
Abstract
Wheat cultivars exposed to optimal photoperiod and vernalization treatments still exhibit differences in flowering time, referred to as earliness per se (Eps). We previously identified the Eps-A m 1 locus from Triticum monococcum and showed that the allele from cultivated accession DV92 significantly delays heading time and increases the number of spikelets per spike relative to the allele from wild accession G3116. Here, we expanded a high-density genetic and physical map of the Eps-A m 1 region and identified the wheat ortholog of circadian clock regulator EARLY FLOWERING 3 (ELF3) as a candidate gene. No differences in ELF3 transcript levels were found between near-isogenic lines carrying the DV92 and G3116 Eps-A m 1 alleles, but the encoded ELF3 proteins differed in four amino acids. These differences were associated with altered transcription profiles of PIF-like, PPD1, and FT1, which are known downstream targets of ELF3. Tetraploid wheat lines with combined truncation mutations in the A- and B-genome copies of ELF3 flowered earlier and had less spikelets per spike than the wild-type control under short- and long-day conditions. Both effects were stronger in a photoperiod-sensitive than in a reduced photoperiod-sensitive background, indicating a significant epistatic interaction between PPD1 and ELF3 (P < 0.0001). By contrast, the introgression of the T. monococcum chromosome segment carrying the Eps-A m 1 allele from DV92 into durum wheat delayed flowering and increased the number of spikelets per spike. Taken together, the above results support the hypothesis that ELF3 is Eps-A m 1. The ELF3 alleles identified here provide additional tools to modulate reproductive development in wheat.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
More than 700 million tons of wheat is grown every year in very diverse environments, providing a major source of calories and proteins to the human population. The ability of wheat to adapt to these different conditions has been favored by a rapidly evolving genome combined with the buffering effect of recent polyploidization events (Dubcovsky and Dvorak 2007). This plasticity, coupled with strong selection pressures during the expansion of agriculture and wheat cultivation throughout the world, contributed to a large genetic diversity in genes regulating reproductive development. This diversity has been used by wheat breeders to optimize the utilization of available natural resources during plant growth and grain filling and to maximize grain yield in different environments.
In temperate cereals, such as wheat and barley, the initiation of the reproductive phase is regulated by the integration of two main seasonal signals: photoperiod (day length) and vernalization (prolonged exposure to low temperatures). The photoperiod response is mainly regulated by PPD1, a member of the PSEUDO-RESPONSE REGULATOR (PRR) gene family, which upregulates the expression FT1 under long days (Turner et al. 2005). FT1 encodes a mobile protein, homologous to the Arabidopsis FLOWERING LOCUS T (FT) (Yan et al. 2006), which is transported from the leaves to the shoot apical meristem (SAM) (Corbesier et al. 2007; Tamaki et al. 2007). Once in the apex, FT1 forms a protein complex that binds the promoter of the meristem identity gene VRN1 (Li et al. 2015) and upregulates its expression above the critical threshold required to initiate the transition from the vegetative to the reproductive stage (Distelfeld et al. 2009). During the fall, the expression of FT1 in the leaves is downregulated by the flowering repressor VRN2, preventing flowering before winter (Yan et al. 2004). During the winter, cold temperatures induce the expression of VRN1 in the leaves and result in the repression of VRN2 (Chen and Dubcovsky 2012). In the absence of VRN2, the increase of day length during the spring results in the induction of FT1 and the acceleration of the transition to the reproductive stage (Distelfeld et al. 2009).
Even when the vernalization and photoperiod requirements have been fulfilled, there are still residual differences in flowering time among wheat accessions, which are usually referred to as earliness per se (Eps) (Flood and Halloran 1984; Hoogendoorn 1985; Worland and Law 1986; Slafer 1996). These differences are affected by temperature (Slafer and Rawson 1994, 1995a, b) and are important for the fine-tuning of flowering and the adaptation of wheat to different environments. Several Eps loci have been mapped on different wheat and barley chromosomes (Scarth and Law 1984; Hoogendoorn 1985; Miura and Worland 1994; Laurie et al. 1995; Worland 1996; Bullrich et al. 2002; Griffiths et al. 2009; Gawronski and Schnurbusch 2012; Zikhali et al. 2014, 2015), but only a few of the underlying Eps genes have been identified so far (Comadran et al. 2012; Faure et al. 2012; Zakhrabekova et al. 2012; Gawronski et al. 2014).
The temperate cereal homolog of the Arabidopsis circadian clock regulator LUX ARRHYTHMO (LUX) was proposed as a promising candidate gene for the Earliness per se 3 (Eps-A m 3) locus in Triticum monococcum L. (2n = 14, genomes AmAm; Mizuno et al. 2012; Gawronski et al. 2014) and for the colinear early maturity 10 (eam10) locus in barely (Hordeum vulgare L., 2n = 14, genomes HH; Gawronski and Schnurbusch 2012; Campoli et al. 2013). A complete deletion of LUX in the eps-A m 3 mutant and a mutation in a highly conserved region of the LUX protein in eam10 were linked with early flowering under both long-day (LD) and short-day (SD) conditions. The early flowering in both mutants was associated with arrhythmic transcript patterns of central and output circadian clock genes under SD and constant ambient conditions and the upregulation of PPD1 and FT1 transcript levels (Mizuno et al. 2012; Campoli et al. 2013; Gawronski et al. 2014). The effects of the LUX mutations were larger in the presence of the photoperiod-sensitive allele Ppd-H1 than in the presence of the photoperiod-insensitive allele ppd-H1, suggesting that the effect of LUX was mediated in part by the negative regulation of Ppd-H1 (Mizuno et al. 2012; Campoli et al. 2013). The eps-A m 3 mutant also showed temperature-dependent variation in spike development (Gawronski et al. 2014).
The early maturity 8 (eam8, also known as mat-a) mutants in barley show similar phenotypic effects as the eam10 mutants described above. The eam8 gene encodes a homolog of the Arabidopsis circadian clock gene EARLY FLOWERING 3 (Faure et al. 2012; Zakhrabekova et al. 2012), which is known to interact with LUX and EARLY FLOWERING 4 (ELF4) to form a trimeric protein complex known as the “evening complex” (Nusinow et al. 2011). In Arabidopsis, the evening complex functions as a transcriptional regulator that represses the expression of growth-promoting transcription factors PHYTOCHROME-INTERACTING FACTOR 4 (PIF4) and PIF5 (Nusinow et al. 2011), as well as PSEUDO-RESPONSE REGULATOR (PRR) genes PRR7 and PRR9 (Dixon et al. 2011; Helfer et al. 2011; Kolmos et al. 2011; Chow et al. 2012; Herrero et al. 2012). The participation of ELF3 and LUX in the same protein complex is consistent with the similar phenotypic effects observed in loss-of-function mutations in either of the two genes, and in their similar epistatic interactions with PPD1 (Faure et al. 2012; Mizuno et al. 2012; Campoli et al. 2013). Both mutants show early flowering under LD and SD and a transcriptional upregulation of PPD1, suggesting that in the temperate cereals the evening complex acts as a repressor of PPD1 (Faure et al. 2012; Zakhrabekova et al. 2012, Mizuno et al. 2012; Campoli et al. 2013).
In Triticum aestivum, a deletion of the chromosomal region including ELF3 was linked to the earliness per se locus Eps-D1, which causes early flowering and altered expression of circadian clock gene GIGANTEA (GI) (Zikhali et al. 2015). The earliness per se gene Eps-A m 1 from T. monococcum was mapped on the distal region of chromosome arm 1AmL, which is colinear to both eam8 and Eps-D1 (Bullrich et al. 2002; Valarik et al. 2006; Zakhrabekova et al. 2012). The Eps-A m 1 allele from cultivated T. monococcum ssp. monococcum (accession DV92) was associated with delayed heading time and increased number of spikelets per spike relative to the allele from wild T. monococcum ssp. aegilopoides (accession G3116). The effect of this locus was larger when plants were grown at 16 °C compared to 23 °C, suggesting a role of temperature on the modulation of the effects of this gene (Bullrich et al. 2002; Appendino and Slafer 2003; Lewis et al. 2008).
In this study, we developed a high-density genetic and physical map of the Eps-A m 1 region and show that ELF3 is a strong candidate gene for Eps-A m 1. We found four amino acid changes between Eps-A m 1 alleles that were associated with differences in the transcription profiles of circadian clock genes and ELF3 downstream targets. Tetraploid wheat lines carrying loss-of-function mutations in both the A- and B-genome copies of elf3 showed early flowering and reduced spikelet number and exhibited significant epistatic interactions with PPD1 for both traits under SD and LD conditions. By contrast, introgression of the Eps-A m 1 allele from cultivated T. monococcum accession DV92 into durum wheat significantly delayed flowering time and increased the number of spikelets per spike.
Materials and methods
Materials
We developed diploid wheat near-isogenic lines (NILs) for the Eps-A m 1 alleles by backcrossing six times the Eps-A m 1 locus from the wild T. monococcum ssp. aegilopoides accession G3116 (winter growth habit) into the cultivated T. monococcum ssp. monococcum accession DV92 (spring growth habit). The Eps-A m 1 allele from G3116 is associated with early heading and is referred to as Eps-A m 1-e, whereas the allele from DV92 is associated with late heading and is referred to as Eps-A m 1-l (Lewis et al. 2008). After six backcrosses to DV92, these NILs are expected to be more than 98 % identical to the recurrent parent. We confirmed that both NILs have a spring growth habit determined by a nonfunctional vrn-A m 2 allele (Yan et al. 2004) and that both are photoperiod sensitive (early flowering under LD and extremely late flowering under SD) (Appendino and Slafer 2003).
The tetraploid wheat cultivar Kronos (Triticum turgidum ssp. durum, 2n = 28, genomes AABB) was used to generate the TILLING population analyzed in this study (Uauy et al. 2009). This population included 1536 EMS-mutagenized Kronos lines, with DNAs organized in 384 pools of four individual DNAs (Uauy et al. 2009). Kronos carries the Vrn-A1c allele for spring growth habit (Fu et al. 2005; Chen and Dubcovsky 2012) and the Ppd-A1a allele for reduced photoperiodic response (Chen et al. 2014). Plants carrying the Ppd-A1a allele are usually referred in the wheat literature as “photoperiod insensitive” because they flower earlier under SD than plants carrying the wild-type Ppd-A1b allele, reducing the differences in flowering between LD and SD. However, we prefer the term “reduced photoperiodic response” because plants carrying the Ppd-A1a allele still flower significantly earlier when grown under LD than when grown under SD, demonstrating that this allele is still able to respond to differences in photoperiod (Chen et al. 2014).
High-density genetic map of Eps-A m 1
Seeds for the high-density map were generated from BC6 and BC6F2 T. monococcum NILs heterozygous for Eps-A m 1 flanking genes MOT1 (MODIFIER OF TRANSCRIPTION 1) and SMP (SM-LIKE PROTEIN). The isogenic background of these lines reduced genetic variability and facilitated the precise mapping of the Eps-A m 1 locus as a Mendelian trait rather than as a quantitative trait locus (QTL). Young seedlings were screened with molecular markers for MOT1 and SMP, and only plants carrying recombination events between these two markers were phenotyped.
Previous studies have shown that the differences in flowering time between the Eps-A m 1-e and Eps-A m 1-l alleles can be maximized by exposing the plants to fluorescent lights at a low fluency (160 μM m−2 s−1), a long-day photoperiod (16 h of light and 8 h of darkness), and constant cool temperatures (16 °C) (Lewis et al. 2008). The same environmental conditions were used in this study to phenotype the progeny tests of the plants carrying the closest recombination events to Eps-A m 1. Some preliminary experiments were performed under greenhouse conditions (20–25 °C, natural day length supplemented with incandescent lamps to extend photoperiod to 16 h), where the differences were smaller but still significant. Heading times were registered from sowing to spike emergence.
Molecular markers used for the construction of the high-density genetic map are listed in Supplementary Table S1. DNA extraction and PCR procedures were performed as described before (Kippes et al. 2014). Markers were developed using BAC sequences from the T. monococcum physical map (see next section) and sequences from wheat genes homologous to Brachypodium distachyon genes located in the region colinear to Eps-A m 1 (Faricelli et al. 2010). Eps-A m 1 flanking genes FTSH4 (FTSH PROTEASE 4) and SMP were used to search a genomic database of B. distachyon (http://www.phytozome.net) and establish the colinear target region. The genes detected in this region were then used as queries to search a T. aestivum L. (2n = 2x = 42, AABBDD) database of flow-sorted chromosomes arms developed by the International Wheat Genome Sequencing Consortium (IWGSC 2014, http://www.wheatgenome.org/). A reverse BLASTP search from the best wheat candidate to the Brachypodium proteome was done, to confirm that the correct homolog was found.
Physical map
A BAC library from T. monococcum ssp. monococcum accession DV92 (Lijavetzky et al. 1999) was screened with PCR markers developed for the Eps-A m 1 region (Supplementary Table S2). DNA was isolated from the selected BAC clones using the QIAGEN® Large-Construct Kit (Qiagen, USA) and then fragmented in a Covaris ultrasonicator (peak power, 175; duty factor, 10 %; cycles/burst, 200; time, 110 s). Libraries for Illumina sequencing were prepared using the KAPA LTP Library Preparation Kit Illumina platforms (KAPA Biosystems, USA). The quality of the libraries was assessed in an Agilent 2100 Bioanalyzer instrument using the High Sensitivity DNA Kit (Agilent Technologies, USA). Libraries were sequenced using 100-bp paired-end reads on Illumina HiSeq2000. Individual BAC coverage was higher than 100×.
Sequence assembly was performed using CLC 6.5 Beta 4 software (CLC-Bio, USA). Once sequences were assembled, repetitive elements were identified using the Triticeae Repeat Sequence Database (http://wheat.pw.usda.gov/ITMI/Repeats/blastrepeats3). The nonrepetitive sequences were then annotated using a combination of tools, including comparative genomics analyses, BLAST searches of other annotated grass genomes, and the annotation pipeline Triannot (Leroy et al. 2011). Nonrepetitive sequences of the newly sequenced BACs were used to develop new primers to rescreen the BAC library. This process was reiterated several times to expand the physical map of the Eps-A m 1 region. Individual BAC sequences have been deposited in GenBank (Supplementary Table S3).
Characterization of Eps-A m 1 candidate genes and its potential targets
Candidate genes completely linked to Eps-A m 1 in the high-density genetic map were amplified from both parental lines of the T. monococcum mapping population (DV92 and G3116) and sequenced. GenBank accession numbers are listed in Supplementary Table S3; primers used are listed in Supplementary Table S4. The predicted protein products of the three candidate genes were compared between the parental lines using BLASTP. Amino acid polymorphisms are described using a letter indicating the amino acid in the T. urartu (PI 428198) protein, followed by the position of that amino acid from the initial methionine in the T. urartu reference protein (Supplementary Figs. S1–S3), and by a letter describing the derived amino acid. For the three candidate proteins characterized in this study, T. urartu carries the ancestral state in all the analyzed amino acid positions. Therefore, in this study, the first letter in a polymorphism corresponds to the ancestral stage and the last letter to the derived stage. Ancestral stages were inferred from the alignment of the predicted wheat proteins with the corresponding orthologs from Hordeum vulgare, B. distachyon, Sorghum bicolor, Setaria italica, Oryza sativa, Zea mays, and Arabidopsis (Supplementary Figs. S1–S3). Multiple alignments were performed using ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/). The potential effect of amino acid changes was predicted using multiple tools including BLOSUM62 matrix scores (Henikoff and Henikoff 1992), PROVEAN scores (http://provean.jcvi.org; Choi et al. 2012), and PolyPhen scores (http://genetics.bwh.harvard.edu/pph2/index.shtml; Adzhubei et al. 2010).
The expression levels of the Eps-A m 1 candidate genes and some of the known targets of the main candidate protein were characterized using quantitative reverse transcription PCR (qRT-PCR). T. monococcum NILs carrying the Eps-A m 1-e (early flowering) and the Eps-A m 1-l (late flowering) alleles were grown in a growth chamber under continuous temperature (16 °C) and long-day photoperiod (16 h fluorescent lights at 160 μM m−2 s−1). Five weeks after sowing, when plants carrying the Eps-A m 1-e allele were at the terminal spikelet stage and plants carrying the Eps-A m 1-l allele were at the double ridge stage, tissue was collected from young leaves every 4 h during a 24-h period. Six individual plants were sampled at each time point.
RNA samples were extracted using the Spectrum Plant Total RNA Kit (Sigma-Aldrich). First-strand cDNAs were synthesized from 1 μg of total RNA using the High Capacity Reverse Transcription kit (Applied Biosystems). Quantitative PCR was performed using SYBR Green and a 7500 Fast Real-Time PCR system (Applied Biosystems). Primers used for SYBR GREEN quantitative PCR are listed in Supplementary Table S5. ACTIN was used as an endogenous control. Transcript levels for all genes are expressed as linearized fold-ACTIN levels calculated by the formula 2(ACTIN CT − TARGET CT) ± standard error (SE) of the mean. The resulting number indicates the ratio between the initial number of molecules of the target gene and the number of molecules of ACTIN.
Screening of a tetraploid wheat TILLING population
The complete coding regions of the A- and B-genome copies of candidate gene ELF3 were obtained from available sequences from T. aestivum cultivar Chinese Spring (IWGSC 2014, http://www.wheatgenome.org), and from T. turgidum ssp. durum cultivar Kronos transcriptome (Krasileva et al. 2013; http://wheat.pw.usda.gov/GG2/WheatTranscriptome/). Genome-specific primers were developed (Supplementary Table S6) and tested using Chinese Spring nullisomic-tetrasomic lines N1AT1D and N1BT1D, and confirmed by DNA sequencing.
A screening of DNAs from 1536 TILLING lines was performed using 384 pools, each including four DNAs. PCR amplification products of the target region were digested with CelI, as described before (Uauy et al. 2009). Individual mutant lines were identified within the selected pools, and the PCR amplification products were sequenced to characterize the mutations. Lines carrying mutations that introduced premature stop codons in the A- or the B-genome copies of the candidate gene ELF3 were selected and were then intercrossed to combine the mutations in both homoeologs. The resulting lines were backcrossed to the non-mutagenized variety Kronos for two generations to reduce the number of background mutations. Mutations were selected by sequencing in each backcross generation. BC2 plants heterozygous for mutations in the two ELF3 homoeologs were selected and self-pollinated. Plants with no functional copies of ELF3 (elf3-null) and control plants homozygous for the wild-type ELF3 alleles (ELF3-WT) were selected from the segregating BC2F2 plants.
Epistatic interactions between ELF3 and PPD1
The tetraploid elf3-null lines were also crossed to a Kronos near-isogenic line carrying the wild-type photoperiod-sensitive PPD-A1b allele, developed before (Pearce et al. 2013). A BC2 line heterozygous for ELF3 and PPD-A1 was identified using molecular markers and was self-pollinated to produce a BC2F2 population segregating for both genes. A screening of the segregating population identified lines homozygous for the four possible allelic combinations: ELF3-WT/PPD-A1a, ELF3-WT/PPD-A1b, elf3-null/PPD-A1a, and elf3-null/PPD-A1b. Primers used to genotype ELF3 are listed in Supplementary Table S6. Markers used for PPD-A1 have been described before (Wilhelm et al. 2009).
BC2F3 plants carrying the four allelic combinations were grown under both SD (8 h light) and LD (16 h light) photoperiods using fluorescent lights and a constant temperature of 16 °C. Heading time and number of spikelets per spike were determined for each plant, and the effects of the individual alleles and their epistatic interactions were statistically evaluated using two by two factorial ANOVAs. Statistical analyses were performed using SAS program version 9.4.
Introgression of the Eps-A m 1-l locus into tetraploid wheat
The chromosome region including Eps-A m 1 from T. monococcum accession DV92 (2n = 14, AmAm) was introgressed into T. turgidum ssp. durum cultivar CBW0112 (2n = 28; AABB). To generate this material, we first intercrossed T. turgidum ssp. durum cultivar Langdon and T. monococcum ssp. monococcum cultivar DV92 and treated the ABAm hybrid with colchicine to double the chromosome number. We backcrossed the resulting AABBAmAm amphyploid to CBW0112 for five generations, selecting the Eps-A m 1 allele in each crossing cycle with flanking markers wg241 and NDK3 (NUCLEOSIDE DIPHOSPHATE KINASE 3) (Supplementary Table S1). A schematic representation of the process is presented in Supplementary Fig. S4.
CBW0112 BC5F3 near-isogenic lines homozygous for the Eps-A m 1 segment from DV92 were selected for phenotypic characterization. Sister lines homozygous for the Eps-A1 allele from the A-genome of tetraploid wheat were used as negative controls. Lines were grown under controlled conditions (16 °C constant temperature, 16 h fluorescent lights), and heading time and spikelet number were registered.
When lines were at BC3F2, seven SSR markers and three CAP markers previously mapped to wheat chromosome 1A were tested to determine the size of the introgressed 1Am chromosome segment (Supplementary Table S7, Supplementary Fig. S4).
Results
Redefinition of the target chromosome region for Eps-A m 1
In a previous study, the T. monococcum Eps-A m 1 locus was mapped linked to genes MOT1 and FTSH4 (Faricelli et al. 2010; Fig. 1a). Two years later, two independent studies in barley identified ELF3 as the gene responsible for the eam8 mutation (Faure et al. 2012; Zakhrabekova et al. 2012), which maps very close to MOT1 and FTSH4. Since both Eps-A m 1 and eam8 loci affect similar traits (Lewis et al. 2008; Faure et al. 2012) and are physically close to each other (only 28 kb apart in Brachypodium), we decided to add ELF3 to our previous map and revisit the critical recombination events (Fig. 2a, b).

a T. monococcum high-density genetic map based on the analysis of 10,000 gametes, showing the previous candidate region for Eps-A m 1 in light gray (Faricelli et al. 2010). The reevaluation of critical NIL 502 resulted in a new candidate region for Eps-A m 1, indicated in darker gray. b Graphical genotypes and heading times of a progeny test of critical NIL 502. Different colors indicate chromosome regions homozygous for the DV92 allele (dark gray), homozygous for the G3116 allele (light gray), or heterozygous (diagonal gray lines). Heading time for each line is shown as the mean of at least five plants ± standard error of the mean. Values followed by different letters are significantly different from each other (P < 0.01). This progeny test confirmed that the Eps-A m 1 locus in NIL 502 was distal to the MOT1/FTSH4 locus

a Triticum monococcum high-density genetic map. Genetic distances are based on the analysis of 5800 gametes. Putative genes are color-coded and indicated as circles. b Triticum monococcum physical map. Sequenced BACs are indicated as blue bars and assembly gaps as striped bars. c Triticum aestivum 1AL IWGSC contigs colinear to the Eps-A m 1 region. Contig names were shortened, and only the contig number is shown. d Brachypodium distachyon region colinear to Eps-A m 1
We sequenced ELF3 in the parental lines DV92 (AC270217.1) and G3116 (GenBank KU570055) and used one of the discovered SNPs to develop a codominant marker (Supplementary Table S1). Using this marker, we mapped ELF3 completely linked to ADK1 (similar to rice putative kinase ADK1, AAT44307) in the same T. monococcum segregating population used before to map Eps-A m 1 (Fig. 1a, Faricelli et al. 2010). In this population, a single recombination event in NIL 502 (Fig. 1a, b) separated MOT1/FTSH4 from ADK1/ELF3. NIL 502 carries the G3116 alleles for MOT1/FTSH4 and the DV92 alleles for ADK1/ELF3 (Fig. 1b), so if this line flowers early it would indicate that Eps-A m 1 is linked to MOT1/FTSH4 and if it flowers late it would indicate that Eps-A m 1 is linked to ADK1/ELF3.
NIL 502 was initially classified as an early flowering line (Lewis et al. 2008), but a reexamination of the original progeny test showed that it had an intermediate flowering time when compared to the early and late heading NILs. To determine if the intermediate flowering time was the result of more than one gene affecting flowering segregating in this region or simply the pleiotropic effect of other genes still segregating in the genetic background of this line, we backcrossed NIL 502 twice to a NIL carrying the G3116 alleles for all markers in this region. The resulting BC2F2 plants were homozygous G3116 for MOT1/FTSH4 and segregated for the distal ADK1/ELF3 genes (Fig. 1b). These lines showed a clear segregation for heading time that was perfectly associated to the ADK1/ELF3 alleles (Fig. 1b). In addition, the homozygous recombinant lines showed almost identical flowering time as the parental line DV92 (Fig. 1b). These two results demonstrated that NIL 502 carries the Eps-A m 1 allele for late flowering and that, therefore, Eps-A m 1 is linked to ADK1/ELF3 rather than to MOT1/FTSH4 as suggested before (Faricelli et al. 2010). Erratum notes are being submitted to the original papers correcting the phenotyping of NIL 502 (Lewis et al. 2008; Faricelli et al. 2010).
High-density genetic map of Eps-A m 1 in T. monococcum
To further characterize the relationship between Eps-A m 1 and ELF3, we developed a new high-density genetic map for the region between MOT1 and SMP (Fig. 2a). Using these two flanking markers, we screened 2900 BC6F2 NILs (5800 gametes) and identified 22 lines with recombination events in the target region. Based on the heading time of BC6F3 progenies of the selected recombinants grown under controlled environment conditions, the Eps-A m 1 locus was mapped 0.20 cM proximal to SMP and 0.18 cM distal to MOT1 (Fig. 2a). The 10 recombination events found between MOT1/FTSH4 and Eps-A m 1 clearly demonstrated that neither MOT1 nor FTSH4 are viable candidate genes for Eps-A m 1. This result is consistent with the conclusion obtained from the reanalysis of the progeny of the critical NIL 502 reported above.
The ELF3 gene was completely linked to the flowering phenotype in all the recombinant lines, suggesting that it is a viable candidate gene for Eps-A m 1. To further delimit the Eps-A m 1 candidate region, we designed additional markers based both on B. distachyon genes flanking ELF3 and on nonrepetitive sequences obtained from the T. monococcum BACs identified during the construction of the physical map (see next section, Fig. 2b).
The B. distachyon orthologs of wheat Eps-A m 1 flanking genes FTSH4 (Bradi2g14330) and SMP (Bradi2g14260) define a 47-kb genomic region including the Brachypodium orthologs of ADK1 (Bradi2g14310), ELF3 (Bradi2g14290), and five additional annotated genes (Table 1 and Fig. 2d). These Brachypodium genes were used as queries to identify the closest homologs in the T. aestivum draft genome sequence (IWGSC 2014). Three wheat orthologs were identified using this strategy, in addition to ADK1 and ELF3, and were mapped to the target region (Fig. 2c). The wheat ortholog of Bradi2g14280 was not detected in the T. aestivum database but was later found in one of the BAC sequences and was incorporated into the high-density genetic map (Fig. 2). The closest wheat homolog of Bradi2g14320 was found in wheat chromosome 2AS and, therefore, was eliminated from further analyses. One additional gene, designated as GRIK1-like, was found in the same T. aestivum contig carrying the ELF3 gene (IWGSC-contig 3795905), but was not present in the Brachypodium colinear region. GRIK1-like encodes a serine/threonine-protein kinase similar to GRIK1 (GEMINIVIRUS REP INTERACTING KINASE 1) from Arabidopsis.
After the inclusion of the additional markers in the high-density genetic map, the Eps-A m 1 region was delimited to a 0.1-cM interval flanked by ADK1/CK1-like (CASEIN KINASE I ISOFORM DELTA-LIKE) on the proximal side and ACA7 (ALPHA CARBONIC ANHYDRASE 7-LIKE) on the distal side. In the high-density mapping population, Eps-A m 1 was completely linked to ELF3, GRIK1-like, and PWWP1 (PWWP domain-containing protein) (Fig. 2a), which are described briefly below and in more detail in the “Discussion” section.
ELF3 is a circadian clock gene involved in the regulation of flowering time in Arabidopsis, rice, legumes, and barley (Hicks et al. 2001; Matsubara et al. 2012; Saito et al. 2012; Weller et al. 2012; Faure et al. 2012; Zakhrabekova et al. 2012). GRIK1-like encodes a putative serine/threonine kinase that is homologous to Arabidopsis GRIK1. This Arabidopsis gene is part of a cascade that coordinates the metabolic requirements of rapidly growing cells and geminivirus-infected cells (Shen et al. 2009). PWWP1 encodes a putative PWWP domain-containing protein with no clear homologs in Arabidopsis. This domain is found in proteins that are involved in cell division, growth, and differentiation, but the actual function of this gene is currently unknown. Among the three Eps-A m 1 candidate genes, we prioritized ELF3 for further characterization based on its known role in the regulation of flowering in several plant species.
Physical map of the Eps-A m 1 region
A BAC library from T. monococcum accession DV92 including 276,000 clones (5.6-fold coverage, Lijavetzky et al. 1999) was screened with markers for flanking genes FTSH4 and SMP and for the linked candidate gene ELF3.
The chromosome walk from the proximal site started from the distal border of BAC 707L18, which included genes MOT1, FTSH4, and ADK1. Six BACs were mapped to this region, covering ~270 kb. Gene CK1-like, an ortholog of Bradi2g14301, was also detected in this region (Fig. 2b).
On the distal side of Eps-A m 1, a screening using primers for the SMP gene yielded BACs 102H21 and 73J03, which covered ~197 kb. Genes SMP, ACA7, and NLE (NOTCHLESS), orthologous to Bradi2g14260, Bradi2g14270, and Bradi2g14275, were detected in close proximity, within an 8.6-kb region present in both BACs (Fig. 2b). Although the physical distance between genes NLE and SMP is only of 346 bp, we detected 10 recombination events between them. The analysis of the sequence between these two genes revealed the presence of trinucleotide repeats, which have been associated before to fragile sites and chromosomal instability hot spots that can lead to high recombination frequencies (Aguilera and Gómez-González 2008).
A third screening of the T. monococcum BAC library was performed using PCR markers for ELF3, which was completely linked to Eps-A m 1. Two BACs were sequenced, and from their borders, chromosome walks were initiated in both directions. A total of nine BACs were mapped to this contig, covering ~570 kb. Genes ELF3, GRIK1-like, and PWWP1 were identified in this region (Fig. 2b).
Nine of the 10 genes annotated in the colinear region in Brachypodium are included in the T. monococcum physical map, and the last one (Bradi2g14320) has its closest homolog in a different wheat chromosome. Since all the genes annotated in the colinear region in Brachypodium have been accounted for, any additional wheat genes found in the two gaps still present in the T. monococcum physical map (Fig. 2b) are not expected to be colinear with Brachypodium.
In summary, we constructed and sequenced an ~1-Mb physical map of the MOT1-SMP region, which includes 10 wheat genes. The resulting gene density of roughly one gene per 100 kb found in the Eps-A m 1 region is very similar to the average of one gene per 96 kb found before for an 18.2-Mb region in wheat chromosome 3B (Choulet et al. 2010). The T. monococcum genes were distributed in small islands including one to three genes separated by large regions of repetitive elements, which is similar to distributions found in other wheat genome regions (Gottlieb et al. 2013).
Characterization of Eps-A m 1 candidate proteins
To help us prioritize or discard any of the three T. monococcum candidate genes completely linked to Eps-A m 1, we compared the predicted protein sequences encoded by the parental lines, DV92 and G3116, using primers listed in Supplementary Table S4. We also analyzed their expression profiles in NILs carrying the DV92 or G3116 Eps-A m 1 alleles using the qRT-PCR primers listed in Supplementary Table S5.
The predicted ELF3 protein showed four polymorphisms between the two parental lines: V364L, G681R, G700D, and G718A, which are predicted to have intermediate to large effects on protein function based on either BLOSUM62, PROVEAN, or PolyPhen-2 scores (Table 2). None of these polymorphisms were located within the conserved blocks defined by Liu et al. (2001) (Supplementary Fig. S1). Comparison of the T. monococcum sequences with homologous proteins from other species indicates that DV92 has the derived amino acids for the V364L and G681R polymorphisms and G3116 has the derived amino acids at G700D and G718A (Supplementary Fig. S1). The ancestral ELF3 haplotype (haplotype-A) is present in T. urartu, in the A- and B-genome of tetraploid wheat and, with single amino acid changes, in the D-genome of hexaploid wheat, barley, Brachypodium, and sorghum (Supplementary Fig. S1 and Table S8). The G3116 haplotype (haplotype-B) was frequent among T. monococcum ssp. aegilopoides accessions and among T. monococcum ssp. monococcum accessions with a winter growth habit (Supplementary Table S8), whereas the DV92 haplotype (haplotype-C) was predominant among T. monococcum ssp. monococcum accessions with a spring growth habit. Interestingly, five out of the seven T. monococcum ssp. monococcum accessions carrying ELF3 haplotypes A or B were collected in Turkey, where this species was initially domesticated (Heun et al. 1997).
To analyze the linkage between ELF3 haplotypes and Eps1 alleles, we evaluated a segregating BC1F2 population generated by crossing T. monococcum ssp. monococcum accession PI 355522 (ELF3 haplotype-B) to DV92 (ELF3 haplotype-C) and then backcrossing the F1 to DV92. These lines were grown in a controlled environment (16 °C and 16 h of light), and heading time and spikelet number were registered. This experiment confirmed that PI 355522 carries the Eps-A m 1-e allele for early flowering and reduced number of spikelets (Supplementary Table S9).
In a separate experiment, we characterized 10 additional T. monococcum accessions from Supplementary Table S8 under saturated vernalization (6 weeks at 4 °C) and photoperiod conditions (16 h fluorescent light). Under these conditions, we found that plants from the four T. monococcum accessions carrying the ELF3 haplotype-B headed more than 1 month earlier than plants from the six accessions carrying the ELF3 haplotype-C (Supplementary Table S10). These results suggest that the four haplotype-B lines carry the Eps-A m 1-e allele and the six haplotype-C lines carry the Eps-A m 1-l, but this linkage was not tested in this study.
The GRIK1-like protein showed three polymorphisms between G3116 and DV92 (D77E, A228S, and D351N, Supplementary Table S8), which are all predicted to have limited effects on protein function based on BLOSUM62, PROVEAN, and PolyPhen-2 scores (Table 2). Polymorphisms A228S and D351N are both located within the serine/threonine protein kinase catalytic domain, whereas D77E is located outside this conserved domain (Fig. S2). Comparisons with homologous proteins from other grass species indicate that DV92 carries the ancestral allele at these three amino acid positions (Supplementary Fig. S2 and Table S8). The D351N polymorphism was detected only in G3116 and T. monococcum ssp. monococcum PI 355522. The D77E and A228S polymorphisms were present in the same accessions that carry the ELF3 haplotype-B (Supplementary Table S8).
The predicted PWWP1 proteins showed two indel polymorphisms and five amino acid changes between G3116 and DV92. These changes were all located outside the conserved PWWP domain (Supplementary Fig. S3). The indels were detected in variable regions of the protein, so it was not possible to determine the ancestral and derived states. Among the five amino acid changes observed in PWWP1, DV92 has the derived state for T294M and V701A, and G3116 has the derived state for S251W, S327I, and G542C (Supplementary Fig. S3). The derived amino acid polymorphisms in G3116 are predicted by BLOSUM62, PROVEAN, and PolyPhen-2 to have larger impact on protein function than those in DV92 (Table 2). The derived amino acids in PWWP1 found in G3116 were also present in other T. monococcum accessions carrying the ELF3 A- and B-haplotypes but not in those carrying the ELF3 haplotype-C. All the accession from this last group showed the V701A polymorphism (Supplementary Table S8).
Characterization of the expression profiles of Eps-A m 1 candidate genes
To test if the Eps-A m 1 phenotype was caused by differential expression of any of the candidate genes, transcript levels of ELF3, GRIK1-like, and PWWP1 were measured in T. monococcum Eps-A m 1-e and Eps-A m 1-l NILs grown under controlled environmental conditions (Fig. 3a–c). Samples were collected from leaves because preliminary studies showed significant differences in PPD1 and FT1 transcript levels between Eps-A m 1 alleles in this tissue. Although the leaf samples were collected from plants at the same chronological age (5 weeks after sowing), the shoot apical meristems were more advanced in the early flowering NILs (Eps-A m 1-e allele, terminal spikelet stage) than in the late flowering NILs (Eps-A m 1-l allele, double ridge stage). No significant differences in the transcription profiles of the three candidate genes were detected between Eps-A m 1 alleles during a 24-h time course (Fig. 3a–c).

Expression analysis of candidate genes (a–c), flowering genes (d–f), and circadian clock genes (g–l). T. monococcum NILs carrying the Eps-A m 1 allele from DV92 (Eps-A m 1-l, blue dashed lines) or G3116 (Eps-A m 1-e, red lines) were grown under controlled conditions (16 °C; 16 h light). Each point represents means ± standard error of the mean of six individual plants (*P < 0.05, **P < 0.01, ***P < 0.001)
Among these three genes, ELF3 showed transcript levels (Fig. 3a) that were almost an order of magnitude higher than those observed for GRIK1-like and PWWP1 (Fig. 3b, c). We also analyzed the expression profiles of the A-genome homoeologs of these three genes in hexaploid wheat using previously published RNA-seq data (IWGSC 2014) and WheatExp (Pearce et al. 2015). GRIK1-like showed relatively uniform transcript levels across different tissues and developmental stages (Supplementary FigS5a). PWWP1 showed low transcript levels across multiple tissues and developmental stages with a 4–8-fold increase in the last stages of grain development (Supplementary Fig. S5b). Finally, ELF3 showed similar expression levels across tissues and developmental stages, with a 2-fold increase in the latest stage of grain development (Z85, Supplementary Fig. S5c).
Validation of ELF3 as a candidate gene for Eps-A m 1
Studies in barley have shown that loss-of-function mutations in ELF3 accelerate the transition from vegetative to reproductive stages and the duration of spike development in this species (Faure et al. 2012). Since these two traits were also affected by the Eps-A m 1 alleles, we prioritized the validation of ELF3 as a candidate gene for Eps-A m 1. These validation studies included (i) the comparison of the expression of known downstream targets of ELF3, (ii) the determination of the effect of the introgression of the Eps-A m 1 allele in tetraploid wheat, and (iii) the determination of the effect of the elimination of all functional copies of ELF3 in tetraploid wheat.
-
(i)
Expression profiles of downstream genes: Previous studies of the barley elf3 mutants showed significant differences in the transcription profiles of PPD1, FT1, and central circadian clock genes, so the same genes were compared between the Eps-A m 1 alleles. The peak of the PPD1 expression in the NIL carrying the Eps-A m 1-l allele occurred 4 h earlier (Zeitgeber time ZT4) than in the NIL carrying the Eps-A m 1-e allele (ZT8), and those differences were highly significant (P < 0.01, Fig. 3d). The NILs carrying the Eps-A m 1-e allele also showed a higher peak than Eps-A m 1-l before dark, but those differences were not significant. The changes in PPD1 were also reflected in changes in the expression of FT1, a downstream target of PPD1. NILs carrying the Eps-A m 1-e showed significantly higher transcript levels of FT1 than NILs carrying the Eps-A m 1-l allele for most of the day (Fig. 3e). This result is consistent with the early flowering of the Eps-A m 1-e NIL.
Since ELF3 is involved in circadian clock regulation in both Arabidopsis and barley (Kolmos et al. 2011; Faure et al. 2012), we analyzed the expression patterns of several clock genes. Significant differences were detected in at least one time point in the PRR genes TOC1 (TIMING OF CAB EXPRESSION1) (Fig. 3h), PRR73 (Fig. 3j), and PRR59 (Fig. 3l). No significant differences between Eps-A m 1 alleles were detected for CCA1 (CIRCADIAN CLOCK ASSOCIATED 1) (Fig. 3g), GI (Fig. 3i), and PRR95 (Fig. 3k). In Arabidopsis, the evening complex, formed by the ELF3, ELF4, and LUX proteins, binds to the promoters of PIF4 and PIF5 to repress their transcription at dusk (Nusinow et al. 2011). We identified a T. monococcum PIF-like gene (TmoDV92v1_076750, Fox et al. 2014) that showed a significant 4-h shift between NILs carrying the different Eps-A m 1 alleles (Fig. 3f). As in PPD1, the peak of expression of PIF-like occurred 4 h earlier in the NILs carrying the Eps-A m 1-l allele (ZT8) than in those carrying the Eps-A m 1-e allele (ZT12) (Fig. 3f).
In summary, the set of flowering genes differentially expressed between the Eps-A m 1 alleles is consistent with an effect of ELF3. It should be noted that many of the changes observed in the T. monococcum NILs are not as severe as those reported for the elf3 null mutants in barley. This is not an unexpected result since the barley mutations are loss-of-function mutations (Faure et al. 2012; Zakhrabekova et al. 2012), whereas the T. monococcum ELF3 alleles differ only in four amino acid changes and are both likely functional alleles.
-
(ii)
Determination of the effect of the introgression of the Eps-A m 1 allele into tetraploid wheat: The GRIK1-like proteins encoded by the diploid T. monococcum DV92 and the tetraploid Kronos A-genome copy alleles are identical, but the ELF3 and PWWP1 proteins are not. Therefore, a significant effect of the introgression of the DV92 Eps-A m 1-l allele into tetraploid wheat would suggest that GRIK1-like is not likely a good candidate gene for Eps-A m 1.
Under controlled environmental conditions (16 °C, 16 h fluorescent light), BC5F3 lines of tetraploid wheat CBW0112 carrying a distal 1Am chromosome segment (Supplementary Fig. S4) including the DV92 Eps-A m 1-l allele flowered 6.4 days later (P < 0.0001; Fig. 4a) and had 1.1 more spikelets (P < 0.0001; Fig. 4b) than sister lines without the T. monococcum introgression. These results suggest that GRIK1-like is an unlikely candidate gene for Eps-A m 1. In addition, they suggest that the DV92 Eps-A m 1-l allele includes a gene(s) that actively delays flowering time and increases the number of spikelets per spike that is effective both in diploid and tetraploid wheat.
Fig. 4 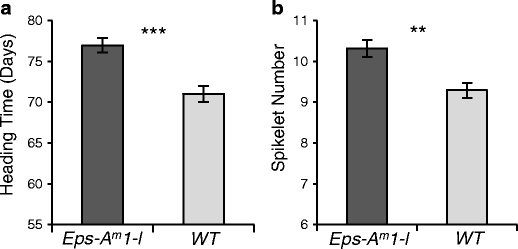
Effect of the introgression of the T. monococcum chromosome segment carrying the Eps-A m 1-l allele from DV92 into tetraploid wheat on heading time (a) and spikelet number (b). Bars represent the mean of at least 15 plants ± standard error of the mean. Asterisks indicate significant differences (**P < 0.01, ***P < 0.001)
-
(iii)
Loss-of-function mutants of ELF3 in tetraploid wheat: We hypothesized that if ELF3 was the correct candidate gene for Eps-A m 1, and the ELF3 allele from DV92 was actively delaying flowering and increasing the number of spikelets per spike, then an elf3-null mutant should show the opposite phenotypic effects. We selected mutations that generated premature stop codons in the A and B homoeologs of ELF3 in the TILLING population of the tetraploid wheat variety Kronos (Uauy et al. 2009). These mutations eliminate the last 241 (A-genome) or 244 (B-genome) amino acids of the C-terminal region of the ELF3 protein and are most likely loss-of-function alleles (Fig. 5a). These lines were intercrossed and then backcrossed to the recurrent parent Kronos for two generations, to reduce the number of background mutations. ELF3-WT and elf3-null homozygous BC2F2 plants (both homozygous for PPD-A1a) were evaluated under controlled environmental conditions (16 °C, 16 h fluorescent light). Under these conditions, the elf3-null Kronos plants flowered on average 7.2 days earlier than sister lines carrying wild-type ELF3 (P < 0.0001; Fig. 5b) and had on average 3.8 less spikelets per spike (P < 0.0001; Fig. 5c). This experiment confirmed that, in tetraploid wheat, the elf3-null mutants have the opposite effect to the introgression of the DV92 Eps-A m 1-l allele.
Fig. 5 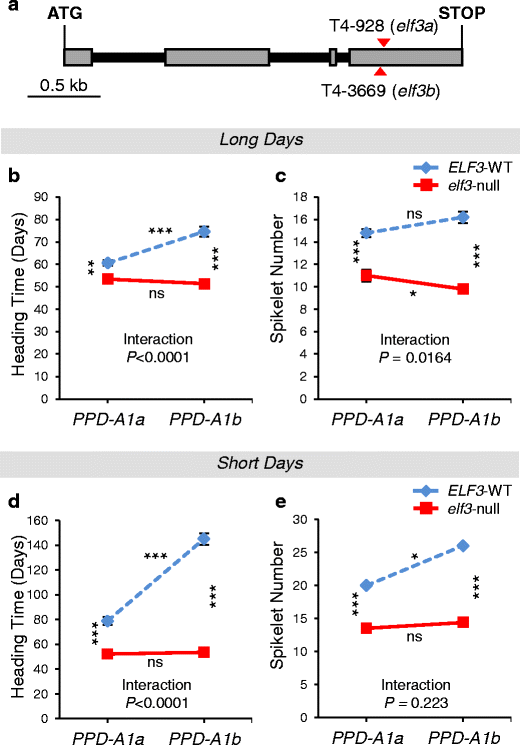
a Schematic representation of the ELF3 gene. Positions of the selected mutations introducing premature stop codons on the A- and B-genome copies of the gene are indicated with red triangles. Exons are shown as gray rectangles and introns as black lines. b–e Heading time and spikelet number of tetraploid BC2F3 lines carrying four possible homozygous combinations of ELF3 and PPD1 alleles. Plants were grown either under LD (b, c) or SD conditions (d, e). ELF3 alleles: ELF3-WT (blue dashed lines), elf3-null (red lines). PPD1 alleles: PPD-A1a (allele with reduced photoperiodic response); PPD-A1b (photoperiod sensitive allele). Asterisks between lines indicate significant differences between ELF3 alleles; asterisks on top and below lines indicate significant differences between PPD1 alleles (*P < 0.05, **P < 0.01, ***P < 0.001, ns nonsignificant)


Epistatic interactions between ELF3 and PPD1
In barley, loss-of-function mutations and wild-type alleles of ELF3 and PPD1 showed significant epistatic interactions (Faure et al. 2012). To test if similar epistatic interactions were present in tetraploid wheat, we intercrossed the elf3-null line carrying the PPD-A1a allele for reduced photoperiodic response with a Kronos near-isogenic line carrying the photoperiod-sensitive PPD-A1b allele. The four different combinations of ELF3 and PPD-A1 homozygous alleles were selected from segregating BC2F3 plants and were evaluated under both LD and SD conditions (16 °C, fluorescent light).
Under LD, highly significant differences (P < 0.0001) were found between elf3-null plants and ELF3-WT sister lines both in heading time and in the number of spikelets per spike (Fig. 5b, c). For both traits, the differences between ELF3-alleles were larger in the presence of the photoperiod-sensitive PPD-A1b allele than in the presence of the PPD-A1a allele with reduced photoperiodic response (23.2 vs. 7.2 days and 6.4 vs. 3.8 spikelets per spike, Fig. 5b, c). These differences resulted in significant interactions between ELF3 and PPD1 for both heading time (P < 0.0001) and number of spikelets per spike (P < 0.05). These interactions can be visualized as nonparallel lines in the graphs presented in Fig. 5b, c. The 14-day difference in heading time between the PPD-A1a and PPD-A1b alleles (P < 0.0001) in the presence of the wild-type ELF3 allele disappeared in the elf-null mutant background (Fig. 5b).
Under SD conditions, there was also a significant epistatic interaction for heading time between ELF3 and PPD1 alleles (P < 0.0001) but not for spikelet number (P = 0.2230, Fig. 5d, e). In general, the effects of the PPD1 and ELF3 alleles were stronger under SD than under LD. In the presence of the wild-type allele of ELF3, plants carrying the photoperiod-sensitive PPD-A1b allele flowered on average more than 66 days later than the PPD-A1a sister lines (Fig. 5d). When the experiment was terminated 150 days after sowing, there were still three plants with this genotype that did not flower and that were excluded from the statistical analysis. By contrast, in the presence of the elf3-null mutant alleles, no significant differences were observed between plants carrying the PPD-A1a (52.2 days) and PPD-A1b (53.6 days) alleles (Fig. 5d). Heading times for elf3-null lines under SD were comparable to those observed under LD conditions (51.4 to 53.6 days).
In summary, loss-of-function mutations in elf3 were sufficient to abolish the differences in flowering time associated with the presence of the PPD-A1a or PPD-A1b alleles, or with different photoperiods.
Discussion
Characterization of the Eps-A m 1 candidate region
T. monococcum is a different species from T. urartu, the A-genome donor to the polyploid wheat species (Johnson and Dhaliwal 1976). These two diploid species diverged from each other before the origin of the polyploid wheat species (Dubcovsky and Dvorak 2007), so domestication and selection processes operated independently on their respective genomes. Therefore, it is not surprising that the cloning of several important agronomic genes from T. monococcum has revealed alleles not detected before in the polyploid wheat species. These include novel alleles for spring growth habit in the vernalization genes VRN1 and VRN2 (Yan et al. 2003, 2004); novel stem rust resistance genes, such as Sr35 (Saintenac et al. 2013); and new earliness per se alleles, such as those detected in Eps-A m 3 (Gawronski et al. 2014), and in Eps-A m 1 in this study.
Map-based cloning approaches are easier to perform in T. monococcum than in the polyploid wheat species because of its diploid nature and the relatively higher levels of polymorphism (Dubcovsky et al. 1996). In this study, we used a highly polymorphic segregating population from a cross between a cultivated (DV92) and a wild T. monococcum accession (G3116) to map Eps-A m 1 within a 0.1-cM interval in the distal region of T. monococcum chromosome arm 1AmL (Fig. 2). A large deletion in the colinear region of chromosome arm 1DL was also associated with differences in earliness per se in hexaploid wheat (Zikhali et al. 2015). In barley, 85 out of the 195 early flowering mutants characterized by the Scandinavian barley mutation program (Lundqvist 2014) belong to the mat-a group (=eam8), that was mapped on the distal region of chromosome arm 1HL colinear with Eps-A m 1 (Zakhrabekova et al. 2012; Faure et al. 2012). Taken together, these results indicate the presence of a conserved earliness per se gene in the distal region of the long arm of homoeologous group one in both barley and wheat.
The sequencing of the Eps-A m 1 candidate region revealed 10 T. monococcum genes, including nine that were present in the colinear region in B. distachyon (Fig. 2). The only other B. distachyon gene present in this region was homologous to a wheat gene located in a different chromosome region, suggesting that the current list of T. monococcum candidate genes includes all the conserved genes present in this region. However, we cannot rule out the possibility of noncolinear wheat genes in the two unsequenced gaps of the current T. monococcum physical map. Among the 10 T. monococcum genes detected in this region, three were completely linked to Eps-A m 1: GRIK1-like, PWWP1, and ELF3 (Fig. 2). These three candidate genes are described in more detail below.
Eps-A m 1 candidate genes
None of the three linked genes showed significant transcriptional differences between the T. monococcum NILs carrying the Eps-A m 1-l and Eps-A m 1-e alleles (Fig. 3a–c), suggesting that the differences in heading time and spikelet number between these alleles are more likely associated to the observed differences in their respective proteins. Therefore, we present below a detailed discussion of the different amino acid polymorphisms identified in this study.
GRIK1-like
The putative serine/threonine kinase encoded by this gene has been proposed to be part of a gene network that coordinates the metabolic requirements of rapidly growing cells (Shen et al. 2009). For all three amino acid polymorphism identified in GRIK1-like, the cultivated T. monococcum accession DV92 carries the ancestral haplotype. Since the predicted GRIK1-like protein in DV92 is identical to the one predicted for the A-genome of tetraploid wheat (Supplementary Fig. S2 and Table S8c), the significant effect of the Eps-A m 1-l introgression into tetraploid wheat suggests that GRIK1-like is an unlikely candidate gene for Eps-A m 1.
PWWP1
Not much is known about the PWWP1 protein except for the presence of the PWWP domain (pfam 00855). This domain is found in numerous proteins involved in cell division, growth, and differentiation. It binds to Histone-4 methylated at lysine-20, H4K20me, suggesting that it is a methyl-lysine recognition motif. The PWWP1 proteins encoded by the DV92 and G3116 alleles differ in five amino acids. The S251W and T294M polymorphisms are unlikely causal mutations for Eps-A m 1 because they are not polymorphic in the DV92 × PI 355522 population, which shows a clear segregation for heading time and spikelet number (Supplementary Table S9). The S327I and G542C polymorphisms are also unlikely causal mutations for Eps-A m 1 because these two amino acids are identical between DV92 and the A-genome of tetraploid wheat, and still the introgression of the DV92 Eps-A m 1allele into tetraploid wheat is associated with significant differences in heading time and number of spikelets per spike (Fig. 4). We currently have no evidence to rule out V701A as a candidate polymorphism for Eps-A m 1. However, this change is predicted to have a limited effect on protein structure and function (Table 2) and, therefore, is an unlikely causal polymorphism for the observed phenotypic differences.
ELF3
In Arabidopsis, ELF3 mediates the circadian gating of light responses and regulates light input to the clock (McWatters et al. 2000; Covington et al. 2001). In this species, ELF3, ELF4, and LUX form a trimeric protein complex named evening complex that directly represses circadian clock genes PRR7 and PRR9 (Dixon et al. 2011; Helfer et al. 2011; Kolmos et al. 2011; Chow et al. 2012; Herrero et al. 2012). The evening complex regulates the expression of growth-promoting transcription factors PIF4 and PIF5, gating hypocotyl growth in the early evening (Nusinow et al. 2011). ELF3 also affects the evening loop of the circadian clock by regulating GI protein turnover (Yu et al. 2008). Mutations in ELF3 have multiple pleiotropic effects. For example, the natural A362V mutation in Arabidopsis results in a constitutive shade avoidance phenotype and shortened circadian periods (Coluccio et al. 2011) and affects temperature-induced hypocotyl elongation (Raschke et al. 2015). Loss-of-function mutations of elf3 result in even stronger phenotypes including arrhythmic circadian outputs under continuous light and dark and early flowering under both SD and LD photoperiods (Covington et al. 2001; Thines and Harmon 2010; Zagotta et al. 1996). Arabidopsis plants overexpressing ELF3 show delayed flowering under LD, supporting the hypothesis that this gene acts as a flowering repressor (Liu et al. 2001).
A role of ELF3 in the regulation of flowering is also well established in barley and supported by indirect evidence in wheat. The characterization of 87 early flowering mat-a mutants in barley resulted in the identification of more than 20 independent ELF3 alleles encoding for defective proteins. These results provided convincing evidence that the early flowering of the barley eam8/mat-a mutants is caused by mutations in ELF3 (Faure et al. 2012; Zakhrabekova et al. 2012). In hexaploid wheat, the large deletion in the distal region of the 1DL chromosome arm associated with accelerated flowering (under both SD and LD) was shown to include the ELF-D3 gene. In addition, a S674G polymorphism in ELF-B3 was linked to a QTL for heading time in the double haploid population of Avalon × Cadenza (Zikhali et al. 2015). These results, together with the complete linkage between Eps-A m 1 and ELF3 in the T. monococcum high-density mapping population described in this study, provide strong support to the hypothesis that ELF3 is Eps-A m 1.
The barley elf3-null mutants showed an accelerated transition from vegetative to reproductive growth and accelerated spike development (Faure et al. 2012). These effects are opposite to those associated with the presence of the T. monococcum Eps-A m 1-l allele. The T. monococcum plants carrying the Eps-A m 1-l allele exhibit a delayed transition from vegetative to reproductive growth and delayed spike development that results in a significant increase in the number of spikelets per spike, both in diploid (Lewis et al. 2008) and tetraploid wheat (Fig. 4). These phenotypic differences are modulated by temperature, with stronger effects observed at 16 than at 23 °C (Lewis et al. 2008). The modulation of Eps-A m 1 effects by temperature provides an additional link between Eps-A m 1 and ELF3 because ELF3 plays a central role in the thermal entrainment of the clock in Arabidopsis, and the elf3 mutant shows no evidence of temperature entrainment of the circadian clock in the dark (Thines and Harmon, 2010). In addition, a single amino acid change in the ELF3 protein was linked to variation in thermoresponsive growth in Arabidopsis through the differential regulation of PIF4 expression and its downstream targets (Raschke et al. 2015). A complementary study showed that the binding of ELF3 to the target promoters is temperature dependent, providing a mechanism by which temperature directly controls ELF3 activity (Box et al. 2015).
Epistatic interactions between ELF3 and PPD1
PPD1 is the major photoperiod gene in wheat (Beales et al. 2007; Wilhelm et al. 2009) and barely (Turner et al. 2005). However, the natural mutations that originated the photoperiod-insensitive allele in barley are different from the mutations that originated the reduced photoperiod-sensitive alleles in wheat. In barley, the photoperiod-insensitive ppd-H1 carries four amino acid changes, including one in a conserved amino acid of the CCT domain, and that is the most likely causal basis of the ppd-H1 phenotype (Turner et al. 2005). The ppd-H1 allele is unable to accelerate flowering under long days but shows no differences with PPD-H1 under SD. By contrast, the reduced photoperiod sensitivity mutations in wheat are associated with deletions in the promoter region that result in elevated expression of the PPD1 gene and its downstream FT1 target and accelerated flowering under SD (Beales et al. 2007; Wilhelm et al. 2009). Given the different nature of these PPD1 mutations, it is interesting to compare the epistatic interactions between ELF3 and PPD1 in both species.
Significant epistatic interactions between ELF3 and PPD1 were detected both under LD and SD in wheat, but only under LD in barley (Supplementary Table S11, Faure et al. 2012). The similar effects of the barley ppd-H1 and PPD-H1 alleles under SD likely explain the absence of significant epistatic interactions under these conditions. By contrast, in wheat, both PPD1 alleles have significant effects under LD and SD, although the effect under LD is largely reduced. In this study, when plants carrying the wild-type ELF3 allele were grown under fluorescent lamps at 16 °C, the differences between the PPD-A1a and PPD-A1b alleles were 14 days, compared with 66.3 days under SD (Supplementary Table S11). However, in previous studies using stronger sodium halide lights (~260 μM m−2 s−1) and higher temperatures (day 20 °C/night 17 °C), the differences between the same PPD1 alleles in the same Kronos background were only 4 days under LD (Chen et al. 2014). These results suggest that the light and temperature conditions selected for this study enhanced the differences between the PPD1 alleles and likely the phenotypic differences between the ELF3 alleles.
That an important part of the ELF3 effect on flowering is mediated by its effect on PPD1 is evident in the elf3 mutants in rice. In rice, SD accelerates flowering (short-day plant) and PPD1 (=PRR37) acts as a repressor of FT-like genes (Zhao et al. 2012; Koo et al. 2013) rather than as a promoter of flowering as observed in the long-day cereals wheat and barley. Loss-of-function mutations of elf3 in rice result in late flowering time under both SD and LD conditions (Matsubara et al. 2012; Saito et al. 2012), opposite to the effect observed in barley and wheat. This reversion can be explained in part by the opposite effect of PPD1/PRR37 in these two species, although the upregulation of the LD floral repressor GHD7 in the rice elf3 mutant also contributes to its late flowering. Interestingly, an amino acid change in the conserved block III of the rice ELF3 protein accelerates flowering under natural day and LD conditions, without affecting circadian rhythms (Matsubara et al. 2012). Since the evening complex interacts with multiple proteins (Huang et al. 2016), it would be interesting to test the effect of the ELF3 natural mutations discovered in rice and in T. monococcum accession DV92 on these protein-protein interactions.
Although the interaction between ELF3 and PPD1 plays an important role in the regulation of flowering, there seems to be an additional PPD1-independent effect of ELF3 on flowering time. In barley, even in the presence of the photoperiod-insensitive ppd-H1 allele, mutations in ELF3 result in a significant acceleration of flowering under both LD (13.3 days) and SD (25.3 days) (Supplementary Table S11, Faure et al. 2012). However, we cannot rule out the possibility that the ppd-H1 mutations is hypomorphic and has sufficient residual effect when expressed at the increased level observed in the eam8/mat-a mutants (Faure et al. 2012).
Applications and practical implications of the novel ELF3 alleles
The significant increase in spikelet number associated with the Eps-A m 1-l allele (ELF3 haplotype-C) may have favored the selection of this allele during the domestication of T. monococcum, explaining its high frequency among the cultivated T. monococcum ssp. monococcum accessions (Supplementary Table S8). However, since T. monococcum is currently cultivated in very limited regions of the world (Troccoli and Codianni 2005), we decided to transfer the Eps-A m 1-l allele into tetraploid wheat. The significant increase in number of spikelets per spike (1.1 more spikelets) represents an encouraging result. However, this preliminary experiment was performed under controlled environmental conditions that are different from the ones observed in natural environments. The potential impact of this introgression on tetraploid wheat yield will require replicated field experiments in multiple environments. We are currently introgressing this T. monococcum chromosome segment into different tetraploid and hexaploid backgrounds to perform these experiments.
Although the results from the Eps-A m 1-l allele introgression into tetraploid wheat are very preliminary for practical application, this experiment provided some valuable information. In previous studies using diploid wheat segregating populations, it was not possible to determine which of the Eps-A m 1 alleles represented the ancestral stage. We did not know if the Eps-A m 1-l allele delayed flowering time or if the Eps-A m 1-e allele accelerated the transition to the reproductive stage (Lewis et al. 2008). We speculated that the Eps-A m 1-l allele was the derived state based on its high frequency among the more modern cultivated accessions of T. monococcum. The significant delay in heading time and increased spikelet number observed in the tetraploid lines carrying the T. monococcum DV92 allele support this hypothesis. It also suggests that the Eps-A m 1-l has a stronger effect than the replaced Eps-A1 allele from tetraploid wheat and that Eps-A m 1-l is a hyperactive allele.
However, the results from tetraploid wheat do not provide conclusive evidence that the effects were caused by the polymorphisms in ELF3, because a large number of T. monococcum genes were introgressed together with ELF3 into the translocated 1AmL chromosome segment. To provide a more conclusive answer to this question, we are mutagenizing the tetraploid line with the T. monococcum introgression with EMS. If we are able to identify loss-of-function mutations in the introgressed ELF3, and these mutations abolish the flowering delay and spikelet number increase associated with the translocated 1AmL chromosome segment, we will be able to conclude that the observed differences were caused by ELF3.
In addition to the T. monococcum introgression line with delayed flowering, the early flowering elf3 tetraploid mutants generated in this study may also represent a valuable germplasm for environments that require a very short life cycle and that are exposed to very long days during their growing season. In barley, the elf3 mutations (eam8/mat-a) have been used commercially in Scandinavia, in regions with very short growing seasons and long photoperiods (Lundqvist 2014). The Swedish cultivar ‘Mari’ carrying the elf3 mutation has been grown as far north as Iceland (Lundqvist 2014). Faure et al. (2012) proposed the intriguing hypothesis that the disruption of the circadian clock observed in the elf3 mutants may be adaptative in regions that exhibit extreme variation in light period during the year (e.g., >20 h of light per day). Although the current production of wheat at high latitudes is limited, global warming may open new geographic areas to wheat cultivation. In this context, it would be interesting to test the performance of elf3 wheat mutants in high-latitude regions of North America, Europe, and Asia.
Even if the elf3 loss-of-function wheat mutants find a special niche in high-latitude regions, it is unlikely that they will be useful in other regions because of the negative pleiotropic effects in the number of spikelets per spike. In our controlled environment experiments, the elf3 Kronos mutants showed significant reductions in the number of spikelets per spike both under both LD (up to 6.4 spikelets) and SD photoperiods (up to 11.6 spikelets, Supplementary Table S1). By contrast, the Eps-A m 1-l allele is associated with a positive effect on spikelet number and was selected in regions where commercial polyploid wheat species are being currently grown. This may increase the probability of this new ELF3 allele of being adaptative in current wheat production regions. Even if this novel hyperactive ELF3 allele is useful, it still remains to be tested if the other linked T. monococcum genes present in the translocated chromosome segments are associated to negative effects on agronomic performance or quality.
Control of flowering time is an important feature of plant adaptation, and the transition between the vegetative and reproductive growth needs to occur in a precise seasonal window to maximize wheat grain yield potential. The new ELF3 alleles introduced in this study expand the genetic tools available to wheat breeders to manipulate wheat flowering time and maximize its adaptation to novel and changing environments.
References
Adzhubei IA, Schmidt S, Peshkin L, Ramensky VE, Gerasimova A, Bork P, Kondrashov AS, Sunyaev SR (2010) A method and server for predicting damaging missense mutations. Nat Methods 7:248–249
Aguilera A, Gómez-González B (2008) Genome instability: a mechanistic view of its causes and consequences. Nat Rev Genet 9:204–217
Appendino ML, Slafer GA (2003) Earliness per se and its dependence upon temperature in diploid wheat lines differing in the major gene Eps-A m 1 alleles. J Agr Sci 141:149–154
Beales J, Turner A, Griffiths S, Snape JW, Laurie DA (2007) A Pseudo-Response Regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor Appl Genet 115:721–733
Box MS, Huang BE, Domijan M, Jaeger KE, Khattak AK, Yoo SJ, Sedivy EL, Jones DM, Hearn TJ, Webb AA, Grant A (2015) ELF3 controls thermoresponsive growth in Arabidopsis. Curr Biol 25:194–199
Bullrich L, Appendino ML, Tranquilli G, Lewis S, Dubcovsky J (2002) Mapping of a thermos-sensitive earliness per se gene on Triticum monococcum. Theor Appl Genet 105:585–593
Campoli C, Pankin A, Drosse B, Casao CM, Davis SJ, von Korff M (2013) HvLUX1 is a candidate gene underlying the early maturity 10 locus in barley: phylogeny, diversity, and interactions with the circadian clock and photoperiodic pathways. New Phytol 199:1045–1059
Chen A, Dubcovsky J (2012) Wheat TILLING mutants show that the vernalization gene VRN1 down-regulates the flowering repressor VRN2 in leaves but is not essential for flowering. PLoS Genet 8:e1003134
Chen A, Li C, Hu W, Lau MY, Lin H, Rockwell NC, Martin SS, Jernstedt JA, Lagarias JC, Dubcovsky J (2014) PHYTOCHROME C plays a major role in the acceleration of wheat flowering under long-day photoperiod. Proc Natl Acad Sci U S A 111:10037–10044
Choi Y, Sims GE, Murphy S, Miller JR, Chan AP (2012) Predicting the functional effect of amino acid substitutions and indels. PLoS ONE 7:e46688
Choulet F, Wicker T, Rustenholz C, Paux E, Salse J, Leroy P, Schlub S, Le Paslier MC, Magdelenat G, Gonthier C, Couloux A, Budak H, Breen J, Pumphrey M, Liu SX, Kong XY, Jia JZ, Gut M, Brunel D, Anderson JA, Gill BS, Appels R, Keller B, Feuillet C (2010) Megabase level sequencing reveals contrasted organization and evolution patterns of the wheat gene and transposable element spaces. Plant Cell 22:1686–1701
Chow BY, Helfer A, Nusinow DA, Kay SA (2012) ELF3 recruitment to the PRR9 promoter requires other evening complex members in the Arabidopsis circadian clock. Plant Signal Behav 7:170–173
Coluccio MP, Sanchez SE, Kasulin L, Yanovsky MJ, Botto JF (2011) Genetic mapping of natural variation in a shade avoidance response: ELF3 is the candidate gene for a QTL in hypocotyl growth regulation. J Exp Bot 62:167–176
Comadran J, Kilian B, Russell J, Ramsay L, Stein N, Ganal M, Shaw P, Bayer M, Thomas W, Marshall D, Hedley P, Tondelli A, Pecchioni N, Francia E, Korzun V, Walther A, Waugh R (2012) Natural variation in a homolog of Antirrhinum CENTRORADIALIS contributed to spring growth habit and environmental adaptation in cultivated barley. Nat Genet 44:1388–1392
Corbesier L, Vincent C, Jang SH, Fornara F, Fan QZ, Searle I, Giakountis A, Farrona S, Gissot L, Turnbull C, Coupland G (2007) FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 316:1030–1033
Covington MF, Panda S, Liu XL, Strayer CA, Wagner DR, Kay SA (2001) ELF3 modulates resetting of the circadian clock in Arabidopsis. Plant Cell 13:1305–1316
Distelfeld A, Li C, Dubcovsky J (2009) Regulation of flowering in temperate cereals. Curr Opin Plant Biol 12:178–184
Dixon LE, Knox K, Kozma-Bognar L, Southern MM, Pokhilko A, Millar AJ (2011) Temporal repression of core circadian genes is mediated through EARLY FLOWERING 3 in Arabidopsis. Curr Biol 21:120–5
Dubcovsky J, Dvorak J (2007) Genome plasticity a key factor in the success of polyploid wheat under domestication. Science 316:1862–1866
Dubcovsky J, Luo M-C, Zhong G-Y, Bransteiter R, Desai A, Kilian A, Kleinhofs A, Dvorak J (1996) Genetic map of diploid wheat, Triticum monococcum L., and its comparison with maps of Hordeum vulgare L. Genetics 143:983–999
Faricelli ME, Valárik M, Dubcovsky J (2010) Control of flowering time and spike development in cereals: the earliness per se Eps-1 region in wheat, rice, and Brachypodium. Funct Integr Genomics 10:293–306
Faure S, Turner AS, Gruszka D, Christodoulou V, Davis SJ, von Korff M, Laurie DA (2012) Mutation at the circadian clock gene EARLY MATURITY 8 adapts domesticated barley (Hordeum vulgare) to short growing seasons. Proc Natl Acad Sci U S A 109:8328–8333
Flood RG, Halloran GM (1984) Basic development rate in spring wheat. Agron J 76:260–264
Fox SE, Geniza M, Hanumappa M, Naithani S, Sullivan C, Preece J, Tiwari VK, Elser J, Leonard JM, Sage A, Gresham C (2014) De novo transcriptome assembly and analyses of gene expression during photomorphogenesis in diploid wheat Triticum monococcum. PLoS One 9:e96855
Fu D, Szucs P, Yan L, Helguera M, Skinner J, von Zitzewitz J, Hayes PM, Dubcovsky J (2005) Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol Genet Genomics 273:54–65
Gawronski P, Ariyadasa R, Himmelbach A, Poursarebani N, Kilian B, Stein N, Steuernagel B, Hensel G, Kumlehn J, Sehgal SK, Gill BS, Gould P, Hall A, Schnurbusch T (2014) A distorted circadian clock causes early flowering and temperature-dependent variation in spike development in the Eps-3A m mutant of einkorn wheat. Genetics 196:1253–1261
Gawronski P, Schnurbusch T (2012) High‐density mapping of the earliness per se-3Am (Eps‐3A m) locus in diploid einkorn wheat and its relation to the syntenic regions in rice and Brachypodium distachyon L. Mol Breed 30:1097–1108
Gottlieb A, Muller HG, Massa AN, Wanjugi H, Deal KR, You FM, Xu XY, Gu YQ, Luo MC, Anderson OD, Chan AP, Rabinowicz P, Devos KM, Dvorak J (2013) Insular organization of gene space in grass genomes. PLoS One 8:e54101
Griffiths S, Simmonds J, LeveringtonM WY, Fish L, Sayers L, Alibert L, Orford S, Wingen L, Herry L, Faure S, Laurie D, Bilham L, Snape J (2009) Meta-QTL analysis of the genetic control of ear emergence in elite European winter wheat germplasm. Theor Appl Genet 119:383–395
Helfer A, Nusinow DA, Chow BY, Gehrke AR, Bulyk ML, Kay SA (2011) LUX ARRHYTHMO encodes a nighttime repressor of circadian gene expression in the Arabidopsis core clock. Curr Biol 21:126–33
Henikoff S, Henikoff JG (1992) Amino acid substitution matrices from protein blocks. Proc Natl Acad Sci U S A 89:10915–10919
Herrero E, Kolmos E, Bujdoso N, Yuan Y, Wang M, Berns MC, Uhlworm H, Coupland G, Saini R, Jaskolski M, Webb A, Gonçalves J, Davis SJ (2012) EARLY FLOWERING4 recruitment of EARLY FLOWERING3 in the nucleus sustains the Arabidopsis circadian clock. Plant Cell 24:428–443
Heun M, Schaferpregl R, Klawan D, Castagna R, Accerbi M, Borghi B, Salamini F (1997) Site of einkorn wheat domestication identified by DNA fingerprinting. Science 278:1312–1314
Hicks KA, Albertson TM, Wagner DR (2001) EARLY FLOWERING 3 encodes a novel protein that regulates circadian clock function and flowering in Arabidopsis. Plant Cell 13:1281–1292
Hoogendoorn J (1985) A reciprocal F1 analysis of the genetic control of ear emergence, number of leaves and number of spikelets in wheat. Euphytica 34:545–555
Huang H, Alvarez S, Bindbeutel R, Shen Z, Naldrett MJ, Evans BS, Briggs SP, Hicks LM, Kay SA, Nusinow DA (2016) Identification of evening complex associated proteins in Arabidopsis by affinity purification and mass spectrometry. Mol Cell Proteomics 1:201–217
IWGSC The International Wheat Genome Sequencing Consortium (2014) A chromosome-based draft sequence of hexaploid bread wheat (Triticum aestivum) genome. Science 345:1251788
Johnson BL, Dhaliwal HS (1976) Reproductive isolation of Triticum boeticum and Triticum urartu and the origin of the tetraploid wheats. Am J Bot 63:1088–1094
Kippes N, Zhu J, Chen A, Vanzetti L, Lukaszewski A, Nishida H, Kato K, Dvorak J, Dubcovsky J (2014) Fine mapping and epistatic interactions of the vernalization gene VRN-D4 in hexaploid wheat. Mol Genet Genomics 289:47–62
Kolmos E, Herrero E, Bujdoso N, Millar AJ, Tóth R, Gyula P, Nagy F, Davis SJ (2011) A reduced-function allele reveals that EARLY FLOWERING3 repressive action on the circadian clock is modulated by phytochrome signals in Arabidopsis. Plant Cell 23:3230–3246
Koo BH, Yoo SC, Park JW, Kwon CT, Lee BD, An G, Zhang Z, Li J, Li Z, Paek NC (2013) Natural variation in OsPRR37 regulates heading date and contributes to rice cultivation at a wide range of latitudes. Mol Plant 6:1877–1888
Krasileva KV, Buffalo V, Bailey P, Pearce S, Ayling S, Tabbita F, Soria M, Wang S, Consortium I, Akhunov E, Uauy C, Dubcovsky J (2013) Separating homeologs by phasing in the tetraploid wheat transcriptome. Genome Biol 14:R66
Laurie DA, Pratchett N, Bezant J, Snape JW (1995) RFLP mapping of five major genes and eight quantitative trait loci controlling flowering time in a winter x spring barley (Hordeum vulgare L.) cross. Genome 38:575–585
Leroy P, Guilhot N, Sakai H, Bernard A, Choulet F, Theil S, Reboux S, Amano N, Flutre T, Pelegrin C, Ohyanagi H, Seidel M, Giacomoni F, Reichstadt M, Alaux M, Gicquello E, Legeai F, Cerutti L, Numa H, Tanaka T, Mayer K, Itoh T, Quesneville H, Feuillet C (2011) TriAnnot: a versatile and high performance pipeline for the automated annotation of plant genomes. Front Plant Sci 3:1–14
Lewis S, Faricelli ME, Appendino ML, Valárik M, Dubcovsky J (2008) The chromosome region including the earliness per se locus Eps-A m 1 affects the duration of early developmental phases and spikelet number in diploid wheat. J Exp Bot 59:3595–3607
Li C, Lin H, Dubcovsky J (2015) Factorial combinations of protein interactions generate a multiplicity of florigen activation complexes in wheat and barley. Plant J 84:70–82
Liu XL, Covington MF, Fankhauser C, Chory J, Wagner DR (2001) ELF3 encodes a circadian clock-regulated nuclear protein that functions in an Arabidopsis PHYB signal transduction pathway. Plant Cell 13:1293–1304
Lijavetzky D, Muzzi G, Wicker T, Keller B, Wing R, Dubcovsky J (1999) Construction and characterization of a bacterial artificial chromosome (BAC) library for the A genome of wheat. Genome 42:1176–1182
Lundqvist U (2014) Scandinavian mutation research in barley—a historical review. Hereditas 151:123–131
Matsubara K, Ogiso-Tanaka E, Hori K, Ebana K, Ando T, Yano M (2012) Natural variation in Hd17, a homolog of Arabidopsis ELF3 that is involved in rice photoperiodic flowering. Plant Cell Physiol 53:709–716
McWatters HG, Bastow RM, Hall A, Millar AJ (2000) The ELF3 zeitnehmer regulates light signaling to the circadian clock. Nature 408:716–720
Miura H, Worland AJ (1994) Genetic control of vernalization, day length response and earliness per se by homoeologous group-3 chromosomes in wheat. Plant Breed 113:160–169
Mizuno N, Nitta M, Sato K, Nasuda S (2012) A wheat homologue of PHYTOCLOCK 1 is a candidate gene conferring the early heading phenotype to einkorn wheat. Genes Genet Syst 87:357–367
Nusinow DA, Helfer A, Hamilton EE, King JJ, Imaizumi T, Schultz TF, Farré EM, Kay SA (2011) The ELF4-ELF3-LUX complex links the circadian clock to diurnal control of hypocotyl growth. Nature 75:398–402
Pearce S, Vanzetti LS, Dubcovsky J (2013) Exogenous gibberellins induce wheat spike development under short days only in the presence of VERNALIZATION 1. Plant Physiol 163:1433–1445
Pearce S, Vazquez-Gross H, Herin SY, Hane D, Wang Y, Gu YQ, Dubcovsky J (2015) WheatExp: an RNA-seq expression database for polyploid wheat. BMC Plant Biol 15:299
Raschke A, Ibañez C, Ullrich KK, Anwer MU, Becker S, Glöckner A, Trenner J, Denk K, Saal B, Sun X, Ni M, Davis SJ, Delker C, Quint M (2015) Natural variants of ELF3 affect thermomorphogenesis by transcriptionally modulating PIF4-dependent auxin response genes. BMC Plant Biol 15:197
Saintenac C, Zhang W, Salcedo A, Rouse MN, Trick HN, Akhunov E, Dubcovsky J (2013) Identification of wheat gene Sr35 that confers resistance to Ug99 stem rust race group. Science 341:783–786
Saito H, Ogiso-Tanaka E, Okumoto Y, Yoshitake Y, Izumi H, Yokoo T, Matsubara K, Hori K, Yano M, Inoue H, Tanisaka T (2012) Ef7 encodes an ELF3-like protein and promotes rice flowering by negatively regulating the floral repressor gene Ghd7 under both short- and long-day conditions. Plant Cell Physiol 53:717–728
Scarth R, Law CN (1984) The control of the day-length response in wheat by the Group-2 chromosomes. Plant Breed 92:140–150
Shen W, Reyes MI, Hanley-Bowdoin L (2009) Arabidopsis protein kinases GRIK1 and GRIK2 specifically activate SnRK1 by phosphorylating its activation loop. Plant Physiol 150:996–1005
Slafer GA (1996) Differences in phasic development rate amongst wheat cultivars independent of responses to photoperiod and vernalization. A viewpoint of the intrinsic earliness hypothesis. J Agr Sci 126:403–419
Slafer GA, Rawson HM (1994) Sensitivity of wheat phasic development to major environmental factors: a re-examination of some assumptions made by physiologists and modellers. Aust J Plant Physiol 21:393–426
Slafer GA, Rawson HM (1995a) Base and optimum temperatures vary with genotype and stage of development in wheat. Plant Cell Environ 18:671–679
Slafer GA, Rawson HM (1995b) Intrinsic earliness and basic development rate assessed for their response to temperature in wheat. Euphytica 83:175–183
Tamaki S, Matsuo S, Wong HL, Yokoi S, Shimamoto K (2007) Hd3a protein is a mobile flowering signal in rice. Science 316:1033–1036
Thines B, Harmon FG (2010) Ambient temperature response establishes ELF3 as a required component of the core Arabidopsis circadian clock. Proc Natl Acad Sci U S A 107:3257–3262
Troccoli A, Codianni P (2005) Appropriate seeding rate for einkorn, emmer, and spelt grown under rainfed condition in southern Italy. Eur J Agron 22:293–300
Turner A, Beales J, Faure S, Dunford RP, Laurie DA (2005) The pseudoresponse regulator Ppd-H1 provides adaptation to photoperiod in barley. Science 310:1031–1034
Uauy C, Paraiso F, Colasuonno P, Tran RK, Tsai H, Berardi S, Comai L, Dubcovsky J (2009) A modified TILLING approach to detect induced mutations in tetraploid and hexaploid wheat. BMC Plant Biol 9:115
Valarik M, Linkiewicz AM, Dubcovsky J (2006) A microcolinearity study at the earliness per se gene Eps-A m 1 region reveals an ancient duplication that preceded the wheat-rice divergence. Theor Appl Genet 112:945–957
Weller JL, Liew LC, Hecht VF, Rajandran V, Laurie RE, Ridge S, Wenden B, Vander Schoor JK, Jaminon O, Blassiau C, Dalmais M, Rameauc C, Bendahmanef A, Macknightb RC, Lejeune-Hénautd I (2012) A conserved molecular basis for photoperiod adaptation in two temperate legumes. Proc Natl Acad Sci U S A 109:21158–21163
Wilhelm EP, Turner AS, Laurie DA (2009) Photoperiod insensitive Ppd-A1a mutations in tetraploid wheat (Triticum turgidum Desf.). Theor Appl Genet 118:285–294
Worland AJ (1996) The influence of flowering time genes on environmental adaptability in European wheats. Euphytica 89:49–57
Worland AJ, Law CN (1986) Genetic analysis of chromosome 2D of wheat. I. The location of genes affecting height, day-length insensitivity, hybrid dwarfism and yellow-rust resistance. Plant Breed 96:331–345
Yan L, Fu D, Li C, Blechl A, Tranquilli G, Bonafede M, Sanchez A, Valarik M, Yasuda S, Dubcovsky J (2006) The wheat and barley vernalization gene VRN3 is an ortholog of FT. Proc Natl Acad Sci U S A 103:19581–19586
Yan L, Loukoianov A, Blechl A, Tranquilli G, Ramakrishna W, SanMiguel P, Bennetzen JL, Echenique V, Dubcovsky J (2004) The wheat VRN2 gene is a flowering repressor down-regulated by vernalization. Science 303:1640–1644
Yan L, Loukoianov A, Tranquilli G, Helguera M, Fahima T, Dubcovsky J (2003) Positional cloning of wheat vernalization gene VRN1. Proc Natl Acad Sci U S A 100:6263–6268
Yu JW, Rubio V, Lee NY, Bai S, Lee SY, Kim SS, Liu L, Zhang Y, Irigoyen ML, Sullivan JA, Zhang Y, Lee I, Xie Q, Paek NC, Deng XW (2008) COP1 and ELF3 control circadian function and photoperiodic flowering by regulating GI stability. Mol Cell 32:617–30
Zagotta MT, Hicks KA, Jacobs CI, Young JC, Hangarter RP, Meeks-Wagner DR (1996) The Arabidopsis ELF3 gene regulates vegetative photomorphogenesis and the photoperiodic induction of flowering. Plant J 10:691–702
Zakhrabekova S, Gough SP, Braumann I, Muller AH, Lundqvist J, Ahmann K, Dockter C, Matyszczak I, Kurowska M, Druka A, Waugh R, Granerd A, Steind N, Steuernageld B, Lundqviste U, Hansson M (2012) Induced mutations in circadian clock regulator Mat-a facilitated short-season adaptation and range extension in cultivated barley. Proc Natl Acad Sci U S A 109:4326–4331
Zhao J, Huang X, Ouyang X, Chen W, Du A, Zhu L, Wang S, Deng XW, Li S (2012) OsELF3-1, an ortholog of Arabidopsis EARLY FLOWERING 3, regulates rice circadian rhythm and photoperiodic flowering. PLoS One 7:e43705
Zikhali M, Leverington-Waite M, Fish L, Simmonds J, Orford S, Wingen LU, Goram R, Gosman N, Bentley A, Griffiths S (2014) Validation of a 1DL earliness per se (eps) flowering QTL in bread wheat (Triticum aestivum). Mol Breed 34:1023–1033
Zikhali M, Wingen LU, Griffiths S (2015) Delimitation of the Earliness per se D1 (Eps-D1) flowering gene to a subtelomeric chromosomal deletion in bread wheat (Triticum aestivum). J Exp Bot. doi:10.1093/jxb/erv458
Acknowledgments
This project was supported by the National Research Initiative Competitive Grants 2009-35304-05091, 2011-67013-30077, and 2011-68002-30029 (Triticeae-CAP) from the USDA National Institute of Food and Agriculture, by the Howard Hughes Medical Institute and the Gordon and Betty Moore Foundation grant GBMF3031, and by the grants PICT Raíces 2006-1744 and 2011-509 from FONCYT (Argentina). We are grateful to Mariana Padilla for excellent technical support.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 479 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Alvarez, M.A., Tranquilli, G., Lewis, S. et al. Genetic and physical mapping of the earliness per se locus Eps-A m 1 in Triticum monococcum identifies EARLY FLOWERING 3 (ELF3) as a candidate gene. Funct Integr Genomics 16, 365–382 (2016). https://doi.org/10.1007/s10142-016-0490-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10142-016-0490-3