-0706.2008.17309.x
Hedenström A, Alerstam T (1997) Optimum fuel loads in migratory birds: distinguishing between time and energy minimization. J Theor Biol 189:227–234. doi:10.1006/jtbi.1997.0505
Hume ID, Biebach H (1996) Digestive tract function in the long-distance migratory garden warbler, Sylvia borin. J Comp Physiol B 166:388–395. doi:10.1007/s003600050024
Karlsson H, Nilsson C, Bäckman J, Alerstam T (2012) Nocturnal passerine migrants fly faster in spring than in autumn: a test of the time minimization hypothesis. Anim Behav 83:87–93. doi:10.1016/j.anbehav.2011.10.009
Klaassen M (1996) Metabolic constraints on long-distance migration in birds. J Exp Biol 199:57–64. http://www.ncbi.nlm.nih.gov/pubmed/9317335
Klaassen M, Hoye BJ, Nolet BA, Buttemer WA (2012) Ecophysiology of avian migration in the face of current global hazards. Phil Trans Soc Lond B 367:1719–1732. doi:10.1098/rstb.2012.0008
Kokko H (1999) Competition for early arrival in migratory birds. J Anim Ecol 68:940–950. doi:10.1046/j.1365-2656.1999.00343.x
Lupi S, Goymann W, Cardinale M, Fusani L (2016) Physiological conditions influence stopover behaviour of short-distance migratory passerines. J Ornithol 157:583–589. doi:10.1007/s10336-015-1303-5
Maggini I, Bairlein F (2010) Endogenous rhythms of seasonal migratory body mass changes and nocturnal restlessness in different populations of Northern Wheatear Oenanthe oenanthe. J Biol Rhythms 25:268–276. doi:10.1177/0748730410373442
Maggini I, Bairlein F (2011) Body condition and stopover of trans-Saharan spring migrant passerines caught at a site in southern Morocco. Ring Migr 26:31–37. doi:10.1080/03078698.2011.586591
McWilliams SR, Guglielmo C, Pierce B, Klaassen M (2004) Flying, fasting, and feeding in birds during migration: A nutritional and physiological ecology perspective. J Avian Biol 35:377–393
McWilliams SR, Karasov WH (2005) Migration takes guts. digestive physiology of migratory birds and its ecological significance. In: Marra P, Greenberg R (eds) Birds of two worlds. Smithsonian Institution Press, Washington DC, pp 67–79
Mettke-Hofmann C, Lorentzen S, Schlicht E, Schneider J, Werner F (2009) Spatial neophilia and spatial neophobia in resident and migratory Warblers (Sylvia). Ethology 115:482–492. doi:10.1111/j.1439-0310.2009.01632.x
Pilastro A, Spina F (1997) Ecological and morphological correlates of residual fat reserves in passerine migrants at their spring arrival in southern Europe. J Avian Biol 28:309–318. doi:10.2307/3676944
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org
Salewski V, Schaub M (2007) Stopover duration of Palearctic passerine migrants in the western Sahara—independent of fat stores? Ibis 149:223–236. doi:10.1111/j.1474-919X.2006.00608.x
Sandberg R, Moore FR (1996) Fat stores and arrival on the breeding grounds: reproductive consequences for passerine migrants. Oikos 77:577–581. doi:10.2307/3545949
Schaub M, Jenni L, Bairlein F (2008) Fuel stores, fuel accumulation, and the decision to depart from a migration stopover site. Behav Ecol 19:657–666. doi:10.1093/beheco/arn023
Schmaljohann H, Liechti F, Bruderer B (2007) Songbird migration across the Sahara: the non-stop hypothesis rejected! Proc R Soc B 274:735–739. doi:10.1098/rspb.2006.0011
Schmaljohann H, Korner-Nievergelt F, Naef-Daenzer B, Nagel R, Maggini I, Bulte M, Bairlein F (2013) Stopover optimization in a long-distance migrant: the role of fuel load and nocturnal take-off time in Alaskan northern wheatears (Oenanthe oenanthe). Front Zool 10:26. doi:10.1186/1742-9994-10-26
Smith AD, McWilliams SR (2014) What to do when stopping over: behavioral decisions of a migrating songbird during stopover are dictated by initial change in their body condition and mediated by key environmental conditions. Behav Ecol 25:1423–1435. doi:10.1093/beheco/aru148
Smolinsky JA, Diehl RH, Radzio TA, Delaney DK, Moore FR (2013) Factors influencing the movement biology of migrant songbirds confronted with an ecological barrier. Behav Ecol Sociobiol 67:2041–2051. doi:10.1007/s00265-013-1614-6
Spina F, Volponi S (2008) Atlante della Migrazioni degli Uccelli in Italia. II. Passeriformi. Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA), Ozzano dell’Emilia (Bologna)
Tsvey A, Bulyuk VN, Kosarev V (2007) Influence of body condition and weather on departures of first-year European robins, Erithacus rubecula, from an autumn migratory stopover site. Behav Ecol Sociobiol 61:1665–1674. doi:10.1007/s00265-007-0397-z
van Gils JA, Munster VJ, Radersma R, Liefhebber D, Fouchier RAM, Klaassen M (2007) Hampered foraging and migratory performance in swans infected with low-pathogenic avian influenza A virus. PLoS One 184:e184. doi:10.1371/journal.pone.0000184
Yong W, Moore FR (1993) Relation between migratory activity and energetic condition among thrushes (Turdinae) following passage across the Gulf of Mexico. Condor 95:934–943. doi:10.2307/1369429
Acknowledgements
Open access funding provided by University of Veterinary Medicine Vienna. We thank Fernanda Machado Tahamtani and Marco Cianchetti Benedetti for help during fieldwork. Three anonymous reviewers provided useful comments to a previous version of the manuscript. This is publication N. 60 of the Progetto Piccole Isole. All experimental procedures were authorized by the Regione Lazio and are in accordance with Italian law.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This study was supported by the Max Planck Society (Prof. M. Gahr), by the University of Ferrara (FAR 2011 and 2013), and by the Vigoni program of the Italian Ministry of University and Research (MIUR) and the German Academic Exchange Service (DAAD).
Conflict of interest
The authors declare that that they have no conflict of interest.
Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Communicated by N. Chernetsov.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Lupi, S., Maggini, I., Goymann, W. et al. Effects of body condition and food intake on stop-over decisions in Garden Warblers and European Robins during spring migration. J Ornithol 158, 989–999 (2017). https://doi.org/10.1007/s10336-017-1478-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-017-1478-z