
Abstract
Large carnivores can reduce ungulate numbers by predation and via induced risk effects alter ungulate behavior, indirectly affecting lower trophic levels. However, predator-induced risk effects probably act at different spatial scales, which have often been ignored in trophic cascade studies. We studied how a fine-scale risk factor (distance from tree logs) affects ungulate browsing intensity and how this is modified over a landscape-scale risk gradient (distance from human settlements to wolf core) in the Białowieża forest, Poland. We found that landscape-and fine-scale risk factors strongly interacted in determining the strength and magnitude of carnivore-induced risk effects on lower trophic levels. In low-risk areas, tree logs reduced browsing intensity in small patches (approx. 4–6 m from logs), whereas in high-risk areas browsing intensity was reduced up to at least 16 m from tree logs. Moreover, the magnitude of these effects changed, with the strongest reduction in browsing intensity around tree logs in high-risk areas (up to 37%) and the smallest in low-risk areas (< 20%). Overall, the results of this study indicate that perceived risk factors act at different spatial scales, where impediments (objects blocking view and escape routes) act as a risk factor at a fine scale and carnivore distribution shapes perceived risk at the landscape scale. Moreover, these risk factors strongly interact, thereby determining the functional role of large carnivores in affecting ecosystem processes. These interactive effects should be incorporated in predator-induced trophic cascade studies to understand patterns of tree regeneration in ecosystems where large carnivores and herbivores live together.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Highlights
-
Wolves and tree logs affect deer distribution and behavior at two spatial scales.
-
Tree logs reduce sapling browsing more in risky compared to low-risk areas.
-
Landscape- and fine-scale risk factors determine the potential for tree regeneration.
Introduction
Large carnivores play a major role in ecosystem functioning through suppressing ungulate herbivory which in turn affects abundance and distributions of several fauna taxa, and vegetation development (Ripple and others 2014). These indirect trophic cascading effects of large carnivores on vegetation are induced by reduced ungulate population densities via predation and by affecting ungulate behavior and spatial distribution via perceived risk effects. The risk effects of carnivores on prey behavior have been suggested to be stronger than the reduction in prey population for inducing trophic cascades (Kotler and Holt 1989; Werner and Peacor 2003; Preisser and others 2005; Verdolin 2006; Creel and Christianson 2008; Valeix and others 2009b).
Risk effects of large carnivores seem to act at two spatial scales: carnivore distribution and activity patterns mainly determine ungulate density and distribution via predation risk on a large spatial scale (the landscape scale), whereas the presence of valleys, ridges and impediments (for example, structural objects that limit view and/or escape possibilities for prey) can affect ungulate density, distribution, behavior and perceived predation risk on a fine spatial scale (Halofsky and Ripple 2008; Kuijper and others 2015; Painter and others 2015). However, there is an ongoing debate about the existence and importance of these fine-scale risk factors compared to landscape-scale risk factors (Halofsky and Ripple 2008; Kauffman and others 2010, 2013; Winnie 2012, 2014; Beschta and Ripple 2013; Beschta and others 2014, 2018; Painter and others 2015). Although there is agreement that at a large-scale wolves change ungulate distribution from a closed toward an open landscape, there is disagreement whether fine-scale risk factors result in patchy tree recovery.
Moreover, risk effects are often dependent on landscape structures (impediments, valleys and ridges) and the strength of risk effects is modified by predator presence (Creel and others 2008; Eisenberg and others 2014; Kuijper and others 2015). Thus, risk factors operating at different spatial scales are likely to strongly interact. These interacting effects of structural fine-scale and landscape-scale risk factors in affecting ungulate behavior and their impact on the vegetation has received little attention in studies on trophic cascading effects of large carnivores (but see Kuijper and others 2015; Beschta and others 2018).
At the landscape scale, ungulates may alter their spatial distribution to reduce potential predation risk (Creel and others 2005; Valeix and others 2009a; Thaker and others 2011; Hopcraft and others 2012; White and others 2012; Latombe and others 2014), often resulting in a shift from high-quality forage but risky habitat to safer habitats with lower quality forage (Creel and others 2005; Fortin and others 2005). Which habitat type ungulates perceive as safe is to a large extent influenced by their main predator’s hunting strategy (that is, courser or ambush) and how they can themselves react to this to reduce predation risk (Creel and others 2005; Shrader and others 2008; Hopcraft and others 2012; Kuijper and others 2014; Wikenros and others 2015). Ungulates generally perceive foraging in open areas as less risky when their main predator uses an ambush strategy (Shrader and others 2008; Valeix and others 2009a), whereas closed habitats are perceived as less risky in case of cursorial/coursing predators (Creel and others 2005). These landscape-scale behavioral changes in response to predation risk can lead to cascading effects on woody plant vegetation (Creel and Christianson 2009; Ripple and Beschta 2012; Ford and others 2014; Flagel and others 2016).
At fine spatial scales, ungulates may adjust their behavior near objects or landscape elements that increase (perceived) predation risk, such as impediments or dense vegetation that reduce visibility or block escape routes independent of a predator’s hunting strategy (Halofsky and Ripple 2008; Kuijper and others 2015; Schmidt and Kuijper 2015; Stears and Shrader 2015). At high-risk sites, ungulates generally increase vigilance levels at the cost of foraging time. For example, wapiti (Cervus canadensis) are more vigilant near escape impediments (Halofsky and Ripple 2008). Likewise, red deer (Cervus elaphus) in the Białowieża Primeval Forest (BPF) in Poland avoid tree logs and increase their vigilance levels near fallen tree logs, especially when situated inside the core of a wolf territory (Kuijper and others 2015), indicating that deer perceive tree logs as risky. These behavioral changes seem to play an important role in landscape-level patterns of tree regeneration in the BPF. The increased perceived predation risk near tree logs protects saplings by reducing browsing intensity, allowing them to grow taller and faster above the browsing line (Smit and others 2012; Kuijper and others 2013, 2015).
What until now received little attention is that above-mentioned landscape-scale and fine-scale risk factors likely strongly interact. However, some of studies do suggest that the behavioral response of ungulate prey species to structural fine-scale risk factors strongly depends on landscape-scale patterns of predator presence (White and others 1998; Crosmary and others 2012; Kuijper and others 2015). As a result, the potential for trophic cascading effects of large carnivores is likely context-dependent with expected interacting effects of risk factors occurring at different spatial scales (Kuijper and others 2015; Painter and others 2015). In this study, we focus on how structural fine-scale risk factors (tree logs) and landscape-scale risk factors (perceived risk gradient related to wolf activity) interact in promoting tree regeneration. More specifically, we studied how patterns of ungulate browsing depend on the distance from tree logs and how these effects are modified by landscape-scale patterns of perceived risk of large carnivores.
Methods
Study area
This study was conducted in the Białowieża National Park (BNP; 52°45′N, 23°50′E), Poland from April to July 2015. The BNP (103 km2) is a strictly protected part of the Białowieża Primeval Forest (BPF, 600 km2) where hunting, logging or motorized traffic is not allowed and can only be entered with a permit. Due to this hands-off policy in the BNP there is a considerable amount of fallen trees that cover the forest floor (Bobiec 2002). In the BNP tree logs are, besides small rivers and swampy areas, the only terrain features present that act as impediments for ungulates since boulders, large river valleys and mountain ridges are absent.
The BPF is a temperate lowland forest of rich multispecies tree stands that consists of a mosaic of forest types, but is dominated by a deciduous oak-lime-hornbeam (Tilio-Carpinetum) forest. The climate is continental with a mean temperature of 6.8 °C and a mean annual precipitation of 641 mm. The BPF contains a varied native ungulate and carnivore assemblage: red deer (Cervus elaphus, 4.7 individuals km−1), wild boar (Sus scrofa, 3.2 ind. km−1), roe deer (Capreolus capreolus, 0.8 ind. km−1), bison (Bison bonasus, 0.8 ind. km−1) and moose (Alces alces, 0.06 ind. km−1) co-occur with wolf (Canis lupus, 2–3 ind. per 100 km) and lynx (Lynx lynx, 1–3 ind. per 100 km). In this study, we focus on the wolf–red deer interaction since red deer are the main ungulate species in the study area in terms of density and total biomass, the main browser of all occurring ungulate species and the main prey for wolves (Gębczyńska and Krasińska 1972; Gębczyńska 1980; Jędrzejewska and others 1997; Jędrzejewski and others 2000, 2002).
Wolf activity is the highest in the wolf core, where wolves have their dens and raise their pups (Jędrzejewski and others 2007), but wolves are active in the whole area (Schmidt and others 2009). The activity patterns and den sites of wolves are mainly determined by human presence, that is, wolves avoid humans in space and time (Theuerkauf and others 2003a, b), resulting in safer sites for red deer near humans (that is, human shield effect; Berger 2007). Red deer tend to occur in higher densities close to humans, in contrast to far from human settlements and close to wolves, at least within the national park where no hunting occurs (Kuijper and others 2015). Therefore, inside the National Park, red deer perceive a landscape-scale risk gradient with low perceived risk relatively close to human settlements to high perceived risk in high wolf use areas (Kuijper and others 2015).
A field test of fine-scale versus landscape-scale risk factors
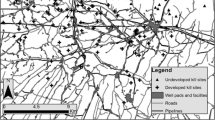
Along this perceived landscape-scale risk gradient, we searched for tree logs, our fine-scale risk factor (Figure 1). Selected tree logs had a length of 15.1 ± 0.57 m (mean ± SE, range = 12–23 m) and a height of 103 ± 3 cm (range = 77–147 cm), with an average volume of 12.4 ± 0.96 m3. Kuijper and others (2015) showed that tree logs greater than 12 m long and greater than 1 m in height resulted in increased vigilance in red deer and therefore form a significant impediment to block the view or escape possibilities. Near the tree log a minimum of 30 saplings had to grow in a range of 16 m from the tree log to allow for a good estimate of browsing intensity. In this study, we focused on saplings between 50 and 200 cm tall as this covers the preferred browsing height of red deer (Renaud and others 2003; Kuijper and others 2013). We confirmed that no other tree log of similar size was present within 50 m radius, therefore saplings were not physically protected by other logs and thus we could measure the distance effect of a single tree log on browsing intensity. In total 26 locations were selected, widely distributed across the forest (distance between locations: mean = 4.31 km; range = 0.41–9.22 km) and all situated within the deciduous or mixed-deciduous forest type (Figure 1).

Locations of the sampled tree logs situated on a perceived predation risk gradient, with low risk close to human settlement and high risk far from human settlements, close to wolves (Kuijper and others 2015). The gradient was divided in four classes of distance from human settlements: filled circle—tree logs located within 1–2.4 km (low risk, n = 6); circle—tree logs located within 2.5–3.9 km (intermediate low risk, n = 9); square—tree logs located within 4.0–5.4 km (intermediate high risk, n = 6); and filled square—tree logs located within 5.5–6.9 km (high risk, n = 5).
Browsing intensity
At each location, we measured a maximum of 10 saplings per strip in 16 adjacent strips of 1 m wide parallel to the tree log (closest strip < 1 m from log; farthest strip 15–16 m from log), to analyze the effect of distance from tree log in 1-m classes (Figure 2). By measuring saplings up to a maximum of 16 m from the tree log we aimed to cover the area with the strongest effects; our previous studies suggested that risk effects operate at the scale of several meters (Kuijper and others 2013, 2014). We started measuring saplings at the center of the tree log and moved to the edges of the strip until we had measured 10 saplings per strip. Sample area per strip was on average 13.4 ± 0.20 m2 (± SE, range = 1.1–23 m2), depending on sapling density. In addition, we sampled a control strip at approximately 50 m from the log (sampling area mean ± SE = 2.7 ± 0.17 m2). We assumed this to be a ‘no-risk distance’ from the tree log as Halofsky and Ripple (2008) observed that deer were no longer vigilant at greater than 30 m from an impediment. Within a 50 m radius of the control no tree logs larger than 100 cm high and 12 m long were present.

Setup of the field study. Up to a distance of 16 m from a tree log of > 12 m long and > 0.8 m high we measured the height, diameter and browsing intensity of 10 saplings 50–200 cm in plot of 1 m wide parallel to the tree log. At 50 m from the tree log—the control plot—we measured the same variables for a maximum of 10 saplings. We measured visibility at a height of 50 and 150 cm from the ground at 8 m from the tree log and on the control plot (filled circle). To calculate canopy openness, we took photos with a fish-eye-lens at 2, 6, 10 and 14 m from the tree log and in the middle of the control plot (filled triangle).
For each sapling between 50 and 200 cm in height (mean ± SE = 81.9 ± 0.6 cm, range: 50–200 cm), diameter and browsing intensity were measured. Browsing intensity was defined as the proportion of the 10 top branches that showed browsing marks (following Kuijper and others 2013) as the top branches are more likely to be browsed, and top branch browsing is the main factor slowing down tree growth (Kuijper and others 2010).
Habitat visibility and canopy openness
We measured visibility per location to be able to correct for possible differences between locations, as visibility is an important determinant of perceived predation risk (Underwood 1982; Ripple and Beschta 2006; Sahlén and others 2016). Visibility was measured as the distance at which the laser of a handheld rangefinder (Bresser 4 × 21 Rangefinder.800, accuracy ± 1 m up to 200 m and ± 5 m from 200 m onwards) hit any object. We repeated this three times, and slightly varied the position of the rangefinder for each measurement, to avoid hitting the same object three times in a row. We measured visibility at 8 m distance from the tree log as an average estimate of visibility for the whole range from 0 to 16 m from the tree log, and at the center of the control strip (Figure 2) in every cardinal and subcardinal direction (resulting in eight directions). Visibility was measured with the rangefinder kept at a height of 50 and 150 cm, the assumed minimum and maximum height adult red deer scans the surrounding while foraging or being vigilant. For both heights (50 and 150 cm), the average of the three visibility measurements was calculated and used for statistical analysis.
Canopy openness, a measure of light availability, was measured to correct for potential differences between locations as a possible confounding factor affecting growth rate. An upward canopy photograph with a fish-eye lens at 1 m above ground level was taken and analyzed with Gap Light Analyzer (GLA, version 2.0) to calculate the percentage of canopy light. We took measurements at 2, 6, 10 and 14 m and at the control at 50 m to test if canopy openness changed with increasing distance from the tree log (Figure 2).
Statistical methods
Data were analyzed with R, version 3.2.3 (https://cran.r-project.org/). We used a generalized mixed effect model with a binomial distribution and a logit-link function from the lme4-package (glmer), with browsing intensity as a response variable. We tested if browsing intensity was affected by the interaction between distance from a tree log and distance from human settlements. The model included as explanatory variables distance from tree log, distance from human settlements, the interaction term between distance from tree log and distance from human settlements, visibility at 50 cm, visibility at 150 cm, and canopy openness, and location as random factor. We used backward stepwise regression to find the minimal adequate model, based on p value and Akaike information criterion (Appendix S1, Table S1). With the least-squares means of the minimal adequate model (calculated with the lsmeans-package), we performed a pairwise comparison of the browsing intensity between each distance from the tree log class and the control class per distance from human settlements class. Sampled tree logs are grouped in four distance classes from human settlements (1.0–2.4, 2.5–3.9, 4.0–5.4 and 5.5–7.0 km). Within locations the measured saplings are grouped per 2 m distance from tree logs (0–2, 2–4, 4–6, 6–8, 8–10, 10–12, 12–14, 14–16 m and the control class) to get proper sample sizes with comparable sapling numbers between the different classes for statistical analysis (Appendix S1: Table S2).
In total, we found 12 different tree species with sapling numbers unequally distributed over the species (Appendix S1: Table S2). Hornbeam was by far the most dominant species (mean = 68%, range = 40.8–90.1%) followed by lime (14.5%), whereas for five species we found less than 10 individuals. Therefore, we could not investigate whether distance from tree log or distance from human settlements affect tree species differently due to contrasts in palatability. Sapling densities (number of saplings per m2) are comparable between locations and there is no support that sapling density changes with increased distance from the tree log or distance from human settlements (Appendix S1: Table S3). High sapling densities can attract more deer, leading to a higher browsing intensity. Alternatively, browsing intensity per sapling could be diluted at high sapling density. However, due to the comparable sapling densities, we did not have to add sapling density as covariable in our models.
For graphical representation we calculated the browsing intensity log response ratio (Borenstein and others 2009) as measure for effect size to determine how much the browsing intensity differed between the control class and each distance from tree log class, per distance from human settlements class. The log response ratio calculates the proportional difference between the mean browsing intensity at each distance from tree log class and the mean browsing intensity on the control class. Positive values indicate a higher browsing intensity compared to the control class, while negative values indicate a lower browsing intensity compared to control class.
With a t test we tested whether the visibility at 50 and 150 cm differed between the control strip and at 8 m from the tree log and whether the canopy openness changed with increased distance from the tree log.
Results
Interactive effects between fine-scale and landscape-scale risk factors
The fine-scale effect of tree logs on browsing intensity interacted strongly with the landscape-scale risk gradient (χ2 (24) = 100.19, p < 0.001): in areas with high perceived risk, browsing intensity was reduced more and at a larger distance from tree logs. In low-risk areas the browsing intensity was reduced up to a distance of approximately 4–6 m from tree logs, whereas in high-risk areas a tree log reduced browsing intensity up to a distance of at least 16 m (Figure 3; Appendix S1, Table S4). Moreover, browsing intensity near tree logs was reduced within the first 4 m with 20.3 ± 10.8% (mean ± SE) in low-risk areas compared to a maximum reduction of 37%, and more than 30% within the first 8 m from a tree log in high-risk areas (Figure 3). The browsing intensity was not significantly affected by visibility at 50 cm (χ2 (1) = 0.569, p = 0.451), or by the canopy openness (χ2 (1) = 1.128, p = 0.288), but was significantly affected by the visibility at 150 cm (χ2 (1) = 24.301, p < 0.001).

Difference in browsing intensity between the control class and distance from tree log class over the perceived risk gradient (based on Kuijper and others 2015; see Figure 1 for classes of perceived risk). In areas with low-risk tree logs reduce browsing intensity within the first 4 m (indicated with the dashed line) and with a maximum of 20%. With increased risk browsing intensity is more reduced (maximum of − 37%) compared to control class and the distance till which tree logs reduce browsing intensity increase still 16 m.
Habitat characteristics
The visibility at the control strip was not significantly different from the visibility measured at 8 m from the tree log (visibility at 50 cm: F(1) = 2.326, p = 0.134; visibility at 150 cm: F(1) = 0.452, p = 0.504). The canopy openness did not significantly change with increased distance from the tree log (F(4) = 0.53, p = 0.714).
Discussion
Although many studies have shown the importance of risk factors in explaining the functional role of large carnivores, few have addressed how risk factors operating at different spatial scales can interact with one another. We found an interaction between structural fine-scale (tree log) and landscape-scale (perceived risk gradient) risk factors in determining the strength and magnitude of carnivore-induced risk effects on lower trophic levels. In low perceived risk areas, tree logs reduced browsing intensity in small patches (ca. 4–6 m from logs), whereas in high-risk areas browsing intensity was reduced in larger patches (over 16 m from tree logs). Moreover, the perceived landscape-scale risk affected the magnitude of these effects around tree logs, with the smallest reduction in browsing intensity in low-risk areas (< 20%) and the strongest reduction in high-risk areas (up to 37%). These results suggest that risk factors operate at different spatial scales and strongly interact and determine potential tree regeneration patterns in ecosystems where large carnivores and herbivores co-occur.
In this study, we focus on the wolf–red deer interaction since red deer is the dominant ungulate species in terms of density, the main browser (Gębczyńska and Krasińska 1972; Gębczyńska 1980) and forms 60–80% of the wolves diet (with roe deer as second prey; Jędrzejewski and others 2000, 2002) in the BPF. Yet we cannot ignore the possible influences of other ungulate species (roe deer, European bison and moose) on sapling browsing, as well as of impacts of the lynx as second large predator in our study system. As a typical ambush predator, the lynx probably reinforces the perceived risk near an impediment for both red deer and roe deer, which strengthens our findings of lower browsing intensity near logs. In contrast, we suggest the occurrence of European bison and moose does not affect our findings. Bison and moose are rarely preyed upon (< 1%) by both wolf and lynx (Jędrzejewska and Jędrzejewski 1998) and therefore are not assumed to perceive a carnivore-induced landscape of fear in this system with high and low-risk sites that will affect their foraging patterns (Hayward and others 2015).
The importance of fine-scale risk factors determining ungulate browsing
Reduced browsing pressure is expected near impediments that potentially block the view of approaching large carnivores or block escape routes. The few studies investigating the effect of structural fine-scale risk factors on ungulate behavior show an avoidance of such impediments, an increase in vigilance, and a reduction in foraging near such risk factors (Halofsky and Ripple 2008; Iribarren and Kotler 2012; Kuijper and others 2015). As a result, a lower browsing intensity is often observed on woody saplings associated with impediments (Smit and others 2012; Kuijper and others 2013; Beschta and others 2018). In contrast, Winnie (2012) found that impediments promoted tree regeneration only via physical protection against ungulate browsing and not via increased perceived risk. The role of structural fine-scale risk factors in affecting the effect of large carnivores on ecosystem functioning remains therefore strongly debated (Beschta and Ripple 2013; Kauffman and others 2013; Beschta and others 2014; Winnie 2014). In our study area red deer increase vigilance levels and avoid fine-scale risk factors (Kuijper and others 2015) and in the present study we show that these lower densities and behavioral changes result in reduced browsing pressure near fine-scale risk factors. However, we cannot distinguish whether this reduced browsing pressure near tree logs is the result of reduced deer density, a reduction in foraging, or a combination of both. All tree saplings were fully accessible to ungulate browsing as all used locations contained only one tree log and no other impediments within 50 m. Physical protection therefore could not have played a role, and we argue that tree logs truly impose a ‘fear effect’ that deer try to avoid. As we found no change in visibility with increased distance from tree logs, we argue that blocking escape routes are the main reason why deer perceive foraging near tree logs as risky. Maybe deer can jump quite easily over an obstacle that is 1 m high, but the tree log also has a width of about 1 m and has lateral branches stretching further that can complicate the jump. Due to the generally low visibility in this closed-canopy forest, deer in these habitats might rely more on other cues indicating predation risk (Kuijper and others 2014) rather than visual cues.
The difference in results between Winnie (2012) and our study can possibly be explained by the difference in behavior of both predator and prey in the half-open system they studied compared to the closed-canopy forest in the present study. In a closed-canopy forest, wolves can use an ambush strategy when visibility is low (Petterson and Cuccie 2003) and use cover as concealment while hunting (Kunkel and Pletscher 2001). In contrast, in the more open landscapes wolves predominantly course their prey. Wolves that use an ambush strategy to kill their prey are expected to create more predictable risk effects and thereby stronger behavioral responses in prey on risky sites than wolves using a coursing strategy (Preisser and others 2007). In the BPF lynx probably reinforces the effect of wolves on deer behavior since it is a typical ambush hunter that sits and waits in concealment for suitable prey (mainly roe deer in the BPF). As a result, red deer and roe deer in closed-canopy forest may perceive foraging near tree logs as more risky, causing a stronger reduction in browsing intensity compared to deer foraging in open, high-visibility habitats. Moreover, due to the dominant habitat types the diet of European red deer (C. elaphus) consists of 70% woody species in our closed-canopy forest (Gębczyńska 1980), whereas the diet of the closely related American wapiti (C. canadensis) comprises more graminoids than woody species (66% vs 29%, Christianson and Creel 2007). However, the percentage woody species in the diet of deer in both systems likely depends on winter severity, as snow depth can affect the availability of graminoids resulting in an increase in the consumption of woody species between years. Given the general difference in diet composition, suppression of tree regeneration is probably stronger by red deer than by wapiti. In comparison with the GYE which contains big boulders, steep ridges and large rivers acting as impediments, tree logs are the only impediments present in the Białowieża forest. We therefore argue that in a closed-canopy forest system structural fine-scale risk factors such as tree logs are of major importance in facilitating successful tree regeneration.
The extent to which fine-scale risk factors reduce browsing pressure strongly interacts with risk factors operating at the landscape scale. We found increased reduction in browsing pressure near tree logs in high-risk areas. This result corresponds with Kuijper and others (2015) who observed that red deer avoid areas with high perceived risk and increase their vigilance level when a fine-scale risk factor is present. The avoidance of tree logs could, besides avoiding risk, be a matter of convenience—deer walk around the patch because it blocks their route—causing lower deer densities and therefore reduced browsing near tree logs. When food is abundant, deer might avoid tree logs as a matter of convenience, but when food is scarce increased risk might prevent deer from foraging close to the logs. In our study, we showed that the radius of these ‘patches of fear’ imposed by structural fine-scale risk factors enlarge with increased risk at the landscape scale. Moreover, these behavioral changes lead to a lower browsing pressure and create ‘windows of opportunity’ for tree regeneration near structural fine-scale risk factors in high-risk areas.
For successful recruitment, a tree has to survive all stages from seed survival via seed germination and seedling establishment to sapling growth until it grows into the canopy. Earlier studies in the Białowieża forest showed that abiotic factors are the main factor determining seedling establishment and hence the sapling density of small-size classes (< 50 cm). Ungulate herbivory does not affect their numbers (Kuijper and others 2010), which explains the equal sapling densities that occurred over the landscape-scale risk gradient in the present study, despite differences in deer browsing pressure. During these early stages of tree recruitment spatial discordance occurs, where seed survival is highest in areas without dead wood lying on the forest floor, and seedling germination and sapling establishment (< 50 cm) is the highest in the presence of dead wood (van Ginkel and others 2013). However, for tree saplings to grow into taller size classes (> 50 cm), ungulate herbivory becomes the main factor determining their density (Kuijper and others 2010) and intense ungulate browsing keeps trees in recruitment bottlenecks (Churski and others 2017, Cromsigt and Kuijper 2011). Once trees grow beyond the ungulate browsing line of 2 m (Kuijper and others 2013), they can escape this herbivore-driven recruitment bottleneck (Churski and others 2017). Our study suggests that saplings within 8 m of tree logs, and in high wolf use areas have a reduced browsing intensity and hence the highest chance to escape this herbivore-driven bottleneck, and grow beyond the browsing line into the tree canopy. Overall, we show that the variation in perceived predation risk, results in strong spatial heterogeneity in browsing intensity, causing improved odds for successful tree regeneration in high-risk areas, implying a trophic cascade.
Interactive effects of spatial scales and consequences for trophic cascades
Most studies on behaviorally mediated effects of large carnivores on their ungulate prey species focused on risk effects operating at the landscape level. Landscape-scale risk factors affect spatiotemporal ungulate activity and distribution (Laundré and others 2001; Hernández and Laundré 2005; Riginos and others 2008; Valeix and others 2009b; Thaker and others 2011; Periquet and others 2012) with consequences for woody vegetation (Kauffman and others 2010; Ford and others 2014; Beschta and Ripple 2016). In contrast, the role of fine-scale risk factors is still strongly debated (Halofsky and Ripple 2008; Kauffman and others 2010, 2013, Winnie 2012, 2014; Beschta and Ripple 2013; Beschta and others 2014). However a recent study shows that fine-scale risk factors seem to reduce browsing intensity only in areas with frequent wolf visits (Beschta and others 2018). This result is in line with the present study, in which we show the importance of structural fine-scale risk factors for promoting tree regeneration in old-growth forests which are characterized by large amounts of dead wood (Bobiec 2002). Large carnivores apparently create a landscape of fear in which the size of ‘patches of fear’ is determined by the interaction between risk factors operating at different spatial scales. For an improved understanding of how large carnivores indirectly affect vegetation in ecosystems, it is crucial to consider these interactive effects between fine- and landscape-scale risk factors. As trade-offs exist between food quality and risk effects (McArthur and others 2014) future studies should aim to disentangle these interactive effects of spatial scales, and include tree palatability, for a full understanding of how forest composition and dynamics are shaped by large carnivores.
References
Berger J. 2007. Fear, human shields and the redistribution of prey and predators in protected areas. Biol Lett 3:620–3.
Beschta RL, Ripple WJ. 2013. Are wolves saving Yellowstone’s aspen? a landscape-level test of a behaviorally mediated trophic cascade: comment. Ecology 94:1420–5.
Beschta RL, Ripple WJ. 2016. Riparian vegetation recovery in Yellowstone: the first two decades after wolf reintroduction. Biol Conserv 198:93–103.
Beschta RL, Eisenberg C, Laundré JW, Ripple WJ, Rooney TP. 2014. Predation risk, elk, and aspen: comment. Ecology 95:2669–71.
Beschta RL, Painter LE, Ripple WJ. 2018. Trophic cascades at multiple spatial scales shape recovery of young aspen in Yellowstone. For Ecol Manag 413:62–9.
Bobiec A. 2002. Living stands and dead wood in the Białowieża forest: suggestions for restoration management. For Ecol Manag 165:125–40.
Borenstein M, Hedges LV, Higgins JPT, Rothstein HR. 2009. Introduction to meta-analysis.
Christianson DA, Creel S. 2007. A review of environmental factors affecting elk winter diets. J Wildl Manag 71:164–76.
Churski M, Bubnicki JW, Jędrzejewska B, Kuijper DPJ, Cromsigt JPGM. 2017. Brown world forests: increased ungulate browsing keeps temperate trees in recruitment bottlenecks in resource hotspots. New Phytol 214:158–68.
Creel S, Christianson D. 2008. Relationships between direct predation and risk effects. Trends Ecol Evol 23:194–201.
Creel S, Christianson D. 2009. Wolf presence and increased willow consumption by Yellowstone elk: implications for trophic cascades. Ecology 90:2454–66.
Creel S, Winnie J, Maxwell B, Hamlin K, Creel M. 2005. Elk alter habitat selection as an antipredator response to wolves. Ecology 86:3387–97.
Creel S, Winnie John A J, Christianson D, Liley S. 2008. Time and space in general models of antipredator response: tests with wolves and elk. Anim Behav 76:1139–46.
Cromsigt JPGM, Kuijper DPJ. 2011. Revisiting the browsing lawn concept: evolutionary interactions or pruning herbivores? Persp Plant Ecol Evol Syst 13:207–15.
Crosmary WG, Valeix M, Fritz H, Madzikanda H, Côté SD. 2012. African ungulates and their drinking problems: hunting and predation risks constrain access to water. Anim Behav 83:145–53.
Eisenberg C, Hibbs DE, Ripple WJ, Salwasser H. 2014. Context dependence of elk (Cervus elaphus) vigilance and wolf (Canis lupus) predation risk. Can J Zool 92:727–36.
Flagel DG, Belovsky GE, Beyer DE. 2016. Natural and experimental tests of trophic cascades: gray wolves and white-tailed deer in a Great Lakes forest. Oecologia 180:1183–94.
Ford AT, Goheen JR, Otieno TO, Bidner L, Isbell LA, Palmer TM, Ward D, Woodroffe R, Pringle RM. 2014. Large carnivores make savanna tree communities less thorny. Science 346:346–9.
Fortin D, Beyer HL, Boyce MS, Smith DW, Duchesne T, Mao JS. 2005. Wolves influence elk movements: behavior shapes a trophic cascade in Yellowstone National Park. Ecology 86:1320–30.
Gębczyńska Z. 1980. Food of the roe deer and red deer in the Białowieża Primeval Forest. Acta Theriol 25:487–500.
Gębczyńska Z, Krasińska M. 1972. Food preferences and requirements of the European Bison. Acta Theriol 17:105–17.
Halofsky JS, Ripple WJ. 2008. Fine-scale predation risk on elk after wolf reintroduction in Yellowstone National Park, USA. Oecologia 155:869–77.
Hayward MW, Ortmann S, Kowalczyk R. 2015. Risk perception by endangered European bison Bison bonasus is context (condition) dependent. Landsc Ecol 30:2079–93.
Hernández L, Laundré JW. 2005. Foraging in the ‘landscape of fear’ and its implications for habitat use and diet quality of elk Cervus elaphus and bison Bison bison. Wildl Biol 11:215–20.
Hopcraft JGC, Anderson TM, Pérez-Vila S, Mayemba E, Olff H. 2012. Body size and the division of niche space: food and predation differentially shape the distribution of Serengeti grazers. J Anim Ecol 81:201–13.
Iribarren C, Kotler BP. 2012. Patch use and vigilance behaviour by Nubian ibex: the role of the effectiveness of vigilance. Evol Ecol Res 14:223–34.
Jędrzejewska B, Jędrzejewski W. 1998. Predation in vertebrate communities. The Bialowieza Primeval Forest as a case study. Berlin: Springer. p 452p.
Jędrzejewska B, Jędrzejewski W, Bunevich AN, Milkowski L, Krasinski ZA. 1997. Factors shaping population densities and increase rates of ungulates in Bialowieza Primeval Forest (Poland and Belarus) in the 19th and 20th centuries. Acta Theriol 42:399–451.
Jędrzejewski W, Schmidt K, Theuerkauf J, Jędrzejewska B, Kowalczyk R. 2007. Territory size of wolves Canis lupus: Linking local (Białowieża Primeval Forest, Poland) and Holarctic-scale patterns. Ecography 30:66–76.
Jędrzejewski W, Jędrzejewska B, Okarma H, Schmidt K, Zub K, Musiani M. 2000. Prey selection and predation by wolves in Białowieża Primeval Forest, Poland. J Mammal 81:197–212.
Jędrzejewski W, Schmidt K, Theuerkauf J, Jędrzejewska B, Selva N, Zub K, Szymura L. 2002. Kill rates and predation by wolves on ungulate populations in Białowieża Primeval Forest (Poland). Ecology 83:1341–56.
Kauffman MJ, Brodie JF, Jules ES. 2010. Are wolves saving Yellowstone’s aspen? a landscape-level test of a behaviorally mediated trophic cascade. Ecology 91:2742–55.
Kauffman MJ, Brodie JF, Jules ES. 2013. Are wolves saving Yellowstone’s aspen? a landscape-level test of a behaviorally mediated trophic cascade: reply. Ecology 94:1425–31.
Kotler BP, Holt RD. 1989. Predation and competition: the interaction of two types of species interactions. Oikos 54:256–60.
Kuijper DPJ, Cromsigt JPGM, Jędrzejewska B, Miścicki S, Churski M, Jędrzejewski W, Kweczlich I. 2010. Bottom-up versus top-down control of tree regeneration in the Białowieża Primeval Forest, Poland. J Ecol 98:888–99.
Kuijper DPJ, de Kleine C, Churski M, Van Hooft P, Bubnicki JW, Jędrzejewska B. 2013. Landscape of fear in Europe: wolves affect spatial patterns of ungulate browsing in Białowieża Primeval Forest, Poland. Ecography 36:1263–75.
Kuijper DPJ, Verwijmeren M, Churski M, Zbyryt A, Schmidt K, Jędrzejewska B, Smit C. 2014. What cues do ungulates use to assess predation risk in dense temperate forests? PLoS ONE 9:e84607.
Kuijper DPJ, Bubnicki JW, Churski M, Mols B, Van Hooft P. 2015. Context dependence of risk effects: wolves and tree logs create patches of fear in an old-growth forest. Behav Ecol 26:1558–68.
Kunkel K, Pletscher DH. 2001. Winter hunting patterns of wolves in and near Glacier National Park, Montana. J Wildl Manag 65:520–30.
Latombe G, Fortin D, Parrott L. 2014. Spatio-temporal dynamics in the response of woodland caribou and moose to the passage of grey wolf. J Anim Ecol 83:185–98.
Laundré JW, Hernández L, Altendorf KB. 2001. Wolves, elk, and bison: reestablishing the ‘landscape of fear’ in Yellowstone National Park, U.S.A. Can J Zool 79:1401–9.
McArthur C, Banks PB, Boonstra R, Forbey JS. 2014. The dilemma of foraging herbivores: dealing with food and fear. Oecologia 176:677–89.
Painter LE, Beschta RL, Larsen EJ, Ripple WJ. 2015. Recovering aspen follow changing elk dynamics in Yellowstone: evidence of a trophic cascade? Ecology 96:252–63.
Periquet S, Todd-Jones L, Valeix M, Stapelkamp B, Elliot N, Wijers M, Pays O, Fortin D, Madzikanda H, Fritz H, Macdonald DW, Loveridge AJ. 2012. Influence of immediate predation risk by lions on the vigilance of prey of different body size. Behav Ecol 23:970–6.
Petterson, RO. and Cuccie, P. 2003. The wolf as a carnivore. In: Mech LD, Boitani L, Eds. Wolves: behaviour, ecology and conservation. The University of Chicago Press, pp 104–31.
Preisser EL, Bolnick DI, Benard ME. 2005. Scared to death? the effects of intimidation and consumption in predator–prey interactions. Ecology 86:501–9.
Preisser L, Orrock L, Schmitz J. 2007. Predator hunting mode and habitat domain alter nonconsumptive effects in predator–prey interactions. Ecology 88:2744–51.
Renaud PC, Verheyden-Tixier H, Dumont B. 2003. Damage to saplings by red deer (Cervus elaphus): effect of foliage height and structure. For Ecol Manag 181:31–7.
Riginos C, Grace JB, Top-down VS, Grace B. 2008. Savanna tree density, herbivores, and the herbaceous community: bottom-up vs. top-down effects. Ecology 89:2228–38.
Ripple WJ, Beschta RL. 2006. Linking wolves to willows via risk-sensitive foraging by ungulates in the northern Yellowstone ecosystem. For Ecol Manag 230:96–106.
Ripple WJ, Beschta RL. 2012. Trophic cascades in Yellowstone: the first 15 years after wolf reintroduction. Biol Conserv 145:205–13.
Ripple WJ, Estes JA, Beschta RL, Wilmers CC, Ritchie EG, Hebblewhite M, Berger J, Elmhagen B, Letnic M, Nelson MP, Schmitz OJ, Smith DW, Wallach AD, Wirsing AJ. 2014. Status and ecological effects of the world’s largest carnivores. Science 343:1241484.
Sahlén E, Noell S, DePerno CS, Kindberg J, Spong G, Cromsigt JPGM. 2016. Phantoms of the forest: legacy risk effects of a regionally extinct large carnivore. Ecol Evol 6:791–9.
Schmidt K, Kuijper DPJ. 2015. A ‘death trap’ in the landscape of fear. Mamm Res 60:275–84.
Schmidt K, Jędrzejewski W, Okarma H, Kowalczyk R. 2009. Spatial interactions between grey wolves and Eurasian lynx in Białowieża Primeval Forest, Poland. Ecol Res 24:207–14.
Shrader AM, Brown JS, Kerley GIH, Kotler BP. 2008. Do free-ranging domestic goats show ‘landscapes of fear’? patch use in response to habitat features and predator cues. J Arid Environ 72:1811–19.
Smit C, Kuijper DPJ, Prentice D, Wassen MJ, Cromsigt JPGM. 2012. Coarse woody debris facilitates oak recruitment in Bialowieza Primeval Forest, Poland. For Ecol Manag 284:133–41.
Stears K, Shrader AM. 2015. Increases in food availability can tempt oribi antelope into taking greater risks at both large and small spatial scales. Anim Behav 108:155–64.
Thaker M, Vanak AT, Owen CR, Ogden MB, Niemann SM, Slotow R. 2011. Minimizing predation risk in a landscape of multiple predators: effects on the spatial distribution of African ungulates. Ecology 92:398–407.
Theuerkauf J, Jędrzejewski W, Schmidt K, Gula R. 2003a. Spatiotemporal segregation of wolves from humans in the Bialowieza Forest (Poland). J Wildl Manag 67:706–16.
Theuerkauf J, Rouys S, Jedrzejewski W. 2003b. Selection of den, rendezvous, and resting sites by wolves in the Bialowieza Forest, Poland. Can J Zool 81:163–7.
Underwood R. 1982. Vigilance behaviour in grazing African antelopes. Behaviour 79:81–107.
Valeix AM, Loveridge AJ, Davidson Z, Murindagomo F, Macdonald DW, Ecology S, Jan N, Chamaill S, De Lyon U, Mendel BG. 2009a. Behavioral adjustments of African herbivores to predation risk by lions: spatiotemporal variations influence habitat use. Ecology 90:23–30.
Valeix M, Fritz H, Loveridge AJ, Davidson Z, Hunt JE, Murindagomo F, Macdonald DW. 2009b. Does the risk of encountering lions influence African herbivore behaviour at waterholes? Behav Ecol Sociobiol 63:1483–94.
van Ginkel HAL, Kuijper DPJ, Churski M, Zub K, Szafrańska P, Smit C. 2013. Safe for saplings not safe for seeds: Quercus robur recruitment in relation to coarse woody debris in Białowieża Primeval Forest, Poland. For Ecol Manag 304:73–9.
Verdolin JL. 2006. Meta-analysis of foraging and predation risk trade-offs in terrestrial systems. Behav Ecol Sociobiol 60:457–64.
Werner EE, Peacor SD. 2003. A review of trait-mediated indirect interactions in ecological communities. Ecology 84:1083–100.
White CA, Olmsted CE, Kay CE. 1998. Aspen, elk, and fire in the Rocky Mountain national parks of North America. Wildl Soc Bull 26:449–62.
White PJ, Proffitt KM, Lemke TO. 2012. Changes in elk distribution and group sizes after wolf restoration. Am Midl Nat 167:174–87.
Wikenros C, Kuijper DPJ, Behnke R, Schmidt K. 2015. Behavioural responses of ungulates to indirect cues of an ambush predator. Behaviour 152:1019–40.
Winnie JA. 2012. Predation risk, elk, and aspen: tests of a behaviorally mediated trophic cascade in the Greater Yellowstone Ecosystem. Ecology 93:2600–14.
Winnie JA. 2014. Predation risk, elk and aspen: reply. Ecology 95:2671–4.
Acknowledgements
We would like to thank Ryan Leroux for help with data collection and the managers of the Białowieża National Park for access to the study site. We would also like to thank the reviewers for their critical but valuable comments that improved this paper. The work of HALVG was supported by the Ubbo Emmius Fund of the University of Groningen and the Academy Ecology Fund. In addition, the work of HALVG and DPJK was supported by funding of the National Science Centre, Poland (Grant No: 2015/17/B/NZ8/02403).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
van Ginkel, H.A.L., Kuijper, D.P.J., Schotanus, J. et al. Wolves and Tree Logs: Landscape-Scale and Fine-Scale Risk Factors Interactively Influence Tree Regeneration. Ecosystems 22, 202–212 (2019). https://doi.org/10.1007/s10021-018-0263-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-018-0263-z