
Abstract
This study aims to investigate the antimicrobial and antibiofilm activity of berberine chloride (BBR) and vancomycin (VAN) as well as synergistic combinations of BBR with VAN against Clostridioides difficile strains. The effect of different concentrations of BBR on strain motility was also assessed. Twelve C. difficile strains (two reference C. difficile 630, ATCC 9689, and one control M120, and 9 clinical C. difficile strains belonging to the PCR-ribotype (RT027)) were collected and investigated for their susceptibility to BBR and VAN in planktonic and biofilm forms. Both the minimum inhibitory concentration (MIC) and the minimum bactericidal concentration (MBC) of BBR for the C. difficile strains were found to vary over a broad range (256–1.024 mg/L and 256–16.384 mg/L, respectively). The MIC and MBC of VAN also varied greatly, ranging from 0.25 to 4.0 mg/L for MIC and 0.25 to 64.0 mg/L for MBC. The synergistic effect of the sub-MIC (1/2 MIC) BBR with VAN reduced of MICs of VAN against the planktonic forms of ten C. difficile strains. The sub-MIC of BBR enhanced the biofilm formation of one strain and was found to be statistically significant. In addition, the sub-MIC of BBR with VAN surprisingly enhanced the biofilm formation of one C. difficile strain. The effect of inhibition of motility in the presence of BBR was statistically significant for 3 clinical strains (p < 0.05). Altogether, BBR exhibited strong antimicrobial activity against C. difficile, and the analysis of the combination of BBR with VAN showed a synergistic effect.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Clostridioides difficile (formerly Clostridium difficile) is one of the most common multi-drug-resistant organisms in hospital-acquired infections and is associated with high morbidity and mortality [1,2,3]. The main virulent factors of C. difficile are two toxins: toxin A (TcdA) and toxin B (TcdB). These virulence factors are glycosyltransferases that inactivate Rho, Ras, and Rac (GTP-binding proteins), resulting in the damage of the colonic epithelium and, subsequently, diarrhea [4]. Some strains possess a third toxin, called binary toxin (CDT) [1]. The increase of the incidence of Clostridioides difficile infections (CDI) is predominantly due to hyperepidemic strains of the genotype NAP1/BI/RT027/toxinotype III, which emerged at the beginning of the new millennium [5]. In a survey of CDI organized in Polish hospitals (2011–2013), C. difficile strains belonging to RT027 were the most prevalent PCR-ribotype in all hospitals involved in the study [6]. Strains belonging to RT027 are characterized by a higher production of toxin A (TcdA) and toxin B (TcdB) in vitro and the presence of binary toxin genes (cdtA and cdtB) [1, 4]. These proteins are the main virulent factors in pathogenesis of CDI. Other factors, such as fibronectin-binding protein A, surface layer proteins (SPLs), cell-wall proteins (CWPs), flagella, and cysteine protease Cwp84, have also been shown to be involved in the adhesion and colonization of the gut by C. difficile. Among these, Cwp84, flagella, LuxS protein, and surface layer proteins (SLPs) are especially associated with biofilm formation. A microbial biofilm is defined as a structured consortium of microbial cells surrounded by the self-produced matrix. Biofilms contribute to the tolerance of C. difficile to antibiotics, including those used as the first and second lines of the treatment for patients with CDI [7]. Metronidazole and vancomycin are the drugs of choice for the treatment of CDI; however, they are associated with a high incidence of relapse [8]. Although most C. difficile isolates are still susceptible to metronidazole and vancomycin, the resistance and reduced sensitivity of C. difficile to these drugs have been recently reported [9, 10]. Recurrent or persistent CDI have also been linked to the ability to produce biofilms [11]. Given the poor efficacy of standard treatment for frequent recurrences of CDIs, researchers have been actively searching treatment alternatives for several decades.
Berberine is a polyphenolic compound and a plant alkaloid that is isolated from Coptis chinensis (Chinese goldthread), Hydrastis canadensis (goldenseal), Rhizoma coptidis, Cortex phellode, and berberis [12]. Berberine chloride (BBR) is a quaternary ammonium isoquinoline alkaloid and is the most commonly available salt form of berberine [13]. Berberine is strong yellow in color and emits a strong yellow fluorescence under ultraviolet light [13]. BBR exhibits many biological functions and has been used to treat gastroenteritis, bacterium-associated diarrhea, and intestinal parasitic infections [14, 15]. The effect of berberine on C. difficile and in the treatment of CDI has been previously discussed in the literature [16,17,18]. Berberine in combination with ciprofloxacin was found to reduce the formation of biofilms by multi-resistant Salmonella sp. strains [19]. In this study, we investigated the antimicrobial and antibiofilm activity of synergistic combinations of berberine (BBR) and vancomycin (VAN) against C. difficile strains and the activity of BBR and VAN individually, as well as the effect of different concentrations of BBR on strain motility.
Materials and methods
Bacterial strains
Twelve C. difficile strains were used in this study: two reference strains (C. difficile 630 (RT012) and ATCC 9689 (RT001)), one control strain (C. difficile M120 (RT078)), and nine clinical strains consisting of clinical isolates and toxigenic strains belonging to the PCR-ribotype (RT027). The original numbers of all the clinical C. difficile strains were anonymized and designated a number from 4 to 12. The clinical C. difficile strains were previously isolated from patients with diarrhea admitted to Polish hospitals between 2012 and 2013. All C. difficile strains were collected in the Anaerobic Laboratory, in the Department of Medical Microbiology, at the Medical University of Warsaw. C. difficile strains were stored at − 70 °C in a Microbank™ bacterial storage system (Pro-Lab Diagnostics, UK) until use in the experiment. C. difficile 630 and M120 were kindly provided as a gift from Prof. Brendan Wren, in the Department of Pathogen Molecular Biology, at the London School of Hygiene and Tropical Medicine, London, UK. Strain ATCC9689 was purchased from bioMérieux (Marcy l’Etoile, France). The strains were thawed before use in experiments, cultured on Columbia blood agar solid medium (bioMérieux, Marcy l’Etoile, France) at 37 °C for 24 h under anaerobic conditions. All isolates were cultivated in brain–heart infusion (BHI; Difco, USA) medium at 37 °C under anaerobic conditions, unless stated otherwise.
Chemicals
Berberine chloride (BBR) (chemical formula C20H18CINO4) purchased from Sigma-Aldrich Co., Ltd. (St. Louis, MO, USA) was dissolved in DMSO (Biomus, Poland) and filtered through a 0.22-μm Millipore filter (Corning, USA). Vancomycin (VAN) was obtained from Sigma-Aldrich Co., Ltd. (St. Louis, MO, USA). BHI medium was obtained from (BIOMAXIMA, Lublin, Poland).
Determination of the minimal inhibitory concentration and minimal bactericidal concentration of berberine chloride and vancomycin for C. difficile strains
The minimal inhibitory concentrations (MICs) of berberine hydrochloride (BBR) and vancomycin (VAN) for the strains were determined using the broth microdilution method in a 96-well plate (Nunc, Denmark). An initial stock solution was prepared by dissolving BBR and VAN in dimethyl sulfoxide (DMSO, Biomus, Poland). Dilutions of 16.384, 8.192, 4.096, 2.048, 1.024, 900, 640, 512, 256, 128, 64, and 32 mg/L of BBR were prepared in BHI medium (Difco, USA). Vancomycin was used at the following dilutions: 64, 32, 16, 8, 6, 4, 2, 1, 0.5, 0.25, 0.125, and 0.0625 mg/L. Wells containing 180 μL of dilution were inoculated with 20 μL of suspension 3 McFarland turbidity strains C. difficile and incubated at 37 °C for 48 h under anaerobic conditions. The positive control (P) was BHI medium with 20 μL of suspension 3 McFarland strains C. difficile, and the negative control (N) was BHI medium. All strains were tested in triplicate. Following incubation, optical density at 600 nm was measured using a microplate reader (Bio-Rad, USA). The minimal bactericidal concentration (MBC) was determined by plating the cell suspensions in 96-well plates used for MIC tests onto Columbia agar containing 5% sheep blood (Beckton Dickinson, Heidelberg, Germany) and incubated at 37 °C for 48 h under anaerobic conditions. The bacterial growth was then visually observed.
Effect of berberine on MIC of vancomycin in C. difficile planktonic growth
The experiments were planned to stimulate situations in which bacteria are exposed to 1/2 MIC values (sub-MICs) of BBR with VAN. After the determination of the MICs of BBR, sub-MIC was added to the BHI medium with different concentrations of VAN. The synergism of BBR with VAN for the twelve strains was determined by the broth microdilution method in a 96-well plate. The following sub-inhibitory concentrations of BBR were prepared for individual strains: strains 2, 3, 5, 6, 7, 9, and 10: 128 mg/L of BBR; strains 11 and 12: 450 mg/L of BBR; strains 4 and 8: 320 mg/L of BBR; and strain 1: 512 mg/L of BBR, with the following concentrations of vancomycin: 64, 32, 16, 8, 4, 2, 1, 0.5, 0.25, 0.125, and 0.0625 mg/L. The solutions thus prepared were added at 180 μL to wells from 20 μL of a C. difficile strain suspension with a turbidity of 3 McFarland and incubated at 37 °C for 48 h. The positive control (P) was BHI medium with 20 μL suspension of 3 McFarland turbidity strains C. difficile, and the negative control (N) was BHI medium. All strains were tested in triplicate. After incubation, the growth of the bacteria was visualized to determine the synergistic MIC.
Effect of sub-inhibitory concentration of berberine and vancomycin on biofilm formation
Biofilm formation was tested according to the methods described previously [20]. All C. difficile strains were incubated overnight in a BHI medium (Difco, USA) at 37 °C. One hundred and eighty microliters of the BHI broth was pipetted into each well of a 96-well flat-bottomed microplate (Nunc, Denmark). Subsequently, 20 μL of C. difficile was added (three wells for each strain). Wells with BHI broth without the inoculum were used as the controls. The following sub-MICs of BBR were prepared for individual strains 2, 3, 5, 6, 7, 9, and 10: 128 mg/L; for strains 11 and 12: 450 mg/L; for strains 4 and 8: 320 mg/L; and for strain 1: 512 mg/L. Similarly, studies on biofilm production under the influence of vancomycin were carried out. Vancomycin sub-MICs were used for individual strains: 0.125 mg/L for strains 6 and 9; 0.25 mg/L for strains 3, 5, 7, 8, 10, and 11; 0.5 mg/L for strain 12; 1.0 mg/L for strain 2; and 2.0 mg/L for strains 1 and 4.
Plates were incubated at 37 °C for 48 h under anaerobic conditions for biofilm formation. After 48 h, the liquid phase was aspired using a sterile pipette, washed twice with phosphate buffer saline (PBS) (Biomed, Poland), and air-dried at 37 °C for 15 min. Each well was then stained with crystal violet (CV) (Analab, Poland) for 10 min. The CV was removed and the wells were washed eight times with PBS. After air-drying for 15 min at 37 °C, the CV within the biofilms was dissolved in ethanol and the absorbance was measured at 620 nm (A620) using a Bio-Rad 550 Microplate Reader (Bio-Rad, USA). All strains were tested six times. The average values for each C. difficile strain were calculated. The effect of berberine and vancomycin on C. difficile biofilms was determined using CV staining of cells in the biofilms in microtiter plates, based on the study by Stepanovic et al. [21], with modifications, as described in the previous section.
Determination of synergism for berberine with vancomycin in biofilm formation
In order to determine the formation of biofilm under the influence of sub-inhibitory doses of BBR with VAN, the following combinations of medium and agent concentrations were used: for strain 2: 128 mg/L of BBR and 1.0 mg/L of VAN; for strains 3, 5, 7, and 10: 128 mg/L of BBR and 0.25 mg/L of VAN; for strain 12: 450 mg/L of BBR and 0.5 mg/L of VAN; for strain 4: 320 mg/L of BBR and 2.0 mg/L of VAN; for strain 6 and 9: 128 mg/L of BBR and 0.125 mg/L of VAN; for strain 8: 320 mg/L of BBR and 0.25 mg/L of VAN; for strain 11: 450 mg/L of BBR and 0.25 mg/L of VAN; and for strain 1: 512 mg/L of BBR and 2.0 mg/L of VAN. All C. difficile strains were incubated overnight in a BHI medium at 37 °C. One hundred and eighty microliters of the BHI broth was pipetted into each well of a 96-well flat-bottomed microplate (Nunc, Denmark). Subsequently, 20 μL of C. difficile (three wells for each strain) was added to the wells. Wells with BHI broth without the inoculum were used as the controls. The plates were incubated at 37 °C for 48 h under anaerobic conditions for biofilm formation. After 48 h, the liquid phase was aspired using a sterile pipette, washed twice with phosphate buffer saline (PBS) (Biomed, Poland), and air-dried at 37 °C for 15 min. Each well was then stained with crystal violet (CV) (Analab, Poland) for 10 min. The CV was removed and the wells were washed eight times with PBS. After air-drying for 15 min at 37 °C, the CV within the biofilms was dissolved in ethanol and the absorbance was measured at 620 nm (A620) using a Bio-Rad 550 Microplate Reader. All strains were tested six times. The average values for each C. difficile strains were calculated.
Confocal laser scanning microscopy
Biofilms were visualized via confocal laser scanning microscopy (CLSM) according to methods previously described by Piotrowski et al. [22]. Only two strains, C. difficile 630 and clinical strain 4, were selected for the visualization of biofilm under CLSM, as they showed the highest vancomycin minimal inhibitory concentration (MIC = 4.0 mg/L) among the strains tested. Biofilms were grown on sterile 10-mm-diameter glass-bottom dishes (Nunc, Denmark). Overnight cultures of C. difficile were diluted in fresh BHI with or without the sub-inhibitory concentration of VAN, BBR, and VAN with BBR. The biofilms were grown on sterile glass-bottomed flasks. As a result, 2500 μL of BHI medium and 500 μL of overnight cultures of the two strains were obtained.
The biofilms were grown by culturing the strains as follows: BHI medium supplemented with 2 mg/L of VAN for strains 630 and 4; BHI medium supplemented with 512 mg/L of BBR for strain 630; and BHI medium supplemented with 320 mg/L of BBR for strain 4. The biofilms were allowed to grow for 48 h at 37 °C under anaerobic conditions. The mature biofilms were then washed twice using 10 mM MgSO4 before staining with acridine orange (10 μ/mL) for 30 min in the dark. The dishes were washed twice with 10 mM MgSO4. Imaging was performed using a Nikon A1R MP microscope with a Nikon Ti Eclipse series (Nikon, Japan) using a × 60 objective lens and immersion oil. Images were acquired at 2040 × 2048 pixels using a Z-step of 0.1 μm. Acridine orange was detected using an excitation wavelength of 488 nm and emission wavelength of 500–550 nm. Images were processed and analyzed using NIS-Elements AR v. 4.10 software.
Effect of a sub-inhibitory concentration of berberine on the motility of C. difficile in vitro
The effects of BBR on the motility of C. difficile were investigated in 12 strains. Among them, 3 strains were used as a control: motile 630, non-motile M120, and motility ATCC 9689 [23]. The 9 other strains consisted of clinical isolates and toxigenic strains belonging to the PCR-ribotype (RT027). After thawing, all strains were seeded on Columbia agar medium (bioMérieux, Marcy l’Etoile, France) and incubated for 48 h in an anaerobic atmosphere. For each strain, a single colony calibrated with 1 μL of C. difficile culture was plated onto pre-reduced 0.4% agar plates with BHI medium without BBR (as a control of motility) and with different concentrations of BBR according to the 1/2 MIC value of all the individual strains (512, 450, 320, or 128 mg/L), followed by incubating at 37 °C under anaerobic conditions. Readings were carried out after 24 h and 48 h. The inhibition of motility was assessed by measuring the diameter of the spot on the soft agar and comparing the results with the control plates (without BBR). The readings were performed independently by two separate individuals.
Statistical analysis
At least three independent replicates of each 96-well-plate experiment were performed. Statistical analysis was performed using Statistica software (version 13, StatSoft, Poland). The normal distribution of the values was confirmed using the Shapiro–Wilk test. The effect of a sub-inhibitory concentration of BBR on the inhibition of the motility of the C. difficile strains was assessed using the Mann–Whitney U test. Differences in biofilm formation were calculated via Kruskal–Wallis one-way analysis of variance (ANOVA). Differences were considered as statistically significant for p values < 0.05.
Results
MIC and MBC values of C. difficile with berberine and vancomycin
The summary of results is in Table 1. Susceptibility testing showed that the MIC values of BBR ranged from 256 to 1024 mg/L (median ca. 491.3 mg/L). Likewise, the MBC values of BBR ranged from 256 to 16.384 mg/L (median ca. 1.879 mg/L). Susceptibility testing showed that the MIC values of VAN ranged from 0.25 to 4.0 mg/L (median ca. 1.2 mg/L). Similarly, the MBC values of VAN ranged from 0.25 to 64.0 mg/L (median ca. 7.3 mg/L).
Berberine decreased the MIC of vancomycin in the plankton forms of C. difficile growth
Our results showed that the sub-MIC values of BBR significantly reduced the MIC values of VAN for ten strains. In the case of the VAN-resistant reference strain, strain 630 (MIC = 4 mg/L) and clinical strain 4 (MIC = 4 mg/L), the MICs of VAN under the influence of BBR decreased significantly from 4.0 to 0.5 mg/L and 1.0 mg/L, respectively. The MIC value of VAN of the reference strain ATCC 9689 was found to decrease from an MIC value of 2.0 mg/L to 0.0625 mg/L. For the M120 control strain, and for one clinical strain (no. 11), the MICs of VAN did not decrease and the value remained the same, i.e., 0.5 mg/L. All results of the synergism of BBR with VAN are presented in Table 1.
Effect of the sub-inhibitory concentration of berberine and vancomycin and berberine with vancomycin on biofilm formation
The treatment of 12 C. difficile strains with a sub-inhibitory concentration of BBR or VAN did not cause any significant differences in biofilm formation in strains, as follows: strain 630: BBR (p = 1.0) and VAN (p = 0.09); strain ATCC9689: BBR (p = 0.39) and VAN (p = 0.22); strain M120: BBR (p = 1.0) and VAN (p = 0.64); strain 4: BBR (p = 0.37) and VAN (p = 0.86); strain 6: BBR (p = 0.27) and VAN (p = 1.0.); strain 7: BBR (p = 1.0) and VAN (0.54); strain 8: BBR (p = 1.0) and VAN (p = 1. 0); strain 9: BBR (p = 1.0) and VAN (p = 1.0); strain 10: BBR (p = 0.22) and VAN (p = 1. 0); strain 11: BBR (p = 0.11) and VAN (p = 1.0); and strain 12: BBR (p = 1.0) and VAN (p = 0.92). For strain 5, a statistically significant p value was observed for BBR (p = 0.02), but not VAN (p = 0.99).
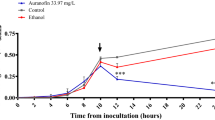
However, statistically significant differences in biofilm formation were observed for strain no. 5 (p = 0.02) treated with a sub-inhibitory concentration of BBR with VAN. In addition, treatment with sub-MICs of BBR and VAN was found to induce biofilm formation of strains no. 1 (630) and no. 11 but it was not statistically significant (p = 0.05) (Fig. 1).

Average biofilm formation by examined C. difficile strains with different concentrations of BBR, VAN, and the synergism of VAN and BBR. Legend: CTR, control; BBR, berberine; VAN, vancomycin; COM, combination of berberine and vancomycin
Confocal laser scanning microscopy
After investigating the influence of VAN and BBR on all twelve C. difficile strains, two strains were selected for biofilm testing by confocal microscopy. The two strains selected had the highest MIC values of VAN among the strains. For the biofilm testing of strain 630, a sub-inhibitory dose of VAN (2 mg/L) and BBR (512 mg/L) was used. This strain in the control produced a biofilm with a very homogeneous, carpet-like, dense structure (Fig. 2A). The generation of biofilm by strain 630 in the presence of 2 mg/L VAN was also very dense, but irregular with a high 3D architecture containing microaggregates (Fig. 2B). In the presence of a sub-inhibitory dose of BBR, the biofilm produced by strain 630 was very different to that produce by the control and biofilm with VAN. In this case, the biofilm was very thin and irregular and also contained microaggregates (Fig. 2C). The biofilm of strain 630 in 2 mg/L of VAN and 512 mg/L of BBR was thick, heterogeneous, and irregular with a small amount of microaggregates (Fig. 2D). Sub-inhibitory doses of VAN (2 mg/L) and BBR (320 mg/L) were also used to test a second strain of C. difficile: strain 4. This strain produced an irregular, heterogeneous biofilm control with a fairly high 3D architecture containing microaggregates (Fig. 2E). Under the influence of a sub-inhibitory dose of VAN, this strain produced a very thick, but rare, irregular biofilm with a 3D high architecture containing microaggregates (Fig. 2F). In the presence of BBR, this strain produced a biofilm with a regular structure and fairly high 3D architecture (Fig. 2G). The biofilm of strain 4 with 2 mg/L of VAN and 320 mg/L BBR was rather homogeneous and thick with a very high 3D architecture containing microaggregates (Fig. 2H).

Effects of VAN and BBR and VAN with BBR on reference C. difficile strain 630 and clinical RT027 (no. 4) on biofilm formation. Representative confocal microscopy images of horizontal (xy) and vertical (xz) projections of C. difficile biofilm structures. Slices viewed with maximum intensity projection. Legend: A, control; B, VAN 2 mg/L; C, BBR 512 mg/L; D, VAN 2 mg/L with BBR 512 mg/L; E, control; F, van 2 mg/L; G, BBR 320 mg/L; H, VAN 2 mg/L with BBR 320 mg/l
Sub-inhibitory concentrations of berberine inhibit motility of C. difficile strains
Three clinical C. difficile isolates (4, 11, and 12) showed inhibition of motility after the addition of sub-inhibitory (1/2 MIC) doses of BBR (320, 450, and 450 mg/L, respectively) to the BHI agar plates. This effect was statistically significant for strain 4 after 48 h (p < 0.01), for strain 11 (24 h, p = 0.016; 48 h, p = 0.02), and strain 12 (24 h, p = 0.01; 48 h, p < 0.01) (Table 2). The addition of BBR at 1/2 MIC on strains 630 (512 mg/L) and ATCC 9689 (128 mg/L), as well as strains 5 (128 mg/L), 6 (128 mg/L), 7 (128 mg/L), 8 (320 mg/L), 9 (128 mg/L), and 10 (128 mg/L), did not result in a statistically significant inhibition (p > 0.05). The effect of the sub-MICs of BBR on the inhibition of motility of selected C. difficile strains on soft agar is presented in Fig. 3.

Effect of a sub-inhibitory concentration of berberine on the motility of C. difficile in vitro. Legend: A, control (without BBR); B and C, with sub-MICs of BBR: 128 mg/L or 450 mg/L, respectively
Discussion
C. difficile is the leading multidrug-resistant pathogen in hospital-acquired diarrhea [1]. Currently, CDI is managed via the use of the two conventional antimicrobials, metronidazole and vancomycin, for the treatment of mild to moderate CDI and severe CDI, respectively. Fidaxomicin is a good alternative, especially in patients at risk of relapse [8]. However, several issues are associated with the use of these agents, including a high recurrence rate of 20–25%. C. difficile has developed resistance to several different classes of antimicrobials; however, the rate of resistance varies widely depending on the geographic regions and policies of antimicrobial use [24]. Natural products derived from food and other plant extracts have great antimicrobial potential against multi-drug-resistant microorganisms [25]. Berberine is a natural isoquinoline alkaloid that is increasingly drawing attention due to its multiple therapeutic effects, among others, on cancer, diabetes, hyperlipidemia, and cardiovascular diseases [26,27,28,29]. In traditional Chinese medicine, berberine has been widely used to treat bacterial diarrhea and gastroenteritis [30]. It has been previously indicated that, within the same species of bacteria, some strains may present distinct sensitivities to antimicrobial agents. Our research revealed that the MIC of berberine against C. difficile strains ranged from 256 to 1.024 mg/L. In contrast, Tan et al. showed that the MIC values of berberine varied from 64 to 512 mg/L against Staphylococcus aureus [31]. Wang et al. found that the MIC value of berberine for C. difficile spores was 640 mg/L [17]. In this study, a higher MBC was observed, possibly due to the fact that some C. difficile cells are found in the form of spores, which can wait later. It is likely that berberine will not enter the core of dormant spores due to the core’s extreme impermeability, consistent with the dormant spores’ resistance to antibiotics. The level of berberine accumulated at the berberine MICs in the individual germinated spores was heterogeneous for C. difficile. These values were 25–50-fold higher than the MIC values. However, berberine did not affect the germination of C. difficile spores, but did block the outgrowth of germinated spores. In our study, a higher MBC (in strain 1) was observed, at 16.384 mg/L. A certain amount of bacteria in special spores of C. difficile was able to survive and persist at high berberine concentrations. Our study tested the sensitivity of 9 clinical strains (RT027) to berberine. As a result, we found that berberine presented significant antibacterial activity against all strains. Furthermore, synergism was observed in berberine at 1/2 MICs combined with vancomycin. The great diversity of MICs among the C. difficile strains indicates the importance of determining the MIC value. Zuo et al. found that berberine significantly lowered the MIC values of a series of antibiotics against S. aureus [32].
In a previous study, the antimicrobial effect of berberine chloride in combination with various anti-staphylococcal drugs on reference CoNS strains was found to vary greatly depending on the bacterial strain and drug used. The most significant synergistic effects towards CoNS strains were noted when berberine was combined with linezolid, cefoxitin, and erythromycin [33]. In another study, the combined use of fusidic acid (FA) and berberine chloride (BBR) was found to result in an in vitro synergistic action against 7 out of 30 clinical methicillin-resistant Staphylococcus aureus (MRSA) strains [34]. In the present study, we examined the sub-MICs of berberine and sub-MICs of vancomycin for biofilm formation in C. difficile strains. The sub-MICs of BBR and VAN separately did not significantly increase biofilm formation in most strains except one. However, the sub-inhibitory concentration (1/2 MIC) of BBR with VAN was unexpectedly found to enhance biofilm formation in one clinical C. difficile strain. This is a form of adaptation by C. difficile to highly stressful environments produced by berberine with vancomycin, wherein the organism tended to live in its biofilm form instead of its planktonic form.
The flagella allow bacteria to move and contribute to bacterial colonization and pathogenesis by promoting adhesion to host cells, providing motor-driven nutrients and promoting biofilm formation [35]. The clinical strains tested in this study showed significant motility. Here, three clinical strains were found to have an inhibition of motility after the addition of sub-MIC (1/2 MIC) doses of BBR. The mechanism of action of berberine on the motility of C. difficile strains under the influence of sub-inhibitory doses of BBR is interesting: sub-inhibitory doses of BBR reduced the motility of clinical strains to varying degrees. To the best of our knowledge, this is the first study to focus on the biological effect of berberine both alone and in combination with vancomycin on C. difficile and biofilm formation.
In conclusion, specific biological substances exhibit antimicrobial properties. Our work demonstrates the antimicrobial ability of berberine against C. difficile. Importantly, we observed that sub-MICs of BBR with VAN can enhanced biofilm formation in some C. difficile strains.
References
Rupnik M, Wilcox MH, Gerding DN (2009) Clostridium difficile infection: new developments in epidemiology and pathogenesis. Nat Rev Microbiol 7:526–536. https://doi.org/10.1038/nrmicro2164
Kuijper EJ, Barbut F, Brazier JS, Kleinkauf N, Eckmanns T, Lambert ML et al (2008) Update of Clostridium difficile infection due to PCR ribotype 027 in Europe, 2008. Euro Surveill 13:18942
Lachowicz D, Pituch H, Obuch-Woszczatyński P (2015) Antimicrobial susceptibility patterns of Clostridium difficile strains belonging to different polymerase chain reaction ribotypes isolated in Poland in 2012. Anaerobe 31:37–41. https://doi.org/10.1016/j.anaerobe.2014.09.004
Smits WK, Lyras D, Lacy DB, Wilcox MH, Kuijper EJ (2016) Clostridium difficile infection. Nat Rev Dis Primers 2:16020. https://doi.org/10.1038/nrdp.2016.20
Krutova M, Kinross P, Barbut F, Hajdu A, Wilcox MH, Kuijper EJ, survey contributors (2018) How to: surveillance of Clostridium difficile infections. Clin Microbiol Infect 24:469–475. https://doi.org/10.1016/j.cmi.2017.12.008
Pituch H, Obuch-Woszczatyński P, Lachowicz D, Wultańska D, Karpiński P, Młynarczyk G, van Dorp SM, Kuijper EJ, Polish Clostridium difficile Study Group (2015) Hospital-based Clostridium difficile infection surveillance reveals high proportions of PCR ribotypes 027 and 176 in different areas of Poland, 2011 to 2013. Euro Surveill:20. https://doi.org/10.2807/1560-7917.ES.2015.20.38.30025
Ðapa T, Leuzzi R, Ng YK, Baban ST, Adamo R, Kuehne SA, Scarselli M, Minton NP, Serruto D, Unnikrishnan M (2013) Multiple factors modulate biofilm formation by the anaerobic pathogen Clostridium difficile. J Bacteriol 195:545–555. https://doi.org/10.1128/JB.01980-12
Ooijevaar RE, van Beurden YH, Terveer EM, Goorhuis A, Bauer MP, Keller JJ, Mulder CJJ, Kuijper EJ (2018) Update of treatment algorithms for Clostridium difficile infection. Clin Microbiol Infect 24:452–462. https://doi.org/10.1016/j.cmi.2017.12.022
Freeman J, Vernon J, Pilling S, Morris K, Nicholson S, Shearman S, Longshaw C, Wilcox MH (2018) Pan-European longitudinal surveillance of antibiotic resistance among prevalent Clostridium difficile Ribotypes Study Group. Clin Microbiol Infect 24:724–731. https://doi.org/10.1016/j.cmi.2017.10.008
Baghani A, Ghourchian S, Aliramezani A, Yaseri M, Mesdaghinia A, Douraghi M (2018) Highly antibiotic-resistant Clostridium difficile isolates from Iranian patients. J Appl Microbiol 125:1518–1525. https://doi.org/10.1111/jam.14035
Vuotto C, Donelli G, Buckley A, Chilton C (2018) Clostridium difficile biofilm. Adv Exp Med Biol 1050:97–115. https://doi.org/10.1007/978-3-319-72799-8_7
Cicero AF, Baggioni A (2016) Berberine and its role in chronic disease. Adv Exp Med Biol 928:27–45. https://doi.org/10.1007/978-3-319-41334-1
Neag MA, Mocan A, Echeverría J, Pop RM, Bocsan CI, Crişan G, Buzoianu AD (2018) Berberine: botanical occurrence, traditional uses, extraction methods, and relevance in cardiovascular, metabolic, hepatic, and renal disorders. Front Pharmacol 9:557. https://doi.org/10.3389/fphar.2018.00557
Silvester AJ, Aseer KR, Yun JW (2018) Dietary polyphenols and their roles in fat browning. J Nutr Biochem 64:1–12. https://doi.org/10.1016/j.jnutbio.2018.09.028
Rabbani GHBT, Knight J, Sanyal SC, Alam K (1987) Randomized controlled trial of berberine sulfate therapy for diarrhea due to enterotoxigenic Escherichia coli and Vibrio cholera. J Infect Dis 155:979–984. https://doi.org/10.1093/infdis/155.5.979
Lv Z, Peng G, Liu W, Xu H, Su J (2015) Berberine blocks the relapse of Clostridium difficile infection in C57BL/6 mice after standard vancomycin treatment. Antimicrob Agents Chemother 59:3726–3735. https://doi.org/10.1128/AAC.04794-14
Wang S, Setlow B, Setlow P, Li YQ (2016) Uptake and levels of the antibiotic berberine in individual dormant and germinating Clostridium difficile and Bacillus cereus spores as measured by laser tweezers Raman spectroscopy. J Antimicrob Chemother 71:1540–1546. https://doi.org/10.1093/jac/dkv504
Jiang H, Zhang D, He J, Han X, Lin J, Lan Y, Xiong X, Yu L, Yang M, Han L (2018) A novel method to mask the bitter taste of berberine hydrochloride: powder surface modification. Pharmacogn Mag 14:253–260. https://doi.org/10.4103/pm.pm_114_17
Shi C, Li M, Muhammad I, Ma X, Chang Y, Li R, Li C, He J, Liu F (2018) Combination of berberine and ciprofloxacin reduces multi-resistant Salmonella strain biofilm formation by depressing mRNA expressions of luxS, rpoE, and ompR. J Vet Sci 19:808–816. https://doi.org/10.4142/jvs.2018.19.6.808
Piotrowski M, Karpiński P, Pituch H, van Belkum A, Obuch-Woszczatyński P (2017) Antimicrobial effects of Manuka honey on in vitro biofilm formation by Clostridium difficile. Eur J Clin Microbiol Infect Dis 36:1661–1664. https://doi.org/10.1007/s10096-017-2980
Stepanovic S, Vukovic D, Dakic I, Savic B, Svabic-Vlahovic M (2000) A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J Microbiol Methods 40:175–179
Piotrowski M, Wultańska D, Obuch-Woszczatyński P, Pituch H (2019) Fructooligosaccharides and mannose affect Clostridium difficile adhesion and biofilm formation in a concentration-dependent manner. Eur J Clin Microbiol Infect Dis 38:1975–1984. https://doi.org/10.1007/s10096-019-03635-7
Stabler RA, He M, Dawson L, Martin M, Valiente E, Corton C, Lawley TD, Sebaihia M, Quail MA, Rose G, Gerding DN, Gibert M, Popoff MR, Parkhill J, Dougan G, Wren BW (2009) Comparative genome and phenotypic analysis of Clostridium difficile 027 strains provides insight into the evolution of a hypervirulent bacterium. Genome Biol 10:R102. https://doi.org/10.1186/gb-2009-10-9-r102
Spigaglia P (2016) Recent advances in the understanding of antibiotic resistance in Clostridium difficile infection. Ther Adv Infect Dis 3:23–42. https://doi.org/10.1177/2049936115622891
Roshan N, Riley TV, Knight DR, Steer JH, Hammer KA (2019) Natural products show diverse mechanisms of action against Clostridium difficile. J Appl Microbiol 126:468–479. https://doi.org/10.1111/jam.14152
Tong L, Xie C, Wei Y, Qu Y, Liang H, Zhang Y, Xu T, Qian X, Qiu H, Deng H (2019) Antitumor effects of berberine on gliomas via inactivation of caspase-1-mediated IL-1β and IL-18 release. Front Oncol 9:364. https://doi.org/10.3389/fonc.2019.00364
Wang Y, Yan A, Li S, Liu B, Li H, Yan Y (2019) Efficacy and safety of berberine in the treatment of type 2 diabetes with insulin resistance: protocol for a systematic review. Medicine (Baltimore) 98:e16947. https://doi.org/10.1097/MD.0000000000016947
Zhang LS, Zhang JH, Feng R, Jin XY, Yang FW, Ji ZC, Zhao MY, Zhang MY, Zhang BL, Li XM (2019) Efficacy and safety of berberine alone or combined with statins for the treatment of hyperlipidemia: a systematic review and meta-analysis of randomized controlled clinical trials. Am J Chin Med 47:751–767. https://doi.org/10.1142/S0192415X19500393
Feng X, Sureda A, Jafari S, Memariani Z, Tewari D, Annunziata G, Barrea L, Hassan STS, Šmejkal K, Malaník M, Sychrová A, Barreca D, Ziberna L, Mahomoodally MF, Zengin G, Xu S, Nabavi SM, Shen AZ (2019) Berberine in cardiovascular and metabolic diseases: from mechanisms to therapeutics. Theranostics 9:1923–1951. https://doi.org/10.7150/thno.30787
Chen C, Yu Z, Li Y, Fichna J, Storr M (2014) Effects of berberine in the gastrointestinal tract - a review of actions and therapeutic implications. Am J Chin Med 42:1053–1070. https://doi.org/10.1142/S0192415X14500669
Tan J, Wang J, Yang C, Zhu C, Guo G, Tang J, Shen H (2019) Antimicrobial characteristics of berberine against prosthetic joint infection-related Staphylococcus aureus of different multi-locus sequence types. BMC Complement Altern Med 19:218. https://doi.org/10.1186/s12906-019-2558-9
Zuo GY, Li Y, Han J, Wang GC, Zhang YL, Bian ZQ (2012) Antibacterial and synergy of berberines with antibacterial agents against clinical multi-drug resistant isolates of methicillin-resistant Staphylococcus aureus (MRSA). Molecules 17:10322–10330. https://doi.org/10.3390/molecules170910322
Wojtyczka RD, Dziedzic A, Kępa M, Kubina R, Kabała-Dzik A, Mularz T, Idzik D (2014) Berberine enhances the antibacterial activity of selected antibiotics against coagulase-negative Staphylococcus strains in vitro. Molecules 19:6583–6596. https://doi.org/10.3390/molecules19056583
Liang RM, Yong XL, Duan YQ, Tan YH, Zeng P, Zhou ZY, Jiang Y, Wang SH, Jiang YP, Huang XC, Dong ZH, Hu TT, Shi HQ, Li N (2014) Potent in vitro synergism of fusidic acid (FA) and berberine chloride (BBR) against clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA). World J Microbiol Biotechnol 30:2861–2869. https://doi.org/10.1007/s11274-014-1712-2
Baban ST, Kuehne SA, Barketi-Klai A, Cartman ST, Kelly ML, Hardie KR, Kansau I, Collignon A, Minton NP (2013) The role of flagella in Clostridium difficile pathogenesis: comparison between a non-epidemic and an epidemic strain. PLoS One 8:e73026. https://doi.org/10.1371/journal.pone.0073026
Acknowledgments
The authors would like to thank Prof. Brendan Wren, Department of Pathogen Molecular Biology, London School of Hygiene and Tropical Medicine, UK, for kindly providing the C. difficile 630 and M120 control strains. We would like to thank Dr. Bohdan Paterczyk, Faculty of Biology, University of Warsaw, for help in CLSM imaging. We wish to thank a Polish clinical microbiologist for C. difficile strains.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical approval
Clinical C. difficile strains were collected as part of routine Polish hospital surveillance. Ethical approval and informed consent were not required.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wultańska, D., Piotrowski, M. & Pituch, H. The effect of berberine chloride and/or its combination with vancomycin on the growth, biofilm formation, and motility of Clostridioides difficile. Eur J Clin Microbiol Infect Dis 39, 1391–1399 (2020). https://doi.org/10.1007/s10096-020-03857-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10096-020-03857-0