
Abstract
The livestock sector supports billions of people worldwide, yet when poorly managed, can have substantial negative effects on soils and ecosystem functions. Despite numerous studies and reviews of the effects of livestock activity on hydrological functions, a global synthesis of their effects on different biotic (vegetation, herbivore type) and abiotic (climate, soil, spatial scales) factors remains elusive. This makes it difficult to provide ecologically based advice on how best to manage grazing to minimise environmental damage and maximise hydrological functions. We used a global meta-analysis to examine the effects of livestock activity on hydrological inflows (infiltration, soil moisture), and outflows (runoff, sediment) using a dataset compiled from 3044 contrasts of grazed and ungrazed data from 129 publications between 1935 and 2020. Overall, we found that livestock activity increased sediment production (+ 52 ± 19.8%; mean ± 95% CI), reduced infiltration (− 25 ± 5.2%), but had no significant effect on either runoff (+ 27 ± 29.6%) or soil moisture (− 5 ± 5.9%). These impacts varied markedly among herbivore types and intensity, with greatest reductions in infiltration at high and low grazing contrasts, and more pronounced negative effects of grazing when sheep and cattle grazed together than either alone. The impact of livestock activity also varied with climatic region, soil texture, and the interaction between aridity and plant cover. Our study demonstrates the nuanced effects of livestock activity, with herbivore type and intensity, and environmental context modifying the hydrological outcomes. Livestock grazing is unlikely to result in positive environmental outcomes for infiltration and sediment production.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Highlights
-
Livestock activity increases sediment production but reduces infiltration.
-
Hydrological effects of livestock activity vary with herbivore type and intensity.
-
Impacts of livestock activity depend on vegetation community, climatic region and soil texture.
Introduction
The livestock sector is a substantial global industry, accounting for 40% of GDP, and employing 1.3 billion people, of which 1 billion are from some of the world’s poorest societies (Steinfeld and others 2006). Grazing of livestock on native pastures accounts for 70% of all agricultural land and 30% of Earth’s land surface (Steinfeld and others 2006). It provides billions of people with multiple resources such as milk, meat, hide, fuel and fertiliser (dung), security, transportation, and the potential to accumulate capital (Campos and others 2016). Livestock grazing will continue to exhibit an upward trend globally (Asner and others 2004). Yet despite its importance, poorly managed livestock activity is often blamed for contributing to widespread land and water degradation (for example, desertification, water pollution), habitat destruction (for example, land degradation in grassland, rangeland) and contributes heavily to global CO2 emissions (Fleishner 1994; Eldridge and others 2016; Sanderman and others 2017). Predicted drier climates and increasing demand for water to support an ever-growing population and demand for agricultural products will place increasing pressure on dwindling water supplies and ecosystem stability in water-limited regions such as drylands, where grazing of livestock is the predominant land-use (Steinfeld and others 2006; Huang and others 2016).
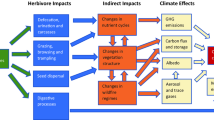
One of the most important impacts of overgrazing is its effect on soil hydrological processes; the movement of water through the soil profile (infiltration) or across the surface (runoff), its capture and storage in the soil (soil moisture), and the entrainment and transport of eroded sediment in runoff (Thurow 1991). Livestock grazing directly and indirectly impacts hydrological processes by direct removal of plants through herbivory and effects on soils by trampling (Figure 1). For example, herbivory can positively and negatively alter plant community composition, favouring short-statured annual species with fibrous root systems over taller perennials with deep-rooted systems (Clay and others 2005), and enhanced the richness of both native and exotic grasses and forbs (Stahlheber and D’Antonio 2013; Sasha and others 2017; Figure 1; Pathway 2). Indirectly, this reduces the capacity of soils to conduct water and can increase the likelihood that surface moisture is evaporated from the uppermost soil layers (for example, Thurow 1991; Pathway 6 via 2). Grazing-induced trampling not only reduces vegetation cover (Pathway 3) but also disrupts the connectivity of surface soil macropores (Marquart and others 2019), reducing infiltration (Pathway 9 via 1), or compacting the soil layers, exposing the surface to raindrop impact, and leading to reduced surface stability and greater water erosion (Gifford and Hawking 1978; Pathway 7 via 1). Grazing can also exert direct effects on hydrological processes by removing biocrusts (Eldridge 1998), communities of lichens and mosses (Eldridge and others 2020a), disaggregating erodible soil particles and increasing their susceptibility to raindrop activity and movement by overland flow (Teague and others 2011; Pathway 8). Direct trampling of the soil surface can also lead to the development of depositional crusts that enhance runoff (Castellano and Valone 2007; Pathway 8). Effects as broad as altering microbial community composition through changes in plant or litter composition could have a suppressive effect on organic matter decomposition and therefore hydrological processes, particularly infiltration or soil water holding capacity, or the ability to resist erosion (Kuske and others 2012; Zheng and others 2018; Pathways 4, 5, 7 and 8). These scenarios suggest that the effects of livestock on hydrological processes are far-reaching, complex and likely to be strongly nuanced, across different communities, contexts and environments (Eldridge and others 2016).

Flowchart of the hydrological response to herbivore activity (herbivory, trampling) via changes in vegetation and soil. Text outside the boxes is the moderators (syn. covariates) used to explore potential effects of activity on inflows (infiltration, soil moisture storage) and outflows (runoff, sediment).
The deleterious effects of livestock activity on hydrology compared to the ungrazed natural systems have been widely reported in the literature (for example, Gifford and Hawkins 1978; McCalla and others 1984; Thurow 1991; Holt 1997; Marquart and others 2019). However, such effects are unlikely to be felt uniformly across different environmental and management contexts. For example, grazing effects might differ among soils of different texture, given that loamy and clay soils have different levels of erodibility and compatibility, and therefore different susceptibility to soil detachment and sediment production (Gifford and Hawkins 1978). Soil moisture retention depends on soil texture, with sands expected to be less erodible by water than clay soils due to their high levels of saturated hydraulic conductivity. Similarly, the idiosyncratic effects of trampling, grazing behaviour, dentition and plant selectivity of different herbivores influence any effects on soils and soil hydrology (Eldridge and others 2016). Effects may also differ across different plant community types (vegetation assemblages) due to variation in plant forms and the differential responses to herbivory and trampling. Finally, any effects are likely to be strongly influenced by livestock grazing intensity, with stronger effects under increasing levels of grazing (Alderfer and Robinson 1947; Thurow 1991; Lai and Kumar 2020). Further, some components of hydrology such as hydrological outflows (runoff and sediment production) can yield important information on potential effects of rainfall intensity (Climate factor in the Universal Soil Loss Equation USLE; Wischmeier and Smith 1960) and plant cover (Management factor in USLE) on runoff and sediment production, when these are assessed directly with rainfall simulators of known intensity. Of the vast number of publications on the impacts of grazing, including reviews (Gifford and Hawkins 1978; Basche and DeLonge 2019) and recent meta-analyses (Union of Concerned Scientists 2017; Hao and He 2019; Lai and Kumar 2020), none has attempted to provide a systematic global synthesis of hydrological impacts, nor account for idiosyncratic effects of biotic (vegetation community, herbivore type, herbivore grazing intensity, plant cover) and abiotic (aridity level, soil texture, spatial scale, rainfall intensity) factors. Our review attempts such an assessment using a rigorous meta-analytical approach.
Here, we examine the global effects of livestock activity on soil hydrological processes using a dataset compiled from 129 studies published all between 1935 and 2020. We focused on four broad hydrological measures; two inflows and two outflows. Hydrological inflows comprised (1) infiltration, various measures of water flow through the soil such as hydraulic conductivity and ponded infiltration, and (2) soil moisture. Hydrological outflows included (3) runoff water lost from a site, that is, the difference between water applied and infiltration, assuming minimal interception), and (4) sediment production, the concentration or mass of soil entrained in runoff water. We hypothesized that grazing would lead to overall reductions in hydrological inflows (that is, infiltration, soil moisture) and therefore, more hydrological outflows (that is, runoff and erosion), but such an effect would likely vary with biotic (for example, vegetation assemblages, herbivore type, grazing intensity) and abiotic (aridity level, soil texture, spatial scale, rainfall intensity) factors, which explains the nuanced hydrological responses reported to date. Although some of our hypotheses are intuitive, that is, that more grazing leads to reduced infiltration and therefore, more runoff and erosion (Gifford and Hawkins 1978; Lai and Kumar 2020), we are unaware of any studies that have attempted a quantitative, rigorous global synthesis despite the suspected negative effects of increasing livestock activity on hydrological processes. This study is, to our knowledge, the first global meta-analysis of hydrological consequences of grazing based on a systematic, meta-analytical approach accounting for the effects of herbivore activity, herbivore type, climatic context, vegetation assemblage and study scale. We hope that the results of this review lead to a better understanding of hydrological effects of livestock activity and therefore better environmental outcomes in systems where livestock grazing is a substantial land-use component.
Methods
Database Construction
We used a systematic meta-analytical approach (Nakagawa and Santos 2012) to evaluate the general impacts of grazing on hydrological processes and explore the factors driving the variation in such impacts. Meta-analyses are used widely in ecology to synthesize evidence from a large number of studies, to test hypotheses, and to evaluate ecological outcomes at global scales (Gurevitch and others 2018). Although the overall effect of any analysis may be non-significant, specific studies can show either significant positive or negative effects. Thus, while our study may show overall negative effects, specific studies, in particular locations, would be expected to deviate from the general trend depending on differences in management and environmental context.
We searched Web of Science for studies published from January 1935 to August 2020 on the hydrological impact of grazing across the globe. We used the following search string: Topic = (grazing OR trampling OR livestock OR treading) NEAR/5 (hydrology OR infiltration OR sorptivity OR runoff OR run-off OR erosion OR sediment OR hydraulic conductivity OR sediment yield). We supplemented this dataset by including papers from reviews and unpublished government reports. This resulted in 585 unique studies, which were then screened using the PRISMA procedure (Preferred Reporting Items for Systematic Reviews and Meta-Analysis, Figure S1 in Appendix S1: Liberati and others 2009). While there are many studies on the impacts of livestock on hydrology, we restricted our analysis of studies that contained: (1) quantitative results of experiments or trials conducted under natural field conditions, for example, excluding feed lot studies; (2) data that were relevant to hydrological responses only (for example, excluding non-hydrological responses to grazing activity); (3) data on livestock grazing impacts (that is, not combined with additional management impacts); (4) data for different levels of grazing (that is, no grazing, low grazing, moderate grazing and high grazing). By grazing, we refer to any effects of livestock including soil disturbance, pugging, vegetation destruction, and herbivory. We restricted herbivore activity to domestic livestock (cattle, sheep and goats). We did not consider native ungulates (for example, zebra, deer, giraffe) nor macropods (kangaroos), but acknowledge that disturbance by these animals potentially has direct and indirect impacts on hydrological processes. For example, the effects of feral horses on hydrological processes have been discussed elsewhere (Eldridge and others 2020b). Based on these criteria, studies were restricted to 129 publications (Appendix S2).
Data Collection
From each study, we recorded location, the intensity of grazing activity (that is, ungrazed, low, moderate or high grazing), sample size, the mean and the standard deviation for each hydrology-related measure, for all available measures of each grazing treatment. Data from multiple time-points were averaged, and data from multiple depths included as presented by the authors. We also extracted data on environmental conditions to further explore the impacts of grazing under different environmental contexts. Aridity (1—precipitation/potential evapotranspiration) was derived from the Consortium for Spatial Information (CGIAR-CSI, 2020) means of the 1950–2000 period (Zomer and others 2008), and soil properties (for example, texture categories, surface layer sand content) were obtained from the HWSD database (resolution 1 km; HWSD 2019). Numeric data from figures were extracted using the software Engauge Digitizer V 4.1. Our final database consisted of 14 hydrological attributes (Appendix S3), which were categorised into four broad hydrological variables, inflows (infiltration, soil moisture), and outflows (sediment, runoff).
Estimating Mean Effect Size
We calculated a log response ratio (LnRR) as the effect size to determine the effect of grazing on hydrological attributes: LnRR = ln(x̅t/x̅c) (Hedges and others 1999) where x̅t is the mean value of the hydrological variable in the grazing plot (that is, treatment), and x̅c is the mean value of the hydrological variables in either the ungrazed plot or the plot with the lower intensity of grazing (defined as the control). Thus, positive values indicate an increase in the value of a given hydrological attribute with greater livestock grazing intensity and vice versa. Our final dataset contained 3044 contrasts between grazing intensity treatments and controls.
Meta-regression Model Approach
Random effects models were used to estimate mean effect sizes for grazing impacts. All of our models included random factors and a variance matrix structure to account for three potential sources of non-independence: (1) the effect size within the same study, (2) the residuals of data record among different studies, and (3) shared controls among observations from a single study that reported multiple levels of grazing but only a single control (for example, lower grazing level or ungrazed plot). To control for any potential lack of independence within and between studies, we included two random factors in our models, a unique identifier for each study to account for the within study variance, and the order of the data within the data file to account for the between-study variance and difference in sample size. To control for the potential influence of shared controls, which would cause non-independence among data with the same control (ungrazed), we included a coded group used to identify shared controls (Nakagawa and Santos 2012). To ensure the successful fitting of either random model, we excluded LnRR data in which variance was either too small (≤ 0.0001) or too large (≥ 1000), which is standard practice in meta-analyses (Nakagawa and Santos 2012).
Our modelling approach involved three stages. We first ran four intercept-only (null) models for (1) infiltration, (2) soil moisture, (3) sediment, (4) runoff using LnRR as our response variable and included the two random factors and the variance matrix described above. The null models allowed us to estimate the mean effect sizes for an overall response for each response variable. The significance of the estimated effect sizes was examined using a t-test, testing whether the estimated effect size was significantly different from zero in the random effect model at P < 0.05. Because our meta-analysis (intercept) models had high levels of heterogeneity (I2 > 0.95), we then ran meta-regression models which used a range of moderators such as, climate, soil texture, experiment scale, plant community, herbivore type, and grazing intensity as the fixed effect, to explore the impact of these moderators on the four hydrological response variables, respectively. The six moderators (Figure S2 in Appendix S4) were as follows: (a) climate zones (that is, arid, semiarid, dry subhumid, humid; these climate zones were divided based on aridity values), (b) soil texture categories (that is, clay, loam, sand), (c) experiment scale where data were measured (fine, < 0.05 m2, generally permeameters or micro-rainfall simulators; medium, 0.05–10 m2, larger rainfall simulators; large, > 10 m2, instrumented watersheds), (d) plant community type (that is, grassland, shrubland, woodland, forest), (e) herbivore type (cattle, goat, sheep, and mixed, that is, cattle and sheep), (f) grazing intensity levels (that is, ungrazed to high grazing, low to high, ungrazed to moderate, moderate to high, low to moderate, ungrazed to low). For any category, modelling requires data from at least two different studies. As runoff and sediment are the negative outcomes, and infiltration and soil moisture are positive outcomes of grazing that most concern land managers, ranchers, pastoralists and conservationists, we further explored how these factors drive hydrological responses by fitting linear models to our data. For runoff and sediment, our models included aridity (continuous value), soil sand content, plant cover (log response ratio of the plant cover in plots under higher grazing compared with lower grazing), rainfall intensity (mm h−1), grazing intensity (1 = ungrazed cf. low, 2 = low cf. moderate, 3 = moderate cf. high, 4 = ungrazed cf. to moderate, 5 = low cf. high, 6 = ungrazed cf. high grazing) as predictors and we also included an interaction between aridity and all other predictors (that is, plant cover, rainfall intensity, grazing intensity) to evaluate whether the impact of these drivers changes with increasing aridity. For infiltration and soil moisture, however, data were not available for plant cover or rainfall intensity, largely due to the way that these two hydrological response variables are measured. We therefore ran similar linear models, but excluded plant cover and rainfall intensity for these attributes.
To assess any potential publication bias, we used funnel plots, Egger regression and 'trim and fill' approaches (Nakagawa and Santos 2012) across the whole dataset, and for the four hydrological attributes (see Appendix S5). Meta-analysis was performed in the ‘metafor’ package (Viechtbauer 2010), and linear models were fitted in R 3.4.1 version (R Core Team 2013).
Results
Our review of the literature yielded 585 references from which we identified 129 publications with empirical and herbivore specific data on the effects of herbivore activity on hydrological responses. From these publications, we extracted 3044 contrasts of an effect of livestock grazing on four broad hydrological variables from five continents (Asia, Europe, Australia, North America, South America, Africa; Figure 2). Most data reported information on water flow through the soil (infiltration, sorptivity; 77%; n = 2334 contrasts; Appendix S3), followed by sediment production (13%; n = 397), soil moisture (5%; n = 166) and runoff (5%; n = 147). Most studies (69%) were from semiarid areas (Figure S2a in Appendix S4) and either grasslands (56%) or woodlands (36%; Figure S2b), but evenly distributed among clays, loams and sandy soils (Figure S2c). Studies focused mainly on the effects of cattle (55%) or sheep (26%; Figure S2d), with a relatively even distribution across the six different grazing contrasts (Figure S2e). Sixty-seven percent of observations were conducted at the fine (< 0.05 m2) spatial scales (Figure S2f).

The global distribution of sites (red circles) used in the meta-analysis and the percentage of comparisons from each continent shown in different colours. Circle size represents the number of studies from each region.
Overall, we found significantly greater sediment production (+ 52%; P < 0.001) but lower infiltration (− 25%; P < 0.001) across all grazing contrasts. The effects of grazing on runoff were marginally significantly positive (+ 27%, P = 0.07), but there was no significant grazing effect on soil moisture (− 5%, P = 0.12; Figure 3).

a mean value of the log response ratio (± 95% CI) and the number of contrasts used in the analyses of each hydrological response variable, b–e frequency distribution of effect sizes for each hydrological response variable. Note that 120 observations were removed automatically from the analyses due to anomalous variances (variances ≤ 0.0001 or ≥ 1000).
Effects of Grazing Intensity and Herbivore Type
The response of hydrological functions to changes in grazing intensity varied with the individual attribute. For example, linear modelling showed that infiltration declined marginally with increasing grazing pressure, but was reduced most strongly (− 37 to − 41%) when grazing increased from Ungrazed to Low, or Ungrazed to High. At intermediate levels of grazing (for example, Moderate to High), reductions in infiltration were less pronounced (Figure 4). There was a trend of greater sediment loss and runoff with increasing grazing intensity (Figure 4). Soil moisture was significantly affected by grazing intensity at only intermediate levels of intensity (Moderate cf. High; Figure 4). Herbivore type also had some effects on infiltration, with a more pronounced effect when sheep and cattle grazed together than when either grazed alone. Cattle reduced soil moisture content more than sheep, but their effects on sediment production were equivocal (Figure 4).

Effects of livestock grazing, as measured with the log response ratio (lnRR ± 95% CI), on four hydrological processes: infiltration, soil moisture (moisture), runoff and sediment production (sediment). Results are separated by different levels of each of the six moderators (1) Aridity (arid, semiarid, dry subhumid, humid), (2) Community (forest, woodland, shrubland, grassland), (3) Soil texture (sand, loam, clay), (4) Herbivore type (cattle, sheep, goats, mixed), (5), Grazing intensity (U-H = ungrazed to high grazing; L–H = low to high; U-M = ungrazed to moderate; M-H = moderate to high; L-M = low to moderate; U-L = ungrazed to low) and (6) Measurement scale (fine, medium, large). Significant results are indicated by whether the 95% CI spans the x = 0 line. Positive values show that increasing grazing increased the value of that hydrological response variables, while negative values show that increasing grazing reduced it. ND = no data. Note: some attributes with a significant effect may appear non-significant due to the size of the symbol, which intersects the x = 0 line.
Effects of Abiotic Moderators
We also found some soil texture effects. For example, the suppression of infiltration was more pronounced on sandy (− 43%) than loamy or clay (− 23 to − 25%) soils, while increasing sediment production was greater on loams (+ 67%) than clays (+ 30%). Loamy soils had lower soil moisture (− 8%) and greater runoff (+ 34%) than those from other texture groups (Figure 4). Linear models also showed a significant but small increase in infiltration with increasing aridity (P = 0.006).
There was no evidence of livestock activity effects on any hydrological responses across different climatic zones (Figure 4). However, the suppression of infiltration by livestock activity was more pronounced in forests (− 51%) than woodlands, shrublands or grasslands (− 10 to − 28%; Figure 4). There were no clear trends for runoff or soil moisture, but a suggestion that the effects of livestock activity on increased sediment were more pronounced in grasslands and shrublands (+ 51 ~ + 66%) than woodlands (+ 38%). Finally, because some studies were conducted at different spatial scales, a consequence of different study methods (for example, large plot-level simulations cf. micro-infiltrometers), we examined potential scale-related effects. Our analyses revealed no appreciable effect of scale on the relationship between livestock activity and hydrological responses.
We also found some significant interactions (Figures S4 and S5 in Appendix S6), for example, the effect of livestock activity on sediment production increased with increasing rainfall intensity at more arid sites but showed the opposite response in more humid sites (1.85 ± 1.58, estimate ± 95% CI; P = 0.024; Figure S4a). There was also an effect of runoff. As aridity increased, the impact of vegetation cover on runoff changed from negative to positive (0.66 ± 0.46, P = 0.008; Figure S4b). Linear modelling showed no effects of our moderators on soil moisture.
Discussion
We found that livestock activity, which included effects of herbivory and soil disturbance (that is, trampling), had substantial effects on hydrological processes. Overall, there were strong and significant increases in sediment production, reductions in infiltration, but marginal significant increases in runoff. Moreover, we found that herbivore type and the intensity of livestock activity had some impacts on hydrological responses, with greater reductions in infiltration at high compared with low livestock activity, and more pronounced effects when mixed herbivores (sheep and cattle) grazed together than when either grazed alone. Additionally, our results also suggest a context-dependency of herbivore activity on hydrology, varying with vegetation community, climatic region and soil texture. Together our results demonstrate substantial deleterious effects of livestock activity on hydrological inflows and outflows, particularly at higher levels of livestock intensity, but with no overall changes in available soil moisture.
Hydrological Effects Differ with Herbivore Type and Intensity
Consistent with our findings and abundant historical and contemporary literature (Rauzi 1963; Rhoades and Locke 1964; Gifford and Hawkins 1978; Thurow and others 1986; Lai and Kumar 2020), increasing livestock activity led to significant declines in infiltration, and increases in runoff and sediment production, particularly at high grazing intensities, and irrespective of the specific environmental conditions. Further, there is a suggestion of a greater reduction in infiltration when cattle and sheep grazed together than when either grazed alone. Herbivore-specific effects are not unexpected, given marked differences in foraging behaviour, plant selectivity, dentition, and body mass among different herbivore types. These factors would have direct effects on surface compaction, soil disturbance and erosion, and indirect effects via changes in plant cover composition or structure. Both sheep and cattle tend to move along clearly defined pathways, but these vary in size, extent and depth due to differences in herbivore body size (Andrew 1988; Dougill and Cox 1995). Further, larger body-sized herbivores such as cattle tend to consume large amounts of highly fibrous material, particularly lower digestibility grasses (Squires 1981). Sheep are typically more selective, with a greater preference for shorter forbs and herbs, given their upper incisor, which allows them to crop material closer to the ground than cattle. It is likely therefore, that when cattle and sheep graze together, that they exert additive and potentially synergistic effects on vegetation structure and soils, and therefore, on hydrological inflows and outflows (Eldridge and others 2017).
We also found a greater reduction in soil moisture content under grazing by cattle than by sheep, but their effects on sediment production were equivocal. This could be due to herbivore-specific trampling effects. For example, cattle have a greater body mass and five-times greater hoof area than sheep and goats (Greenwood and McKenzie 2001) and exert a higher pressure (98 kg cm−1) than sheep and goats (71 kg cm−1; Ssemakul 1983). Greater grazing pressure would reduce porosity and therefore hydraulic conductivity by reducing the positive effects of invertebrate-derived macropores on the surface (pathways 9 via 1; Figure 1) with attendant increases in runoff (pathway 7 via 1). Lower porosity would prevent movement and storage of water in the uppermost layers, thus resulting in greater reductions in soil moisture. Specific herbivore effects could influence hydrological processes by favouring annual species with weakly developed root systems of low hydrological conductance compared with perennial, macropore-producing grasses and perennial forbs, which are effective conductors of water (Basche and Edelson 2017; Basche and DeLonge 2019). For example, cattle and sheep has been shown to increase plant richness (Socher and others 2013; Gao and Carmel 2020) and we have shown previously that the effects of grazing on plant composition were mediated through the simplification of habitat structure (Eldridge and others 2016). Notwithstanding the oft-reported reduction in perennial grasses under grazing, in some situations, herbivore activity can promote perennial plants over annuals (for example, Papanikolaou and others 2011), thereby maintaining soil hydrological processes.
Our analysis found that runoff increased (value became more positive) as the intensity of livestock activity increased, and there were some ill-defined effects of grazing intensity on infiltration. Increased infiltration under grazing has been reported elsewhere (for example, Franzluebbers and others 2012), but generally only under conditions of low grazing intensity or specific grazing management practices (Briske and others 2008). Based on our global dataset, a progression from ungrazed to low grazing was associated with less runoff, but surprisingly this did not correspond to increased infiltration. Low levels of grazing have been shown to reduce plant cover sufficiently to promote overland flow (Alderfer and Robinson 1947), so suppression of runoff likely relates to animal effects on the soil, perhaps greater surface detention resulting from pugging (Greenwood and McKenzie 2001), and more rainfall interception by vegetation and/or litter, rather than greater infiltration in the soil. Runoff from heavily grazed landscapes has been shown to be almost an order of magnitude greater than that under light grazing (Rauzi and Hanson 1966). Although grazing could also reduce the accumulation of organic residues, promote the establishment of plants that prevent runoff (Carson and Peterson 1990) and exert positive effect on diversity in productive systems, these effects may only apply if the competitive dominant species are palatable and accessible to herbivores. Thus, the effects of trampling on runoff are generally exacerbated under high levels of grazing activity across the globe (Weltz and Wood 1986). We also detected the highest reduction in infiltration under greatest grazing intensity. Apart from the direct effect on vegetation and soil porosity, grazing will also likely have indirect effects on hydrology by reducing the recruitment of woody plants (Lohbeck and others 2020), which have been shown to conduct large amounts of infiltration, particularly preferential flow paths for deep percolation to the water table (Dreccer and Lavado 1993).
Although our meta-analysis shows that livestock grazing generally has negative effects on hydrology across the globe, we acknowledge that there are many situations where livestock can have positive effects on hydrological function. Some grazing management systems are more hydrologically friendly than others. Conventional set stocking grazing management systems, for example, whereby livestock remain in a single paddock (allotment) year-round are likely to result in stronger declines in soil structure, infiltration and moisture availability, with more runoff and sediment production than systems where animal numbers are adjusted according to seasonal conditions (Thurow 1991; Webber and others 2010; Sirimarco and others 2018). Conversely, systems based on the periodic movement of livestock among paddocks (for example, rotational or multi-paddock grazing) are more likely to enhance hydrological function (Hillenbrand and others 2019; Döbert and others 2021) when grazing intensity is low or moderate. However, the data limitation on different management systems prevents us from disentangling these nuance effect. Rotational systems that use high-intensity livestock grazing for extended periods, however, are likely to result in greater runoff and lower infiltration than those using low or moderate livestock grazing (Briske and others 2008).
The mechanisms behind these grazing management effects likely relate to changes in plant species composition due to different plant preference by livestock. Livestock grazing could increase plant richness and therefore hydrological function by releasing subordinate species from competitive exclusion by larger plants (Milchunas and others 1988). Similarly, greater litter cover resulting from the grazing of large dominant plants would be expected to improve hydrological function by retaining moisture near the soil surface (Deutsch and others 2010). Altogether, these increases in perennial plants, resulting from either climate change, variable grazing management regime or livestock preference would be expected to improve soil porosity, thereby increasing infiltration, and reducing runoff and sediment production (Basche and DeLonge 2019).
Hydrological Effects of Livestock Grazing Activity Vary with Climate
Despite the strong increase in sediment production with increasing livestock intensity (Figure 4), this effect varied with rainfall intensity. Specifically, increasing rainfall intensity was associated with a slight increase in sediment production in arid environments, but strongly reduced sediment production in humid areas. This result is not unexpected given the strong links between climatic zone and plant cover. Areas of high rainfall are likely to support greater plant cover, thereby reducing the likelihood of raindrop detachment and sediment transport. Despite higher rainfall intensity and erosivity in more mesic areas, it is likely that overland flow, the vehicle for sediment transport, is lower due to greater opportunities for infiltration, more surface litter, greater plant biomass and therefore greater porosity of mesic soils, and thus, lower sediment transport rates (Verheijen and others 2009). Conversely, soils in more arid areas have lower levels of organic carbon (Eswaran and others 1993), which, combined with a sparser plant cover and a greater proportion of interspace to perennial vegetation patch, might result in higher levels of runoff and sediment production. We also found that the effect of increasing plant cover on runoff was more pronounced in mesic than arid areas, and this is likely related to the type of cover that predominates in more arid areas. Arid and semiarid areas are typified by fertile patches dominated by perennial vegetation, separated by interspaces often with an assortment of biological soil crusts (biocrusts; Weber and others 2016). Thus, while vascular plant cover may be sparse, biocrust cover is typically extensive, and these crusts have a marked effect on hydrological processes (Eldridge and others 2020a). Various mechanisms for this reduction in runoff have been mooted including the fact that they retain soil moisture in the immediate surface horizons, with important implications for the productivity of dryland ecosystems and global hydrological effects (Eldridge and others 2020a).
Any effects of grazing on plants, such as changes in plant functional groups, and therefore hydrological function, could also be climate related. For example, in areas where summer rainfall is predicted to increase under changing climates, grazing could provide greater opportunities for competitive displacement of annuals by summer-growing perennial plants (Clarke 2003). Other landscape-level moderators might include environmental data such as rainfall characteristics, land use in the surrounding area and the connectivity of landscapes.
Caveats and Conclusions
Our study is based on global meta-analysis, which is used widely in ecology to synthesise evidence from a large number of studies, to test hypotheses, and to evaluate the general outcomes at global scales (Gurevitch and others 2018). However, a substantial constraint of all meta-analyses is the inability to capture the nuances of different biophysical, environmental and management factors associated with the data. In our analyses, many studies lacked sufficient data on specific information that prevented us from undertaking more subtle and nuanced statistical analyses. These explanatory attributes included grazing management practices such as the duration and timing of grazing (Trimble and Mendel 1995; Castellano and Valone 2007), the occurrence of feral or native co-herbivores such as deer, kangaroos, zebu (Hester and others 1996), ecosystem productivity (Lezama and others 2014), and the evolutionary history of grazing (Milchunas and others 1988) to name a few.
Nonetheless, despite the many idiosyncratic characteristics associated with these many studies, and the large variability in many of our moderators, we still found clear and unambiguous effects of grazing that promoted sediment production and reduced infiltration, and a strong suggestion of greater runoff; all markedly adverse effects on ecosystem functions, with the effects varying with herbivore type, grazing intensity and interactions with climatic regimes. We acknowledge, however, that log response ratios can be biased when sample sizes are small (n < 20; Nakagawa and others 2017), thus preventing us from exploring relationships for specific components of infiltration (depth to wetting front) and runoff (runoff coefficient, time to runoff, Table S1). Variance estimates can also be erroneous when the scale of study parameters is near zero (Lajeunesse 2015).
In conclusion, we found overall reductions in infiltration, and an increase in sediment production and runoff with increased grazing at the global scale, though more nuanced studies involving particular grazing systems, animal types and environments may well arrive at different conclusions for a specific region. However, we hope that this review stimulates further studies of the impacts of livestock on hydrological processes, particularly those involving different grazing management strategies that might have more positive effects on plants and soils.
References
Alderfer R, Robinson R. 1947. Runoff from pastures in relation to grazing intensity and soil compaction. Journal of the American Society of Agronomy 39:948–958.
Andrew M. 1988. Grazing impact in relation to livestock watering points. Trends in Ecology & Evolution 3:336–339.
Asner GP, Elmore AJ, Olander LP, Martin RE, Harris AT. 2004. Grazing systems, ecosystem responses, and global change. Annual Review of Environment and Resources 29:261–299.
Basche AD, DeLonge MS. 2019. Comparing infiltration rates in soils managed with conventional and alternative farming methods: A meta-analysis. PLoS One 14: e0215702.
Basche AD, Edelson OF. 2017. Improving water resilience with more perennially based agriculture. Agroecology and Sustainable Food Systems 41:799–824.
Briske D, Derner J, Brown J, Fuhlendorf S, Teague R, Gillen B, Ash A, Havstad K, Willms W. 2008. Benefits of rotational grazing on rangelands: an evaluation of the experimental evidence. Rangeland Ecology and Management 61:3–17.
Campos P, Ovando P, Mesa B, Oviedo JL. 2016. Environmental income of livestock grazing on privately-owned silvopastoral farms in Andalusia, Spain. Land Degradation and Development 29:250–261.
Carson WP, Peterson CJ. 1990. The role of litter in an old-field community: impact of litter quantity in different seasons on plant species richness and abundance. Oecologia 85:8–13.
Castellano M, Valone T. 2007. Livestock, soil compaction and water infiltration rate: evaluating a potential desertification recovery mechanism. Journal of Arid Environments 71:97–108.
CGIAR-CSI, 2020. “Global aridity index and potential evapotranspiration climate database v2”, accessed June 26, 2020. https://cgiarcsi.community/2019/01/24/global-aridity-index-and-potential-evapotranspiration-climate-database-v2/
Clarke PJ. 2003. Composition of grazed and cleared temperate grassy woodlands in eastern Australia: patterns in space and inferences in time. Journal of Vegetation Science 14:5–14.
Clay K, Holah J, Rudgers JA. 2005. Herbivores cause a rapid increase in hereditary symbiosis and alter plant community composition. Proceedings of the National Academy of Sciences 102:12465–12470.
Deutsch ES, Bork EW, Willms WD. 2010. Separation of grassland litter and ecosite influences on seasonal soil moisture and plant growth dynamics. Plant Ecology 209:135–145.
Döbert TF, Bork EW, Apfelbaum S, Carlyle CN, Chang SX, Khatri-Chhetri U. … Boyce, M. S. (2021). Adaptive multi-paddock grazing improves water infiltration in Canadian grassland soils. Geoderma, 401, 115314.
Dougill A, Cox J. 1995. Land degradation and grazing in the Kalahari: new analysis and alternative perspectives: Overseas Development Institute London.
Dreccer M, Lavado R. 1993. Influence of cattle trampling on preferential flow paths in alkaline soils. Soil Use and Management 9:143–148.
Eldridge DJ, Delgado-Baquerizo M, Travers SK, Val J, Oliver I. 2017. Do grazing intensity and herbivore type affect soil health? Insights from a semi-arid productivity gradient. Journal of Applied Ecology 54:976–985.
Eldridge DJ, Ding J, Travers SK. 2020a. Feral horse activity reduces environmental quality in ecosystems globally. Biological Conservation 241: 108367.
Eldridge DJ, Poore AG, Ruiz-Colmenero M, Letnic M, Soliveres S. 2016. Ecosystem structure, function, and composition in rangelands are negatively affected by livestock grazing. Ecological Applications 26:1273–1283.
Eldridge DJ, Reed S, Travers SK, Bowker MA, Maestre FT, Ding J, Havrilla C, Rodriguez-Caballero E, Barger N, Weber B. 2020b. The pervasive and multifaceted influence of biocrusts on water in the world’s drylands. Global Change Biology 26:6003–6014.
Eldridge DJ. 1998. Trampling of microphytic crusts on calcareous soils and its impact on erosion under rain impacted flow. Catena 33:221–239.
Eswaran H, Van Den Berg E, Reich P. 1993. Organic carbon in soils of the world. Soil Science Society of America Journal 57:192–194.
Fleischner TL. 1994. Ecological costs of livestock grazing in western North America. Conservation Biology 8:629–644.
Franzluebbers AJ, Paine LK, Winsten JR, Krome M, Sanderson MA, Ogles K, Thompson D. 2012. Well-managed grazing systems: A forgotten hero of conservation. Journal of Soil and Water Conservation 67:100A-104A.
Gao J, Carmel Y. 2020. A global meta-analysis of grazing effects on plant richness. Agriculture Ecosystems and Environment 302: doi:https://doi.org/10.1016/j.agee.2020.107072
Gifford GF, Hawkins RH. 1978. Hydrologic impact of grazing on infiltration: a critical review. Water Resources Research 14:305–313.
Greenwood K, McKenzie B. 2001. Grazing effects on soil physical properties and the consequences for pastures: a review. Australian Journal of Experimental Agriculture 41:1231–1250.
Gurevitch J, Koricheva J, Nakagawa S, Stewart G. 2018. Meta-analysis and the science of research synthesis. Nature 555:175–182.
Hao Y, He Z. 2019. Effects of grazing patterns on grassland biomass and soil environments in China: A meta-analysis. PLoS One, 14: e0215223.
Hedges LV, Gurevitch J, Curtis PS. 1999. The meta-analysis of response ratios in experimental ecology. Ecology 80:1150–1156.
Hester A, Mitchell F, Gordon I, Baillie G. 1996. Activity patterns and resource use by sheep and red deer grazing across a grass/heather boundary. Journal of Zoology 240:609–620.
Hillenbrand M, Thompson R, Wang F, Apfelbaum S, Teague R. 2019. Impacts of holistic planned grazing with bison compared to continuous grazing with cattle in South Dakota shortgrass prairie. Agriculture, Ecosystems & Environment 279:156–168.
Holt J. 1997. Grazing pressure and soil carbon, microbial biomass and enzyme activities in semi-arid northeastern Australia. Applied Soil Ecology 5:143–149.
Huang J, Yu H, Guan X, Wang G, Guo R. 2016. Accelerated dryland expansion under climate change. Nature Climate Change 6:166–171.
HWSD, 2019. “Harmonized world soil database v12”, accessed November 12, 2019. http://www.fao.org/soils-portal/soil-survey/soil-maps-and-databases/harmonized-world-soil-database-v12/en/
Kuske CR, Yeager CM, Johnson S, Ticknor LO, Belnap J. 2012. Response and resilience of soil biocrust bacterial communities to chronic physical disturbance in arid shrublands. The ISME Journal 6:886–897.
Lai L, Kumar S. 2020. A global meta-analysis of livestock grazing impacts on soil properties. PLoS One 15: e0236638.
Lajeunesse MJ. 2015. Bias and correction for the log response ratio in ecological meta-analysis. Ecology 96:2056–2063.
Lezama F, Baeza S, Altesor A, Cesa A, Chaneton EJ, Paruelo JM. 2014. Variation of grazing-induced vegetation changes across a large-scale productivity gradient. Journal of Vegetation Science 25:8–21.
Liberati A, Altman DG, Tetzlaff J, Mulrow C, Gøtzsche PC, Ioannidis JP, Clarke M, Devereaux PJ, Kleijnen J, Moher D. 2009. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. Journal of Clinical Epidemiology 62:e1–e34.
Lohbeck M, Albers P, Boels LE, Bongers F, Morel S, Sinclair F, Takoutsing B, Vågen T-G, Winowiecki LA, Smith-Dumont E. 2020. Drivers of farmer-managed natural regeneration in the Sahel. Lessons for Restoration. Scientific Reports 10:1–11.
Marquart A, Eldridge DJ, Travers SK, Val J, Blaum N. 2019. Large shrubs partly compensate negative effects of grazing on hydrological function in a semi-arid savanna. Basic and Applied Ecology 38:58–68.
McCalla G, Blackburn W, Merrill L. 1984. Effects of livestock grazing on infiltration rates, Edwards Plateau of Texas. Journal of Range Management 37:291–294.
Milchunas DG, Sala OE, Lauenroth WK. 1988. A generalized model of the effects of grazing by large herbivores on grassland community structure. The American Naturalist 132:87–106.
Nakagawa S, Noble DW, Senior AM, Lagisz M (2017). Meta-evaluation of meta-analysis: ten appraisal questions for biologists. BMC Biology Mar 3;15(1):18. doi: https://doi.org/10.1186/s12915-017-0357-7. PMID: 28257642; PMCID: PMC5336618.
Nakagawa S, Santos ES. 2012. Methodological issues and advances in biological meta-analysis. Evolutionary Ecology 26:1253–1274.
Papanikolaou AD, Fyllas NM, Mazaris AD, Dimitrakopoulos PG, Kallimanis AS, Pantis JD. 2011. Grazing effects on plant functional group diversity in Mediterranean shrublands. Biodiversity and Conservation 20:2831–2843.
Rauzi F, Hanson CL. 1966. Water intake and runoff as affected by intensity of grazing. Journal of Range Management 19:351–356.
Rauzi F. 1963. Water intake and plant composition as affected by differential grazing on rangeland. Journal of Soil and Water Conservation 18:114–116.
Rhoades ED, Locke E. 1964. Water intake on a sandy range as affected by 20 years of differential cattle stocking rates. Journal of Range Management 17:185–190.
R Core Team. 2013. R: A language and environment for statistical computing.
Sanderman J, Hengl T, Fiske G. 2017. Soil carbon debt of 12,000 years of human land use. Proceedings of the National Academy of Sciences of the United States of America 114:9575–9580.
Sasha G, Erica S, Michele H, Bartolome JW. 2017. Livestock grazing supports native plants and songbirds in a California annual grassland. PLoS One, 12: e0176367.
Sirimarco X, Barral MP, Villarino SH, Laterra P. 2018. Water regulation by grasslands: A global meta-analysis. Ecohydrology. 2018; 11:e1934.
Socher SA, Prati D, Boch S, Müller J, Baumbach H, Gockel S, Hemp A, Schöning I, Wells K, Buscot F. 2013. Interacting effects of fertilization, mowing and grazing on plant species diversity of 1500 grasslands in Germany differ between regions. Basic and Applied Ecology 14:126–136.
Squires VR. 1981. Livestock Management in the Arid Zone. Publishers: Inkata Press, Melbourne.
Ssemakul J. 1983. A comparative study of hoof pressures of wild and domestic ungulates. African Journal of Ecology 21:325–328.
Stahlheber KA, D’Antonio CM. 2013. Using livestock to manage plant composition: a meta-analysis of grazing in California Mediterranean grasslands. Biological Conservation 157:300–308.
Steinfeld H, Gerber P, Wassenaar T, Castel V, Rosales M, Rosales M, de Haan C. 2006. Livestock's long shadow: environmental issues and options: Food and Agriculture Organisation, Rome.
Teague W, Dowhower S, Baker S, Haile N, DeLaune P, Conover D. 2011. Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agriculture, Ecosystems and Environment 141:310–322.
Thurow T. 1991. Hydrology and erosion. In ‘Grazing Management: An Ecological Perspective’. (Eds R. Heitschmidt and J. Stuth.) pp. 141–159. Timberline Press: Portland, OR.
Thurow TL, Blackburn WH, Jnr Taylor CA. 1986. Hydrologic characteristics of vegetation types as affected by livestock grazing systems, Edwards Plateau, Texas. Journal of Range Management 39:505–509.
Trimble SW, Mendel AC. 1995. The cow as a geomorphic agent—a critical review. Geomorphology 13:233–253.
Union of Concerned Scientists. 2017. Turning Soils into Sponges. https://www.ucsusa.org/sites/default/files/attach/2017/08/turning-soils-into-sponges-full-report-august-2017.pdf
Verheijen FG, Jones RJ, Rickson R, Smith C. 2009. Tolerable versus actual soil erosion rates in Europe. Earth-Science Reviews 94:23–38.
Viechtbauer W. 2010. Conducting meta-analyses in R with the metafor package. Journal of Statistical Software 36:1–48.
Webber DF, Mickelson SK, Ahmed SI, Russell JR, Powers WJ, Schultz RC, Kovar J. 2010. Livestock grazing and vegetative filter strip buffer effects on runoff sediment, nitrate, and phosphorus losses. Journal of Soil and Water Conservation 65:34–41.
Weber B, Bowker M, Zhang Y, Belnap J. 2016. Natural recovery of biological soil crusts after disturbance. Biological soil crusts: an organizing principle in drylands: Springer, pp. 479–498.
Weltz M, Wood MK. 1986. Short-duration grazing in central New Mexico: effects on sediment production. Journal of Soil and Water Conservation 41:262–266.
Wischmeier WH, Smith DD. 1960. A universal soil-loss equation to guide conservation farm planning. Transactions of the 7th International Congress of Soil Science 1: 418–425.
Zheng W, Zeng S, Bais H, LaManna JM, Hussey DS, Jacobson DL, Jin Y. 2018. Plant growth-promoting rhizobacteria (PGPR) reduce evaporation and increase soil water retention. Water Resources Research 54:3673–3687.
Zomer RJ, Trabucco A, Bossio DA, Verchot LV. 2008. Climate change mitigation: A spatial analysis of global land suitability for clean development mechanism afforestation and reforestation. Agriculture, Ecosystems and Environment 126:67–80.
Acknowledgements
We thank Naomi Huynh for commencing this database as part of a student project and Grant Fraser for giving us access to infiltration data from Queensland, Australia. We acknowledge the considerable advice and assistance with the meta-analysis code provided by Shinichi Nakagawa.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Eldridge, D.J., Ding, J. & Travers, S.K. A Global Synthesis of the Effects of Livestock Activity on Hydrological Processes. Ecosystems 25, 1780–1791 (2022). https://doi.org/10.1007/s10021-022-00746-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-022-00746-9